
Abstract
Transfection with either hypoxia-inducible factor-1α (HIF-1α) or heme oxygenase-1 (HO-1) gene can induce neovascularization in ischemic tissues. Although expression of transfected HIF-1α gene occurs rapidly, the expressed HIF-1α protein degrades quickly, limiting its therapeutic efficacy. Meanwhile, expressed HO-1 protein does not rapidly undergo degradation, but gene expression occurs a couple of days after transfection, resulting in apoptosis and a delay in angiogenesis in ischemic tissues at the incipient period of HO-1 gene transfection. We hypothesize that combined delivery of HIF-1α and HO-1 gene will enhance antiapoptosis and neovascularization in ischemic tissue compared with HIF-1α or HO-1 single-gene therapy. To test this hypothesis, ischemic mouse hindlimbs were treated with HIF-1α and/or HO-1 gene therapy. The combined gene therapy proved superior to both single-gene therapies, resulting in rapid expression of HIF-1α gene and long-term maintenance of expressed HO-1 protein. The apoptosis in the ischemic region was significantly less, and angiogenic growth factor secretion and angiogenesis were greater in the combined gene therapy than in either of the single-gene therapies. Our results suggest that a combined gene therapy of HIF-1α and HO-1 enhances the transfection of both genes and improves angiogenesis compared with either single-gene therapy.
Introduction
Gene therapy using hypoxia-inducible factor-1α (HIF-1α) or heme oxygenase-1 (HO-1) is known to promote antiapoptosis and angiogenesis in ischemic tissues.9–14 HIF-1α is a subunit of HIF-1, the major transcription factor involved in the response to hypoxia. HIF-1α upregulates specific genes participating in cell proliferation and vascular development to directly protect cells against ischemic apoptosis.9–11 Similarly, overexpression of the HO-1 gene in ischemic tissue also protects the tissue against oxidative stress produced under hypoxic conditions or inflammatory cytokine-related pathological conditions. Moreover, overexpression of HO-1 enhances blood vessel formation, which can counteract hypoxia-induced injury.12–14
However, HIF-1α gene therapy alone or HO-1 gene therapy alone may not be efficacious for therapeutic angiogenesis due to the rapid degradation of overexpressed protein or delay in exogenous gene expression.15,16 Overexpression of exogenous HIF-1α gene can induce immediate muscle cell protection from ischemic apoptosis. 16 HIF-1α expression in the ischemic tissue may be not high enough to protect the tissue from ischemic damage as a normal response to hypoxia. Further, the expressed HIF-1α protein degrades quickly. 17 Therefore, ischemic tissues do not have a sufficient number of vital tissues to express HIF-1α. Therefore, overexpression of HIF-1α through exogenous gene delivery would enhance the therapeutic efficacy in the ischemic tissue response to hypoxia. 17 Overexpression of HO-1 protein does not degrade quickly under ischemic conditions, but HO-1 gene expression only occurs a couple of days after transfection.18–20 Due to delayed expression of the HO-1 gene, ischemic apoptosis progresses in ischemic regions in the incipient period of HO-1 protein expression, 21 and the therapeutic efficacy of HO-1 gene therapy is limited.
We hypothesized that the combined delivery of the HIF-1α gene and HO-1 gene would enhance antiapoptosis and neovascularization in ischemic tissue, in comparison to either single-gene therapy. Overexpression of an exogenous HIF-1α gene should attenuate ischemic apoptosis in the ischemic region during the incipient period of gene delivery and improve HO-1 gene transfection efficacy, which would, in turn, enhance neovascularization. To test this hypothesis, ischemic mouse hindlimbs were treated with HIF-1α gene therapy alone, HO-1 gene therapy alone, or a combined gene therapy of HIF-1α and HO-1. A control group of mice was left untreated. The patterns of HIF-1α and HO-1 gene expression and apoptosis in the ischemic region were evaluated. Angiogenesis, limb survival, and the secretion of angiogenic paracrine factors were also evaluated.
