
Abstract
Mesenchymal stem cells (MSC) from bone marrow and outgrowth endothelial cells (OEC) from peripheral blood are considered as attractive cell types for applications in regenerative medicine aiming to build up complex vascularized tissue-engineered constructs. MSC provide several advantages such as the potential to differentiate to osteoblasts and to support the neovascularization process by release of proangiogenic factors. On the other hand, the neovascularization process can be actively supported by OEC forming perfused vascular structures after co-implantation with other cell types. In this study the formation of angiogenic structures in vitro was investigated in cocultures of MSC and OEC, cultured either in the medium for osteogenic differentiation of MSC (ODM) or in the medium for OEC cultivation endothelial cell growth medium-2 (EGM2 Bullet Kit). After 2 weeks, cocultures in EGM2 formed more microvessel-like structures compared to cocultures in ODM as demonstrated by immunofluorescence staining for the endothelial marker CD31. Increased expression of CD31 and CD146 in quantitative real-time polymerase chain reaction as well as a higher percentage of CD31- and CD146-positive cells in flow cytometry indicated a beneficial influence of EGM2 on endothelial cell growth and function. In addition, the improved formation of vascular structures in EGM2 correlates with higher levels of the proangiogenic factor vascular endothelial growth factor and platelet-derived growth factor in the supernatant of cocultures as well as in monocultures of MSC when cultivated in EGM-2. Nevertheless, ODM was more suitable for the differentiation of MSC to osteoblastic lineages in the cocultures, whereas EGM2 favored factors involved in vessel stabilization by pericytes. In conclusion, this study highlights the importance of medium components for cell interaction triggering the formation of angiogenic structures.
Introduction
Mesenchymal stem cells (MSC) are considered as an essential cell type contributing to bone repair in adults. After tissue injury, MSC are supposed to migrate to the healing site and to colonize the wound. During the formation of new bone, MSC proliferate and then differentiate in response to different growth factors and cytokines released from their neighbor cells2–4 into chondrogenic and osteogenic lineages. The participation of MSC in repair processes of damaged bone and other tissues such as tendon, cartilage, meniscus, and skin has been demonstrated in several studies.5–11 In addition, the application of MSC in regenerative medicine provides a number of advantages. MSC can be isolated from different sources and tissues such as bone marrow, blood, adipose tissue, liver, and spleen.12–17 Further, MSC are characterized by a high expansion rate and the ability to differentiate into diverse cell types such as adipogenic, osteogenic, chondrogenic, and myogenic lineages.12,18 In addition, they are also able to support the neovascularization process by the release of proangiogenic factors such as angiopoetin-1 (Ang-1), basic fibroblast growth factor (bFGF), and vascular endothelial growth factor (VEGF).19,20 Another attractive feature of MSC for applications in regenerative medicine is their low immunogenicity.21,22
Another cell type implicated in the vascularization of damaged bone tissue is the endothelial progenitors cell (EPC) circulating in the peripheral blood and capable of being recruited to the wound site during bone repair processes.2–4 Although the mechanisms are still critically debated, the mixed population of EPC seems to contain cell types that differentiate to mature endothelial cells and incorporate into microvessels. On the other hand, different cell fractions of EPC support the angiogenic activity of mature endothelial cells through a paracrine mechanism based on the production of growth factors such as VEGF.2–4,23 A very distinct cell type that gives endothelial colonies in heterogeneous EPC cultures are outgrowth endothelial cells (OEC), which are considered as a cell type with several endothelial characteristics, including the ability to contribute to perfused vascular structures in vivo.24–32 OEC are currently being discussed as an attractive source of autologous endothelial cells. They can be isolated from peripheral blood, resemble a homogenous, well-defined population, and, in addition, reveal an attractive expansion capacity.25,26,28,33
The combination of OECs and MSCs could be beneficial for generation of prevascularized tissue-engineered constructs. Several studies report that cocultivation or co-implantation of endothelial cells with MSC or other cells has a positive effect on the formation of vascular structures, which could be observed both in vivo and in vitro.34–36 Cocultivation of endothelial cells and MSC supports the development of tubular structures and complex vascular networks.34,35 Further studies report that MSC in coculture with endothelial cells result in a stabilization of vascular structures by pericyte-like cells. 34 Several studies reveal that MSC influence the formation of angiogenic structures via a paracrine mechanism.37–39 In general, the release of proangiogenic factors, such as VEGF by a series of mesenchymal cells, including primary osteoblasts and smooth muscle cells, together with the direct cellular communication via gap junctions to endothelial cells, are considered to induce or support the angiogenic process.40–44 The detailed mechanisms by which mesenchymal cells accelerate bone repair and angiogenesis are currently under investigation. In this context, in vitro studies using cocultures of OEC and MSC may help to elucidate cell–cell interactions and the underlying paracrine mechanisms. Moreover, they serve as attractive in vitro test systems for the development of new approaches, and might be used to assess new therapeutic compounds, or to assess the impact of biomaterials on cell functionality. 45 In addition, they also enable scientists and clinicians to build up more complex constructs with a view to a fast and successful vascularization.46,47
Despite this promising potential of cocultures, individual cell types mostly require different medium components for their self-maintenance or their functionality. In addition, cell culture medium itself might modulate the cell interaction between different cell types. Thus, the cell culture conditions need to be optimized and the effects on the individual cell types must be carefully characterized. In the present study MSC and OEC were cocultivated in the individual culture media routinely used for the corresponding monocultures. We have investigated the formation of angiogenic structures, the stability of the endothelial cell population, as well as the release of pro-angiogenic growth factors to assess the impact of the cell culture medium on endothelial cell function. Further, the level of osteogenic differentiation was compared. Our study reveals significant differences in endothelial cell population stability and vascular function depending on the cell culture medium, which seems to correlate with the secretion of angiogenic factors in the context of the specific coculture conditions.
