
Abstract
Tissue engineering approaches are currently being investigated for the restoration of myocardial function in heart failure patients, most commonly by combining cells with a substrate to form myocardial-like constructs (MCs). The final properties of these constructs are dependant on the characteristics of both the substrate and the cells used for fabrication. To create a construct with the appropriate mechanical properties required for any future therapeutic, we tailored an acrylated polypropylene glycol triol (aPPGT) substrate to the elastic modulus of heart tissue and then investigated the fabrication of MCs. We first assessed the aPPGT substrate alone in vivo, both under normal conditions and in an infarct model in mice, and found that there was a mild foreign body response with good integration of the substrate into the epicardial surface in mice hearts. We next studied the fabrication and properties of MCs by culturing mouse embryonic cardiomyocytes on the aPPGT substrate. To achieve myocardial-like concentrically contractile constructs, cocultures with supportive stromal cells were found to be essential and both mouse heart-derived stromal cells or bone-derived mouse mesenchymal stromal progenitor cells (mMSCs) could be used. These different stromal cell types produced MCs with different properties. The average beating rate of the constructs formed from mouse heart-derived stromal cells was significantly higher those constructs formed using mMSCs. Conversely, the constructs formed using mMSCs had reduced fibrotic extracellular matrix secretion and increased hepatocyte growth factor expression. Both of these mMSC construct properties may enhance integration and therapeutic efficacy of the construct postimplantation on the surface of the infarcted heart. This study thus demonstrates the formation of MCs using mechanically tailored aPPGT substrate and also demonstrates the effects of different stromal cell populations have on the properties of the resultant MCs, both of which are critical for future applications of tissue engineering in heart failure patients.
Introduction
This apparent requirement for a supportive stromal cell fraction is consistent with the constitution of native heart tissue, where one or even more stromal cells (e.g., fibroblasts) are in direct contact with each cardiomyocyte and comprise the majority of cells in the heart. 28 Resident stromal cells support the heart tissue through a variety of essential functions, including (1) provision of an extracellular matrix (ECM) structure throughout the heart that integrates and distributes mechanical force; (2) sensing and responding to mechanical stimuli; (3) secretion of autocrine and paracrine factors; and (4) modulation of electrical properties (reviewed in Ref.28,29). However, in a diseased state, it is these same cells that may contribute to the progression of heart failure though excessive fibrosis and remodeling events. 30 For these reasons the choice of support cell component for MC fabrication is likely to be just as important as the cardiomyocytes themselves. While fabrication of synthetic materials capable of mimicking key cell functions may be an option, the complexity of creating a dynamic myocardial ECM mimic is currently beyond our capability. The identification of the appropriate supportive stromal cell type is thus an important step toward the delivery of MC therapeutics for patients with heart disease. These supportive stromal cells must be able to sustain functional cardiomyocytes, allow MC integration in vivo, and obviate any adverse events (e.g., excessive fibrosis).
In this study, we have used an acrylated polypropylene glycol triol (aPPGT) substrate (described previously in Ref. 31 ), tailored to the elastic modulus of native myocardial tissue, to fabricate MCs. The use of modulus-matched aPPGT is vital to facilitate integration in vivo and also modulate the contractile phenotype of cardiomyocytes without the provision of external devices in vitro.27,32 The aPPGT substrate was then combined with a cellular component to fabricate MCs, and given the limited accessibility to human cardiomyocytes and supportive heart stromal cells, mouse cells were used as a model. For the formation of the MCs we used a coculture of embryonic mouse heart-derived cardiomyocytes and mouse heart-derived stromal cells (heartMCs). For comparison, we also used a coculture of embryonic mouse heart-derived cardiomyocytes and the more accessible mouse bone-derived multipotent mesenchymal stromal cells (mscMCs). We thereafter compared the formation and properties of the resultant MCs fabricated using the different stromal cell types.
Materials and Methods
All chemicals and reagents were sourced from Sigma unless otherwise noted.
aPPGT fabrication
aPPGT was synthesized, processed, and characterized as per a previous investigation. 31 Briefly, for acrylation 10% (w/v) PPGT in dehydrated dichloromethane (Univar) was combined with acryloyl chloride (Alfa Aesar) and triethylamine, added simultaneously in an equimolar ratio to give the required molar ratio of PPGT to acryloyl chloride. The reaction took place in an inert argon environment for 24 h, before removing the dichloromethane via distillation. The remaining mixture of aPPGT and triethylamine hydrochloride salt was then dissolved in ethyl acetate (Labscan), cooled to −80°C, vacuum filtered, and the ethyl acetate removed via distillation. The aPPGT was then sterile filtered with a 45 μm polytetrafluoroethylene membrane filter (PM Separations). High acrylate formulations were synthesized using a 1.00 acryloyl chloride to hydroxyl group on PPGT reaction mixture and the low acylate formulations were synthesized using a 0.25 acryloyl chloride to hydroxyl group on PPGT mixture. The aPPGT used for substrates in this study was formulated by combining a high acrylate and low acrylate PPGT in a ratio of 0.025:0.975.
