
Abstract
After the clinical insertion of a bone biomaterial, the surrounding osteoblasts would migrate and attach to the implant surface and foster a microenvironment that largely determines the differentiation fate of the comigrated mesenchymal stem cells. Whether the fostered microenvironment is suitable for osteogenic differentiation of mesenchymal stem cells is critical for the subsequent osseointegration. In this study, we determined (1) how the spherical or rod-shaped hydroxyapatite nanoparticles (nHA) incorporated poly(ɛ-caprolactone) (PCL) films (PCL-spherical nHA, PCL-rod nHA) interact with primary human osteoblasts (HOBs); (2) how the microenvironment rendered by their interaction affects osteogenic differentiation of adipose tissue-derived mesenchymal stem cells (ASCs). HOBs were seeded on PCL, PCL-spherical nHA, and PCL-rod nHA films, respectively. When cultured alone, the HOBs on PCL-rod nHA films showed most efficient osteoblastic differentiation compared with those on PCL or PCL-spherical nHA films. When cocultured with ASCs in an indirect coculture system, only the HOBs on PCL-rod nHA films up-regulated the gene expression of Runx2, bone sialoprotein, and osteocalcin of ASCs. Additionally, the HOBs on PCL-rod nHA films showed significant up-regulation of bone morphogenic protein 2 gene and protein expression and induced highest phosphorylated Smad1/5 protein level in ASCs. Treatment of the coculture medium with bone morphogenic protein 2 inhibitor (Noggin) largely abolished the osteogenic differentiation of the ASCs induced by the HOBs on PCL-rod nHA films. In conclusion, HOBs can not only best display their osteoblastic phenotype by culturing on PCL-rod nHA films but also render an optimal osteogenic niche for the differentiation of stem cells.
Introduction
MSCs are a promising cell resource for tissue-engineering purpose due to their innate capacities of selfrenewal and multipotential differentiation such as osteogenic or chondrogenic differentiation under an appropriate stimulation by hormonal and local factors.12–16 The aim of the bone tissue-engineering approach is to transplant an appropriate scaffolding biomaterial alone or combine it with the cells (i.e., MSCs) capable of osteogenic differentiation to the bone defects for repairing and regeneration. For instance, ceramic scaffolds combined with MSCs were shown to induce bone formation in critical-sized bone defect in dogs. 17 However, it is of great importance to know the underlying mechanisms that contribute to the bone formation in the presence of biomaterials.
Different cell types including osteoblasts and MSCs would migrate to the biomaterial site on bone graft implantation,18–20 and the capacity of MSCs to differentiate into functional osteoblasts is largely determined by the implant's composition, structure, and the proper soluble osteogenic signals (cytokines, growth factors, etc.) present at the site of implantation. 21 Among the factors at the implantation site, the soluble osteogenic signals are primarily stemmed from the surrounding osteoblasts and act on the MSCs in a paracrine manner. In vitro, several studies demonstrated that osteoblasts are able to direct MSCs into osteogenic differentiation in an indirect or direct cell-cell coculture system.22–24 It is well known that the surface characteristics of biomaterials including chemical composition and topography largely determine the phenotype and functionality of osteoblasts via autocrine and/or paracrine regulation.25–27 We previously demonstrated that a clinically used scaffold, β-tricalcium phosphate, exerts its osteoconductivity through a bone morphogenic protein 2 (BMP2) autocrine loop. 28 Considering the in vivo scenario of cell interactions between MSCs and osteoblasts after the biomaterial implantation, most studies in the literature failed to take the important roles of biomaterials where the osteoblasts grow into account while investigating the crosstalk between MSCs and osteoblasts.
This work mimicked the in vivo scenario, where the bone implanted biomaterial is populated by MSCs and osteoblasts. We investigated how the different shapes of nHA, when interacting with HOBs, affect their differentiation; and whether and how the microenvironment rendered by HOBs on differentiation substrates affects osteogenic differentiation of adipose tissue-derived mesenchymal stem cells (ASCs). We demonstrated that the PCL films incorporated with rod-shaped nHA (PCL-rod nHA) but not spherical nHA (PCL-spherical nHA) are able to indirectly route ASCs into osteogenic differentiation by directly interacting with HOBs through a manner of BMP2 paracrine regulation.
Materials and Methods
Preparation of PCL film and the incorporation of nHA
Preparation of nHA
The preparation of nHA with different shapes was previously described. 4 Briefly, different shapes of nHA with Ca/P ratios of 1.67 were synthesized using a precipitation reaction between aqueous mixture of ammonium phosphate dibasic ((NH4)2HPO4; Sigma Aldrich reagent grade, ≥98%) and calcium nitrate tetrahydrate (Ca (NO3)2; Sigma Aldrich, >99%) in a beaker at different temperatures for a duration of 4 h. (NH4)2HPO4 solution was added drop wise to Ca(NO3)2 solution under continuous stirring. The concentration of HA was 0.66 mM and kept constant for all the synthesis, whereas the pH was varied from 5 to 12 using buffers and the temperature was varied from room temperature to 70°C. With the change of the pH and temperature, different shapes of nHA were prepared. In all the cases, the resultant white color product was washed several times with distilled water to completely remove the buffer. After drying at 80°C, powders were heated at 450°C to remove remained solvents and buffers. The chemical composition and physical morphology of the obtained powder were determined by X-ray diffraction method and field emission electron microscopy, respectively.
Incorporation of nHA into PCL films
PCL pellets (Sigma Aldrich) were dissolved in chloroform (Sigma Aldrich) at a concentration of 10% w/v, and the same weight of spherical or rod nHA powder was dispersed in 10 mL chloroform by sonication for 15 min and then added to PCL solution. The resultant mixture was stirred for 24 h at room temperature to increase homogenization and then sonicated for 10 min just before casting. Films were prepared by casting the slurries onto glass plates and drying at room temperature under a fume hood for 24 h and then dried for 3 days in an oven at 37°C.