Materials and Methods
Preparation of plasmid DNA
pSV-HIF-1α was prepared as described previously. 22 Briefly, the HIF1a cDNA was amplified by reverse transcription (RT)-polymerase chain reaction (PCR) using total RNA from 293 cells as a template. The sequences of the primers were as follows: 5′-ACCGACGCGTGCGGGGACCGATTCACCAT-3′ (forward primer) and 5′-GCTCTAGAATGAAATTAAGAAAAAGCTCAGTT-3′ (backward primer). The amplified HIF1a cDNA was digested with MluI and XbaI and inserted into pSI (Promega, Madison, WI), which produced pSVHIF1a. The human HO-1 cDNA (Genebank accession number: NM002133) was cloned by RT-PCR using total RNA from HEK 293 cells as a template. The sequences of the primers are as follows: forward primer, 5′-CCCAAGCTTATGGAGCGTCCGCAACCCG-3′; reverse primer, 5′-GCTCTAGAGCATTCACATGGCATAAAGC-3′. HinDIII and XbaI sites were incorporated into the forward and reverse primers, respectively, for cloning convenience. Amplified HO-1 was inserted into pEpo-SV-Luc (pGL3-promoter, Promega) replacing luciferase and resulting in the construction of pEpo-SV-HO-1. Plasmids were confirmed by restriction enzyme analysis and direct sequencing. Plasmids were transformed in Escherichia coli DH5α and amplified in Terrific Broth overnight at 37°C and at 220 rpm. Plasmids were purified using the Maxi plasmid purification kit (Qiagen, Valencia, CA).
Mouse hindlimb ischemia model
Hindlimb ischemia was induced in mice as previously described.23–25 Four-week-old female athymic mice (20–25 g body weight; Orient, Seoul, Korea) were anesthetized with xylazine (10 mg/kg) and ketamine (100 mg/kg). The femoral artery and its branches were ligated through a skin incision using a 6-0 silk suture (Ethicon, Somerville, NJ). The external iliac artery and all of the upstream arteries were then ligated. The femoral artery was excised from its proximal origin as a branch of the external iliac artery to the distal point where it bifurcates into the saphenous and popliteal arteries. All animal treatments and experimental procedures were approved by the Institute of Laboratory Animal Resources in Seoul National University (Institutional Animal Care and Use Committee No. SNU-100203-3).
Treatment of limb ischemia
Immediately after arterial dissection, the mice were randomly divided into six groups (n = 10). Normal mice without arterial dissection served as a positive control. Untreated mice served as a negative control. Empty plasmid and gene-delivering plasmids was suspended in phosphate buffered saline and injected intramuscularly into four sites of the gracilis muscle in the medial thigh with (12.5 μg plasmid/site) or without (25 μg plasmid/site) same amount of HIF-1α gene. HIF-1α gene alone was also injected. The physiological status of the ischemic limbs was followed up to 4 weeks after treatment.
Western blot analysis
Hindlimb muscles were sampled by cutting muscle from the 6-0 silk sutured marks left in proximal origin of ligated femoral arteries to knee joints (∼2 cm × 1 cm). The sampled muscles were divided into several pieces and used for Western blot analyses. Samples obtained from the mouse hindlimb muscles were homogenized using a Dounce homogenizer (50 strokes, 4°C) in ice-cold lysis buffer (15 mM Tris HCl, pH 8.0, 0.25 M sucrose, 15 mM NaCl, 1.5 mM MgCl2, 2.5 mM ethylenediaminetetraacetic acid, 1 mM ethylene glycol-bis(beta-aminoethyl ether)-N,N,N′,N′-tetraacetic acid (EGTA), 1 mM dithiothreitol, 2 mM NaPPi, 1 μg/mL pepstatin A, 2.5 μg/mL aprotinin, 5 μg/mL leupeptin, 0.5 mM phenymethyl sulfonyl fluoride, 0.125 mM Na3VO4, 25 mM NaF, and 10 μM lactacystin). Protein concentration was determined by the micro-bicinchonic acid method of Pierce (Rockford, IL). Western blot analysis was carried out with 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis. After transfer to an Immobilon-P membrane (Millipore, Billerica, MA), the samples were detected with antibodies against HIF-1α (Bethyl Laboratories, Montgomery, TX), HO-1 (Assay designs, Ann Arbor, MI), CD68 for inflammation detection (Abcam, Cambridge, United Kingdom), VEGF (Abcam), FGF2 (Transduction Laboratories, Lexington, United Kingdom), and hepatocyte growth factor (Sigma, St. Louis, MO) and a subsequent incubation with a horseradish peroxidase-conjugated secondary antibody (Santa Cruz Biotechnology, Santa Cruz, CA) for 1 h at room temperature. The blots were developed with an enhanced chemiluminescence detection method (Amersham Bioscience, Piscataway, NJ).