aPPGT tensile testing
aPPGT was fabricated into sheets ∼0.7 mm thick and then cut using a dog-bone-shaped mold (conforming to ISO–37, 0.7×2.0×35 mm) before tensile testing. The samples were tested using a uniaxial tensile test bench (Instron 5567) at a strain rate of 0.06/s. The elastic modulus was determined using a linear regression of the stress–strain curve. The ultimate tensile stress was determined by stress at failure and the elongation to break determined as the strain at failure.
Electrospun fibers
Polycaprolactone (PCL) (Mn 80,000 g/mol) was dissolved in a mixture of chloroform/dimethylformamide (90/10 v/v) at a concentration of 15% (w/v). The polymer solution was ejected through a 21-gauge blunt needle at a flow rate of 2 mL/h. The voltage was set at 10 kV and the deposition distance between the needle and the collector (10.5×10.5 cm2) was 20 cm. The process was run for 1 h deposition, forming an electrospun mat ∼0.7 mm thick.
Substrate fabrication
Strips (∼2 mm wide) of electrospun fibers made from PCL were placed on glass cover-slips and embedded in a solution of aPPGT. The aPPGT was then cross-linked by application of UV, followed by cutting the substrates to size. The sol was then removed and the substrate surfaces oxidized with 0.01 M potassium permanganate for 30 s before a Gelatine solution (Invitrogen), containing 500 ng/mL 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide hydrochloride in the presence 500 ng/mL of N-hydroxysuccinimide, was added (∼3 h) to covalently cross-link gelatine to the surface of the substrates.
Acute myocardial infarction surgery and epicardial implantation in mice
Mice were anesthetized with an intraperitoneal injection of xylazine (10 mg/kg) and ketamine (80 mg/kg), mechanically ventilated, and the chest cavity opened aseptically with a lateral thoracotomy in the third intercostal space. The surface of the heart was exposed by parting the pericardium. In mice subjected to acute myocardial infarction (AMI), the left coronary artery was ligated 1 mm distal to the margin of the left atrium with 7-0 prolene suture and ischemia was confirmed by observing a color change in the myocardium. Substrates were applied to the epicardial surface of the heart and secured with sutures through the electrospun fiber reinforced ends. The chest was then evacuated and closed and the animal allowed to recover. After 5 days (AMI) or 14 days (healthy), the mice were euthanized using CO2 and cervical dislocation, and the hearts removed for analysis. Hearts were embedded in OCT gel before snap freezing in liquid nitrogen, and stored at −80°C.
Heart cell extraction
Heart cells were extracted from C57BL/6 mouse embryos (∼E18). Mice were euthanized by cervical dislocation under approval of the local Animal Ethics Committee. The hearts of the embryos were surgically removed and sliced into 4 pieces and then treated with 200 U/mL collagenase I in PBS (Invitrogen) for 30 min at 37°C followed by a wash in TrypLE (Invitrogen). The TrypLE was replaced every 15 min until no solid tissue could be seen (five to six times) and the supernatant collected. The supernatant was re-suspended in fetal bovine serum (FBS) and centrifuged (400 g, 5 min, 20°C). The cells were then re-suspended in cardiomyocyte media: 10% FBS, 1:4 Media 199:Dulbecco's modified Eagle's medium, and 100 U/mL penicillin and 100 μg/mL streptomycin (all Invitrogen). The cell suspension was filtered through a 40 μm cell strainer (BD Bioscience) and the filtered solution seeded into tissue culture flasks for 45 min to allow the stromal fraction to attach. The remaining enriched cardiomyocyte suspension was removed and designated the cardiomyocyte fraction. The adherent cells were then passaged using TrypLE and designated the stromal fraction.