Materials and Methods
Isolation and expansion of MSCs
Human MSCs were isolated from bone marrow residues loosely associated with human cancellous bone fragments in accordance with rules of the local ethics committee. Bone fragments were washed three times with phosphate-buffered saline (PBS) to collect the loosely associated cell fractions. The washing solution containing loosened bone marrow was filtered (pore size 70 μm) and centrifuged, and the cell pellet was resuspended in Dulbecco's modified Eagle's medium (DMEM-Ham F12; Gibco Life Technologies, Karlsruhe, Germany) supplemented with 20% fetal calf serum (FCS; Gibco Life Technologies), and 1% Penicillin/Streptomycin (Pen/Strep liquid; Gibco Life Technologies). Cells were seeded at a ratio of 2×106 cells/cm2 on collagen (collagen type-I; BD Biosciences, Heidelberg, Germany)-coated flasks (T75). After 7–10 days, colonies of MSC appeared. After reaching confluence MSC were expanded using a splitting ratio of 1:3 and were subsequently cultivated in DMEM-Ham F12 supplemented with 10% FCS and 1% Penicillin/Streptomycin according to several studies.48,49 MSC in passage two were cultivated in osteogenic differentiation medium (ODM) for 2 weeks to induce osteogenic differentiation before starting monoculture and coculture experiments. ODM consists of DMEM-Ham F12 supplemented with 10% FCS, 1% Penicillin/Streptomycin, 10 mM β-glycerol phosphate (Sigma-Aldrich, Steinbach, Germany), 50 μM ascorbate-2-phosphate (Sigma-Aldrich), and 0.1 μM dexamethasone (Sigma-Aldrich).
Isolation and expansion of OECs
OEC were isolated from peripheral blood buffy coats in accordance with local ethical committee directives, including the consent of the individual donors. Mononuclear cells were separated by ficoll (Sigma-Aldrich) density centrifugation as previously published 33 and seeded on 24-culture-well plates coated with collagen type-I (BD Biosciences) at a density of 5×106/well. Cells were cultivated in EGM2 BulletKit (endothelial cell growth medium-2 BulletKit, CC-3162; Lonza, Verviers, Belgium) supplemented with 5% FCS (Gibco Life Technologies), 1% Pen/Strep (Gibco Life Technologies), and growth factors provided by the manufacturer in a humidified atmosphere (37°C, 5% CO2). After 3–4 weeks single colonies of OEC with cobblestone-like morphology and a series of endothelial markers such as CD31 and CD146 appeared. After reaching confluence, OEC were seeded on fibronectin-coated (Millipore, Schwalbach, Germany) plates and expanded over several passages using a splitting ratio of 1:2. For further details on OEC, please refer also to Ref. 50
Coculture of MSC and OEC
For cocultivation, MSC were seeded on 24-culture-well plates coated with fibronectin (Millipore) at a density of 300,000 cells/well and cultivated in ODM. For immunofluorescence staining and quantification of mineralization, MSC were seeded on fibronectin-coated Thermanox coverslips (Nunc, Langenselbold, Germany). After 24 h OEC were added at a density of 200,000 cells/well. The used cell density and the ratio 3:2 for MSC/OEC for coculture experiments was chosen due to previously establish protocols.31,51 Cocultures were either cultivated in EGM2 or in ODM for 2 weeks. After the 2 weeks the influence of the different medium on the process of angiogenesis and osteogenic differentiation was investigated. To analyze the influence of the different culture media on MSC monoculture, MSC were seeded at a density of 300,000 cells/well corresponding to the coculture seeding ratio.
Immunofluorescence staining
After 2 weeks cocultures of MSC and OEC cultured in different media were investigated by immunofluorescent staining for the endothelial marker CD31. For this purpose, cells were fixed with 3.7% paraformaldehyde (Merck, Darmstadt, Germany) and permeabilized using 0.1% Triton® X-100 (Sigma-Aldrich). For staining, anti-human CD31 (M0823; Dako, Hamburg, Germany) was diluted 1:50 in 1% bovine serum albumin (Sigma-Aldrich) in PBS and incubated for 1 h at room temperature. After washing with PBS, cells were labeled with the corresponding secondary antibody: Alexa 488 anti-mouse (A-11029; MoBiTec, Göttingen, Germany) diluted 1:1000 in 1% bovine serum albumin in PBS. After incubation for 1 h at room temperature, cells were washed and cell nuclei were counterstained with Hoechst, diluted 1:1000 (Sigma-Aldrich), and incubated for 20 min at room temperature.