Surface roughness assessment
The surface roughness of the films was quantified using atomic force microscopy (AFM) (Multimode Model MMAFMLN with Nanoscope IIIa controller; Veeco Metrology). A section of the PCL film, PCL-spherical nHA, or PCL-rod nHA (the rougher surface facing upwards) was secured with double-sided adhesive tape onto a stainless steel AFM specimen disk. The film was imaged in tapping mode at a scan rate of 0.5 Hz using Budget Sensor® Tap300Al-G cantilevers (Innovative Solutions). The root mean square (Rq) roughness of six randomly selected locations on the film was obtained (n=6). Each location was 10 μm×10 μm in area with 512×512 measured points in total.
Degradation of the PCL-nHA films
To know how the degradation of nHA affects the ion microenvironment of the culture medium, we measured the Ca2+ and p3− concentrations of the cell culture medium immersed with PCL, PCL-spherical nHA, or PCL-rod nHA for 4 and 14 days. Films with same diameter (n=6 for each group) were selected, washed carefully with 100% ethanol, and then dried at 37°C for 24 h. After soaking, Ca2+ and p3− concentrations in the medium were tested by inductively coupled plasma atomic emission spectroscopy (ICP-OES, Optima 3000 DV).
HOB isolation, culturing, and attachment, and osteoblast differentiation of HOBs on different PCL films
Permission to use discarded human tissue was granted by the Human Ethics Committee of the University of Sydney, and informed consent was obtained. HOBs were isolated from normal human trabecular bone as previously described. 29 Briefly, bone was chopped into 1 mm3 pieces, washed several times in phosphate buffered saline (PBS), and digested for 90 min at 37°C with 0.02% (w/v) trypsin (Sigma Aldrich) in PBS. Digested bone chips were cultured in complete media containing α-Minimal Essential Medium (α-MEM; Gibco Laboratories), supplemented with 10% (v/v) fetal calf serum (FCS; Gibco Laboratories), 2 mM l-glutamine (Gibco Laboratories), 25 mM Hepes Buffer (Gibco Laboratories), 2 mM sodium pyruvate, 30 mg/mL penicillin, 100 mg/mL streptomycin (Gibco Laboratories), and 0.1 mM l-ascorbic acid phosphate magnesium salt (Wako Pure Chemicals). The cells were cultured at 37°C with 5% CO2, and the medium refreshed every 3 days until 80%–90% confluence.
All HOBs used in the experiments were at passage three. At 80%–90% confluence, the cells were trypsinized with the trypsinizer/EDTA (Invitrogen) and subsequently suspended in complete media. Before cell seeding, all the films were decontaminated by soaking them twice in 70% ethanol and 30 min each, followed by rinsing thrice with PBS solution and 5 min each before exposing to ultraviolet light for 30 min.
For examining HOBs attachment on the films, the HOBs were seeded on the different films at a density of 10,000 cells per cm2 for 24 h. Cells were then rinsed thrice in PBS, fixed in 1.25% glutaraldehyde, and postfixed in osmium tetroxide followed by sequential dehydration in graded ethanol. The films were then dried in hexamethyldisilizane and coated with gold for scanning electron microscopy (SEM) analysis using FE-scanning electron microscopy (Zeiss Ultra).
For the evaluation of the differentiation of HOBs on different PCL films, HOBs were seeded at a density of 10,000 cells per cm2 on PCL or PCL-spherical nHA or PCL-rod nHA films and cultured for 1 and 7 days. Osteoblastic differentiation-related gene expression (Collagen I, Runx2, bone sialoprotein, osteocalcin, and BMP2) was analyzed using quantitative real-time polymerase chain reaction (qRT-PCR), and BMP2 protein expression was examined by western blot.
Blocking BMPs signaling in coculture of ASCs and HOBs
ASCs (Invitrogen by Life Technologies) were propagated according to the manufacturer's instructions. The MesenPRO RSTM Basal Medium (Invitrogen by Life Technologies) with the supplementation of L-glutamine and MesenPRO RS Growth Supplement (Invitrogen by Life Technologies) was used for ASCs expansion, and ASCs at passage five were used in the current study. An indirect coculture system was employed to separate HOBs and ASCs by placing a cell culture insert with 0.4 μm pore-size filter (Millipore) onto a well of the six-well plate, and the PCL or PCL-spherical nHA or PCL-rod nHA films were placed on the insert. ASCs were seeded on the wells of plates at a cell density of 5000 cells per cm2, whereas HOBs were seeded at a cell density of 10,000 cells per cm2 on various films on the inserts. In this culture system, ASCs can only perceive the signals of the soluble factors released by the HOBs seeded on different films, as the HOBs on the insert can not migrate through the 0.4 μm pore-size filter and directly contact with ASCs. After they were cocultured for 4 and 14 days, ASCs were collected, and the osteogenic gene expression and Smad1 and p-Smad1/5 protein expression were determined. For BMP2 signaling blocking studies, the culture medium was replaced with media containing Noggin (500 ng/mL; Sigma).
Quantitative real-time polymerase chain reaction
At the predetermined time points (4 and 14 days), total RNA was isolated from HOBs and ASCs by adding Trizol reagent (Sigma) after the medium was removed according to the manufacturer's instructions. First-strand cDNA was synthesized from 0.7 μg total RNA using the Reverse Transcription Kit (Bioline) according to the manufacturer's instructions. Real-time PCR was performed in Rotor-Gene 6000 (Corbett Life Science) by using Immomix (Bioline) according to manufacturer's instructions, and the relative gene expression levels for Runx2, collagen type I, alkaline phosphatase, bone sialoprotein, osteocalcin, and BMP2 were obtained by normalizing them to a house-keeping gene (18S). Primers for the selected genes are listed in Table 1.
BMP2, bone morphogenic protein 2.
