
Abstract
The effect of visible light irradiation on the expression of pluripotent genes (Oct-4, Sox2, and Nanog) in amniotic fluid-derived stem cells (AFSCs) and on the osteogenic differentiation ability of AFSCs was investigated using light-emitting diodes (LEDs) at 0–2 mW/cm2 in various wavelengths : [blue (470 nm), green (525 nm), yellow (600 nm), and red (630 nm)]. Pluripotent gene expression in AFSCs was up-regulated by visible light irradiation from a LED for more than 6 h. Green light irradiation of AFSCs up-regulated the expression of pluripotent genes more significantly than irradiation with other light. The osteogenic differentiation of AFSCs was facilitated by green and blue light irradiation. Facilitated differentiation into osteogenic cells by visible light irradiation was not mediated by reactive oxygen species (ROS); alkaline phosphatase activity (a marker of early osteogenic differentiation) and gene expression of osteopontin (a marker of late osteogenic differentiation) did not change significantly between AFSCs in differentiation medium with or without a ROS scavenger (vitamin C). The mitogen-activated protein kinase/extracellular signal-regulated protein kinase pathway, as well as other unknown signaling pathways, may be responsible for the activation of signaling pathways that facilitate the differentiation of AFSCs into osteogenic cells on light irradiation.
Introduction
Several studies have investigated the effect of light on the regulation of native cell function (e.g., genetically unchanged cells).2–4,11–16 Vinck et al. reported that green light irradiation from a light-emitting diode (LED) enhanced fibroblast growth more than red and infrared (IR) light irradiation.15,16 Liebmann et al. investigated the effects of visible light on the proliferation and differentiation of human keratinocytes and endothelial cells. 1 They found that light at wavelengths of 632–940 nm had no effect on those cells, whereas blue light reduced proliferation, in a dose-dependent manner, by up to 50%. This reduction was attributed to the induction of differentiation through an increase in differentiation markers. A photolytic release of nitric oxide was observed from nitrosated proteins, indicating that the proteins were light acceptors and might be blue light signal transducers. 1
Zhang et al. investigated the gene expression of fibroblasts irradiated with red light (628 nm) using cDNA microarray techniques. 12 The gene expression profiles revealed that more than 100 genes were regulated by red light irradiation, and these genes could be grouped into 10 functional categories. Most of these genes played important roles in the enhancement of cell proliferation and the suppression of apoptosis. Both the p38 mitogen-activated protein kinase (MAPK) signaling pathway and the platelet-derived growth factor signaling pathway were found to be involved in the cell growth induced by red light irradiation. 12
Violet-blue light (380–500 nm) has been reported to disrupt DNA integrity and cellular processes such as mitochondrial function and mitosis, which are probably mediated by light-induced reactive oxygen species (ROS).17–20 It has also been reported that the terminal enzyme of the respiratory chain, cytochrome c oxidase, which is located in the mitochondria, acts as a photoacceptor for the red-to-near IR region in eukaryotic cells; modulation of the redox state of the mitochondria is achieved by the secondary reactions of cell signaling molecules. 21
The effects of light on nerve cells have also been reported by several researchers.6,7,9–11,22–27 Giuliani et al. reported that IR lazer irradiation of a neural cell line protected the cells against oxidative stress. 11 In our previous studies, we found that the neurite outgrowth of nerve-like PC12 cells was suppressed by visible light irradiation from the lamp of an inverted light microscope.22–24 Patterned light irradiation was used for the preparation of regulated, rewritable neuronal networks analogous to “CD-RW” or “CD+RW” devices. 22 The neurite outgrowth of PC12 cells under LED irradiation was also investigated at several wavelengths. When PC12 cells were irradiated with monochromatic LED light, neurite outgrowth was slightly, but not completely, suppressed at 455, 525, 600, 630, 880, and 945 nm. 24 Long, connected neuronal outgrowths (e.g., 3 mm) were observed with 470 nm LED light at 1.8 mW/cm2 intensity. 24 These studies motivated us to see that visible light might have an effect on the function of mesenchymal stem cells (MSCs).
There are only a few studies that have investigated the effect of light irradiation on stem cells.28–33 Tuby et al. reported that low-level lazer irradiation (wavelength=804 nm) promoted proliferation of MSCs as well as cardiac stem cells in vitro. 28 Abramovitch-Gottlib et al. found that low-level irradiation from a He-Ne lazer (wavelength=632.8 nm) stimulates the development of an osteogenic phenotype in MSCs seeded on three-dimensional coralline (Porites lutea) biomatrices. 31
The effects of irradiation with different wavelengths of visible light on osteogenic differentiation of human MSCs have not yet been investigated. Here, we report the effect of visible light irradiation on the expression of pluripotent genes in human amniotic fluid-derived stem cells (AFSCs) and on the differentiation ability of AFSCs under light irradiation from LEDs with several wavelengths of light, including 470, 525, 600, and 630 nm.