In addition, after 2 weeks cocultures of MSC and OEC in ODM and EGM2 were snap-frozen in liquid nitrogen and sectioned at a thickness of 10 μm. These samples were permeabilized using 0.1% Triton X-100 (Sigma-Aldrich) and stained for the endothelial marker vWF and the pericyte-associated marker smooth muscle actin. For staining, anti-human vWF (A0082; Dako) was diluted 1:8000 and sm-actin (M0851; Dako) was diluted 1:100 in 1% bovine serum albumin (Sigma-Aldrich) in PBS and samples were incubated for 30 min at room temperature. After washing with PBS, cells were labeled with the corresponding secondary antibody; Alexa 488 anti-mouse (A-11029; MoBiTec) and Alexa 594 anti-rabbit (A-21207; MoBiTec) were diluted 1:1000 in 1% bovine serum albumin in PBS. After incubation for 30 min at room temperature, cells were washed and cell nuclei were counterstained with Hoechst, diluted 1:1000 (Sigma-Aldrich), and incubated for 10 min at room temperature. All samples were mounted with Gelmount (M01; Biomeda, Foster City, CA) and analyzed using confocal microscopy (LeicaTCS-NT).
Flow cytometry analysis
Flow cytometry analysis for the characterization of isolated MSC was performed using a combination of the markers CD90, CD29, CD44, CD271, CD45, and CD34 as previously published.12,52 In addition, the percentage of total cells stained positive for the endothelial cell markers CD146 and CD31 in cocultures of MSC and OEC cultivated in ODM and endothelial basal medium 2 (EBM2) was compared. Briefly, cells from cocultures were trypsinized (Trypsin-EDTA; Gibco Life Technologies) for 10 min at 37°C and treated with collagenase type IV (Sigma-Aldrich) for at least 20 min at 37°C to separate cells for the fluorescence-activated cell sorting analysis. After incubation, cells were fixed with 3.7% paraformaldehyde (MERCK) for 10 min at room temperature and stained with the different antibodies according to the manufacturer's protocol. Each sample was incubated with anti-CD146–PE (550315; BD Biosciences), anti-CD31-APC (130091759; Miltenyi Biotec, Bergisch-Gladbach, Germany), anti-CD90-FITC (CBL415F; Millipore), anti-CD29-FITC (559883; BD Bioscience), anti-CD271-APC (130091884; Miltenyi Biotec), anti-CD45-FITC (130080202; Miltenyi Biotec), and CD34-FITC (CBL496F; Millipore) and with isotypic control anti-IgG1-PE (555749; BD Biosciences), anti-IgG1-APC (550931; BD Biosciences), anti-IgG1-FITC (555748; BD Bioscience), or anti-IgG2a-FITC (130091837; Miltenyi Biotec) according to the manufacturer's protocol (20 min at 4°C). Analysis was performed using a FACSCalibur (BD Biosciences) and CellQuestPro software (BD Biosciences). Results depicted in a table or in histogram plots are representative for cells gained from at least three donors.
Gene expression analysis
To assess the osteogenic differentiation of MSC monocultures induced by ODM, quantitative real-time polymerase chain reaction (RT-PCR) was performed for the osteoblastic markers alkaline phosphatase (ALP), Collagen 1 (Col1), Osteocalcin, Osteonectin, and Osteopontin. In addition, the influence of EGM2 and ODM in cocultures was investigated by quantitative RT-PCR for several markers as listed in Table 1. For gene expression analysis, total RNA was extracted from MSC monocultures and cocultures using RNeasy Kit (Qiagen, Hilden, Germany) according to the manufacturer's protocol. RNA was transcribed into cDNA using Omniscript RT kit (Qiagen). For quantitative RT-PCR, Applied Biosystems 7300 Real-Time PCR System (Applera Deutschland GmbH, Darmstadt, Germany) was used. About 12.5 μL of QuantiTect™ SYBR® Green PCR Master Mix, 2.5 μL of QuantiTect™ SYBR® Green primer assay, 4 μL of RNase free water (all provided by Qiagen), and 6 μL of cDNA (1 ng/μL) were used for one reaction. Glyceraldehyde 3-phosphate dehydrogenase was used as internal control, and each gene was processed in triplicate. For amplification, the following thermocycler program was performed. Stage 1: 95°C, 15 min; stage 2: 94°C, 15 s; 55°C, 30 s; 72°C, 35 s; stage 2 was repeated for 40 cycles; stage 3 dissociation step. Relative quantification of gene expression was examined using Applied Biosystems Sequence Detection software v.1.2.2 (Applera). The gene expression levels in MSC monocultures cultivated in control medium (DMEM) were compared with ODM and the cocultures cultivated in ODM were compared with EGM-2 (reference value expression in MSC monocultures in control medium and in cocultures in ODM set to 1).
