
Abstract
It is well known that physiological functions and pathological conditions of cells and tissues can be influenced not only by chemical molecules, but also by physical stimuli such as electromagnetic waves. In particular, epidemiological studies suggest possible associations between exposure to electromagnetic fields and an increased risk of tumors and neurodegenerative disorders, such as Alzheimer's disease. However, depending on the dose and on the length of treatment, the electromagnetic stimuli can be harmful or induce a cytoprotective cellular response, suggesting a possible application in medical therapy. In this study, under a tissue engineering viewpoint, we investigated the effects of an electromagnetic wave (magnetic field intensity, 2 mT; frequency, 75 Hz) on a neuronal cellular model characterized by the overexpression of the amyloid precursor protein (APP). After a prolonged electromagnetic treatment, lower mitochondrial activity and proliferation rate, resulting in a higher cellular quiescence, were observed. Focusing on the stress and oxidative pathways, we detected an overall increase of two fundamental proteins, the chaperone heat shock protein HSP70 and the free radical scavenger superoxide dismutase-1 enzyme (SOD-1). Interestingly, we found that the electromagnetic stimulation promotes the nonamyloidogenic processing of APP through an increased expression of the α-secretase ADAM10 and an enhanced release of the soluble neurotrophic factor sAPPα (a product of the ADAM10-mediated cleavage of APP). In conclusion, these findings suggest that the electromagnetic stimulus, if properly administered in terms of dose and timing, is able to induce a cytoprotective response in the cell. Moreover, these results suggest a possible use of this particular physical stimulation to improve the functional capability of the cells to face noxae.
Introduction
The mechanisms by which an electromagnetic radiation can interact with the biological systems and cause cell damages, tumors, and neurodegenerative diseases have been only partially investigated. For instance, it is known that the low-energy EMFs are not able to directly damage cellular proteins or the DNA, but, in complex in vivo/in vitro systems, alterations of blood parameters, changes of cytokine profiles, and effects on the immune system have been documented without a clear understanding of the underlying molecular mechanisms.12–14
At systemic level, a possible explanation of these effects may revolve around the suppression of the pineal hormone melatonin involved in free radical scavenging. 15 At molecular level, an electromagnetic wave can affect the redox status of the cells acting, in particular, on the formation of reactive oxygen species (ROS) and other free radicals, which could, in turn, damage lipids, proteins, carbohydrates, and nucleic acids present inside the cells.14,16 Moreover, the response to EMFs could evoke a general stress reaction, 17 in particular via the expression of the heat shock protein HSP70, whose transcriptional and translational levels increase during an electromagnetic stimulation in several cell lines and organisms.18,19
Interestingly, depending on the EMF dose and on the cell type, the biological effects can induce cytotoxicity or cytoprotection.20–22 The cytoprotection might be induced by a kind of hormetic effect, in which low doses of otherwise adverse chemical/physical stimuli improve the functional ability of cells and tissues to face noxae. 23 As consequence, if a generally accepted definition of tissue engineering relies on suitable methods to improve or replace biological functions, the hormetic effect could be studied and proposed under a tissue engineering viewpoint.
To this regard, a protective action of EMF has been obtained via the electromagnetic modulation of Ca2+ influx with consequent reduction of the apoptosis rate.24,25 Moreover, based on the concept that the hormetic effect may implicate the strengthening of the molecular responses to stress, it has been proposed the use of a long-wave EMF as a possible therapy against neurodegenerative disorders, such as AD, as documented by the delay of the cellular senescence associated with an increased expression of heat shock proteins. 26 Another hormetic action leading to biological improvements has been achieved in an in vivo study on a mouse model of AD, where the cognitive functions ameliorated as a consequence of a high-frequency EMF treatment. 27
On the other hand, in an in vitro AD cellular model characterized by cells stably transfected with the human amyloid precursor protein (APP) carrying the Swedish mutation, an EMF exposure caused a significantly increased secretion of the amyloid peptide (Aβ), whose accumulation is a typical feature of AD pathogenesis. 28 Notably, APP is an integral membrane protein that can undergo two distinct metabolic pathways: amyloidogenic and nonamyloidogenic. The first one, via β- and γ-secretases, originates the Aβ peptide involved in the genesis of amyloid plaques, a key hallmark of AD 29 ; the second one produces the neurotrophic fragment sAPPα after the cleavage operated by an α-secretase. Within this context, the protective or nonprotective actions of an electromagnetic treatment may depend on its influence on the particular metabolic processing of APP.