Western blotting
Western Blotting was performed for determining BMP2 expression in HOBs and p-Smad1/5 and Smad1 expression in ASCs. HOBs/ASCs were washed with ice-cold PBS and lysed for 30 min in ice-cold RIPA lysis buffer (20 mM Tris–HCl, pH 7.5, 1 mM EDTA, 1 mM EGTA, 150 mM NaCl, and 1% Triton X-100, protease inhibitor cocktail [Sigma], and phosphatase inhibitor cocktail 2 [Roche]). Total protein concentration of the sample was measured using the BCA protein assay kit (Bio-Rad). Equal aliquots of protein (15 μg) were heated at 70°C for 10 min in 4×sample buffer (WesternBreeze; Invitrogen) and 10× reducing buffer, and separated on 8%–12% SDS-PAGE gels (WesternBreeze; Invitrogen). The proteins were then transferred to PVDF membranes, washed with 1× TBS-T (20 mM Tris–HCl pH 7.6, 137 mM NaCl, containing 0.1% Tween 20), and blocked for 1 h at room temperature in 1× TBS-T with 1% BSA. The membranes were washed thrice followed by incubation with different primary antibodies (antiβ actin [1:5000; Abcam], antiBMP2 [1:5000; Abcam], antiSmad1 [1:1000; Cell signaling], and antip-Smad1/5 [1:500], cell signaling, respectively) in TBS-T containing 1% BSA overnight at 4°C. After three washes, the membranes were incubated with secondary antibody (WesternBreeze; invitrogen) for 60 min, followed by another three washes before the protein bands were visualized with chemiluminescent reagents (WesternBreeze; invitrogen) in Alpha Innotech Digital Imaging System (Alpha Innotech).
Statistical analysis
The data in this study were obtained from at least four independent experiments and represented with Mean±SE. For statistical analysis, Levene's test was first performed to determine the homogeneity of variance for all the data, and then Tukey HSD post-hoc test was used for the data with homogeneous variants, otherwise Tamhane's T2 Post Hoc test was employed for analysis. SPSS 17.0 program was employed for all statistical analysis, and differences were considered significant if p<0.05.
Results
Film surface roughness and ion release measurements
Surface roughness analysis using AFM showed all three film surfaces possessing relatively smooth patches as well as ridges and grooves (Fig. 1A–C); the distribution of which appeared to be random, with no statistically significant difference in the Rq between the films (Fig. 1D).

Surface roughness analysis and measurement of Ca2+ and P3− concentrations in the cell culture medium immersed with different films. The surfaces of PCL
There were no significant differences in the concentrations of the released Ca2+ and p3− from PCL, PCL-spherical nHA, and PCL-rod nHA films soaked in cell culture medium for 4 days. However, after 14 days of soaking, lower Ca2+ and p3− concentrations were found in the medium soaked with PCL-spherical nHA and PCL-rod nHA films, compared with that of PCL films, but there was no difference between the medium soaked with PCL-spherical nHA and PCL-rod nHA films (Fig. 1E).
SEM analysis of nHA and HOBs attachment
SEM images showed that spherical (Fig. 2B) or rod-shaped (Fig. 2C) nHA well dispersed in PCL film with no apparent aggregation formation. The densities of spherical and rod-shaped nHA were consistent with an average of 25 nHA per microscopic ken (5000× magnification), and the average sizes of spherical and rod-shaped nHA were 50 nm in diameter, 20 nm in width, and 100 nm in length, respectively. Figure 2A showed the PCL film without any incorporation of nHA. The HOBs attached and spread well on all three different films after 24 h of seeding, and they displayed triangular or tubular cell shape (Fig. 2D–F).
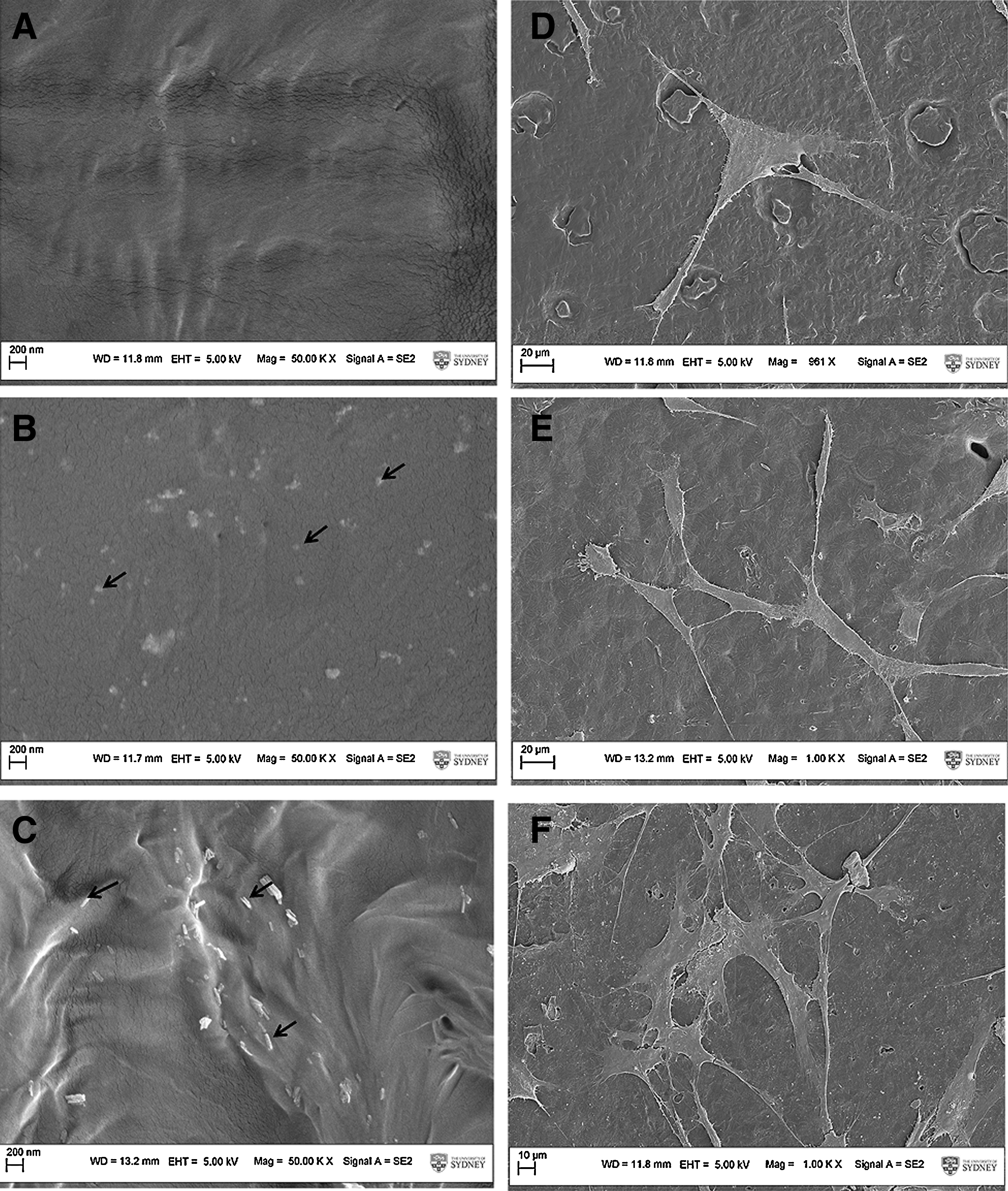
Examinations of nHA shapes and their effects on HOBs attachment. The images showed that either spherical or rod shape of nHA were well dispersed on the surface of PCL films. Arrows indicate the nanoparticles.