Enzyme-linked immunosorbent assay
Culture supernatants of MSC monocultures and cocultures of MSC and OEC, whether cultivated in ODM or EBM2, were collected 3, 7, 10, and 14 days of culture. The total amount of VEGF- and platelet-derived growth factor (PDGF) (concentration per 1 mg protein) was determined using ELISA DuoSets® (DY293B, DY220; R&D Systems, Wiesbaden, Germany) according to the manufacturer's protocol. Concentration was determined in triplicate at OD450 using a microplate reader (GENios plus; Tecan, Crailsheim, Germany) and a wavelength of 450 nm. Protein concentration was determined by the BCA Protein Assay Kit (23225; Thermo Fisher Scientific, Bonn, Germany) according to the manufacturer's protocol. Values for VEGF obtained in cultures in EGM2 were corrected by subtracting the basic VEGF concentration contained within the EGM2 growth medium.
Quantification of mineralization using Alizarin Red
To examine the osteogenic differentiation of MSC monocultures induced by ODM and to verify the osteogenic differentiation of MSC and cocultures of MSC and OEC in ODM or EGM2, alizarin red staining was used and the mineralization was quantified. As additional controls, osteogenic differentiation was also analyzed in EGM2 supplemented with β-Glycerophosphat and in EGM2-SODM, a medium containing ingredients of the bullet kit with the exception of hydrocortisone and acorbic acid, which were replaced by the differentiation factors used in the ODM medium. For a better understanding, different medium compositions are listed in Table 2. Alizarin red staining and quantification of mineralization was performed using the Osteogenesis Quantitation Kit (Millipore) according to the manufacturer's protocol. For quantitative analysis, alizarin red dye was extracted from cells and the concentration was detected in triplicate at OD405 using a microplate reader (TECAN). Mineralization was defined as Alizarin Red in μM. In cocultures consisting of MSC and OEC Alizarin Red was normalized to the DNA content of each sample. DNA content of cocultures in ODM, βGly, and SODM was determined using Quant-iT Pico Green dsDNA Assay Kit (Invitrogen, Darmstadt, Germany) according to manufacturer's protocol. Fluorescence was measured using microplate reader (Tecan) at an excitation/emission wavelength of 485/520 nm. In addition, the ratio of endothelial cells and MSC in the cocultures was analyzed by flow cytometry for the endothelial marker CD31 as described in the above section.
Except hydrocortisone and ascorbid acid.
DMEM, Dulbecco's modified Eagle's medium; FCS, fetal calf serum; ODM, osteogenic differentiation medium; EGM2, endothelial cell growth medium; EBM2, endothelial basal medium 2; FGF, fibroblast growth factor; VEGF, vascular endothelial growth factor; IGF, insulin-like growth factor; EGF, epidermal growth factor.
Determination of ALP activity
To determine the osteogenic differentiation of MSC in cocultures in different cell culture media, the ALP activity in the supernatant was examined using p-nitrophenyl phosphate (pNPP, Sigma Aldrich). About 20 μL of each sample was incubated with 60 μL of substrate solution (0.2% pNPP in 1 M Diethanolamine and 0.5 mM MgCl2, pH 9.8) for 30 min at 37°C. Then, 80 μL of stop solution (0.2 mM EDTA in NaOH, pH 8.0) was added and the absorbance of each sample was detected in triplicate at 405 nm using a microplate reader (Tecan). ALP activity was defined as mM pNP/1 μg DNA.
Statistical analysis
Significant differences in the investigated samples were determined using the nonparametric tests, Wilcoxon matched-pairs signed rank test, and the Mann–Whitney test. Statistical analysis was performed with GraphPad Prism (GraphPad Software). Statistical significance was demonstrated when the p-values were less than 0.05.
Results
Characterization of MSC using FACS analysis
Before using MSC in coculture experiments with OEC, the characteristics of isolated MSC were determined and the percentage of cells stained positively for the endothelial marker CD31 established to assess potential impurities by endothelial cells in the MSC populations. MSC were characterized for the marker CD90, CD29, CD44, CD45, and CD34 using FACS analysis as previously published.12,52 Table 3 represents quantitative data in percentage of total for each marker. Isolated cells under control conditions in DMEM-Ham F12 stained positively for the stromal cell-associated markers, CD29, CD90, and CD44, whereas the MSC precursor marker53–55 CD271 was only present in a subset of cells as indicated in Table 3. On the other hand, cells were negative for the hematopoietic stem cell marker CD34 and the hematopoetic marker CD45 (Table. 3). About 3.2% of total cells were positive for the endothelial marker CD31 (Table. 3). Based on these results, isolated cells can be identified as MSC as previously described in several studies.12,52
Significant p<0.03.
Differentiation of MSC into osteoblast lineage
To induce the differentiation of MSC into osteogenic lineage, MSC were cultivated in ODM for 2 weeks. ODM was supplemented with β-glycerol phosphate, ascorbate-2-phosphate, and dexamethasone, known as factors that promote osteoblastic differentiation and matrix mineralization.56–59
The characteristics of MSC in ODM compared to control medium were investigated using FACS analysis. The percentage of total cells stained positively for the stromal cell-associated markers CD29, CD90, and CD44 and the MSC precursor marker CD271 decreased significantly in MSC cultivated in ODM (Table. 3). The percentage of cells stained positively for the hematopoietic marker CD45, the hematopoetic stem cell marker CD34, and the endothelial marker CD31 did not change in response to ODM.
