
Abstract
Articular cartilage is organized in depth zones with phenotypically distinct subpopulations of chondrocytes that are exposed to different oxygen tensions. Despite growing evidence of the critical role for oxygen in chondrogenesis, little is known about its effect on chondrocytes from different zones. This study evaluates zonal marker expression of human articular chondrocytes from different zones under various oxygen tensions. Chondrocytes isolated from full-thickness, superficial, and middle/deep cartilage from knee replacement surgeries were expanded and redifferentiated under hypoxic (5% O2) or normoxic (20% O2) conditions. Differentiation under hypoxia increased expression of hypoxia-inducible factors 1alpha and 2alpha and accumulation of extracellular matrix, particularly in middle/deep chondrocytes, and favored re-expression of proteoglycan 4 by superficial chondrocytes compared with middle/deep cells. Zone-dependent expression of clusterin varied with culture duration. These results demonstrate that zonal chondrocytes retain important phenotypic differences during in vitro cultivation, and that these characteristics can be improved by altering the oxygen environment. However, transcript levels for pleiotrophin, cartilage intermediate layer protein, and collagen type X were similar between zones, challenging their reliability as zonal markers for tissue-engineered cartilage from osteoarthritis patients. Key factors including oxygen tension and cell source should be considered to prescribe zone-specific properties to tissue-engineered cartilage.
Introduction
Chondrocytes from different zones exhibit distinct morphologies and protein expression. Therefore, engineering tissue with a zonal structure requires a thorough understanding of the in vitro behavior of zonal chondrocytes as well as a set of suitable markers that define their different phenotypes. The flattened cells of the superficial zone have been characterized by expression of specific proteins, such as clusterin 4 and proteoglycan 4 (PRG4). 5 Pleiotrophin (PTN), an embryonic growth and differentiation factor, has also been found in the superficial zone. 6 The more spherical and randomly distributed chondrocytes from the middle layers have been found to specifically express cartilage intermediate layer protein (CILP) 7 and collagen type X (COL10A1), often used as a surrogate indicator of hypertrophy, is usually confined to the calcified, deeper regions of normal articular cartilage. 8
Among zonal marker proteins, the zone-specific functions of PRG4 are most clearly understood. This highly glycosylated protein is secreted into the synovial cavity by cells lining the synovial joint,5,9 where it plays a key role in boundary lubrication.10,11 During osteoarthritis, PRG4 levels in affected joints are down-regulated, which potentially contributes to the progression of the disease. 12 Consistent with this, treatment with recombinant PRG4 decreased cartilage degeneration in a rat model of osteoarthritis. 13 PRG4 also has antiadhesive properties that may inhibit integration of tissue-engineered constructs with native cartilage. 14 Therefore, restoring in vivo stratification may have particular importance for the mechanical and metabolic properties of cartilaginous constructs.
The cells of different zones are exposed to different levels of nutrients due to the avascular nature of articular cartilage. For example, the oxygen partial pressure declines from ∼7% to 10% at the superficial layer of articular cartilage to <1% near the subchondral bone. 15 The response of chondrocytes to oxygen deprivation is regulated by the transcription factors of the hypoxia-inducible factor (HIF) family. 16 HIF-1α appears to play a role in chondrocyte survival and energy generation,17,18 whereas HIF-2α seems to control hypoxia enhanced chondrogenic differentiation. 19 Despite the growing evidence for a critical influence of reduced oxygen on chondrogenesis, 20 little is known about the zone-specific effects of oxygen gradients.
We hypothesized that chondrocytes exhibit a zone-specific response to oxygen concentration during in vitro redifferentiation and that the re-expression of specific markers of cartilage zones can be regulated by varying oxygen concentration. To test our hypothesis, we isolated and expanded human zonal articular chondrocytes and redifferentiated them in micromass pellets under normoxic (20% O2) and hypoxic (5% O2) conditions. Pellets were then assessed for DNA and glycosaminoglycan (GAG) contents, HIF localization, and expression of zonal marker and matrix proteins.