Reverse transcription polymerase chain reaction
Hindlimb muscles were sampled by cutting muscle from the 6-0 silk sutured marks left in proximal origin of ligated femoral arteries to knee joints (∼2 cm × 1 cm). The sampled muscles were divided into several pieces and were homogenized and lysed in TRIzol reagent right after sampling, and RT-PCR was performed. Each sample was homogenized and lysed in TRIzol reagent (Invitrogen, Carlsbad, CA). Total RNA was extracted with chloroform (Sigma) and precipitated with 80% (v/v) isopropanol (Sigma). After the supernatant was removed, the RNA pellet was washed with 75% (v/v) ethanol, air-dried, and dissolved in 0.1% (v/v) diethyl pyrocarbonate-treated water (Sigma). RNA concentration was determined by measuring the absorbance at 260 nm using a spectrophotometer. Reverse transcription was performed using 5 μg of pure total RNA and SuperScriptTM II reverse transcriptase (Invitrogen), and the synthesized cDNA was amplified by PCR. The sequences of the primers for p53 were 5′-ATGACTGCCATGGAGGAGTCACAG-3′ (forward primer) and 5′-TGAGAAGGGACAAAAGATGACAGG-3′ (backward primer), Bcl-xl were 5′-TGGAGTAAACTGGGGGTCGCATCG-3′ (forward primer) and 5′-AGCCACCGTCATGCCCGTCAGG-3′ (backward primer), CD31 were 5′-GACCCAGCAACATTCACAGAT-3′ (forward primer) and 5′-TCTTTCACAGAGCACCGAAGT-3′ (backward primer), smooth muscle alpha actin (SM α-actin) were 5′-TGTGTGGACTCTGGAGATG-3′ (forward primer) and 5′-CTTCTGCATCCTGTCAGCAA-3′ (backward primer), and for MyoD were 5′-TGGGCGATCTGTCAAAGGCGATCTGTCAA-3′ (forward primer) and 5′-AGTTGATCCGTATGCAAGAGTATTCTCATGCACTCTT-3′ (backward primer). PCR was carried out for 35 cycles of denaturing (94°C, 30 s), annealing (58°C, 45 s), and extension (72°C, 45 s) with a final extension at 72°C for 10 min. PCR products were observed by electrophoresis on a 2% (w/v) agarose gel and ethidium bromide staining. The products were analyzed with a gel documentation system (Gel Doc 1000; Bio-Rad, Hercules, CA). β-actin served as an internal control.
Immunohistochemistry
Ischemic limb muscles harvested 28 days after treatment were embedded in optimal cutting temperature compound (O.C.T. compound, TISSUE-TEK® 4583; Sakura Finetek USA, Torrance, CA), frozen, and cut into 10 μm-thick sections at −22°C. All three samples per group were completely sectioned and six slides were selected from the beginning, middle, and end part of each sample. Thirty different pictures were randomly selected from slides stained for caspase-3 for caspase-3-positive cells/4,6-diamidino-2-phenylindole (Vector Laboratories, Burlingame, CA)–positive cells quantification. For the capillaries in ischemic regions, sections were subject to immunofluorescent staining with anti–von Willebrand factor (vWF). Twenty different images per slide from three different samples with ×200 magnification were randomly analyzed. Each image was ∼650 × 550 μm. vWF-positive vessels with single-layered round morphology were counted and calculated. Sections of the capillaries and arterioles in ischemic tissue were immunofluorescently stained with anti-vWF (Abcam) and anti-SM α-actin (Abcam), respectively. Primary antibodies against mouse-specific VEGF (Abcam) were used to examine expression of this angiogenic factor in ischemic tissues. The staining signals for vWF, SM α-actin, and VEGF were observed with FITC-conjugated secondary antibodies (Jackson ImmunoResearch Laboratories, West Grove, PA). The sections were counterstained with 4,6-diamidino-2-phenylindole and examined using a fluorescence microscope (Nikon TE2000, Tokyo, Japan).
Histology
Ischemic limb muscles were retrieved 3, 7, 14, and 28 days after treatment. Specimens were fixed in formaldehyde, dehydrated with a graded ethanol series, and embedded in paraffin. Specimens were sliced into 4-μm sections and stained with hematoxylin and eosin to examine muscle degeneration and tissue inflammation. Masson's trichrome collagen staining was also performed to assess tissue fibrosis in the ischemic regions.
Statistical analysis
Quantitative data are expressed as the mean ± standard deviation. Statistical analysis was performed by analysis of variance using a Bonferroni test. A p-value <0.05 was considered statistically significant.
Results
HIF-1α and HO-1 gene expression
Expression of the HIF-1α and HO-1 genes was enhanced with combined gene therapy (Fig. 1). Western blot analysis showed that human HIF-1α was expressed in the ischemic region after gene delivery, but the amount of human HIF-1α protein in the ischemic region decreased with time. The amount of HIF-1α protein was greater with the combined gene therapy than with single-gene therapy at days 3 and 7. The HO-1 gene was not expressed until 3 days after gene delivery. On day 3, the amount of HO-1 protein was greater with the combined gene therapy than with the single-gene therapy.