To assess the differentiation of MSC into osteoblastic lineages, the expression of the osteoblastic markers, ALP, collagen type-I, osteocalcin, osteonectin, and osteopontin, was determined by quantitative RT-PCR. Additionally, the quantitative mineralization of MSC cultivated in control medium and ODM was examined using alizarin red staining. Results showed significantly increased expression of ALP (+3.7±1.3), osteocalcin (+2.0±0.8), and osteonectin (+2.2±1.0) in MSC cultured in ODM compared to control medium (Fig. 1A). The expression of osteopontin increased slightly (1.7±1.2), whereas the expression of collagen type-I (+0.8±0.3) was comparable in MSC cultivated in ODM and control medium (Fig. 1A). The mineralization quantified by alizarin red was significantly higher in MSC cultivated in ODM (33.0±3.5 vs. 81.8±8.2 μM) (Fig. 1B).

Relative quantification of gene expression for the osteoblastic differentiation markers ALP, Col1, Osteocalcin, Osteonectin, and Osteopontin
The decrease in the percentage of stromal cell-associated markers in MSC after treatment with ODM and the increased expression of osteoblastic markers and mineralization of MSC on the other hand indicate the differentiation of MSC into an osteoblastic lineage in response to ODM.
Enhanced formation of capillary-like structures in cocultures of MSC and OEC
Cocultures consisting of MSC and OEC were cultured either in ODM or in EGM2 supplemented with several growth factors from the kit. In addition, EBM2 basal medium and ODM with the endothelial supplements from kit were used as additional controls. The formation of angiogenic structures depending on the type of culture medium was investigated by immunofluorescent staining for the endothelial marker, CD31 (green), after 2 weeks of coculture (Fig. 2). Cocultures in ODM were characterized by absence of microvessel-like structures or endothelial networks. Only single cells stained positively for CD31, but these cells did not form an interconnected endothelial cell layer (Fig. 2A). However, cocultures in EGM2 resulted in a considerable formation of angiogenic structures (Fig. 2D). These microvessel-like structures formed an extended vascular network with branching points and generated capillary tubes (Fig. 2D). In the ODM-SEC and the EBM2 basal medium single vascular structures were found, which were not comparable to the extensity and complexity of vascular structures found in EGM2 (Fig. 2B, C). In monocultures of OEC in EGM-2 no angiogenic structures were observed when cultured under standard conditions but can be induced in specific assays for angiogenesis.28,51

Immunofluorescence staining for the endothelial marker CD31 (green) in cocultures of OEC and precultivated MSC in response to ODM
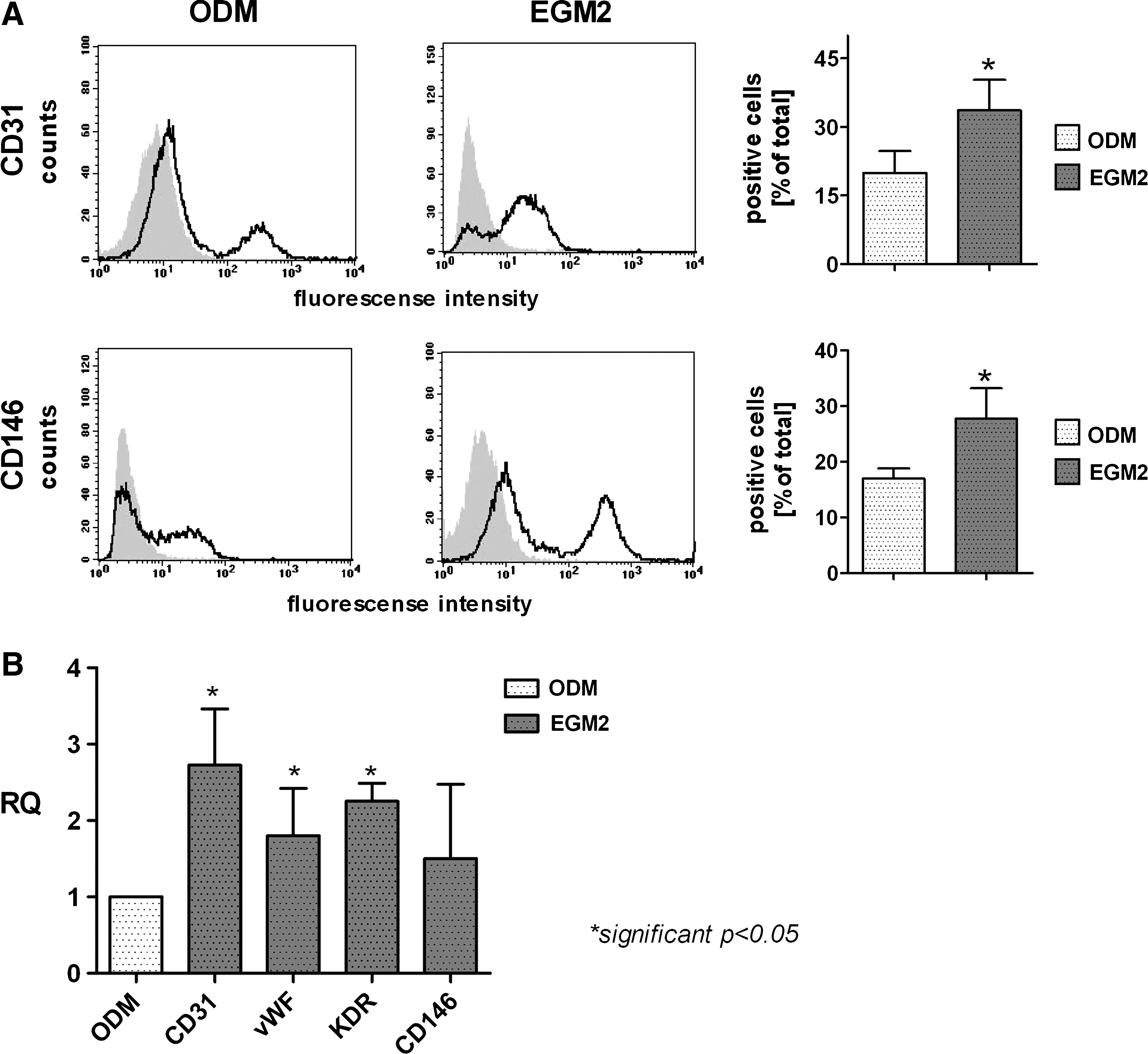
Increased endothelial characteristic in cocultures of MSC and OEC induced by EGM2