Materials and Methods
Cell culture and seeding
The experiments in this study were approved by the Ethics Committees of National Central University, Li Shin Hospital, and the Cathay Medical Research Institute. Fresh, second-trimester amniotic fluid (10 mL of amniotic fluid from each patient, passage zero), obtained for fetal karyotyping by an amniocentesis performed between 16 and 20 weeks of gestation, was used with informed consent from the pregnant women. The amniotic fluid from five donors was mixed and centrifuged at 1200 rpm for 5 min, and the supernatant was discarded. The amniotic fluid cells were resuspended in culture medium A, a 40:60 solution of Dulbecco's modified Eagle's medium (D5648; Sigma-Aldrich, St. Louis, MO), and MCDB 201 (M6770; Sigma-Aldrich) supplemented with 20% fetal bovine serum (04-001-1A, lot 315200; Biological Industries, Kibbutz Beit Haemek, Israel) and 10 ng/mL of fibroblast growth factor-2 (CYT-218; ProSpec-Tany Technogene, Ness-Ziona, Israel). The cells were then cultured in a tissue culture flask (TCPS; Falcon®, BD Biosciences, San Jose, CA) in an incubator with 5% CO2 (passage one). After reaching approximately 80% confluency (approximately 5 days), the cells were trypsinized with 0.25% trypsin-ethylenediaminetetraacetic acid (25200-056; Invitrogen, Carlsbad, CA) followed by centrifugation at 1500 rpm for 5 min. The AFSCs obtained were then cultured in the tissue culture flask in culture medium A (passage two). The subsequent passage of AFSCs was prepared using the same procedures used for passage two. The AFSCs at passage three were exposed to visible light using an LED array at 1 mW/cm2 [wavelength=470 nm (blue), 525 nm (green), 600 nm (yellow), or 630 nm (red; BLAC-CLP; Ikegami Tsusho Co., Ltd., Tokyo, Japan)]; AFSCs were irradiated for 0, 1, 6, 12, 24, 36, and 48 h during growth in culture medium A; and then the gene expression analysis of Oct-4, Sox2, and Nanog was performed as described in the next section. The intensity of the LED light applied to the tissue culture flask was measured using an optical power meter (Q82017A; Advantest Co., Tokyo, Japan).
Differentiation of AFSCs into osteogenic cells
The AFSCs at passage three were cultured for 1 day in a tissue culture flask (35 3001, 35 mm diameter; Falcon, BD Biosciences) in culture medium A with seeding densities of 2×104 cell/cm2 for osteogenic differentiation. The AFSCs were rinsed with phosphate-buffered saline, and the culture medium was replaced with differentiation medium; the cells were cultured in an osteogenic medium (CCM007, Human/mouse StemXVivo Osteogenic Base Media; R&D Systems, Minneapolis, MN) with a supplement (CCM009, human/mouse StemXVivo Osteogenic supplement; R&D Systems) for up to 21 days, with a change of medium twice a week.34–37 The cells were also cultured in the differentiation medium, as just described, with the addition of 100 μM/mL of vitamin C, for the investigation of the effects of ROS on the osteogenic differentiation of AFSCs for up to 28 days, with a change of medium twice a week.
The AFSCs were exposed to visible light using an LED array [wavelength=470 nm (blue), 525 nm (green), 600 nm (yellow), or 630 nm (red; BLAC-CLP; Ikegami Tsusho Co., Ltd.)] at 0, 0.25, 0.50, 0.75, and 1.00 mW/cm2; AFSCs were irradiated for 0 (in dark as a negative control) or 48 h during the first culture period in the differentiation medium, followed by a second culture period in the dark. The intensity of the LED light applied to the tissue culture flask was measured using an optical power meter (Q82017A; Advantest Co.).
The cells were stained with Alizarin red S (A5533; Sigma-Aldrich), safranin O (S8884; Sigma-Aldrich), and von Kossa (S7276; Sigma-Aldrich) to detect the calcium deposits characteristic of osteogenic cells.38–41 Pictures of the stained cells were taken with an inverted phase contrast microscope (Eclipse Ti; Nikon, Tokyo, Japan) and a CCD camera (3.3RTV; MicroPublisher, Qimaging Surrey, BC, Canada). The six pictures (area=500×700 μm) taken of each dish were analyzed using ImageJ software (http://rsb.info.nih.gov/ij/) to calculate the number and surface area of stained and unstained cells. 42 Gene expression of osteopontin was evaluated using the method of analysis described in the next section.