Taking into account all these considerations, our work was aimed to identify, under a tissue engineering point of view, a possible beneficial action of EMF on neuronal cells in aid of pathological conditions. To this purpose, we applied an electromagnetic stimulus (magnetic field intensity, 2 mT; frequency, 75 Hz) to human neuroblastoma cell lines, one of which overexpressing the wild-type (WT) copy of the APP751 gene, and we studied the biological effects of this physical treatment on different parameters. We mainly focused on the evaluation of basic cellular functions (i.e., mitochondrial activity, cell proliferation, and cell cycle phases), as well as of key components of the stress response and the redox system (i.e., HSP70 and SOD-1 proteins), and of the nonamyloidogenic APP pathway (i.e., the α-secretase ADAM10 and the soluble neurotrophic factor sAPPα).
Materials and Methods
Cell cultures and transfections
The SH-SY5Y human neuroblastoma cell lines were grown in Eagle's minimum essential medium supplemented with 10% fetal calf serum, 1% penicillin–streptomycin,
The SH-SY5Y cells, stably transfected with the vector of WT APP isoform of 751 amino acids (APP751) or with the empty vector (pIRES), were prepared as follows: 5×105 SH-SY5Y cells, seeded in a six-well plate, were transfected with 2.5 μg of pIRES or APP751 vectors according to the Lipofectamine™ LTX Reagent's protocol (Invitrogen, Milan, Italy). The antibiotic G418 (Sigma, Milan, Italy) was added at a concentration of 600 μg/mL and drug resistant cells were collected after 1 week. The cells were seeded in 96-well plates with serial dilutions to isolate one cell per well. Wells that contained more than one cell were excluded from further investigation. After 2–3 weeks, surviving clones reached confluence and were then expanded. G418-resistant clones were analyzed by western blotting to confirm the overexpression of APP751. Stably transfected cells were maintained in medium supplemented with G418 at a final concentration of 400 μg/mL.
Construction of the vectors
The Gateway® technology is a universal cloning method (Invitrogen) that takes advantage of the site-specific recombination properties of bacteriophage lambda to rapidly and efficiently insert a gene into multiple vector systems. This technology is composed by an entry vector and a destination vector that can recombine through the att recombination sites. Briefly, following the manufacturer's instructions, the destination empty vector was derived by the insertion of the GWc cassette (Invitrogen) into the pIRES2-EGFP (Clontech, EuroClone, Milan, Italy) obtaining the vector pIRES-GWc-EGFP (Gateway® Destination Vector; Invitrogen). In particular, the GWc cassette, flanked by the attR1 and attR2 recombination sites, contained the chloramphenicol resistance gene (CmR) and the ccdB (procariotic toxic gene), useful to carry on the following steps of cloning and recombination.
To express the gene of the WT APP isoform of 751 amino acids (APP751), the amplified PCR product was cloned into the pCR®8/GW/TOPO® entry vector (Gateway® Entry Vector; Invitrogen) to generate the entry clone. The PCR product was obtained using the PfuUltra High-Fidelity DNA Polymerase (Promega, Milan, Italy) and the following primers: APPfor 5′-ATGCTGCCCGGTTTGGCA-3′ and APPrev 5′-CTAGTTCTGCATCTGCTCAA-3′. The template for the PCR was the cDNA cloned into the vector pcDNA-APP751. 31
Subsequently, the resulting vector, called APP751/GW/TOPO®, was used to transform Mach1™-T1R chemically competent E. coli cells (Invitrogen). The obtained colonies were screened and then sequenced using the provided kit oligonucleotides (Invitrogen). The correct APP751/GW/TOPO® plasmid was recombined in vitro with the destination empty vector pIRES-GWc-EGFP, thus obtaining pIRES-GWc-APP751-EGFP (Invitrogen). The resulting vector and the empty vector were used to transform Mach1™-T1R chemically competent E. coli cells and the obtained colonies were screened by colony PCR.
The plasmids used for the transfections were purified with the HiSpeed® Plasmid Midi Kit (Qiagen, Valencia, CA). The resulting vectors, that is, the empty vector pIRES-GWc-EGFP and the vector pIRES-GWc-APP751-EGFP (called for brevity pIRES and APP751, respectively), were used to transfect the WT SH-SY5Y as previously described.
Electromagnetic bioreactor and cell exposure
An electromagnetic bioreactor was built (Fig. 1).32,33 It consisted of a carrying structure custom-machined in a tube of polymethylmethacrylate. The windowed tube carried two solenoids, the planes of whom were parallel. The cell cultures were 5 cm away from each solenoid plane and the solenoids were powered by a Biostim SPT pulse generator (Igea, Carpi, Italy), a generator of pulsed electromagnetic fields (PEMFs). In this experimental setup, the magnetic field and the induced electric field were perpendicular and parallel to the seeded cells, respectively.

Electromagnetic bioreactor. The windowed tube carried two solenoids, the planes of whom were parallel; in this experimental setup the magnetic field and the induced electric field were perpendicular and parallel to the seeded cells, respectively. The electromagnetic bioreactor applied to the cells a PEMF with the following characteristics: intensity of the magnetic field equal to 2±0.2 mT, amplitude of the induced electric tension equal to 5±1 mV, signal frequency of 75±2 Hz, and pulse duration of about 1.3 ms. PEMF, pulsed electromagnetic field.