Effects of nHA incorporations on HOB differentiation
We next examined whether the shape of nHA affects HOB differentiation. HOBs were seeded on either control PCL films, PCL-spherical nHA, or PCL-rod nHA films for 1 and 7 days. Gene expression of Runx2, collagen type I, bone sialoprotein, and osteocalcin was determined using qRT-PCR. The incorporation of nHA (spherical and rod shaped) significantly enhanced the osteogenic gene expression, but a more pronounced effect was seen in the HOBs on PCL-rod nHA films, as the HOBs on PCL-spherical nHA films expressed significantly higher (p<0.05) Runx2 (Fig. 3A) and bone sialoprotein (Fig. 3C) mRNA levels only after 1 day of culturing but not for 7 days of culturing; whereas bone sialoprotein (Fig. 3C) and osteocalcin (Fig. 3D) gene expression levels of the HOBs on PCL-rod nHA films were also significantly up-regulated (p<0.05) after 7 days of culturing. In addition, the HOBs on PCL-rod nHA films expressed significantly higher (p<0.05) levels of Runx2 (Fig. 3A), collagen I (Fig. 3B), and bone sialoprotein (Fig. 3C) gene expression after 1 day and/or 7 days of culturing, compared with those on PCL-spherical nHA films.

Effects of nHA shapes on HOB differentiation. Comparing with the PCL film control, the nHA incorporation significantly enhanced the osteoblastic gene expression of
Effects of HOBs on different substrates on osteogenic differentiation of ASCs in the coculture system
When ASCs were cultured alone, they expressed the early osteogenic gene expression, such as Runx2 and alkaline phosphatase, but for late osteogenic gene marker, there was a very low level of bone sialoprotein gene expression and no osteocalcin gene expression detected in the ASCs after 4 and 14 days of culturing (Fig. 4A–D). However, when ASCs were cocultured with HOBs, osteogenic gene expression of ASCs was differently induced and the extent of induction was largely dependent on the substrates where HOBs were cultured. The bone sialoprotein (Fig. 4C) and osteocalcin (Fig. 4D) gene expression of ASCs was induced after 4 days of coculturing with the HOBs on PCL or on PCL-spherical nHA films, but they sharply decreased after 14 days. In contrast, when ASCs were cocultured with the HOBs on PCL-rod nHA films, the gene expression of bone sialoprotein and osteocalcin was similarly induced at day 4 but substantially increased after 14 days of coculturing, and also Runx2 gene expression was significantly higher than other groups tested (Fig. 4A, C, D).