Alkaline phosphatase (ALP) activity was assessed according to the manufacturer's instructions (Leukocyte alkaline phosphatase kit 85L-3R; Sigma-Aldrich) as previously described.34,43,44
Gene expression analysis
RNA was extracted from undifferentiated or differentiated AFSCs with the TRI-Reagent kit (Sigma-Aldrich) according to the manufacturer's instructions, and was subsequently treated with DNase (Ambion; Austin, TX) to remove any traces of contaminating DNA, as previously described.45,46 RNA (2 μg) was reverse-transcribed to produce cDNA using reverse transcriptase (SuperScript III First-Stand synthesis system for RT-PCR; Invitrogen, Carlsbad, CA) before being used as a template for polymerase chain reaction (PCR) amplification. Table 1 summarizes the oligomer primer pairs used for the detection of Oct-4, Sox2, Nanog, osteopontin, and GAPDH (glyceraldehyde-3-phosphate dehydrogenase) by PCR amplification. PCR amplification was performed using Taq DNA polymerase (TITANIUM™ Taq PCR Kit [Clontech Laboratories, Inc., Madison, WI]) and a thermocycler (TPersonal, Biometra, GmbH, Goettingen, Germany). The expression of Oct-4, Sox2, Nanog, osteopontin, and GAPDH mRNA was analyzed by electrophoresis; the intensities of the mRNA bands in the electrophoresis gel were measured with an image scanner (EW-97701–86; UVP® BioDoc-It® Imaging Systems, Cole-Parmer, Vernon Hills, IL) and subsequently normalized using the housekeeping gene GAPDH.
Statistical analysis
All of the quantitative results were obtained from triplicate samples. Data were expressed as means±standard deviation of triplicate samples. A Student's t-test (assuming equal variances) was performed to determine the statistical significance between experimental groups. A value of p<0.05 was considered statistically significant.
Results
Effect of light irradiation on the expression of pluripotent genes in AFSCs
The AFSCs were cultured in normal culture medium (culture medium A) under irradiation by LED light for specific time periods (0–48 h). Pluripotent gene (e.g., Oct-4, Sox2, and Nanog)47–49 expression of AFSCs under blue light irradiation was used to determine the optimal irradiation time facilitating the differentiation of AFSCs. Oct-4 and Sox2 are transcription factors essential to maintaining the pluripotent embryonic stem cell (ESC) phenotype. 47 Therefore, the dependence of the expression of Oct-4, Sox2, and Nanog in AFSCs on the duration of light irradiation was investigated with a blue light (470 nm) at a fixed intensity of 1 mW/cm2 and is shown in Figure 1A. Blue light irradiation of AFSCs for more than 6 h produced enhanced gene expression of Oct-4, and that of Sox2 was also found to be enhanced in AFSCs under irradiation after 6 h (p<0.05). Since pluripotent gene expression, especially Nanog, was enhanced after irradiation with blue light for more than 24 h, the light irradiation period was fixed at 48 h in the following experiments. The osteogenic differentiation of AFSCs was enhanced by light irradiation for 48 h, as compared with differentiation after irradiation for 24 or 36 h (data not shown).

The effect of light irradiation on the expression levels of pluripotent genes in AFSCs.
The effect of the wavelength of light on the expression of Oct-4, Sox2, and Nanog in AFSCs was also investigated using blue (470 nm), green (525 nm), yellow (600 nm), and red (630 nm) LED lights. The light intensity was fixed at 1 mW/cm2 with a 48 h irradiation time; the results are shown in Figure 1B. No significantly enhanced expression of pluripotent genes (Oct-4 and Sox2) in AFSCs irradiated with red light was observed, and considerably enhanced expression was found in these genes after irradiation with yellow, green, and blue light (p<0.05). The most significant enhancement of gene expression in this study was observed after irradiation with green light.
Effects of light irradiation intensity on osteogenic differentiation
The AFSCs were cultured in osteogenic differentiation medium under irradiation by LED light for 0 (in dark as a negative control) or 48 h to investigate the effect of light irradiation on the osteogenic differentiation of AFSCs. Figure 2 shows Alizarin red S staining of AFSCs cultured in differentiation medium for 14, 21, and 28 days with blue light irradiation for 0 or 48 h at a light intensity of 1 mW/cm2. A red color after Alizarin red S staining indicates the deposition of calcium caused by the differentiation of AFSCs into osteogenic cells.40,41 Calcium deposition (Alizarin red S staining) was significantly enhanced in AFSCs irradiated with blue light compared with those cultured in the dark (conventional method). The dependence on the intensity of the light to effect osteogenic differentiation (as measured by the Alizarin red S staining area, the ALP activity, and the gene expression intensity of the osteogenic differentiation marker) was investigated in AFSCs irradiated with blue light, and the results are shown in Figure 3. The Alizarin red S staining area (Fig. 3A) and ALP activity (an early marker of osteogenic differentiation, Fig. 3B) 50 in AFSCs were found to be the largest and highest, respectively, at a light intensity of 1 mW/cm2 of blue light irradiation. The expression of the osteopontin gene (a late marker of osteogenic differentiation) was higher in blue light-irradiated AFSCs than in AFSCs cultured in the dark (p<0.05), when the blue light intensity was higher than 1.0 mW/cm2 (Fig. 3C). In addition, both the ALP activity and Alizarin red S staining domains increased after blue light irradiation of AFSCs at a light intensity of more than 1 mW/cm2. Therefore, the light irradiation intensity was fixed at 1 mW/cm2 in subsequent experiments on osteogenic differentiation of AFSCs.