Given the position of the solenoids and the characteristics of the pulse generator, the electromagnetic bioreactor applied to the cells a PEMF with the following characteristics: intensity of the magnetic field equal to 2±0.2 mT, amplitude of the induced electric tension equal to 5±1 mV, signal frequency of 75±2 Hz, and pulse duration of about 1.3 ms.32–39 The magnetic field was measured with a Hall Effect transverse gaussmeter probe (HTD61-0608–05-T, F.W. Bell Line; Sypris Solutions, Louisville, KY) and a gaussmeter (DG-500; Laboratorio Elettrofisico, Milan, Italy), the induced electric tension was measured with a standard coil probe, and the temporal pattern of the electromagnetic signal was evaluated by a digital oscilloscope (LT322, WaveRunner Series; Le-Croy, Chestnut Ridge, NY).
The electromagnetic bioreactor was placed into the cell culture incubator and the exposed cells were continuously stimulated by the PEMF for 18, 36, 54, and 72 h (a duration of 72 h is the maximum culture period without the need of medium change). The control cultures were placed into a different incubator in the absence of PEMF stimulation. The culture medium was not changed during the whole culture period, unless in the case of experiments performed to assess the effect of fresh medium where it was changed daily.
MTT assay
Mitochondrial enzymatic activity was estimated by the MTT assay [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] (Sigma). A cell suspension of 7×103 cells in 100 μL was seeded into each well of 96-well plates. After 18, 36, 54, and 72 h, with or without electromagnetic treatment, 10 μL of MTT (concentration equal to 1 mg/mL) was added to each well. After incubation at 37°C for 4 h, the neoformed purple formazan crystals were solubilized in 100 μL of lysis buffer (20% sodium dodecyl sulfate in 50% dimethylformamide) overnight at 37°C. Absorbance values were measured at 495 nm in a microplate reader (model 550; Bio-Rad Laboratories, Milan, Italy) and the results expressed as absorbance.
Cell proliferation and cell cycle analysis
To assess the proliferation pattern, suspensions of 16.5×105 cells in 5 mL were seeded into ventilated flasks and the cell cultures were exposed to the EMF for 18, 36, 54, and 72 h, as previously described. At the end of the culture period, the cells were trypsinized and counted by an exclusion assay in a Bürker hemocytometer chamber using a 1:2 dilution in 0.04% trypan blue (Sigma). The results were expressed as cell number/mL.
To study the cell cycle at 72 h, suspensions of 16.5×105 cells in 5 mL were seeded into ventilated flasks. After incubation with or without exposition to the electromagnetic stimulus, 5×105 cells were trypsinized, centrifuged at 215 g for 10 min, and then washed with Dulbecco's phosphate-buffered saline (D-PBS) 1× (EuroClone). The cells were resuspended in D-PBS 1×, fixed with 70% [v/v] ethanol solution, and stored at −20°C until further analysis. Cells were then centrifuged at 500 g for 12 min at 4°C, washed with distilled water, resuspended at 37°C in 0.5 mL of D-PBS 1×containing 1 mg/mL RNase A (Sigma), and then incubated for 30 min at 37°C. A volume of 6 μL of 1 mg/mL propidium iodide (Sigma) was added and the cells were incubated for 30 min at 37°C. The DNA content was then analyzed using a flow cytometer (Epics XL; Beckman Coulter, Brea, CA) at 610 nm and the results expressed as percentage of the total cell number.
Western blot analysis
Cells were harvested and homogenized using a teflon/glass homogenizer as previously described. 40 The protein content was measured via the Bradford's method using bovine serum albumine as internal standard. Proteins were diluted in sodium dodecyl sulfate (SDS) protein gel loading solution 2×, boiled for 5 min, separated by 12% SDS-polyacrylamide gel electrophoresis, and then processed as previously described. 41 The antibodies anti-HSP70 (Santa Cruz Biotechnology, Santa Cruz, CA), anti-ADAM10 (Abcam, Cambridge, United Kingdom), and anti-α-tubulin (Sigma) were diluted at 1:1000 in TBST buffer (10 mM Tris-HCl, 100 mM NaCl, 0.1% [v/v] Tween 20, pH 7.5) containing 6% [v/v] milk.
The nitrocellulose membrane signals were detected by chemiluminescence. The experiments were performed at least three times for each different cell preparation using α-tubulin to normalize the data. Densitometric values were obtained using the NIH Image software (http://rsb.info.nih.gov/nih-image) after image acquisition and the statistical analysis was performed on the measurements.