Mouse mesenchymal stromal progenitor cell extraction
C57BL/6 mice were euthanized using CO2 followed by cervical dislocation under approval of the local Animal Ethics Committee. The femurs of the mice were harvested, scraped clean of any muscle or connective tissue, and crushed using a mortar and pestle. The bone fragments were homogenized, filtered through a 40 μm filter (BD Biosciences), and then centrifuged on a 1.073 g/mL Percoll (Amersham) density gradient (500 g, 20 min, 20°C) and the interface collected. The interface was re-suspended in α-minimum essential medium with 20% FBS, 100 U/mL penicillin and 100 μg/mL streptomycin (all Invitrogen), transferred to tissue culture flasks, and incubated at 37°C in a humidified 5% CO2 environment. After 24 h cell culture, nonadherent cells were removed and the remaining cells cultured until 90% confluent (media changed every 3 days). Cells at passage 1 were then fluorescence activated cell sorted on a FACSAria (BD Biosciences) for a Sca-1+/CD45− cell fraction. This ensured that a stromal population, free from contaminating leukocytes, was obtained. Mouse mesenchymal stromal progenitor cells (mMSCs) were expanded until sufficient numbers were achieved. All mMSCs in this study were characterized and used at passages 6–12.
mMSC characterization
To characterize mMSC phenotype, the cells were stained with directly conjugated antibodies against: CD45 (APC), CD90 (FITC), and Sca-1 (PE) (All 5 μg/mL; BD Pharmingen). The samples were run on an LSRII flow cytometer (BD Biosciences) with at least 1×104 counts taken per sample. Trilineage differentiation potential of the mMSCs was verified before use as per previously described methods using human MSCs. 33
Immunofluorescence staining
Samples were fixed in 4% paraformaldehyde for 15 min at 20°C. The samples were stained with primary antibodies (or appropriate IgG controls) against fibronectin (5 μg/mL; Sigma), collagen I (5 μg/mL; Abcam), collagen IV (5 μg/mL; Abcam), laminin (5 μg/mL; Abcam), myosin heavy chain (5 μg/mL; Abcam), cardiac troponin I (19 μg/mL; Abcam), smooth muscle alpha actin (2.5 μg/mL; Sigma), connexin 43 (5 μg/mL; BD Bioscience), CD45 (2.5 μg/mL; BD Pharmingen), CD11b (2.5 μg/mL; BD Pharmingen), or CD90 (2.5 μg/mL; BD Pharmingen). For observation the cells were incubated with goat anti-mouse IgG (fibronectin, myosin heavy chain, cardiac troponin I, and connexin 43), anti-rabbit IgG (collagen I, collagen IV), goat anti-chicken IgG (laminin), or goat anti-rat IgG (CD45, CD11b, and CD90), AlexaFluor 488–, or AlexaFluor 568–conjugated anitbodies (All, 10 μg/mL; Invitrogen). Nuclei were stained using Hoechst33342 (1 μg/mL; Invitrogen) or propidium iodine (1 μg/mL; Invitrogen).
MC seeding
Gelatine-coated aPPGT substrates were seeded with cardiomyocyte fraction cells combined with either the stromal fraction cells or mMSCs at a 3:1 ratio. The substrates were seeded in cardiomyocyte media in ultra low attachment plates (Corning) to ensure that cells only attached to the functionalized aPPGT substrates. A schematic of the MC design is shown in Figure 1.

Acrylated polypropylene glycol triol (aPPGT) substrate-based myocardial-like construct (MC) design. Color images available online at www.liebertonline.com/tea
Beating rates
The beating rates of the MCs were determined in cardiomyocyte media using real-time microscopy on an Olympus IX81 microscope followed by image analysis of the acquired image stack in ImageJ, Version 1.42q. For isoprenaline stimulation experiments, the MCs were incubated in 1 μM isoprenaline in cardiomyocyte media.
Quantitative polymerase chain reaction
Ribonucleic acid (RNA) was extracted using RNeasy Kits (Qiagen). Complementary deoxyribonucleic acid (cDNA) was transcribed using Superscipt III reverse transcriptase (Invitrogen). Platinum SYBR Green qPCR SuperMix-UDG (Invitrogen) was used as the Master-mix and quantitative polymerase chain reaction (qPCR) was run on a 7500 Fast Real-Time PCR System (Applied Biosystems). Results were analyzed using the 2−ΔΔCt method relating gene expression to glyceraldehyde 3-phosphate dehydrogenase (Gapdh) and then to E18 heart expression. Primers were validated using cDNA dilution; sequences are given in the Supplementary Table S1 (Supplementary Data are available online at www.liebertonline.com/tea).
Statistical analysis
Data are presented as mean±standard error of the mean (SEM). To determine statistically significant differences two-tailed Student's t-tests were used with p<0.05 deemed as significant.
Results
aPPGT elastic modulus was tuned to myocardium
The synthesis and characteristic properties of aPPGT substrates have been described previously. 31 As shown in Table 1, the tensile modulus of the aPPGT substrate utilized in this study was tuned to 61 kPa (n=3) by using high acrylate and low acrylate PPGT in a ratio of 0.025:0.975, 31 which is similar to the circumferential modulus of rat ventricular tissue (38–60 kPa 34 ). Other mechanical properties that may affect material performance, including the ultimate tensile strength and elongation to break, were also determined, with recorded values of 66 kPa and 110%, respectively (n=3).