Alizarin red S staining of AFSCs cultured in differentiation medium for 14
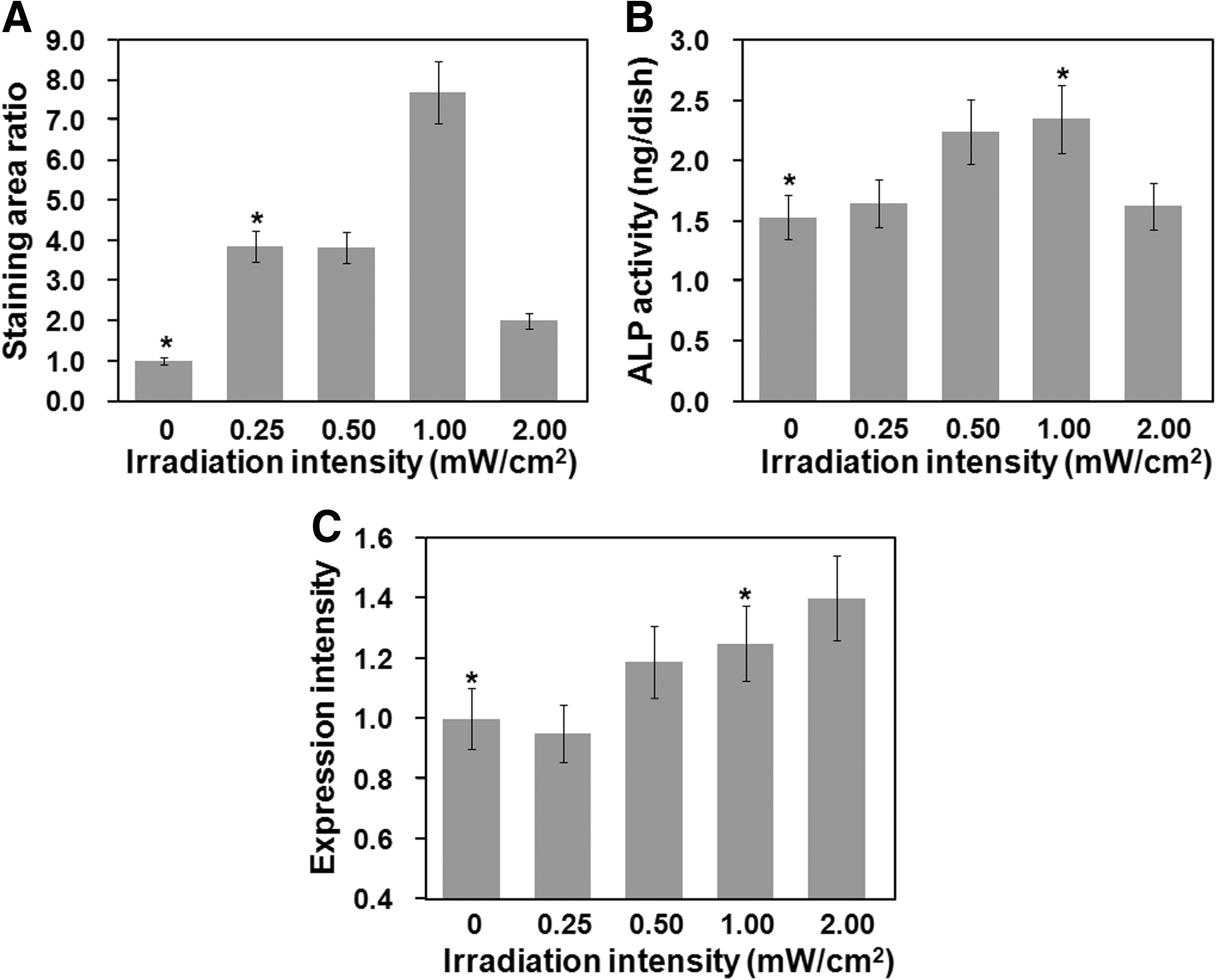
The effect of light irradiation intensity on the osteogenic differentiation of AFSCs.
The effect of the wavelength of light on osteogenic differentiation
The effect of the wavelength of light on the ability of AFSCs to differentiate into osteogenic cells was investigated using LED lights at blue (470 nm), green (525 nm), yellow (600 nm), and red (630 nm) wavelengths, a light intensity of 1 mW/cm2 and an irradiation time of 48 h; the results of these experiments are shown in Figures 4–7. Figure 4 shows the morphology of the AFSCs after 3 weeks of culturing. Figure 5 shows Alizarin red S staining (calcium deposition) of AFSCs cultured in the differentiation medium for 21 days under different wavelengths of light for 0 or 48 h with an intensity of 1 mW/cm2. The Alizarin red S staining area of AFSCs irradiated with blue or green light was found to be greater than that in cells irradiated with red or yellow light, or that of cells cultured in the dark (Fig. 5A and B). Histological evaluation of mineralization was also assessed by von Kossa staining for calcium phosphate deposition (Fig. 6). Intense von Kossa staining was seen in AFSCs irradiated with blue or green light than that in cells irradiated with red or yellow light, or that of cells cultured in the dark.