Detection of the extracellular protein sAPPα
The sAPPα released in the medium was studied using a commercial ELISA kit and following the manufacturer's instructions (IBL International, Hamburg, Germany). Briefly, 100 μL of diluted conditioned medium was added in wells precoated with monoclonal anti-human sAPPα mouse IgG (clone 2B3). The samples were incubated overnight, washed, and then treated with HRP-conjugated anti-human APP mouse IgG (clone R101A4). The color reaction was performed using 3,3′,5,5′-tetramethylbenzidine substrate and the absorbance values were measured at 450 nm in a microplate reader (Bio-Rad Laboratories). All detections were performed in duplicate and the experiments were repeated at least three times. The results were expressed as ng/mL.
Statistics
For the statistical analysis the GraphPad InStat program (GraphPad software, San Diego, CA) was used. The data were analyzed by the analysis of variance, followed, when significant, by an appropriate post hoc comparison test, as specifically indicated in the text or in the legends. Differences were considered statistically significant when p<0.05.
Results
In this study we focused on the SH-SY5Y human neuroblastoma cells since this cellular model has been previously characterized in our lab.30,40,42 In particular, we used three different SH-SY5Y cell lines: the WT, the cells carrying the empty gene vector pIRES (pIRES), and the cells overexpressing the WT APP isoform of 751 amino acids (APP751). All the cell lines were cultured in presence or in absence of a PEMF (magnetic field intensity, 2 mT; frequency, 75 Hz) for 18, 36, 54, and 72 h. The data were analyzed both evaluating the effect of PEMF exposure on the single cell lines and aggregating the groups in treated vs. nontreated independently from the cell line used.
Effects of PEMF on mitochondrial activity and cell proliferation
To evaluate the mitochondrial activity after a continuous exposure to PEMF, the MTT test was performed on WT, pIRES, and APP751 cell lines (Fig. 2A–C). A PEMF exposure of 18, 36, and 54 h did not affect any of the cell lines investigated, as no difference in mitochondrial activity was observed between the controls and the exposed cells. After 72 h of stimulation, the mitochondrial activity decreased in all the single cell lines. This finding was further confirmed by the comparison between the aggregated controls vs. the aggregated exposed cell lines showing a statistically significant reduction only after 72 h of treatment, indicating that this modification is independent of the specific cell line evaluated (absorbance: 0.1688±0.0043 vs. 0.1140±0.0038, two-tailed p<0.0001, n=33–36).
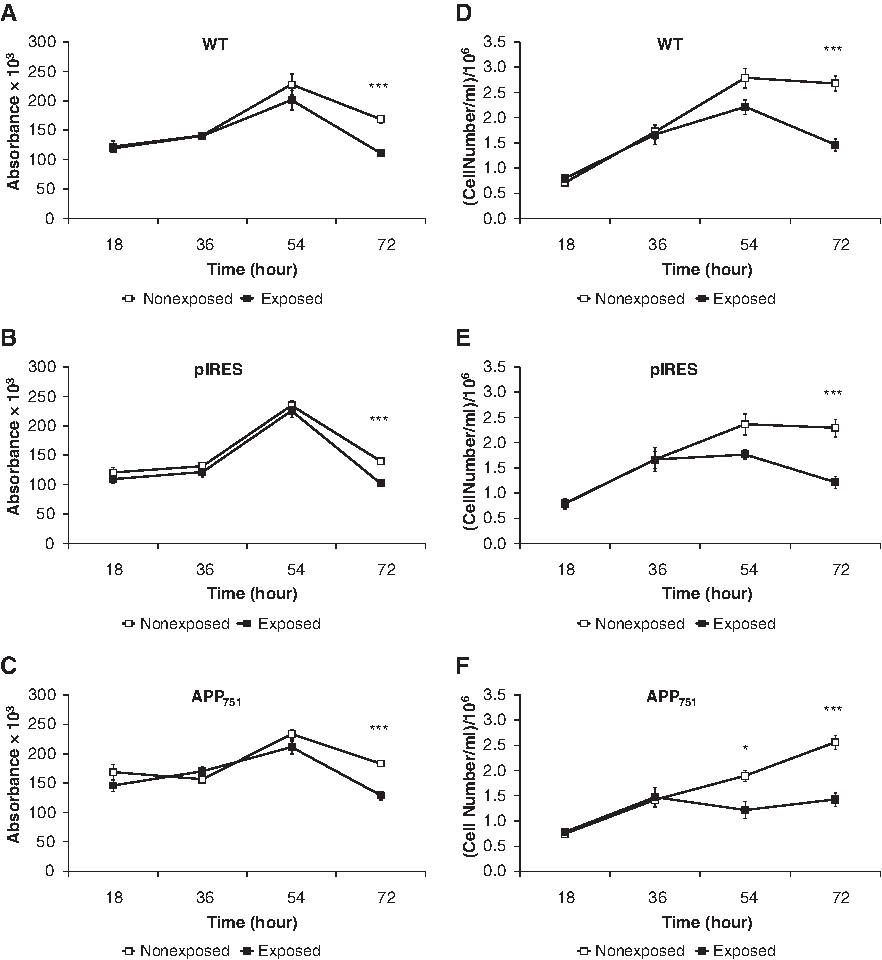
Decrease of mitochondrial activity and cell proliferation after 72 h of PEMF stimulation in WT, pIRES, and APP751 cell lines.