Materials and Methods
Cell isolation
Articular cartilage was obtained with ethics approval from femoral condyles of consenting patients (three female donors, aged 60–70 years) undergoing arthroplasties due to osteoarthritis. Tissue from macroscopically “normal” (International Cartilage Repair Society anatomical grade 0–1
21
) regions of osteoarthritic cartilage were harvested. For zonal cartilage harvest, 6 mm cores were punched, and ∼300–500 μm of the superficial layer was processed separately from the remaining middle/deep tissue. Cartilage slices were minced, washed in Dulbecco's phosphate buffered saline (PBS; Invitrogen, Carlsbad, CA), and digested overnight with 0.15% w/v collagenase type 2 (Worthington, Lakewood, NJ) in serum-free, low-
Cell expansion
Freshly isolated chondrocytes were seeded at 3,000 cells/cm2 in tissue culture flasks (Nunc/Thermo Fisher, Waltham, MA) and cultured in low-glucose basal medium supplemented with 0.1 mM L-ascorbic acid (WAKO Chemical, Osaka, Japan) and 10% fetal bovine serum (Hyclone, Logan, UT) at 37°C in a humidified 5% CO2/95% air incubator. Medium was refreshed twice per week. Subconfluent cells were released from culture flasks by washing twice with PBS and incubating in 0.25% trypsin with 1 mM ethylenediamine tetraacetic acid (EDTA; Invitrogen) at 37°C for 5 min. Cells were counted with a NucleoCounter (Chemometec, Allerød, Denmark) and used for differentiation experiments after three passages. Population doubling times and total population doublings were calculated and compared between groups.
Chondrogenic differentiation
The redifferentiation capacity of the expanded chondrocytes was evaluated by pellet culture in serum-free chondrogenic media (high-glucose basal medium supplemented with 1.25 mg/mL bovine serum albumin (BSA), 10−7 M dexamethasone, 1% v/v insulin, transferrin, selenium, linoleic (ITS+1) (all Sigma), 0.1 mM L-ascorbic acid (WAKO Chemical), and 10 ng/mL transforming growth factor type beta 1 (TGF-β1; Chemicon/MILLIPORE, Billerica, MA). Pellets were formed by centrifugation of 2.5×105 cells per well in polymerase chain reaction (PCR) microplates (Axygen Scientific, Union City, CA) at 600×g and transferred into 24-well plates after day 3. Medium was refreshed twice per week. Pellets were cultured either at ∼20% oxygen in a humidified CO2 incubator or at 5% oxygen in a ProOx C-Chamber (Biospherix, Redfield, NY) inside the same incubator over 28 days. The partial oxygen tension in the ProOx chamber was maintained through an automatically controlled supply of 5% CO2/95% N2, which partially exchanged the gas in the chamber until 5% oxygen was reached.
DNA quantification
The Quant-iT™ PicoGreen® dsDNA assay (Invitrogen) was used to quantify the DNA content in pellet cultures. Briefly, cell pellets were weighed and incubated overnight at 60°C with 0.5 mg/mL proteinase K (Invitrogen) in a phosphate buffer (20 mM Na2HPO4, 30 mM NaH2PO4.H2O, 5 mM EDTA, and pH 7.1). Samples were mixed with the dye solution, and fluorescence was measured with a microplate reader (BMG Labtech, Offenburg, Germany) at 485 nm excitation and 520 nm emission. DNA content was calculated from λ phage DNA standards (Invitrogen).
Quantification of GAGs
The GAG content of the pellets was quantified using the 1,9-dimethylmethylene blue dye (DMMB). Briefly, proteinase K-digested samples were mixed 1 in 10 with 46 μM DMMB chloride (Sigma) dissolved in a glycine buffer (40 mM glycine, 40 mM NaCl, 0.002% m/v sodium azide, and pH 3). Absorbance at 525 nm was measured in a microplate spectrophotometer (Bio-Rad Laboratories, Hercules, CA). GAG content was calculated from chondroitin-6-sulphate standards (Sigma).
mRNA expression analysis by quantitative reverse transcription-PCR
Quantitative reverse transcription ((q)RT)-PCR was applied to quantify mRNA levels in chondrocyte pellet cultures. Total RNA was isolated from pellet cultures using TRIzol® Reagent (Invitrogen) following the manufacturer's protocol with modifications for high proteoglycan content. Briefly, after chloroform-guided phase separation, RNA was precipitated from the aqueous layer with 0.5 volumes of 2-propanol, 0.5 volumes of salt solution (1.2 M NaCl, 0.8 M sodium citrate), and 16 μg/mL linear polyacrylamide (Ambion, Austin, TX) at −80°C overnight. Pelleted RNA was washed with 70% ethanol and resuspended in water. Total RNA was quantified by UV spectroscopy. Only samples that had A260/A280 ratios of at least 1.8 and passed the agarose tris hydroxymethylaminoethane-acetate-EDTA gel analysis were used.