Morphology of AFSCs after growing for 21 days with no light
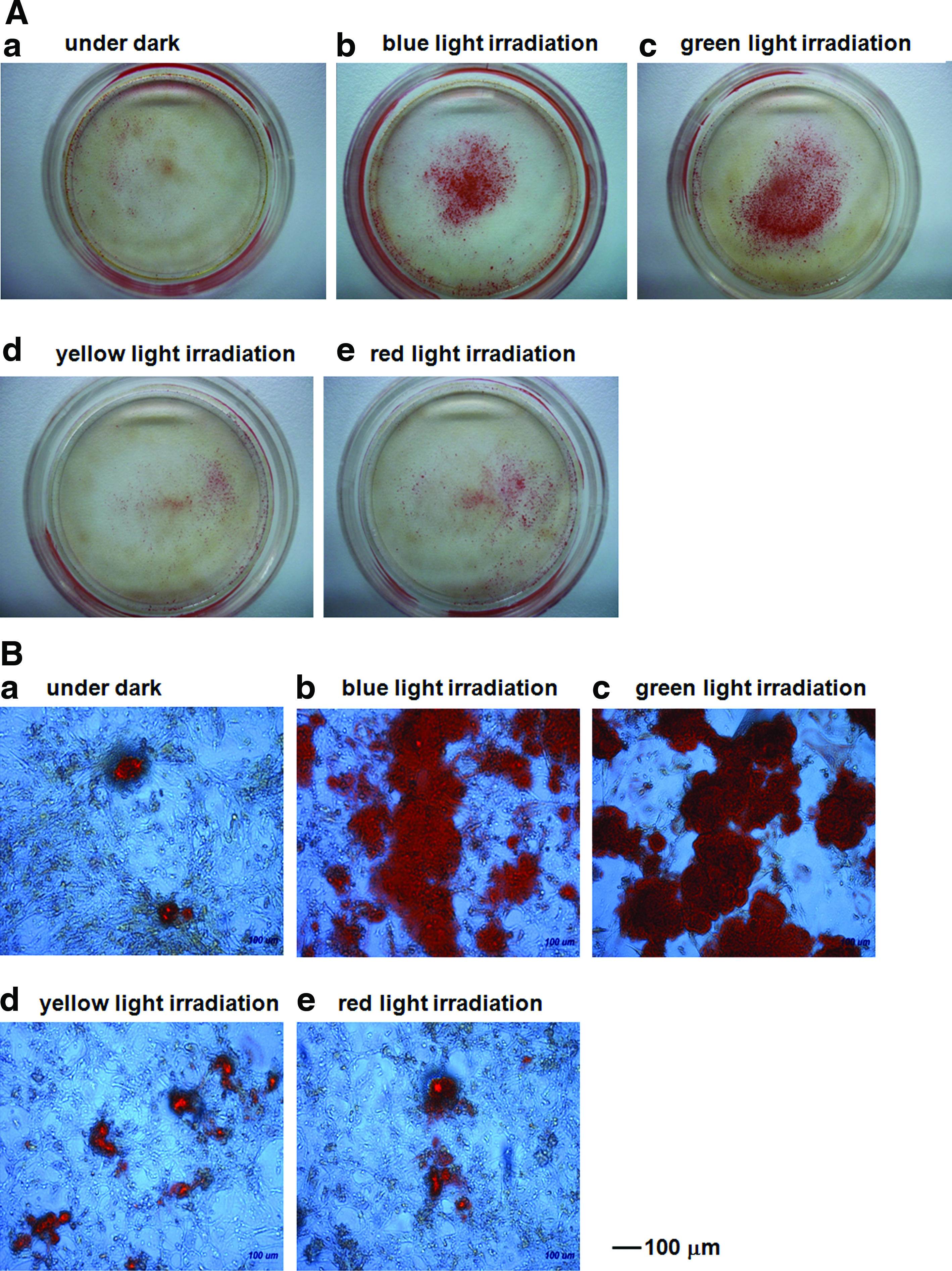
Alizarin red S staining of AFSCs after growth for 21 days with no light
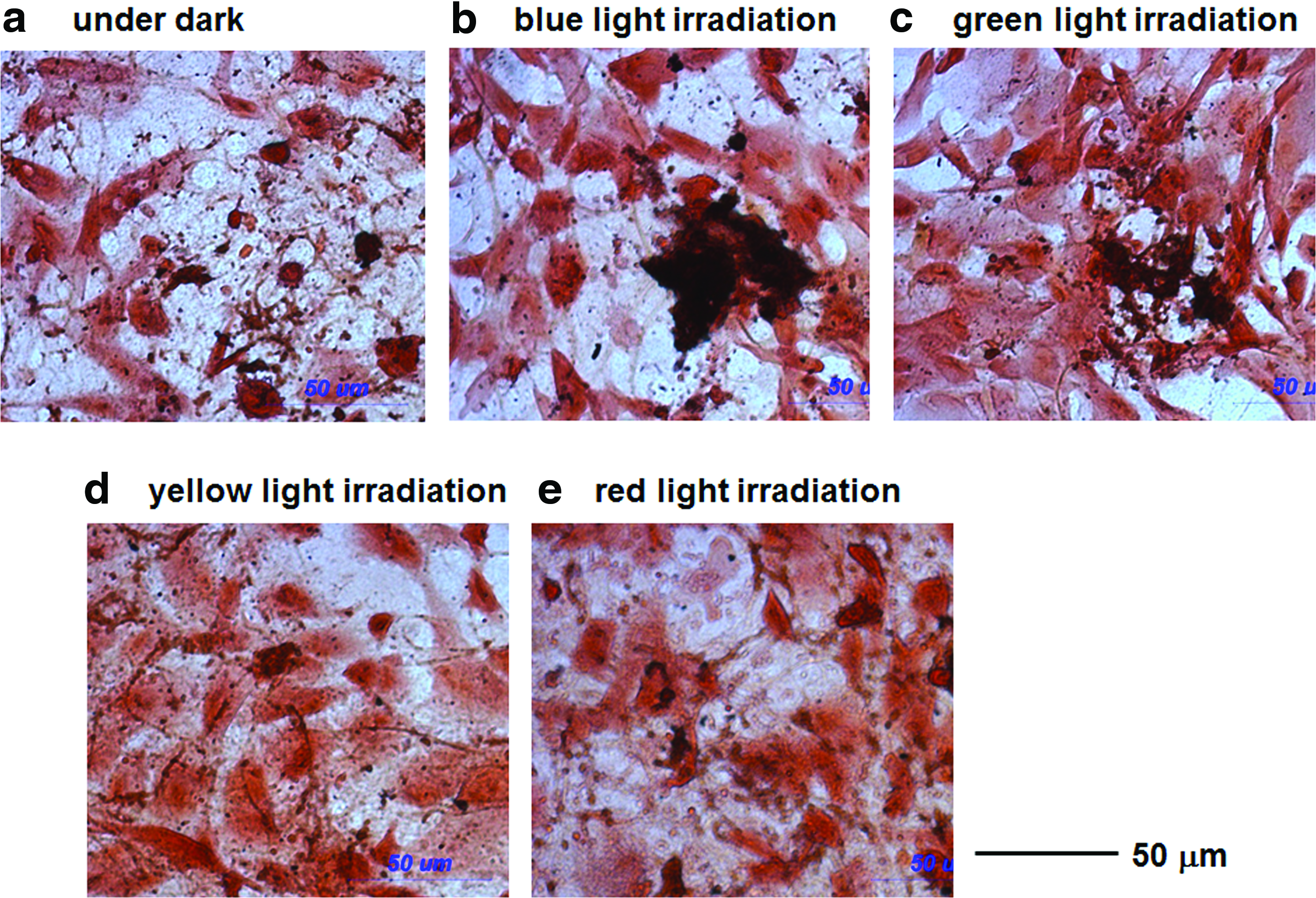
von Kossa and safranin o staining of AFSCs after growth for 21 days with no light

The effect of the wavelength of light on the osteogenic differentiation of AFSCs irradiated with a light intensity of 1 mW/cm2 for 48 h.
Quantitative analysis of Alizarin red S staining, enzymatic activity of ALP, and expression of an osteogenic differentiation marker (osteopontin) was investigated in AFSCs cultured in differentiation medium for 14 days under irradiation with different wavelengths of light for 0 or 48 h at a light intensity of 1 mW/cm2, and the results are shown in Figure 7. The Alizarin red staining area (Fig. 7A) of AFSCs irradiated with blue light or green light was quantitatively higher than that of cells irradiated with red or yellow light, or of cells incubated in the dark (p<0.05). The ALP activity (Fig. 7B) of AFSCs irradiated with blue light was quantitatively higher than that of cells irradiated with yellow light, or of cells incubated in the dark (p<0.05). The gene expression levels of osteogenic differentiation markers (i.e., osteopontin) were found to be higher in AFSCs irradiated with blue light when compared with AFSCs cultured in the dark (p<0.05; Fig. 7C).