To better understand the effect of PEMF on cell proliferation, we performed the trypan blue exclusion test (Fig. 2D–F). After 54 h of PEMF exposure, we observed a significant decrease only in the APP751 cell line, followed by a more pronounced decline after 72 h, also detectable by microscopy (Fig. 3A, B). At this time, the reduction was also evident for both WT and pIRES cell lines. Again, after 72 h of electromagnetic stimulation, when comparing the aggregated controls vs. the aggregated exposed cell lines, the effect was independent of the specific cell line investigated (2.5×106±0.09×106 cells/mL vs. 1.4×106±0.07×106 cells/mL, two-tailed p<0.0001, n=30–33). Considering that WT and pIRES cells showed a similar profile, in the following experiments we studied only the pIRES cell line.
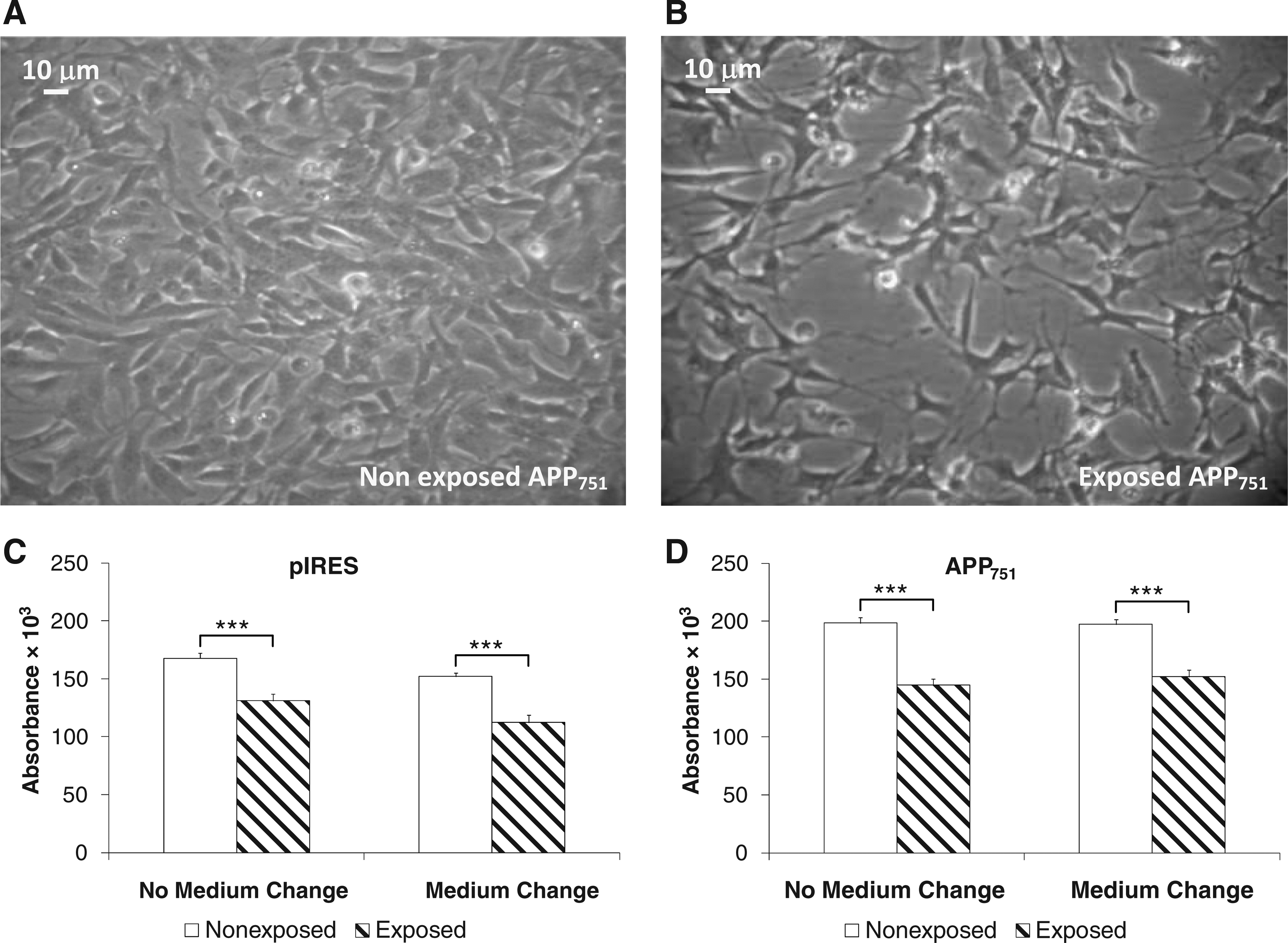
Influence of medium impoverishment in pIRES and APP751 cell lines after 72 h of PEMF exposure.