According to the results from the immunofluorescent staining for CD31 indicating the highest angiogenic activity of OEC in EGM2 and the lowest in the ODM, the relative ratio of OEC in the cocultures depending on culture medium was investigated after 2 weeks by flow cytometry for the endothelial markers CD31 and CD146. The percentage of CD31 (19.9±15.0 vs. 33.6±21.0) and CD146-positive cells (17.0±5.2 vs. 27.7±15.3) is about twice as high in cocultures in EGM2 than in ODM (Fig. 3A). Moreover, results of quantitative RT-PCR showed a significantly increased expression of the endothelial markers CD31 (+2.7±1.5), vWF (+1.8±1.2) and KDR (+2.3±0.6) in cocultures in EGM2 (Fig. 3B). The expression of the additional endothelial marker CD146 (+1.5±1.9) was only marginally increased in response to cultivation in EGM2 compared to ODM (Fig. 3B).
Analysis of angiogenic growth factors influenced by culture medium
To assess in more detail how the medium affects the formation of angiogenic structures by endothelial cells
Results from enzyme linked immunosorbent assay are depicted as relative changes in percentage in comparison to ODM after 3 days of cultivation (Fig. 4A). The release of VEGF and PDGF in EGM-2 was increased in cocultures and in MSC monocultures compared to ODM (Fig. 4A). The release of VEGF tentatively decreased in cocultures cultivated in EGM2 over the investigated time (Fig. 4A). By contrast, the release of PDGF increased in cocultures in response to EGM2 over the time period. In addition, the expression of VEGF on the m-RNA-level as analyzed by quantitative RT-PCR (Fig. 4B) was significantly upregulated in MSC monoculture as well as in coculture in EGM2 compared to ODM after 2 weeks of cultivation, whereas the expression of PDGF showed no significant changes. Further, the expression of the angiogenic factors, ANG-1 and ANG-2, showed a tendency toward upregulation in EGM2 (Fig. 4B).

Analysis of pericyte-associated markers influenced by culture media
A potential effect of the medium on the stabilization of vascular structures by pericytes was investigated by RT-PCR for the pericyte-associated markers, desmin and sm-actin. In addition, cryosections of cocultures in ODM and EGM2 were stained for sm-actin and the endothelial marker vWF, to evaluate the morphology of vessel-like structures, specifically in terms of vessel stabilization by mural cells. Immunofluorescent staining showed that cocultures in ODM are characterized by single cells stained positively for vWF (red) and sm-actin (green) (Fig. 5A). Nevertheless, in ODM no vessel like-structures or organized vessels with associated pericytes were observed. In contrast to this, cross sections of cocultures in EGM2 showed vascular lumina as indicated by the positive staining for vWF. These vessel-like structures were closely associated with cells staining positively for sm-actin (Fig. 5B). Moreover, the expression of desmin (+1.9±1.8) and sm-actin (+1.3±1.1) was tentatively upregulated in cocultures in EGM2 compared to ODM (Fig. 5C).