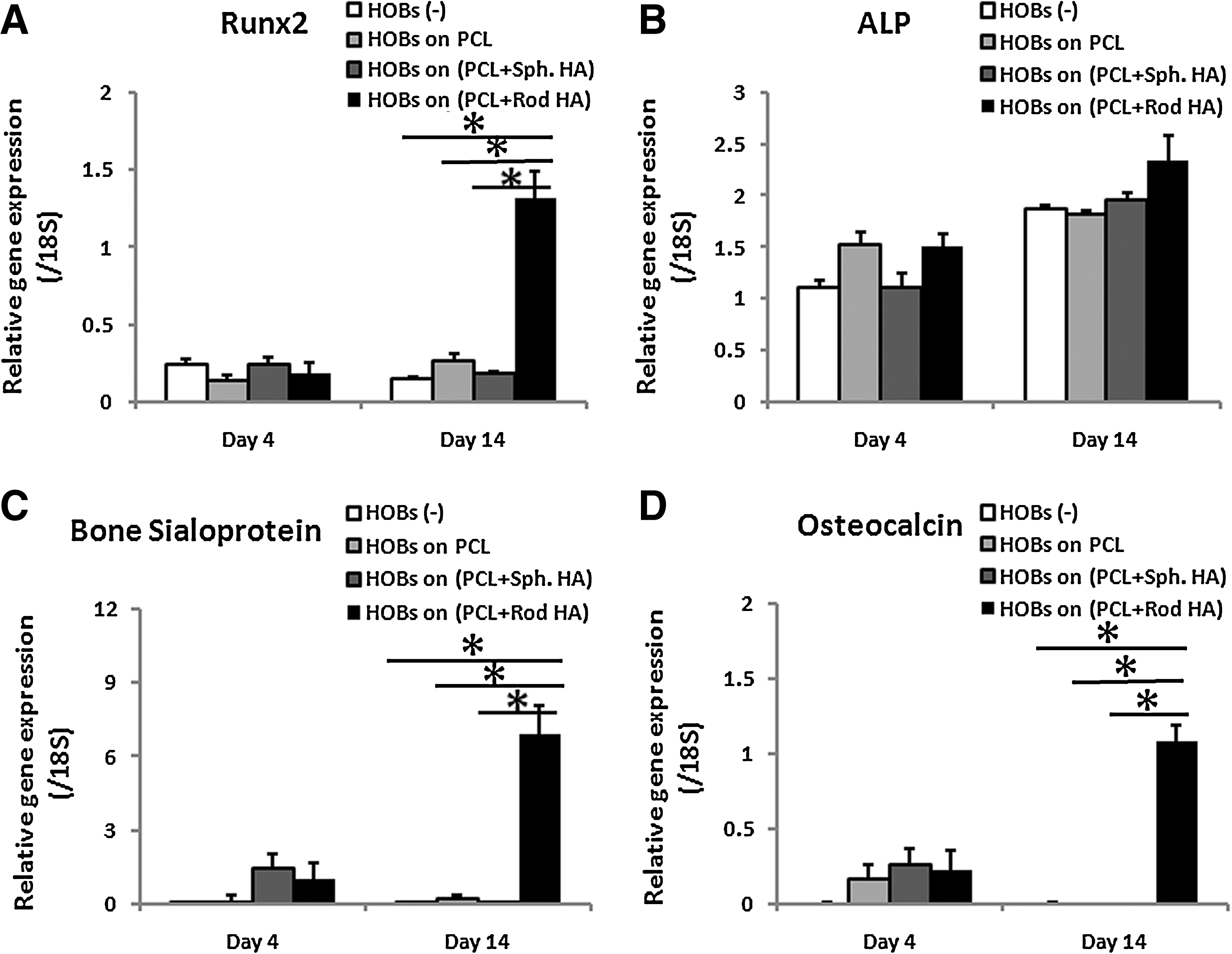
Effects of HOBs interacting with different shapes of nHA on osteogenic differentiation of ASCs in a coculture system. Comparing with the HOBs on PCL or PCL-spherical nHA films, the HOBs on PCL-rod nHA film induced most efficient osteogenic gene expression in ASCs, especially after 14 days of coculturing.
BMP2/Smad1/5 signaling pathway is involved in osteogenic differentiation of ASCs
To gain an insight into how the HOBs interacting with PCL-rod nHA are able to direct ASCs into osteogenic differentiation, we first determined the levels of BMP2 gene and protein expression in the HOBs cultured on different films after 1 and 7 days of culturing. The results showed that BMP2 gene expression (Fig. 5A) was significantly higher (p<0.05) at day 1 in the HOBs cultured on PCL-rod nHA films, but decreased and there were no significant differences at day 7 compared with the BMP2 mRNA levels of HOBs on PCL and PCL-spherical nHA films. However, BMP2 protein levels remained higher (about 1.5-fold) in the HOBs cultured on PCL-rod nHA films (Fig. 5B) at 1 and 7 days. We also determined the BMP2 related Smad1/5 signaling pathway in ASCs when they were cultured alone or with HOBs on different films in the coculture system. Our results demonstrated that the ASCs had a very low level of phosphorylated Smad1/5 protein when cultured alone for 1 and 4 days (Fig. 5C). The phosphorylated Smad1/5 protein expression, however, was dramatically increased by coculturing with HOBs after 4 days of culturing, with the largest increase seen in the ASCs cocultured with the HOBs on PCL rod nHA (Fig. 5C).
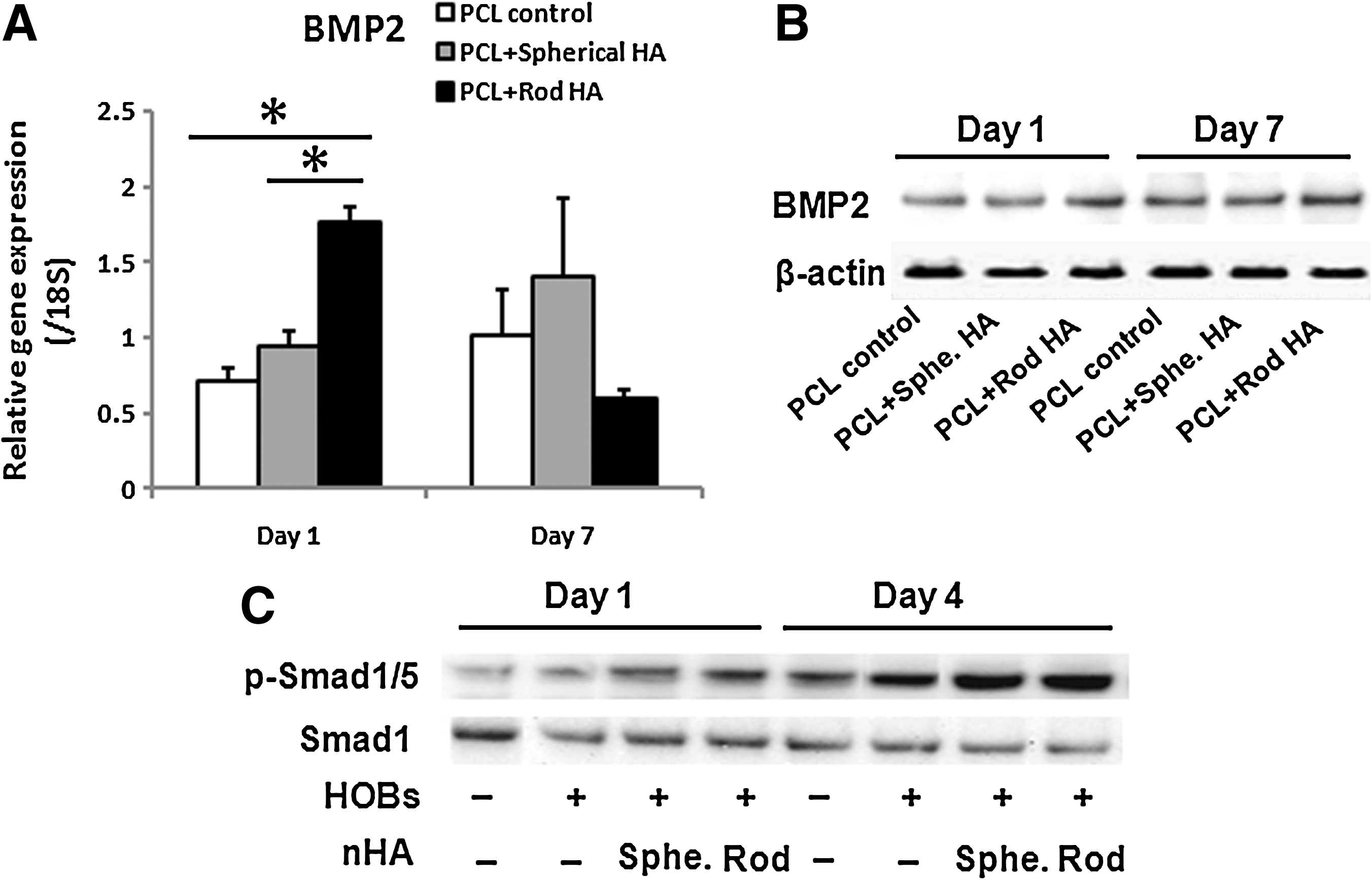
Induction of BMP2 expression in HOBs and activation of Smad1/5 signaling pathway in ASCs in the coculture system.
The results just stated imply that the paracrine BMP2 released from HOBs is involved in directing ASC differentiation into osteogenic lineage through Smad1/5 signaling pathway. To validate this assumption, the coculture medium was supplemented with BMP2 inhibitor (Noggin) and the gene expression of Runx2, bone sialoprotein, osteocalcin, alkaline phosphatase, as well as the Smad1/5 signaling pathway were examined in the ASCs cocultured with the HOBs on PCL-rod nHA films. We found that Noggin treatment significantly (p<0.05) suppressed the inductions of Runx2 (Fig. 6A), bone sialoprotein (Fig. 6C), and osteocalcin (Fig. 6D) gene expression in ASCs. Alkaline phosphatase gene expression, on the other hand, was surprisingly enhanced by the treatment of Noggin (Fig. 6B). Western blot analysis of phosphorylated Smad1/5 protein revealed that the treatment of Noggin did not reduce phosphorylated Smad1/5 protein levels in ASCs (Fig. 6E).