Discussion
It has been reported that illumination with strong white light can give rise to the formation of free radicals, which subsequently leads to specific protein degradation as a consequence of ROS-induced cleavage of the polypeptide backbone by free radicals. 51 Further, blue light was reported to act by forming intracellular ROS, which then affected critical cell functions. 52 Lockwood et al. found that dose-dependent ROS levels were generated in both NHEK and OSC2 cells by blue light irradiation; the induction of intracellular ROS implies that redox effects mediated the cellular responses to blue light. 52
One possible mechanism for the facilitated differentiation of AFSCs into osteogenic cells by visible light irradiation is mediation by ROS, which have been shown to be secondary messengers in cell signal transduction.53,54 It was reported that ROS-enhanced differentiation of human ESCs (hESCs) into bipotent mesendodermal cells occurs via ROS-involved signaling pathways. 55 In particular, expression of the pluripotency markers (Oct-4, Tra 1–60, Nanog, and Sox2) of hESCs was decreased in ROS-inducing conditions, whereas expression of mesodermal and endodermal markers increased. 55 The MAPK family-specific inhibitors were reported to prevent the mesendodermal differentiation of hESCs via ROS induction. 55
We have evaluated the ability of AFSCs to differentiate into osteogenic cells when irradiated with blue light (2 h, 1 mW/cm2) in the presence of free radical scavengers in the differentiation medium. Vitamin C (100 μM) was selected as the free radical scavenger in this study.55,56 No morphological changes in AFSCs were found when using differentiation media with and without free radical scavengers (data not shown). Figure 8A shows that the ALP activity of AFSCs differentiated into osteogenic cells after blue light irradiation with and without a free radical scavenger. Notably, free radical scavenger treatment of AFSCs neither prevented nor promoted osteogenic differentiation when compared with differentiation medium that did not contain a free radical scavenger, as determined by ALP activity (a marker of early osteogenic differentiation). Gene expression of an osteogenic differentiation marker (osteopontin, a marker for late osteogenic differentiation) was investigated in AFSCs after irradiation with blue light in differentiation medium with and without a free radical scavenger, as is shown in Figure 8B. The gene expression of the osteogenic differentiation marker did not change significantly between AFSCs in differentiation medium with and without a free radical scavenger. These results indicate that the facilitated differentiation into osteogenic cells by irradiation with visible light found in this study is not mediated by ROS, but by another mechanism.
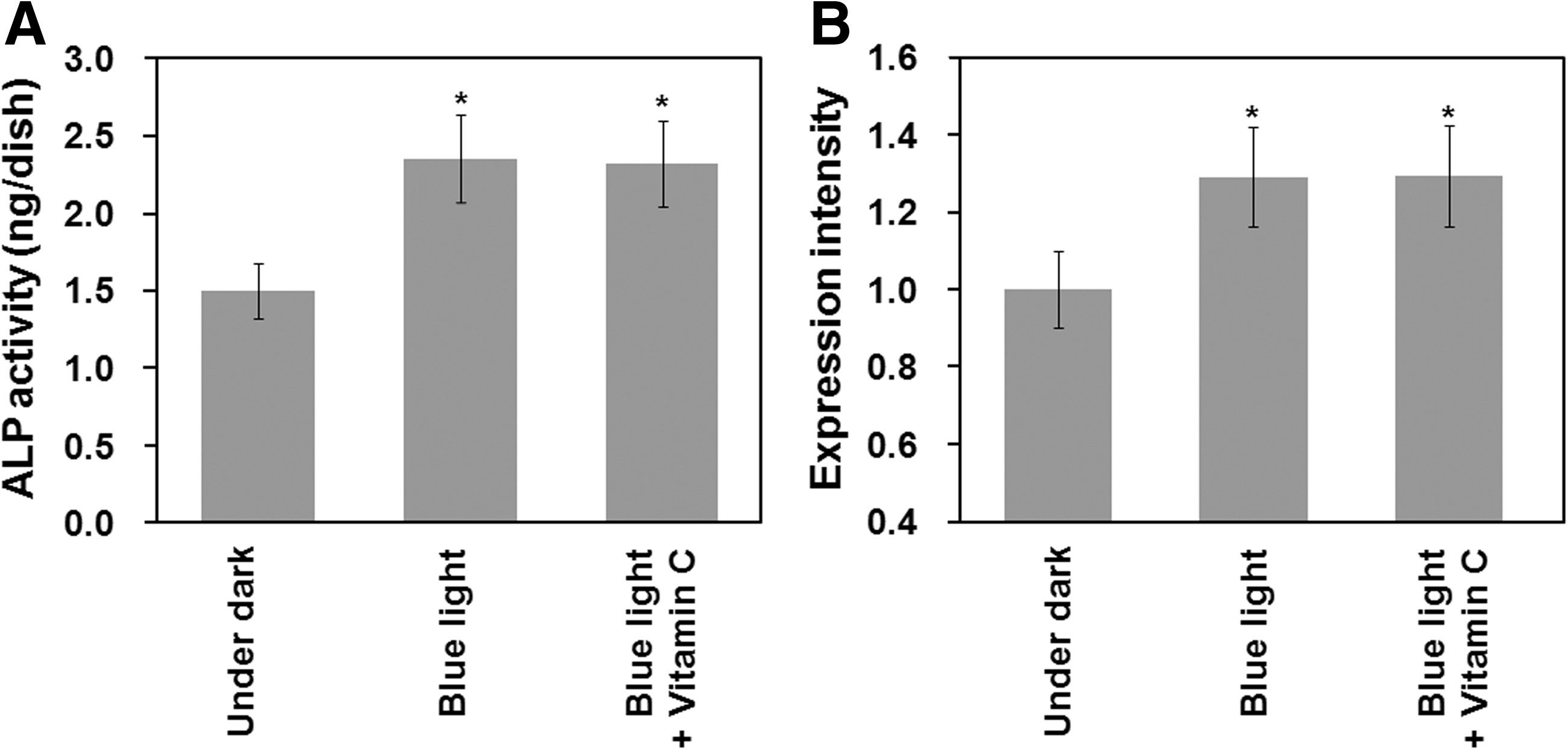
The effect of reactive oxygen species on the facilitated differentiation of AFSCs into osteoblasts after irradiation with blue light at an intensity of 1 mW/cm2 for 48 h.