Taken together, the previous data indicate that the most significant effects on mitochondrial activity and on cell proliferation are evident after 72 h of electromagnetic stimulus. As this effect may depend on the impoverishment of the medium, due to the prolonged culture length, the MTT test was performed after 72 h of PEMF stimulus after a daily medium change. The results showed that the depletion of the medium does not affect the mitochondrial activity, as no statistically significant differences were found in pIRES and APP751 cells cultured in fresh medium in comparison with the same cells grown in exhausted medium (Fig. 3C, D). Therefore, the effect of the solenoid treatment for 72 h, which is also independent of the cell line, is only due to the electromagnetic stimulation.
Cell cycle analysis after 72 h of PEMF exposure
To gain more insight into the cell response to PEMF, we performed a cell cycle analysis with flow cytometry. We focused our attention on the 72 h stimulation as, at this time, the effect of the EMF was more pronounced on both mitochondrial activity and cell proliferation. Regarding to the cell cycle distribution, the treatment caused a statistically significant accumulation of the cells in the G0 phase, which increased by about 7% (Table 1). Moreover, a concomitant strong reduction, by about 45%, of the cells in the S phase was also found after 72 h exposure, whereas the PEMF treatment did not significantly decrease the number of cells in the G2/M phase (Table 1). Comparing the aggregated controls vs. the aggregated exposed cell lines, the effects on the G0 and S phases were confirmed (respectively: 84.9%±0.7% vs. 91.4%±0.7%, two-tailed p<0.0001, n=10; 9.7%±0.6% vs. 4.8%±0.4%, two-tailed p<0.0001, n=10); in addition, a statistically significant reduction of the cells in the G2/M phase after PEMF exposure was found (3.2%±0.5% vs. 1.3%±0.3%, p<0.005, n=10). Collectively, the data indicate that the electromagnetic stimulus causes an increase of the cells in the quiescent status.
Flow cytometry analysis of cell distribution in the different cycle phases G0, G2/M, and S (percentages expressed as mean±SEM) in pIRES and APP751 cell lines. The comparisons were performed between the exposed and the nonexposed cell lines; ap<0.01, bp<0.001, Tukey–Kramer multiple comparison test, n=5.
APP, amyloid precursor protein; PEMF, pulsed electromagnetic field.
Western blot analysis of protein expression: Focus on HSP70 and SOD-1
As reported in the literature, the low-frequency PEMF affects the redox status of the cell.14,16 For this reason, we decided to investigate the expression of two important proteins involved in stress responses: the cytosolic chaperone HSP70 and the enzyme superoxide dismutase-1 (SOD-1).
We observed that the PEMF exposure increases the expression of HSP70 by analyzing both the single cell lines (Fig. 4A) and the aggregated groups (HSP70/α-tubulin ratio: 0.6738±0.0399 [controls] vs. 1.0945±0.0422 [exposed], two-tailed p<0.0001, n=12–14).
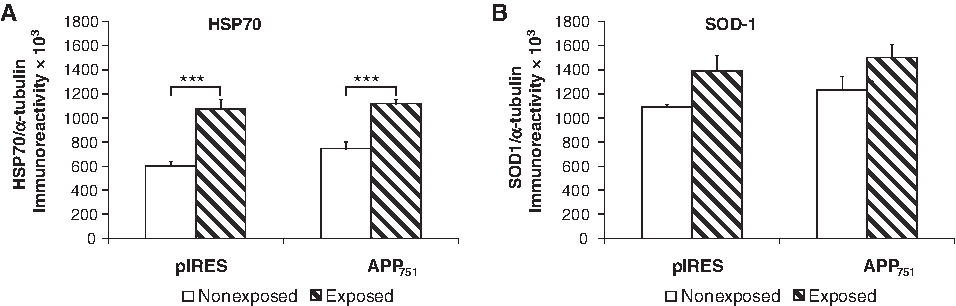
Increase of HSP70 and SOD-1 expression after 72 h of PEMF exposure in pIRES and APP751 cell lines.
An uptrend was also found in SOD-1 levels when the single cell lines were considered, although this rise did not reach the statistical significance (Fig. 4B). However, when comparing the aggregated controls vs. the aggregated exposed cell lines, the effect on SOD-1 levels resulted in a statistically significant increase after the PEMF exposure (SOD-1/α-tubulin ratio: 1.1565±0.0537 [controls] vs. 1.4441±0.0849 [exposed], two-tailed p<0.05, n=11–14).