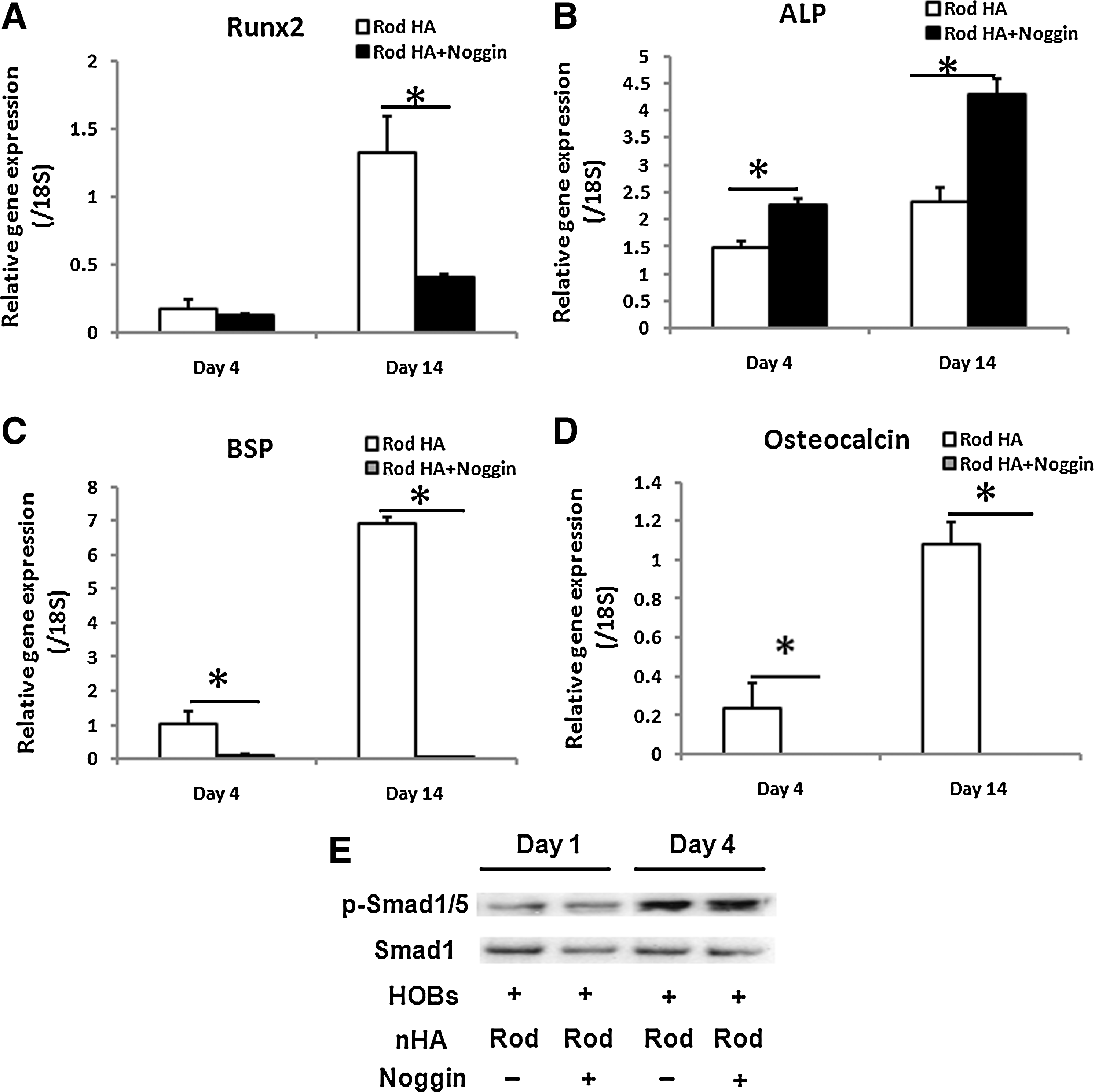
Effects of BMP2 inhibition on osteogenic gene expression and Smad1/5 signaling pathway in the ASCs cocultured with the HOBs on PCL-rod nHA film. The treatment of BMP2 inhibitor (Noggin) in the coculture medium significantly suppressed the induction of Runx2
Discussion
The exact molecular mechanism(s) of how MSCs are directed into different lineages is still lacking, but the mounting in vitro and in vivo evidence suggests that the differentiation fate of MSCs is determined by the local microenvironment niche in an anchorage-dependent manner.30–34 In the current study, we attempted to mimic the in vivo scenario, where the bone implanted biomaterial will be populated by MSCs and osteoblasts. We set up an indirect coculture system with ASCs and HOBs and further manipulated the culture substrate of HOBs in an attempt to reveal how the underlying substrate, in direct contact with HOBs, would indirectly affect the differentiation of ASCs. Our results demonstrated that PCL films incorporated with rod nHA are able to indirectly route ASCs into osteogenic differentiation by directly interacting with HOBs, where the paracrine BMP2 released from HOBs is indispensable for this event.
Generally, adhesion of mammalian cells to biomaterial surface is divided into three steps: protein adsorption, cell–protein interaction, and cell adhesion and/or spreading. The first step, protein adsorption, takes place within seconds of implantation, producing a layer of proteins on the surface of biomaterials, which determines the proceeding steps (cell–protein interaction, cell adhesion and/or spreading); all these three steps are critical for determining cellular functionalities such as cell proliferation and differentiation.35–37 The quantity, conformation, and orientation of the proteins adsorbed on the surface largely depend on both chemical and physical properties of the biomaterial surface. Therefore, many efforts have been directed toward modifying biomaterial's surface in an attempt to optimize the protein adsorption, subsequently modulating the functions of cells.38–41