As shown in Figure 1, substrates were created by embedding electrospun PCL fibers at the peripheral ends (of each strip) of the aPPGT substrate to provide rigidity for handling, maintenance of substrate geometry, increase the fracture toughness, and allow effective suturing (for in vivo experiments).
In vivo implantation of aPPGT substrates demonstrated integration
The in vivo response to the aPPGT substrates (cell-free) was analyzed following epicardial suturing to mouse hearts for either 14 days in a healthy mouse (Fig. 2) or 5 days in an AMI model (Supplementary Fig. S1). In the healthy day 14 mouse, CD45+/CD11b+ cells (monocytes/macrophage subset of leukocytes) and CD90+ cells (indicative of proliferating fibroblasts35,36) were detected in a layer (∼300 μm) surrounding the substrate, along with deposition of collagen I around this layer (∼300 μm) (Fig. 2C, D). Further, this cell layer was present both on the inner (Fig. 2C) and outer (Fig. 2D) side of the substrate, indicating integration into the heart tissue. It should be noted that cells positive for CD45, CD11b, and CD90 were only found adjacent to the implanted substrates (Fig. 2C, D) and in infarcted myocardium (Supplementary Fig. S1B, C), suggesting that cells expressing these markers are only present after AMI or a foreign body response.

Histochemical and immunofluorescence staining of sections of healthy left ventricle wall 14 days after implantation.
Characterization of cells used for MC fabrication
Cardiomyocytes derived from E18 mice hearts were combined in coculture with either mouse heart-derived stromal fraction cells (heartMCs) or mMSCs (mscMCs) to form contractile MCs. The heart-derived cardiomyocyte fraction and stromal fraction used for MC fabrication was characterized using staining for myosin heavy chain (MYH, stains cardiomyocytes) and smooth muscle alpha actin (SM α-actin, stains a variety of