Again, these data corroborate the hypothesis that the effect of the electromagnetic stimulus is independent of the neuroblastoma cell line evaluated.
To assess if these cellular responses might be triggered already at a very early time, we also analyzed the expression profile of both HSP70 and SOD-1 after 18 h of stimulation, but we did not detect any change (data not shown).
Effects of PEMF on the α-secretase ADAM10 and on the release of the neurotrophic factor sAPPα
We focalized on ADAM10 protein since, as already reported, due to its α-secretase activity, it is involved in the nonamyloidogenic pathway of APP. 43
The western blot analysis showed a statistically significant increase in ADAM10 levels after 72 h of electromagnetic stimulation both when considering the single cell lines (Fig. 5A) and when comparing the aggregated controls vs. the aggregated exposed cell lines (ADAM10/α-tubulin ratio: 0.8804±0.0409 [controls] vs. 1.1447±0.0492 [exposed], two-tailed p<0.05, n=18–20). We also investigated the expression of ADAM10 after a very early time (18 h of PEMF exposure), but we did not find any change (data not shown).
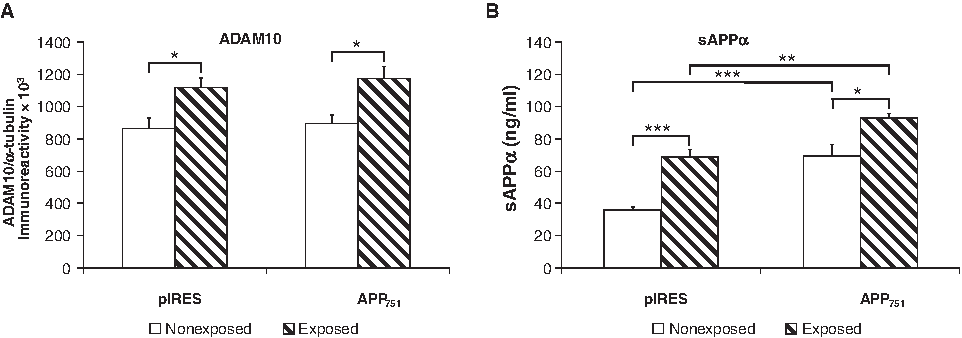
Increase of ADAM10 expression and sAPPα release in the medium after 72 h of PEMF exposure in pIRES and APP751 cell lines.
As mentioned, ADAM10 triggers the proteolysis of APP resulting in the formation of sAPPα, which is a neurotrophic factor. For this reason, we investigated whether the release of the extracellular sAPPα fragment may be altered by 72 h of electromagnetic stimulation. Since the APP751 cell line carried an additional WT copy of the gene for APP751, the corresponding basal levels of sAPPα were significantly higher in comparison with those detected in the pIRES cell line (Fig. 5B). Interestingly, a remarkable increase in the release of sAPPα was noticed after 72 h of PEMF stimulus for both pIRES and APP751 cell lines. In fact, the extracellular sAPPα levels doubled in the pIRES cell line, while increased about 25% in the APP751 cell line, although the levels in the latter remained still significantly more elevated. Again, this difference in the increase after PEMF exposure is probably due to the higher basal levels of sAPPα in the APP751 cell line. The comparison between the aggregated controls vs. the aggregated exposed cell lines confirmed the statistically significant increase of the sAPPα secretion after PEMF exposure (59.2±6.9 vs. 90.6±8.6 ng/mL, two-tailed p<0.01, n=16).
Taken together, these results indicate that the electromagnetic stimulation causes a rise in ADAM10 protein levels and a consequent release of the neurotrophic factor sAPPα.
Discussion
The present work was aimed to shed more light on the response of SH-SY5Y human neuroblastoma cells to a low-frequency PEMF stimulus. We also focused on the cell line overexpressing the WT copy of the APP751 gene to investigate whether this condition may further affect the cellular response.
Our approach relies on the concept that physiological functions or pathological conditions can be influenced by different chemical/physical/structural contexts, because not only the environmental chemistry, but also each mechanical, electromagnetic, and, in general, physical force can modulate the cell phenotype and the tissue development, as first theorized by Ingber.44–54
As a consequence, under a tissue engineering point of view and aiming toward a possible hormetic amelioration of a neuronal tissue in pathological conditions such as AD, we applied an electromagnetic stimulus to different human neuroblastoma cell lines and we studied the biological effects of this physical treatment on various parameters, detecting some basic cellular physiological functions (i.e., mitochondrial activity, cell proliferation, and cell cycle phases) and important members of stress response and of cytoprotective pathways.