In this study HOBs showed different osteoblast differentiation efficiency while they were cultured on the substrates with the incorporations of different shapes of nHA, as suggested by the incorporation of rod-shaped nHA significantly increased Runx2, collagen I, and bone sialoprotein gene expression of HOBs after 1 and/or 7 days of culturing when compared with the PCL or PCL-spherical nHA films. These results are in line with our previous study where we showed that the three-dimensional scaffolds coated with PCL layer incorporated with different shapes of nHA enhanced HOBs differentiation, particularly for needle- and/or rod-shaped nHA. 4 To reveal the underlying mechanism(s) contributing to the superior osteoconductivity of PCL-rod nHA, we compared the degradability of the films by measuring the concentrations of Ca2+ and p3− ions in the culture medium soaked with the various films tested. The results showed that there were lower concentrations of Ca2+ and p3− ions in the medium soaked with PCL-spherical nHA or PCL-rod nHA films than that soaked with PCL films after 14 days of soaking; indicating that the incorporation of nHA may facilitate the formation of calcium phosphate deposition. Notably, there were no differences in Ca2+ and p3− ions concentrations for the medium soaked with PCL-spherical nHA or PCL-rod nHA films, suggesting that the degradation of the films or the Ca2+ and p3− ions microenvironment are unlikely to be the contributing factors to the superior osteoconductivity of PCL-rod nHA films.
It is well known that the geometrical cues from the substrates can significantly influence or even dictate the behavior of the cells growing on them.7,42 The roughness of the different films was measured using AFM, and results showed no significant differences between PCL films and PCL films with the incorporation of spherical or rod-shaped nHA, suggesting that the roughness property of rod nHA PCL films is not responsible for their superior osteoconductivity. Binulal et al. showed that the nanofibers elicit lower protein surface coverage, albeit more effective in terms of hMSCs attachment, spreading and osteogenic differentiation. 43 Similarly, a guided osteogenic differentiation of MSCs can be manipulated by selective sizing of the nanotube dimensions. 7 Therefore, it is plausible to speculate that the rod nHA elicit a panel of optimal proteins adsorption, conformation, and orientation, thereby facilitating HOBs cell attachment, spreading, and differentiation; a possible mechanism that ultimately contributes to the superior osteoconductivity of the PCL-rod nHA. However, further studies are under way to investigate this hypothesis.