Analysis of osteogenic differentiation in cocultures in response to endothelial cell culture medium
The previously described results focused on the formation of angiogenic structures in cocultures of MSC and OEC in response to ODM and EGM2. In addition, the cocultivation in EGM2 might also result in effects on the osteogenic differentiation of MSC. Cocultures in EGM2 showed a significantly decreased mineralization compared to ODM as shown by quantification of alizarin red staining (Fig. 6A). This trend was also observed in different modified forms of the endothelial medium such as EGM2 supplemented with β-glycerophosphate or in EGM2 with supplements for osteogenic differentiation used in the same concentrations as in the ODM (compare Table 2). Similar results were also obtained by determining the ALP activity of cocultures cultivated in the individual medium variants (Fig. 6B) resulting in the highest ALP activity in ODM. Individual values for both ALP activity and Alizarin Red were standardized to total DNA amounts. To estimate the influence of different cell ratios on the osteogenic differentiation assays, the ratios of endothelial cells in the cocultures were determined by flow cytometry for the endothelial marker CD31. These data revealed a ratio of 23.7±16.8% endothelial cells in cocultures in ODM, 43.0%±8.6% in EGM2, 44.7%±10.3% in EGM2 plus β-Glycerophosphat, and 47.2%±7.0% in EGM2 plus SODM. These data suggested that cocultures in EGM-2 or different variations based on the endothelial culture medium result in a much lower differentiation capacity of the MSCs in the system. Similar results were also observed by semiquantitative RT-PCR (data not shown). These data might be associated to some extent with the different cell ratios under the various culture conditions.

Quantification of mineralization using Alizarin Red staining
Discussion
The well-coordinated interaction of different cell types during bone healing and vascularization might be utilized to create complex prevascularized tissues leading to rapid vascularization after implantation.1,2,60 Nevertheless, cellular interactions during angiogenesis and osteogenesis depend on factors that are far from being elucidated, but need further understanding to create tissue-engineered constructs for clinical translation. In this present study cocultures of MSC and OEC were investigated in terms of their angiogenic activation depending on the type of culture medium. The results demonstrate that the formation of angiogenic structures is improved in the endothelial medium EGM2, correlating with a higher release of angiogenic factors by the mesenchymal cells in the coculture and an increase of markers and growth factors associated with vessel stabilization by pericytes. In contrast, the differentiation of MSC toward the osteogenic lineage seems to be less effective in EGM2 and in variants based in the endothelial culture medium, when compared to the ODM.
MSC isolated from bone marrow are defined as nonhematopoietic cells (CD45−, CD14−, CD34−) expressing the markers CD90, CD105, CD73, CD29, and CD44.12,52 Recent studies described CD271 as a marker for an MSC precursor subpopulation used to enrich MSC from bone marrow.53–55 Further, it is considered that bone marrow-derived stem cells contain a population of cells expressing the hematopoietic stem cell marker CD34.61,62 In this study MSC were isolated from bone tissue with residues of associated bone marrow. These cells were characterized and positively tested for CD90, CD29, and CD44 and negative for CD45 and CD34. However, only about 50% of MSC were positive for CD271. These results indicate that isolated MSC possess a heterogeneous population, including subpopulations with the typical phenotype for MSC reported in the literature.12,52–55,59,61
Before using MSC in coculture experiments with OEC, MSC were treated with osteoblastic differentiation medium (ODM) to induce osteogenic differentiation. It is known that bone marrow contains osteogenic precursor cells,63,64 which proliferate and differentiate into osteoblasts in the presence of culture medium components such as ascorbic acid, β-glycerol phosphate, and dexamethasone.49,57,65,66 Accordingly, in the present study MSC showed a higher expression of the osteogenic differentiation markers ALP, osteocalcin, and osteonectin, as well as an increased mineralization after treatment with ODM. Additionally, the percentage of stromal cell- or stem cell-associated markers used to characterize undifferentiated MSC decreased after the treatment with ODM, thus indicating the differentiation of MSC into osteoblastic lineage in ODM.
Several studies have shown that MSC play also a role in angiogenesis, but the mechanisms by which MSC promote wound repair, especially angiogenesis, are currently unresolved and therefore still under investigation. In this context coculture models of MSC with endothelial cells might have a therapeutic advantage, but can be also used as advanced in vitro models to assess the impact of MSC on the vascularization process. In general, it is known that MSC in coculture with endothelial cells have a positive effect on the formation of angiogenic structures in vivo and in vitro.34,35,67 Experiments based on cocultures of MSC and endothelial cells demonstrated an increase in tubular structures and in vascular complexity, as well as in the stabilization of vascular structures by cells with characteristics of pericytes.34,35
In the present study we observed that the formation of microvessel-like structures in cocultures of OEC and MSC was markedly influenced by the culture medium. Angiogenic structures were abundantly formed in EGM-2, a special endothelial medium supplemented with hydrocortisone, FGF, VEGF, insulin-like growth factor, epidermal growth factor, heparin, and ascorbic acid. However, microvessel-like structures were absent in cocultures cultivated in ODM, DMEM/F12 supplemented with β-glycerol phosphate, ascorbate-2-phosphate, and dexamethasone, factors that promote osteoblastic differentiation.56–59 Interestingly, the ODM medium supplemented with the growth factors from the EGM2 Bulletkit, as well as the basal medium from the kit, EBM2, resulted in less prominent vascular structures compared to EGM2 and indicates a significant role of medium components on the cellular interaction. Additionally, the proportion of cells that were positive for the endothelial markers CD31 and CD146, determined by flow cytometry and the expression of diverse endothelial markers at mRNA level, were higher in cocultures cultivated in EGM2 compared to ODM. These results indicate a beneficial influence of the endothelial culture medium in promoting a higher ratio of endothelial cells in the coculture and the angiogenic activation of OEC in vitro. Although MSC are widely accepted as a natural source of angiogenic growth factors, also the cell culture medium seems to have a significant impact on the angiogenic activation of endothelial cells in the coculture.