Low-level light irradiation from LEDs has been demonstrated to enhance cell proliferation and cytokine secretion in several cell types. 57 Light irradiation generated low-energy stimulation of tissues and cells, which resulted in increased cellular activity, increased proliferation of fibroblasts and keratinocytes, and enhanced growth-factor synthesis, angiogenesis, and collagen production.29,58 Li et al. investigated the effects of red-light LED irradiation on the proliferation and osteogenic differentiation of rat MSCs. 29 They found that the growth of rat MSCs was enhanced by red-light irradiation, the effects of which became more obvious at low-cell densities. 29 In their research, increases in both ALP activity and osteocalcin expression of around 10% were found in the rat MSCs receiving red-light irradiation. 29 However, red-light irradiation of AFSCs was not powerful enough to facilitate osteogenic differentiation, which was observed with green and blue light irradiation in this study.
Liebmann et al. investigated the effect of light irradiation on proliferation and differentiation in human skin cells. 1 They found that light wavelengths of 632–940 nm had no effect, but irradiation with light at 412–426 nm had toxic effects at high fluencies; however, light at 453 nm was found to be nontoxic up to a fluence of 500 J/cm2. 1 In the present study, we irradiated cells for 48 h at 1 mW/cm2, which corresponds to 172.8 J/cm2. Further, the wavelengths of light used in this study (470, 525, 600, and 630 nm) were longer than those used in Liebmann's study (453 nm). 1 Therefore, the facilitated differentiation of AFSCs into osteogenic cells by light irradiation is not caused by the toxic effect of light irradiation on AFSCs.
Some researchers have reported that light irradiation activates MAPK/extracellular signal-regulated protein kinase (ERK), which is a signal for cellular proliferation, differentiation, and survival. However, light irradiation was not reported to activate the stress signals of p38 MAPK and c-Jun N-terminal kinase.29,59,60
Although the evidence for the facilitated differentiation of AFSCs into osteogenic cells is clear in this study, the molecular mechanisms of biomodulation by visible light irradiation remain unknown. It has been suggested that the photoreceptor molecules for violet to blue radiation are flavoproteins, which are found in the beginning of the respiratory chain; whereas those for red to near IR radiation are cytochrome c oxidases, found at the end of the respiratory chain.29,61 The activation of photoreceptors generates a series of primary responses in cells, which then lead to secondary reactions and result in increases in cell proliferation and cellular activities.29,61 It has been reported that several genes related to antioxidation and mitochondrial energy metabolism were up-regulated or down-regulated due to light irradiation. 29 The MAPK/ERK pathway, as well as other unknown signal pathways, is likely responsible for the activation of signal pathways that facilitate the differentiation of AFSCs into osteogenic cells on light irradiation.
Conclusion
Pluripotent gene expression in AFSCs was up-regulated when cells were exposed to visible light irradiation from an LED for more than 6 h. Green light irradiation up-regulated pluripotent genes in AFSCs more significantly than irradiation with blue, yellow, or red light.
The differentiation of AFSCs into osteogenic cells was facilitated by green or blue light irradiation from an LED, but not significantly by red or yellow light, although several researchers have reported positive effects on the proliferation and osteogenic differentiation of MSCs using red LED light irradiation.29,31 In this study, the ability of visible light irradiation to facilitate differentiation into osteogenic cells is not mediated by ROS, but by another mechanism. Further study of the effects of light irradiation on the enzymatic activities and signal pathways of AFSCs will elucidate the mechanism of the photoactivation found in this study.
Footnotes
Acknowledgments
This research was partially supported by the National Science Council of Taiwan under Grant Nos. NSC97-2221-E-008-011-MY3, NSC98-2120-M-008-002, and NSC99-2120-M-008-001. This work was also supported by the VGHUST Joint Research Program, Tsou's Foundation (VGHUST97-P3-08 and VGHUST98-P3-11), and the Cathay General Hospital Project (98CGH-NCU-B1, 99CGH-NCU-B2). Grants-in-Aid for Scientific Research (No. 21500436) from the Ministry of Education, Culture, Sports, Science, and Technology of Japan are also acknowledged. The work was performed at National Central University, Jhongli, Taoyuan, Taiwan.
Disclosure Statement
No competing financial interests exist.