The interactions between cells and substrates not only determine the phenotypic characteristics of the cells but also modulate the surrounding microenvironment through autocrine and/or paracrine routes.25,26,28 In this study, the HOBs seeded on PCL films with the incorporation of different shapes of nHA were cocultured with ASCs in an attempt to reveal how the microenvironments stemmed from the interactions between HOBs and different substrates affect the differentiation of ASCs. Our results demonstrated that only the HOBs interacting with PCL-rod nHA are able to direct ASCs into osteogenic differentiation, as indicated by the significant up-regulation of Runx2, bone sialoprotein, and osteocalcin gene expression. These results are in agreement with the recent publication demonstrating a significant up-regulation of osteoblastic phenotype in MSCs, only when the MSCs are cocultured with osteoblasts growing on the surface of acid etched titanium and modified acid etched titanium, but not on tissue culture plastic. 26 It is very interesting to note that there was a decreased gene expression for Runx2, bone sialoprotein, and osteocalcin in the ASCs cocultured with the HOBs on PCL or PCL-spherical nHA films. Others reported that human nucleus pulposus cells in micromass, but not in monolayer condition, maintain their right chondrogenic phenotype and direct human MSCs into chondrogenic differentiation. 44 It is, therefore, vital to employ the “right” culture condition, such as appropriate substrate for studying the functionality of primary tissue cells. In the present study, PCL films with rod-shaped nHA incorporation, mimicking the natural bone structure, may best enable HOBs to exert their biological functionalities and provide the “right” and sustainable signals for ASC osteogenic differentiation, whereas the HOBs on PCL films or PCL with spherical nHA incorporation lack the right phenotype and the sustainable capability for steeling osteogenic differentiation of ASCs. Taken together, it is critical to fabricate the biomaterials for bone tissue engineering that are not only able to promote the osteoblastic differentiation of osteoblasts but also able to co-ordinate with them to render an optimal osteogenic niche for the differentiation of stem cells.
BMPs, members of TGF-β family, play a critical role in the process of bone formation and remodeling. 45 The action of BMPs is mediated by heterotetrameric serine/threonine kinase receptor and the downstream Smad1/5 signaling pathway. Once the signaling pathway is activated, a series of osteogenic-related genes are stimulated. 46 Our group previously showed that beta-tricalcium phosphate, a clinically used bone graft substitute, exerts its osteoconductivity through a BMP2 autocrine loop. 28 To reveal how the rod nHA in direct contact with HOBs can indirectly promote the osteogenic differentiation of ASCs, BMP2 gene and protein expression was analyzed in the HOBs cultured on PCL films with or without the incorporation of rod or spherical nHA. Our results showed that the BMP2 gene and protein expression of HOBs on PCL-rod nHA was significantly higher than that of the HOBs on PCL film or PCL-spherical nHA films at day 1, and that the supplementation of Noggin in the coculture medium largely diminished the osteogenic induction of ASCs cocultured with HOBs on PCL-rod nHA films. These results point out that the BMP2 secreted from HOBs plays a key role and is indispensable for the osteogenic differentiation of ASCs cocultured with the HOBs on PCL-rod nHA films. It is worth noting that BMP2 gene expression of HOBs cultured on PCL or PCL-spherical nHA increased from day 1 to day 7, but their levels decreased when HOBs were cultured on PCL-rod nHA films, and the significant difference of BMP2 gene expression between the HOBs on three different films at day 1 disappeared at day 7. This phenomenon seemed to undermine our conclusion just stated that the BMP2 secreted from HOBs plays a key role in the osteogenic differentiation of ASCs cocultured with the HOBs on PCL-rod nHA. Based on our data of BMP2 protein analysis that BMP2 protein expression in the HOBs cultured on PCL-rod nHA films was higher at both 1 and 7 days than those on PCL or PCL-spherical nHA films, we believe that that there is a feedback loop in the HOBs on PCL or PCL-spherical nHA films, attempting to rescue their osteoblastic phenotype by up-regulating BMP2 gene expression. Although this feedback failed to promote osteoblastic gene expression and BMP2 protein expression of the HOBs on PCL or PCL-spherical nHA films to the similar levels as those of the HOBs on PCL-rod nHA films, this feedback further highlights the crucial role of the “right” substrate for maintaining the osteoblastic phenotype and functionalities of HOBs, as presented by PCL-rod nHA films.
Moreover, we investigated how the downstream of BMP2, Smad1/5 signaling pathway is induced in ASCs by the HOBs on PCL-rod nHA films. Our results demonstrated that the phosphorylated Smad1/5 protein expression was, in general, dramatically increased by coculturing ASCs with HOBs and to the largest extent when cocultured with the HOBs on PCL-rod nHA films. However, the treatment of Noggin in the coculture medium did not significantly decrease the phosphorylated Smad1/5 protein levels, but largely abolished osteogenic differentiation of ASCs. These results indicate that osteogenic differentiation of ASCs induced by BMP2 is independent of the Smad1/5 signaling pathway; and that the Smad1/5 signaling pathway can be stimulated by other factors besides BMPs. Similar results were also reported by others who stated that induced osteogenic differentiation by BMP2 can be independent of the Smad1/5 signaling pathway or the independence of BMP2 in the activation of Smad1 signaling pathway.47,48 Considering the complex crosstalk between Smad1 and Ras/MEK signaling pathways 49 ; a pool of soluble factors may be released from the HOBs in the coculture system. We, therefore, hypothesize that BMP2 is able to induce osteogenic differentiation through other pathways, and Ras/MEK signaling pathway and other growth factors released from HOBs may compensate the signals from BMPs to sustain the Smad1/5 signaling pathway after the treatment of Noggin.
In summary, we demonstrated in this study that HOBs displayed efficient osteoblast differentiation and successfully induced osteogenic differentiation of ASCs in an indirect coculture system when they were grown on PCL-rod nHA films but not on PCL or PCL-spherical nHA films, and that the BMP2 released from the HOBs plays a key role and is indispensable for the osteogenic differentiation of ASCs. It is concluded that the incorporation of rod-shaped nHA might provide an excellent tool for fabricating the biomaterials for bone tissue engineering by not only preserving the phenotype of osteoblasts but also coordinating with them to render an optimal osteogenic niche for the differentiation of stem cells.
Footnotes
Acknowledgments
The authors would like to acknowledge the Australian National Health and Medical Research Council and the Australian Research Council for funding this research. We acknowledge the Australian Center for Microscope and Microanalysis Research Facility for providing the facilities. We are grateful for the assistance provided by Mrs Barbara James in culturing the HOBs and in proof reading this manuscript.
Disclosure Statement
No competing financial interests exist.