Primers used in qRT-PCR are listed in Table 1. Samples with 300 ng of total RNA were treated with DNase I (Invitrogen) and reverse transcribed into complementary DNA (cDNA) with SuperScript™ III first-strand synthesis supermix (Invitrogen) following the manufacturer's protocols. Absolute quantification was carried out using standard curves. 22 All reactions were performed in duplicate in 7.5 μL volumes in a 384-well plate using Express SYBR® GreenER™ qPCR supermix universal (Invitrogen) and a 7900HT fast real-time PCR system (Applied Biosystems, Foster City, CA). Reactions contained 1× qPCR mix, 132 nM of each forward and reverse primers, and 0.2 μL of undiluted cDNA. PCR amplification followed a three step cycling protocol with an initial 5 min incubation at 50°C, 2 min denaturation at 95°C, and 40 cycles of 95°C for 15 s and 60°C for 30 s. Amplification of the correct sequence during real-time reactions was assessed by a postamplification melt curve analysis. Real-time curves were analyzed with ABI Sequence Detection System software version 2.3 (Applied Biosystems). The cDNA copy number for each reaction was then calculated by direct comparison to the known standards and normalized to 18S rRNA. Transcript levels of middle/deep and superficial chondrocytes of each data point were further normalized to mRNA levels of the respective full-thickness controls.
Safranin O staining
Sections from pellet cultures were stained with safranin O for GAG. Briefly, fresh frozen samples embedded in O.C.T. Compound (Sakura Finetek) were cut to yield ∼5 μm thick sections. Sections were fixed with 100% cold acetone and stained using hematoxylin, Fast Green (0.001% w/v), and safranin O (0.1% w/v; all from Sigma). 23 Images were captured with a MicroPublisher 3.3 RTV (QImaging, Surrey, Canada) mounted on a microscope (BX41; Olympus, Tokyo, Japan).
Immunohistochemical and immunofluorescence analysis
To determine the protein expression of PRG4, clusterin, collagen type I (COL1A1), and type II (COL2A1), fresh-frozen sections from chondrocyte pellets and native osteoarthritic cartilage, which had been treated with 0.1 μM monensin (Sigma) in culture medium overnight, 24 were fixed in 100% cold acetone for 15 min. Antigen retrieval was included in the collagen staining procedures by applying 0.1% w/v hyaluronidase (Sigma) in PBS for 20 min at 37°C before blocking sections with 2.5% normal horse serum (Vector Laboratories, Burlingame, CA) for 20 min. Samples were incubated with antibodies against either lubricin (PRG4; 1 h at ∼22°C, 1:200, Abcam ab28484, Sapphire Bioscience, Redfern, Australia), clusterin (2.5 h at ∼22°C, 2G7, 1:200, generous gift from Dr. Brendan Murphy 25 ), collagen type I (overnight at ∼4°C, 1:300, I-8H5, MP Biomedicals, Solon, OH), or type II (overnight at ∼4°C, 1:100, II-II6B3; Developmental Studies Hybridoma Bank [DSHB], Iowa, IA) in a humidified chamber. Control samples were probed in parallel with IgG isotype control antibodies (rabbit IgG, 1:1000, Abcam ab37415, Sapphire Bioscience; mouse IgG, 1:500, Invitrogen). Subsequently, biotinylated secondary anti-rabbit/mouse antibody (Vector Laboratories) was applied to the sections for 30 min, followed by incubation with avidin and biotinylated horseradish peroxidase complex reagent (Vector Laboratories) for 30 min in a humidified chamber. A 5 min wash in PBS was performed after each step. Chromogens were allowed to develop over 5 min by using ImmPACT™ DAB Substrate (Vector Laboratories). Samples were rinsed with tap water for 5 min and counterstained in Mayer's hematoxylin (Amber Scientific, Midvale, Australia). Images of samples were captured, as just described.