stromal cell types and embryonic cardiomyocytes). Staining demonstrated that the cardiomyocyte fraction contained 80% (±4.8% SEM) cardiomyocytes, whereas the stromal fraction contained 6% (±1.0% SEM) cardiomyocytes (Fig. 3A). Striated cardiomyocytes were only found in the cardiomyocyte population, indicating that any cardiomyocytes present in the stromal fraction were immature (Fig. 3B). In the cardiomyocyte fraction, 75% (±3.9% SEM) of the cells displayed SM α-actin staining compared to 53% (±8.3% SEM) in the stromal fraction (Fig. 3A). The SM α-actin cells present in the cardiomyocyte fraction are likely to be embryonic cardiomyocytes, whereas in the stromal fraction they are likely to be a mixture of stromal cell types, including smooth muscle cells, resident heart MSC-like populations, myofibroblasts, and some embryonic cardiomyocytes.
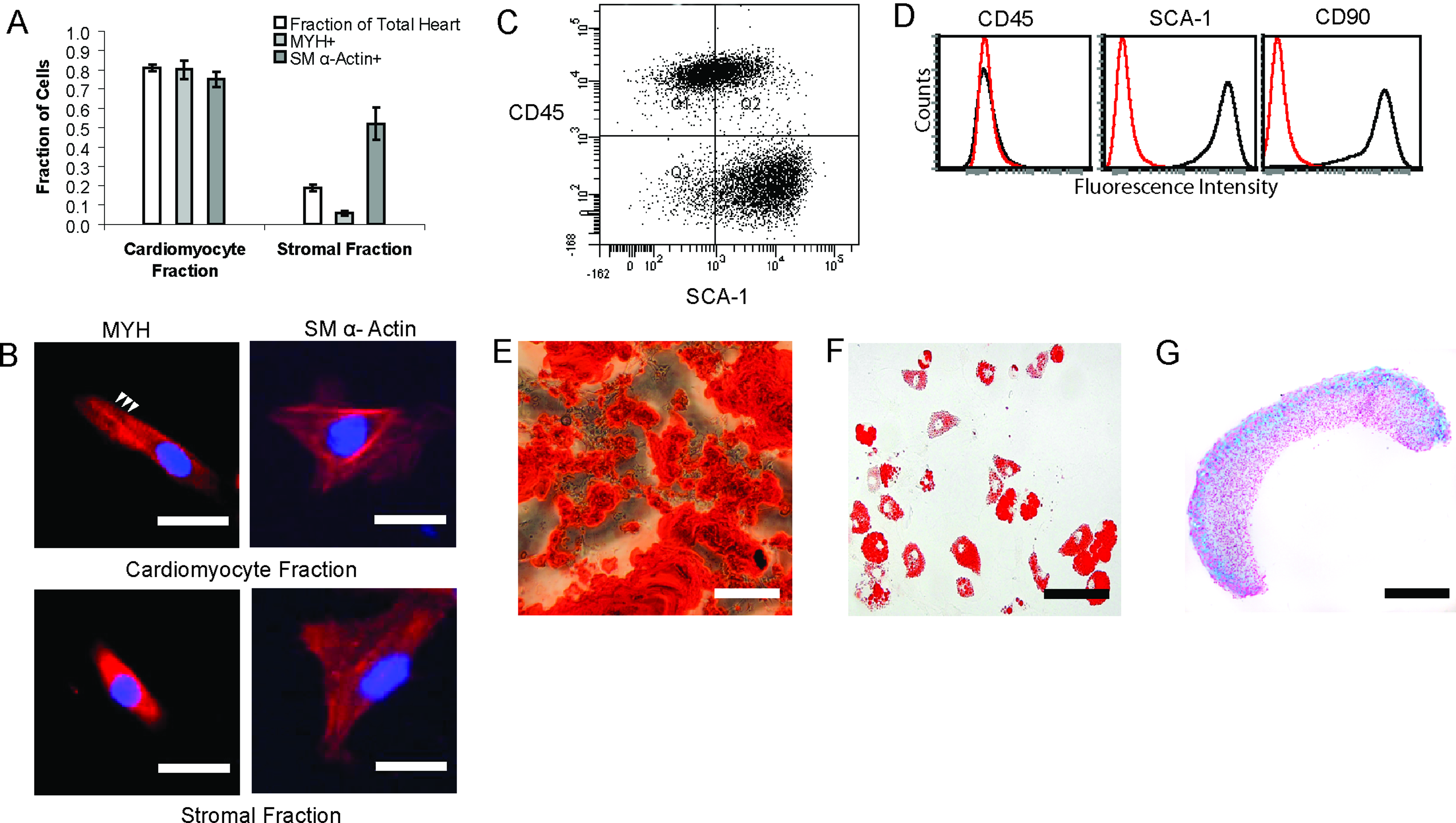
Characterization of cell fractions used for MC fabrication.
The mMSCs used for mscMC fabrication were sorted for Sca-1+/CD45− at passage one (Fig. 3C) and then expanded. After expansion the mMSC population was characterized by expression of CD45−, CD90+, and Sca-1+ using flow cytometry (Fig. 3D) and by trilineage differentiation potential into osteocytes, adipocytes, and chondrocytes (Fig. 3E–G).
The differences in expression of ECM proteins and connexin 43 between the stromal fraction cells and the mMSCs were compared, as these molecules may play critical roles in support of the cardiomyocytes (Fig. 4). The stromal fraction displayed an ECM secretion profile consistent with a fibrotic response, indicated principally by collagen I deposition (Fig. 4A). In comparison, the mMSCs expressed a less fibrotic ECM profile, as indicated by the deposition of predominately fibronectin and laminin (Fig. 4B). Further, only the mMSCs expressed connexin 43 (Fig. 4B), suggesting that both stromal cell types may electrically interact with the cardiomyocytes in different ways. 4