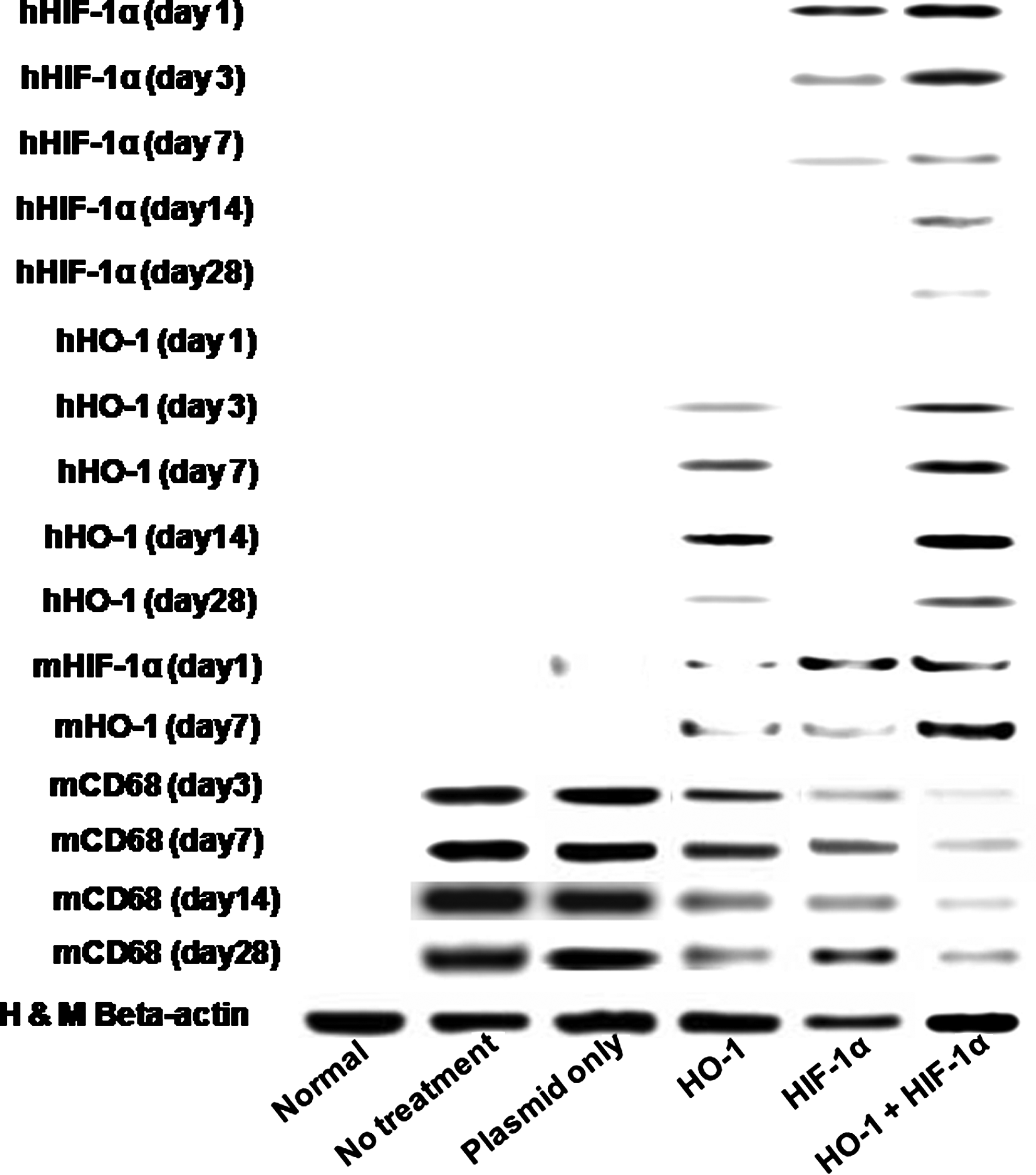
Representative images of Western blot analysis of human HIF-1α and human HO-1 expression in mouse ischemic hindlimbs at 1, 3, 7, and 28 days and mouse HIF-1α and mouse HO-1 expression at 1 and 7 days, mouse CD68 expression after treatments (no treatment, HIF-1α gene therapy alone, HO-1 gene therapy alone, or combined gene therapy of HIF-1α and HO-1). HIF-1α, hypoxia-inducible factor-1α; HO-1, heme oxygenase-1.