In accordance with a previous study on an AD cellular model under similar electromagnetic conditions, 28 we did not find any change in the mitochondrial activity after 18 h of PEMF exposure. However, we observed a decrease after 72 h of exposure (Fig. 2A–C). A similar profile was detected for cellular proliferation (Fig. 2D–F). Moreover, the results indicate that both effects are independent of the cell line investigated, suggesting that these responses are only due to the prolonged electromagnetic stimulation independently upon a modified genetic background.
We also excluded the possible contribution of the medium impoverishment as indirect reason of the observed changes since the mitochondrial activity, after 72 h of PEMF treatment, remained lower in all the cell lines also after a daily change of the medium (Fig. 3C, D).
The results are supported also by the analysis of the cell cycle phases. As a matter of fact, after 72 h of PEMF exposure, we found an accumulation of the cells in the G0 phase with a consequent reduction in the percentage of the cells in the S and G2/M phases. Again, this effect is independent of the specific cell line examined. On the whole, the data suggest that the PEMF elicits an arrest of the cells in the quiescent status (Table 1), as reported in previous works.55,56
Interestingly, the prolonged PEMF exposure for 72 h triggered the induction of peculiar cytoprotective pathways: on one side, the overexpression of proteins implicated in the stress response (HSP70 and SOD-1) (Fig. 4), and on the other side, the activation of the nonamyloidogenic APP pathway (ADAM10 and sAPPα) (Fig. 5).
In agreement with our findings, some studies have reported that the cells respond to the electromagnetic waves through an accumulation of the cytosolic chaperone HSP70,18,19,57 which is involved in many physiological and stress-related pathways,42,58 and through an increase in SOD-1 protein, an ROS and free radical scavenger that is produced as a consequence of alterations of the redox status.14,19
Of relevance, after 72 h of electromagnetic stimulation, independently of the specific cell line, we detected an increase in the nonamyloidogenic processing of APP: as a matter of fact, we documented an enhanced expression of the α-secretase ADAM10 associated with a higher release of the neurotrophic APP metabolite sAPPα. Our results (observed using both WT SH-SY5Y line, cells carrying an empty gene vector, and cells overexpressing the APP751 gene) are different compared to those reported by Del Giudice et al., 28 who found, using cells carrying the human Swedish mutation of APP, that a low-frequency EMF causes an increase of the Aβ1–42 fragment. However, besides the differences in the cell type and dose of the stimulus, the presence of a mutated protein, which spontaneously undergoes an Aβ1–42 amyloidogenic metabolism, may have generated the dissimilarities between ours and Del Giudice et al. findings, although, as mentioned, we obtained comparable results on MTT measurements after 18 h PEMF exposure.
Collectively, our data indicate that the application of an appropriate PEMF, suitable in terms of both dose and timing, induces a kind of hormetic action resulting in some key cytoprotective responses: overexpression of HSP70 and SOD-1 and enhanced secretion of sAPPα.
These effects may be of relevance under a tissue engineering viewpoint since they suggest a possible hormetic use of the PEMF stimulation to improve the functional ability of the neuronal cells to face noxae. In line with this concept, in an interesting work on injured planaria, repeated EMF treatments activate the HSP70 pathway and other repairing mechanisms, leading to an accelerated regeneration, hypothetically modulated via the c-fos pathway. 59 To this regard, according to Pavalko's signaling model, 60 a physical stimulus like PEMF causes an increased production of c-fos,60,61 which is a component of the activator protein-1 (AP-1) transcription factor. In turn, AP-1, implicated in many cellular functions such as differentiation and response to oxidative stress, 16 may modulate the expression of ADAM10 and, consequently, the release of sAPPα. 62
Further, in vitro experiments demonstrated that the continuous exposure to a PEMF, equal to that used in the present study, stimulates both the bone cell proliferation and the biomaterial surface coating with bone extracellular matrix proteins.32–39 In addition, the use of therapeutic EMFs has been suggested to protect myocardium from ischemia reperfusion injury 63 and to treat neurodegenerative disorders such as AD.26,27 Concerning the last research field, it would be interesting to apply the physical parameters used in this study to an AD animal model, similar to the one reported by Arendash et al. 27 to evaluate, in vivo, the effects of a low-frequency electromagnetic stimulus.
In conclusion, our findings support a cytoprotective action of the electromagnetic stimulation, although further studies are required to better understand the effect of PEMF on neuronal cell responses such as differentiation, apoptosis, and their underlying signaling pathways, also applying different doses and times of the PEMF stimulus.
Footnotes
Acknowledgments
The authors are grateful to Dr. R. Cadossi and Dr. S. Setti, who presented us, generously, with the Biostim SPT pulse generator (Igea). This study was supported by INAIL Grant 2010 to Prof. G.R.
Disclosure Statement
No competing financial interests exist.