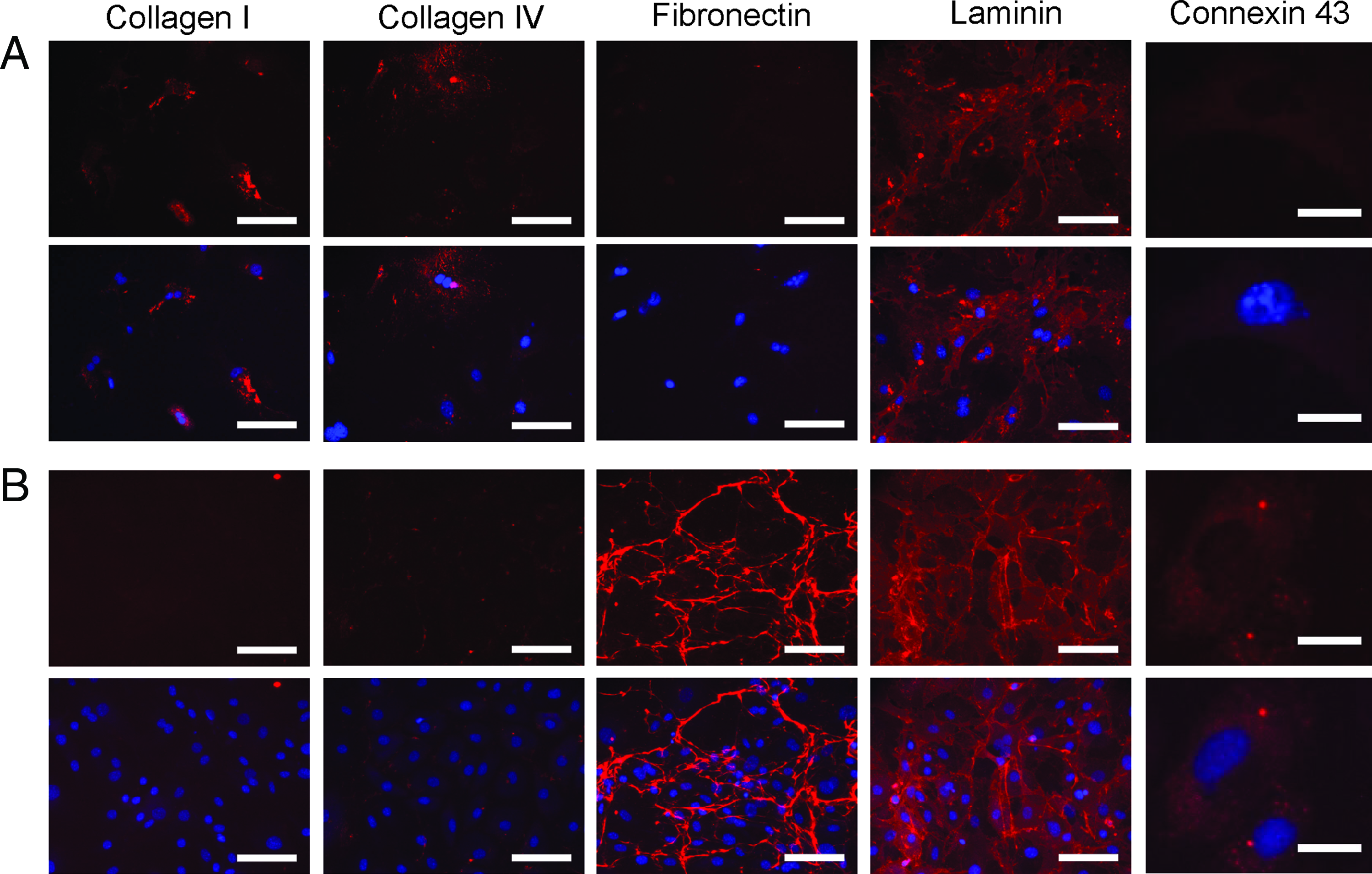
Characterization of extracellular matrix and connexin 43 expression highlights differences between stromal fraction cells and mMSCs.
MC fabrication requires stromal support cells
In the absence of a stromal cell component there was poor attachment and spreading of the cardiomyocytes on the aPPGT substrates, as indicated by cardiac troponin I staining (Fig. 5A). In both the heartMCs (Fig. 5B) and mscMCs (Fig. 5C), spherical aggregates formed in suspension the day after seeding. After 4 days these aggregates attached and spread on the aPPGT substrate and spontaneous beating in both MC types was observed. By day 7, coverage of the substrates was observed on both MC types, indicated by both the phase contrast photomicrographs and the immunofluorescence images of GFP (Fig. 5B, C). Further, both MCs contracted spontaneously (Supplementary Video S1 and S2). It was also noted that in some areas on the mscMCs, cells arranged into fibers (Fig. 5C) and showed spontaneous beating.

Contractile MCs require supportive stromal cells.
Differences in heartMC and mscMC beating rate under normal and isoprenaline stimulated conditions
The heartMCs displayed significantly higher beat rates than mscMCs (Fig. 5D), being 0.7 beat/s (±0.05 SEM) and 0.1 beats/s (±0.03 SEM), respectively. Further, the heartMCs (Supplementary Video S1) beat with more vigour than the mscMCs (Supplementary Video S2). Both MC types had significantly increased beating rates when stimulated with isoprenaline, 2.6 beats/s (±0.44 SEM) and 2.9 beats/s (±0.20 SEM) for the heartMCs and mscMCs, respectively. Further, in the presence of isoprenaline the beating rates for heartMCs and mscMCs were no longer statistically different (p=0.49). Examples of the MC beating rate analysis can be found in Supplementary Figure S2.