Apoptosis
The combined gene therapy of HIF-1α and HO-1 resulted in less apoptosis in ischemic mouse hindlimbs than either single-gene therapy (Fig. 2). Three days after induction of ischemia and gene delivery, apoptosis in the ischemic tissue was detected by caspase-3 staining. Delivery of either the HO-1 or HIF-1α gene decreased the number of apoptotic cells. The number of apoptotic cells on day 3 was significantly less with the combined gene therapy than with HO-1 single-gene therapy (Fig. 2A, C). However, there was no difference in the number of apoptotic cells between the combined gene therapy and HIF-1α single-gene therapy on day 3. On day 7, there were fewer apoptotic cells with the combined gene delivery than with either single-gene therapy (Fig. 2B, D).
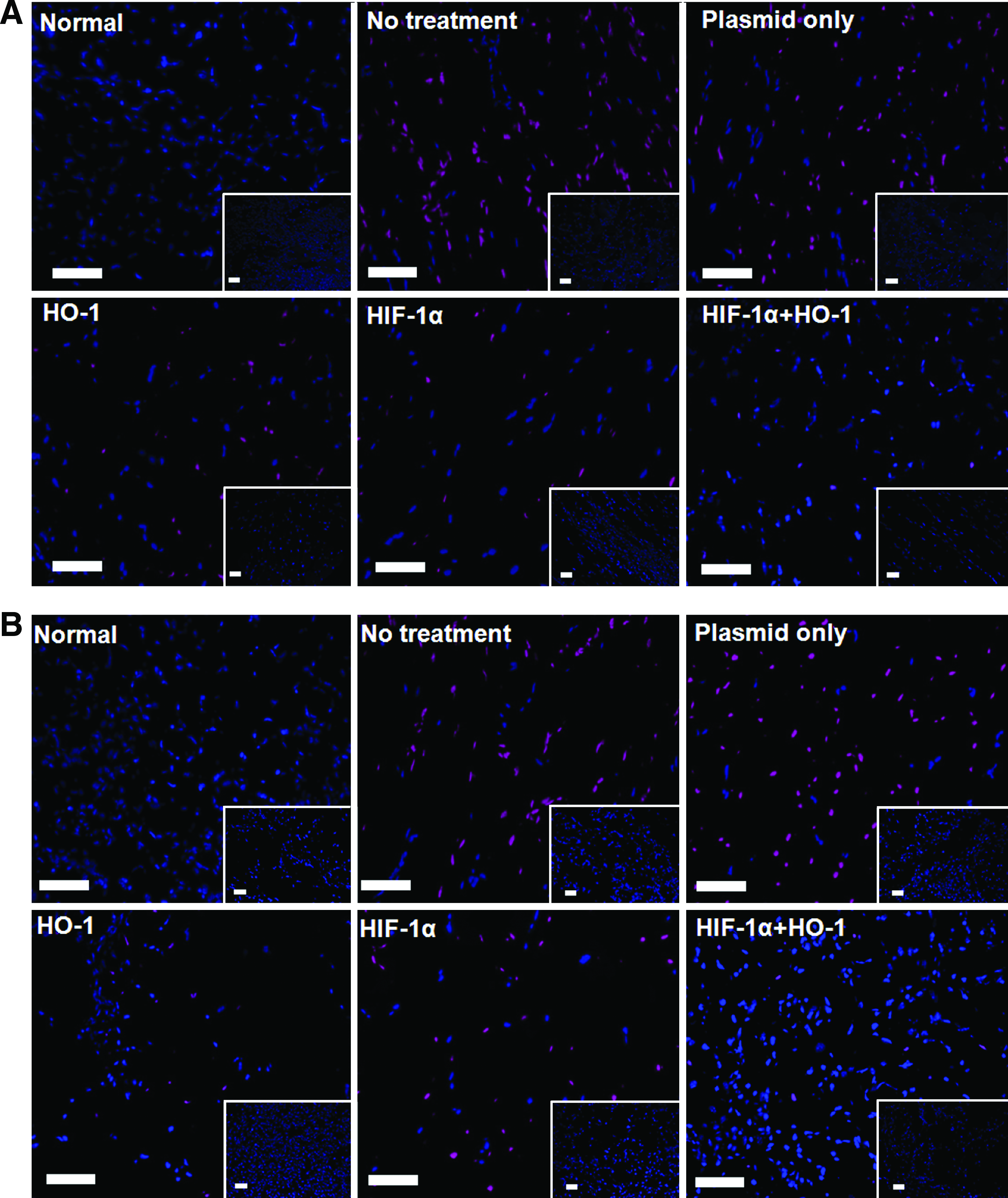
Apoptosis in mouse ischemic hindlimbs at 3 and 7 days after no treatment, plasmid only, HIF-1α gene therapy alone, HO-1 gene therapy alone, or combined gene therapy of HIF-1α and HO-1. Caspase-3 staining results at
RT-PCR analysis for pro-apoptotic and antiapoptotic gene expression was consistent with the caspase-3 staining results (Fig. 2E). mRNA expression of Bcl-xl, an antiapoptotic gene, was greater and that of p53, a pro-apoptotic gene, was less with the combined gene therapy than with either single-gene therapy on day 3. Delivery of either the HO-1 or HIF-1α gene resulted in decreased p53 expression and increased Bcl-xl expression. Expression of p53 and Bcl-xl decreased and increased, respectively, with the combined gene therapy, compared with either single-gene therapy. With HIF-1α gene therapy, p53 increased with time and Bcl-xl decreased with time with HIF 1α treatment alone.