In this context we observed an increased release of VEGF and PDGF in cocultures, as well as in MSC monocultures in response to EGM2 revealing that the secretion of pro-angiogenic growth factors is medium-dependent. Thus, the cell culture medium itself seems to influence the paracrine cell–cell interactions. It is widely accepted that MSC release proangiogenic factors such as Ang-1, bFGF, or VEGF and interact by complex paracrine mechanisms with various cell populations at the wound site.19,20,37 Several studies have shown that MSC-conditioned media enhanced the formation of tubular structures by endothelial cells using chicken chorioallantoic membrane assay in vivo or Matrigel® in vitro.38,39 Nevertheless, the production of growth factors by cells of mesenchymal origin is dependent on external factors, such as cell culture medium as demonstrated in the present study, and can be triggered by physiological conditions such as hypoxia or enhanced by morphogens such as sonic hedgehog, which plays a major role in the developmental process.31,68 Finally, cell interactions guiding angiogenesis are also mediated by direct cell–cell communication via gap junctions.44,69
Further, this study also shows that vessel stabilization is also influenced by the cell culture medium. It is widely accepted that pericytes play a crucial role in the process of vessel stabilization and are essential to built long-lasting vessels after the implantation of prevascularized constructs. Pericytes are situated in the outer wall of microvessels and have stabilizing functions relevant for angiogenesis.70–72 Studies indicate that pericytes migrate to newly formed vascular tubular structures and prevent the regression of such vessels by enwrapping them and acting as stabilizers for the endothelial cell layer. 73 The origin of pericytes is still unknown, although some investigations report that pericytes might derive from MSC that reside along abluminal surfaces of the microvasculature, where they interact directly with vascular endothelial cells.74–76 Several studies demonstrated that MSC differentiate under specific conditions into cells expressing sm-actin, desmin, calponin, and smoothelin, known as pericytes marker.34,76–79 Furthermore, it is known that pericytes express PDGF-β receptors and that MSC and endothelial cells release PDGF-BB, suggesting a paracrine interaction between pericytes, MSC, and endothelial cells.20,80,81 In the present study, we observed an increased expression of sm-actin and desmin in cocultures of MSC and OEC in response to EGM2. The increase in the expression of these smooth muscle cell-related markers correlates with increased PDGF levels in cocultures and monocultures of MSC when cultured in EGM2, indicating an effect of the medium on factors responsible for vessel stabilization. This was additionally underlined by morphological findings revealing the close association of vascular structures with mural cells as indicated in the cross sections stained for sm actin and endothelial markers.
In comparison to ODM, EGM2 and several variants based on the endothelial cell culture medium were less suitable to support the osteogenic differentiation of the MSCs in the coculture, as demonstrated by mineralization and ALP activity assays. The individual cell culture media influenced the cellular ratios of endothelial cells and MSCs within the cultures and thus might partly explain the different outcome in the osteogenic differentiation assays. Nevertheless, differences in MSC differentiation might also derive from factors in the individual growth or basal medium compositions directly influencing MSC differentiation or by factors involved in MSC-OEC interaction. Surprisingly, the supplementation of the EGM2 with the osteogenic differentiation factors, ascorbic acid, β-glycerophosphat, and dexamethasone used as in the ODM as well as supplementation of EGM2 with β-glycerophosphate was less effective in terms of osteogenic differentiation. These data are not consistent with our previous findings for cocultures of OEC with primary osteoblasts where EGM2 had no negative effects on osteogenic differentiation, but are in accordance with reports from the literature that endothelial cells reduce differentiation of MSCs. 82 Therefore, diverse effects of cocultures of endothelial cells with MSC or with more differentiated pOB cannot be excluded at present. Future studies will have to show whether mixtures of both culture media are more suitable in terms of differentiation of MSC in the cocultures or whether the addition of individual factors, which still need to be defined, results in optimal cell growth of endothelial cells and MSCs.
In conclusion, this study provides evidence that MSC functionally interact with OEC in a coculture system and support the formation of vascular structures in vitro. In previous studies based on cocultures of OEC and primary osteoblasts, we already demonstrated that such coculture constructs lead to functional vessels after implantation.83,84 However, the present study highlights the fact that the interaction between the different cell types resulting in the formation of angiogenic structures is dependent on the culture medium, at least partly mediated by influencing the paracrine interactions. The angiogenic activation seems to be associated with a higher release of the proangiogenic factors VEGF and PDGF, in cocultures and MSC monocultures, when cultured in EGM2. In addition, vessel stabilizing components, examined by the expression pattern of desmin and sm-actin, were positively influenced by EGM2.
Footnotes
Acknowledgments
The authors would like to thank BMBF (German-Chinese Young investigator group grant number 0315033), MAIFOR (funded by the University Medical Center, Johannes Gutenberg University), and Expertissues (Contract No
Disclosure Statement
No competing financial interests exist.