Comparison of gene expression in E18 hearts, heartMCs, and mscMCs
Gene expression was analyzed using qPCR for mature cardiomyocyte markers (myosin heavy chain α [Myh6], myosin heavy chain β [Myh7], and connexin 43 [Cx43] [Fig. 6A]), ECM molecules (fibronectin [Fn1], collagen I [Col1a1], collagen III [Col3a1], collagen IV [Col4a1], and laminin γ1 [Lamc1] [Fig. 6B]), and growth factors (fibroblast growth factor 2 [Fgf2], hepatocyte growth factor [Hgf], and insulin-like growth factor 1 [Igf1] [Fig. 6C]). The gene expression was compared between E18 mouse hearts, heartMCs, and mscMCs.

mRNA expression analysis of E18 hearts and MCs using quantitative polymerase chain reaction.
Expression of Myh7/Myh6 ratio was lower in both heartMCs (50-fold) and mscMCs (120-fold) than in E18 hearts (Fig. 6A). There was no significant difference in Cx43 expression between heartMCs and E18 hearts; however, there was a 3-fold higher expression in mscMCs than in heartMCs (Fig. 6A). This may be due to the expression of Cx43 by the mMSCs, which are added as a supportive stromal component in the mscMCs (Fig. 4).
The expression of the different ECM types analyzed, Fn1, Col1a1, Col3a1, Col4a1, and Lamc1, was significantly higher (61-, 2774-, 4-, 3-, and 5-fold, respectively) in heartMCs compared to E18 hearts. The mscMCs also had a significantly higher ECM expression for Fn1, Col1a1, Col4a1, and Lamc1 (44-, 488-, 2-, and 2-fold, respectively) compared to E18 hearts. However, the mscMCs had a significantly lower expression of Col1a1, Col3a1, Col4a1, and Lamc1 compared with heartMCs (5-, 4-, 2-, and 2-fold, respectively).
The expression of Fgf2, Hgf, and Igf1 was significantly higher in both heartMCs (78-, 14-, and 7-fold, respectively) and mscMCs (73-, 38-, and 9-fold, respectively) compared to the E18 hearts. There was a significant difference in the expression of Hgf between the two investigated MC types, with a higher expression (3-fold) in mscMCs compared to the heartMCs.
Discussion
aPPGT substrates 31 were used to provide an appropriate platform for the development of a therapeutic MC. This was done by tailoring aPPGT substrates to the previously published circumferential elastic modulus values of cadaveric rat ventricles (Ecirc) ∼38–60 kPa. 34 This provision of a mechanically matched substrate provides a suitable resistance to the beating movement of the cardiomyocytes, as per previous investigations using mechanically tuneable substrates.27,32 This mechanically matched substrate will also facilitate force transduction of the movement generated by beating heart tissue in vivo, which may facilitate integration. We have also characterized, for the first time, the in vivo response to the aPPGT substrate. The results indicated that the aPPGT substrate was integrated into the epicardial surface of the heart tissue with a low level foreign body response, 37 one that is unlikely to prohibit their in vivo use. However, further studies are required to determine the full extent of the foreign body response over longer periods and whether it will adversely affect its use as a therapeutic component.
We next fabricated MCs using the developed aPPGT substrate in combination with embryonic mouse cardiomyocytes. We found that to form a contractile MC with cardiomyocytes on the aPPGT substrate, a supportive stromal cell component was required. For MC fabrication the heart stromal fraction was used, as this recapitulates the in vivo heart cell composition, and it is consistent with current methodologies where whole heart extracts are used (inclusive of both cardiomyocytes and the stromal fraction cells).20,26 For therapeutic applications, isolation and expansion of human (especially patient derived) heart stromal fractions may be difficult to achieve in the clinic. Further, the potential of cardiac fibroblasts (and also other tissue fibroblasts) to form myofibroblasts (which deposit excessive fibrotic tissue) is clearly un-favorable and could lead to further disease progression. Therefore, we have chosen to also investigate the use of MSCs as a supportive stromal cell component for MC fabrication. The benefits of using MSCs are their immunosuppressive properties, 38 secretion of therapeutic paracrine factors in disease models, 39 demonstrated therapeutic benefit in heart disease models,40,41 and previous/current use in phase II and III clinical trials for treatment of AMI patients (www.clinical trials.gov). In this investigation mouse-derived cells were used, given the difficulties associated with obtaining human heart-derived cardiomyocytes and heart-derived stromal fraction cells.