Angiogenesis
The combined gene therapy enhanced angiogenesis in ischemic limbs as compared with either single-gene therapy alone (Fig. 3). Immunofluorescent staining for mouse vWF and quantification of capillary density at day 28 revealed that the combined gene therapy significantly enhanced (p < 0.05) capillary density (98.4 ± 20.7/mm2) as compared with HIF-1α single-gene therapy (45.2 ± 9.3/mm2) or HO-1 single-gene therapy (25.3 ± 12.1/mm2) (Fig. 3). Immunofluorescent staining for mouse SM α-actin and quantification of arteriole density revealed that arteriole density was significantly enhanced (p < 0.05) with the combined gene therapy (13.2 ± 4.6/mm2) as compared with HIF-1α single-gene therapy (3.3 ± 1.5/mm2) or HO-1 single-gene therapy (2.2 ± 1.5/mm2) (Fig. 3). RT-PCR for mouse specific capillary, arteriole, and myoblast showed that capillary and arteriole appearance and growth were increased with the combined gene therapy as compared with HIF-1α single-gene therapy or HO-1 single-gene therapy (Fig. 3E).
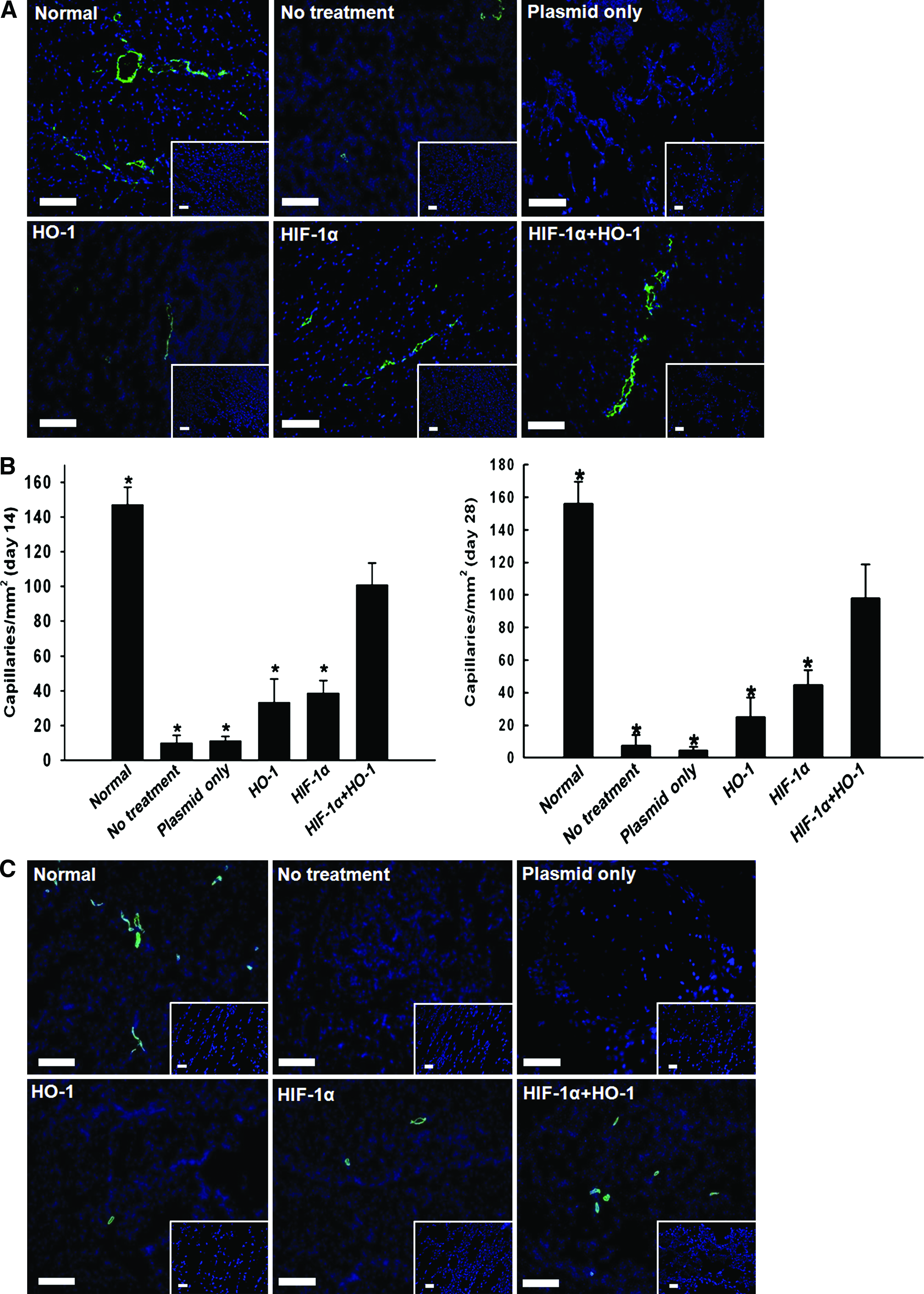
Capillary density, arteriole density, and angiogenic growth factor expression in mouse ischemic hindlimbs 28 days after no treatment, plasmid only, HIF-1α gene therapy alone, HO-1 gene therapy alone, or combined gene therapy of HIF-1α and HO-1.
Angiogenic factor expression
The combined gene therapy enhanced expression of angiogenic paracrine factors in ischemic tissues 28 days after treatment (Fig. 3). Immunofluorescent staining for VEGF revealed that VEGF secretion was greater in mice treated with combined gene therapy than in mice treated with either single-gene therapy (Fig. 3F). Western blot analysis for angiogenic paracrine factors, such as FGF2, hepatocyte growth factor, and VEGF, confirmed that angiogenic factor expression in ischemic muscles was enhanced by the combined gene therapy as compared with either single-gene therapy (Fig. 3G).
Muscle degeneration and fibrosis