It was demonstrated that both stromal fractions used in this study could form concentrically contractile MCs. Further comparisons revealed that the mscMCs displayed a significantly reduced beating rate compared to heart MCs in normal media. However, both MC types increase their beating rates in response to stimulation with an adrenoreceptor agonist, isoprenaline. This suggests that mMSCs exert an inhibitory effect on the beating rate, although we do not currently know the exact mechanism of this effect. It is possible that it is due to the compaction stresses mMSCs exert on the cardiomyocytes and/or electrical interference via coupling through connexin 43 gap junctions. 42 The spheres formed during the initial seeding of the MCs (first 1–2 days) demonstrated limited spontaneous beating activity in mMSC-cardiomyocyte coculture, whereas in the stromal fraction-cardiomyocyte culture, smaller but beating spheres were formed. In the final MCs, the reduction of beating caused by the mMSCs may be beneficial (e.g., the beating rate would be reduced until the delivered cardiomyocytes electrically integrate into the host, thereby reducing arrhythmic events) or detrimental (e.g., the beating rate never recovers and results in possible interference with host electrical behavior) to therapeutic applications and will need to be investigated further.
A comparison of the expression of key cardiomyocyte markers, ECM components, and growth factors was performed and E18 hearts, heartMCs, and mscMCs compared. The levels of Myh6 expression in both the heartMCs and the mscMCs were similar, indicating that both MCs had similar proportions of cardiomyocytes remaining after 7 days. In this study, the analysis of the MCs revealed the Myh7/Myh6 ratio was high in E18 hearts and was significantly lower in both the heartMCs and the mscMCs after 7 days of culture. This indicates that both MC types were capable of producing an environment that was conducive to the maturation of the cardiomyocytes toward a predominately Myh6-expressing phenotype. In previous studies, the Myh7/Myh6 ratio of mRNA and protein expression was shown to decrease over 21 days after birth in rats. 43 It has also been demonstrated that hypothyroidism and diabetes may switch the expression toward a predominant Myh7 expression, which may be a maladaptive response and detrimental to cardiac function.44–46 The cardiomyocytes on the MCs in our study therefore display a phenotype consistent with a healthy and maturing cardiomyocytes in relation to Myh6/Myh7 ratio; however, other markers of hypertrophy may require investigation before use in therapeutic applications.
ECM proteins were also investigated as they form a structural framework that allows the attachment and force transduction of the cardiomyocytes, 28 and may also affect cardiomyocyte integration into host myocardium. 47 Indicative of an attempt to create new tissue, expression of all ECM types analyzed was significantly higher in the heartMCs than in E18 hearts. This included strong expression of structural/fibrotic collagens, Col1a1 and Col3a1, which was less pronounced in mscMCs than in heartMCs. This lower expression of Col1a1 and Col3a1 in the mscMCs could be more beneficial for the delivery of regenerative therapies by allowing for faster and more effective tissue integration 47 and reduced deposition of fibrotic scar tissue.
The expression of growth factors, including Fgf2, Hgf, and Igf1, was significantly higher in both MC types than in E18 hearts. Expression of these growth factors has been previously implicated in the regeneration of damaged heart tissue.26,42,48–51 The observed high expression of Fgf2, Hgf, and Igf1 in both MC types may thus be beneficial for therapeutic applications. The only significant difference observed in this study between the MC types was a 3-fold higher expression of Hgf in the mscMCs than in the heartMCs. Expression of hepatocyte growth factor by MSCs has been previously shown to stimulate angiogenesis. 39 The observed higher expression of Hgf in the mscMCs may thus be advantageous for therapeutic applications by stimulating angiogenesis and vascularization of the implanted construct.
Given these promising findings, each MC type requires comparison in the future in a myocardial infarct model to determine therapeutic differences. It is likely that any therapeutic differences between heartMCs and mscMCs will be due to a combination of factors, including beating rate, ECM production, and growth factor production. Therefore, careful consideration is required to ascertain the combinatory impact of these factors when the different MC types are assessed within an in vivo infarct model.
Conclusion
In this study we have successfully fabricated contractile MCs using a heart tissue modulus-matched aPPGT substrate combined with mouse embryonic cardiomyocytes. Overall, our findings demonstrate that a supportive stromal cell component is required in combination with cardiomyocytes to facilitate the formation of contractile MCs. Heart-derived stromal fraction cells or mMSCs were used as this supportive stromal fraction; however, each give rise to different MC properties. Based on the reduced fibrotic ECM expression and increased Hgf expression, we believe that the mscMCs have greater potential, over the heartMCs, for the restoration of myocardial function in heart failure patients. In future experiments, we aim to test and compare both heartMCs and mscMCs in an in vivo mouse infarction model to assess functional incorporation of the delivered cells and changes in heart function over time.
Footnotes
Acknowledgments
We would like to acknowledge Cedryck Vaquette for the fabrication of the electrospun fibers, Jessica Frith for assistance with article preparation, and funding under ARC Discovery Grants Scheme.
Disclosure Statement
No competing financial interests exist
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.