Histological examination of ischemic limbs retrieved 28 days after treatment indicated that delivery of either the HIF-1α gene or HO-1 gene protected limb muscles from ischemic apoptosis (Fig. 4). Hematoxylin and eosin and Masson's trichrome staining of untreated animals (negative control) revealed serious muscle degeneration in the ischemic tissue and inflammation after ischemia was induced (Fig. 4). In contrast, the muscles of ischemic limbs were notably protected from degeneration and inflammation after treatment with the combined gene therapy of HIF-1α and HO-1. HIF-1α gene therapy alone and HO-1 gene therapy alone also attenuated muscle degeneration and tissue inflammation. However, the attenuation of muscle degeneration and inflammation was more prominent with the combined gene therapy. Western blot analysis for CD68 showed that inflammation was decreased and subsided earlier with the combined gene therapy as compared with HIF-1α single-gene therapy or HO-1 single-gene therapy (Fig. 1).

Histological analysis and hindlimb survival in mice with hindlimb ischemia at 3, 14, and 28 days with no treatment, plasmid only, HIF-1α gene therapy alone, HO-1 gene therapy alone, or combined gene therapy of HIF-1α and HO-1.
Limb salvage
A lack of treatment resulted in rapid and extensive necrosis in ischemic hindlimbs, often leading to complete limb loss via autoamputation by 28 days (Fig. 4). Untreated mice underwent limb loss (85.7%, six out of seven) or severe limb necrosis (14.3%, one out of seven) without limb salvage. Both HIF-1α single-gene therapy and HO-1 single-gene therapy attenuated limb necrosis after treatment, but over 57.14% and 28.6% of the mice treated with these therapies, respectively, ultimately underwent limb loss by 28 days. Combined gene therapy significantly reduced the rate of limb loss in comparison to other treatment groups. Most mice treated with the combined gene therapy either retained their limbs (42.9%, three out of seven) or exhibited necrosis limited to the foot (42.9%, three out of seven). Only one animal lost its limb (14.3%, one out of seven).
Discussion
With combined gene therapy, expression of HIF-1α protein rescued muscle apoptosis before expression of the simultaneously delivered HO-1 gene (Fig. 2A, C, E) and enhanced the transfection efficiency of HO-1 gene at day 3 (Fig. 1). While exogenous HIF-1α protein was secreted within a day after gene delivery, expression of the HO-1 gene occurred on day 3 (Fig. 1), which is consistent with a previous report. 18 During the incipient period of HO-1 gene delivery, when HO-1 is not expressed, severe apoptosis occurs in ischemic tissue (Fig. 2A, C, E), which decreases the HO-1 gene transfection efficiency (Fig. 1). In contrast, apoptosis in the ischemic region was significantly reduced during the incipient period with the combined gene therapy, which is likely due to HIF-1α expression during the incipient period (Fig. 1), as compared with HO-1 single-gene therapy (Fig. 2A, C, E). The enhanced HO-1 transfection efficiency with combined gene therapy, as compared with single-gene therapies, was likely due to enhanced muscle cell viability resulting from HIF-1α gene expression. Without HIF-1α gene delivery, hindlimb muscle cells underwent severe ischemic apoptosis during the incipient period (day 3) of ischemia (Fig. 2A, C, E). Mouse HIF-1α was also expressed at considerably high levels at day 1 in both HIF-1α single-gene therapy group and the combined gene therapy group (Fig. 1). This could be due to ischemic tissue protection by human HIF-1α gene therapy. Although HO-1 gene expression resulted in reduced ischemic apoptosis, the simultaneously transfected HIF-1α gene played a more critical role during the incipient period of ischemia (Fig. 2A, C, E). At day 7, apoptosis in the ischemic region was significantly less with the combined gene therapy than with the HIF-1α single-gene therapy (Fig. 2B, D, E). Since HIF-1α protein degrades rapidly, 16 the amount of expressed HIF-1α protein decreased with time with HIF-1α gene therapy (Fig. 1), resulting in increased apoptosis in the ischemic region on day 7 as compared with day 3 (Fig. 2). With the combined gene therapy, high level expression of HO-1 could compensate for the low level of HIF-1α protein on day 7 (Fig. 1), resulting in decreased apoptosis with the combined gene therapy compared with HIF-1α single-gene therapy on day 7 (Fig. 2B, D, E). Mouse HO-1 was also expressed at a high level at day 7 in the combined therapy group (Fig. 1). This could be due to ischemic tissue protection from ischemic damage by human HIF-1α at the initial period and by human HO-1 at later stage.
The combined gene therapy enhanced the angiogenic efficacy compared with either single-gene therapy. The combined gene therapy also significantly increased capillary and arteriole densities (Fig. 3), and paracrine factor secretion (Fig. 3F, G), as compared with the single-gene therapies. Ischemic limb salvage was also improved in the combined gene therapy (Fig. 4). This may be partially due to greater expression of the HO-1 gene with the combined gene therapy than with the single-gene therapies. Enhanced expression of angiogenic growth factors in the ischemic tissue (Fig. 3) is another contributor to the enhanced angiogenesis observed after combined gene therapy. HIF-1α and HO-1 induce VEGF expression.26–28 The combined HIF-1α and HO-1 gene therapy also enhanced angiogenic growth factor expression (Fig. 3), and this may partially be due to the increased efficiency of HO-1 gene transfection and decreased apoptosis in the ischemic region. High levels of pro-angiogenic factor (i.e., VEGF) expression in unaffected blood supply mice (normal mice) were also found in previous studies.29,30
A sustained delivery of HIF-1α gene could improve the therapeutic angiogenesis efficacy as compared with injection of HIF-1α gene-harboring plasmid in solution. Since HIF-1α protein expressed from exogenous HIF-1α gene degrades fast, HIF-1α gene therapy alone may not exert substantial therapeutic angiogenesis. A sustained delivery of HIF-1α using a polymeric scaffold could circumvent the rapid degradation of HIF-1α and lead to a more sustained supply of the protein, which may improve its efficacy independent of co-delivery of HIF-1α and HO-1gene. 31
In comparison to single-gene therapies, the combined gene therapy of HIF-1α and HO-1 in ischemic hindlimbs protects hindlimb muscles from ischemic apoptosis and enhances angiogenesis and limb survival. HIF-1α gene delivery rescues limb muscle apoptosis before expression of a simultaneously delivered HO-1 gene and potentially enhances HO-1 gene transfection efficacy. This study shows that a combined gene therapy of HIF-1α and HO-1 is more effective than either single-gene therapy in terms of the angiogenic efficacy with ischemic disease.
Footnotes
Acknowledgments
This work was supported by a grant (#A050082) from the Korean Health 21 R&D Project, Ministry of Health, Welfare and Family Affairs, Republic of Korea, and a grant (#SC3220) from the Stem Cell Research Center of the 21st Century Frontier Program, the Ministry of Education, Science and Technology, Republic of Korea.
Disclosure Statement
No competing financial interests exist.