To determine the protein expression of HIF-1α and HIF-2α, fresh-frozen sections from chondrocyte pellets were fixed in 100% cold acetone for 15 min, blocked with 2% w/v BSA/0.1% v/v Triton X-100 in PBS for 20 min, and probed with antibodies against either HIF-1α or HIF-2α (1 h at ∼22°C, 1:100, Novus Biologicals NB100-105/NB100-122, Sapphire Bioscience). Subsequently, samples were incubated with AlexaFluor®488-labelled secondary antibodies (for HIF-1α, anti-mouse, Invitrogen; for HIF-2α, anti-rabbit, Cell Signaling, Danvers, MA; both 1:400) and 5 μg/mL 4′,6-diamidino-2-phenylindole (Sigma) for 1 h at room temperature. Control sections were treated similarly without incubation with a primary antibody. Images were captured with fixed exposure time on an Axio Imager.A1 microscope with epi-fluorescence attachment (Carl Zeiss, Jena, Germany).
Western blot analysis
To test for the amount of PRG4 protein secreted by chondrocytes into the media, conditioned media was collected and pooled from pellets of zonal chondrocytes during week 4 of differentiation culture. Proteins in the media were concentrated by acetone precipitation, and their amounts were quantified by Bradford assay (Sigma). Protein samples, a control of recombinant human lubricin (∼240 kDa, characterized in, 13 generous gift of Dr. Eric Vanderploeg, Pfizer Inc.) and a molecular-weight marker standard (Bio-Rad Laboratories) were mixed nonreduced with NuPAGE® lithium dodecylsulfate Sample Buffer (Invitrogen), electrophoresed on a NuPAGE Novex 3%–8% Tris-Acetate gel (Invitrogen; 100 μg protein or 2.5 μg LUB1 per lane) and transferred onto a nitrocellulose membrane (PALL, Pensacola, FL). The membrane was blocked with 2% w/v skim dry milk in a solution of Tris-buffered saline/Tween-20 and probed with a lubricin antibody (over night at 4°C, 1:1000, Abcam ab28484) and horseradish peroxidase-conjugated anti-rabbit antibody (45 min at ∼22°C, 1:4000, Abcam ab6721/Sapphire Bioscience). The membrane was then incubated in ECL substrate solution (Pierce/Thermo Fisher), and protein bands were visualized in a ChemiDoc™ system (Bio-Rad Laboratories).
Statistical analysis
Statistical analyses were performed using Minitab 15.1 software (Minitab Inc., State College, PA). Analysis of variance was conducted using a general linear model in which the donor was considered a random effect. To test the effect of cell harvest site or oxygen levels during differentiation, data from day 0 were excluded. The effect of cultivation time was tested for each differentiation condition separately including day 0 data. In addition, data sets from each individual time point were tested separately for statistical significance. If significant differences were detected between conditions, then Tukey's post hoc tests were performed. Statistically significant differences were considered to be present at p<0.05.
Results
Propagation of zonal chondrocytes
Population doubling times were not significantly different between chondrocytes from full-thickness (3.5±0.2 days), superficial zone (3.8±0.3 days), or middle/deep zones (4.1±0.04 days) during monolayer propagation (p=0.1). Chondrocytes from the different experimental groups had gone through similar population doublings before redifferentiation (full-thickness: 8.1±0.7; superficial 7.5±1.7; middle/deep: 6.9±1.0; p=0.1).
Expression of HIF-1α and HIF-2α and redifferentiation of zonal chondrocytes
The effect of different oxygen tensions was assessed by immunofluorescence staining for HIF-1α and HIF-2α. Immuno-reactivity for both HIFs appeared to be weaker and more diffuse in sections from normoxia samples compared with those from hypoxia samples; here shown for full-thickness chondrocytes (Fig. 1). Staining was observed to be localized in the cytosol of the cells cultured under ambient oxygen. In contrast, in chondrocytes differentiated under 5% O2, most intense staining was peripheral to their nuclei. Under low oxygen, immuno-reactivity for both HIFs was found to be more strongly associated with the nuclei in middle/deep cells, compared with full-thickness and superficial zone chondrocytes. Staining intensity for HIF-1α was highest near the center of the pellets cultured under hypoxia.
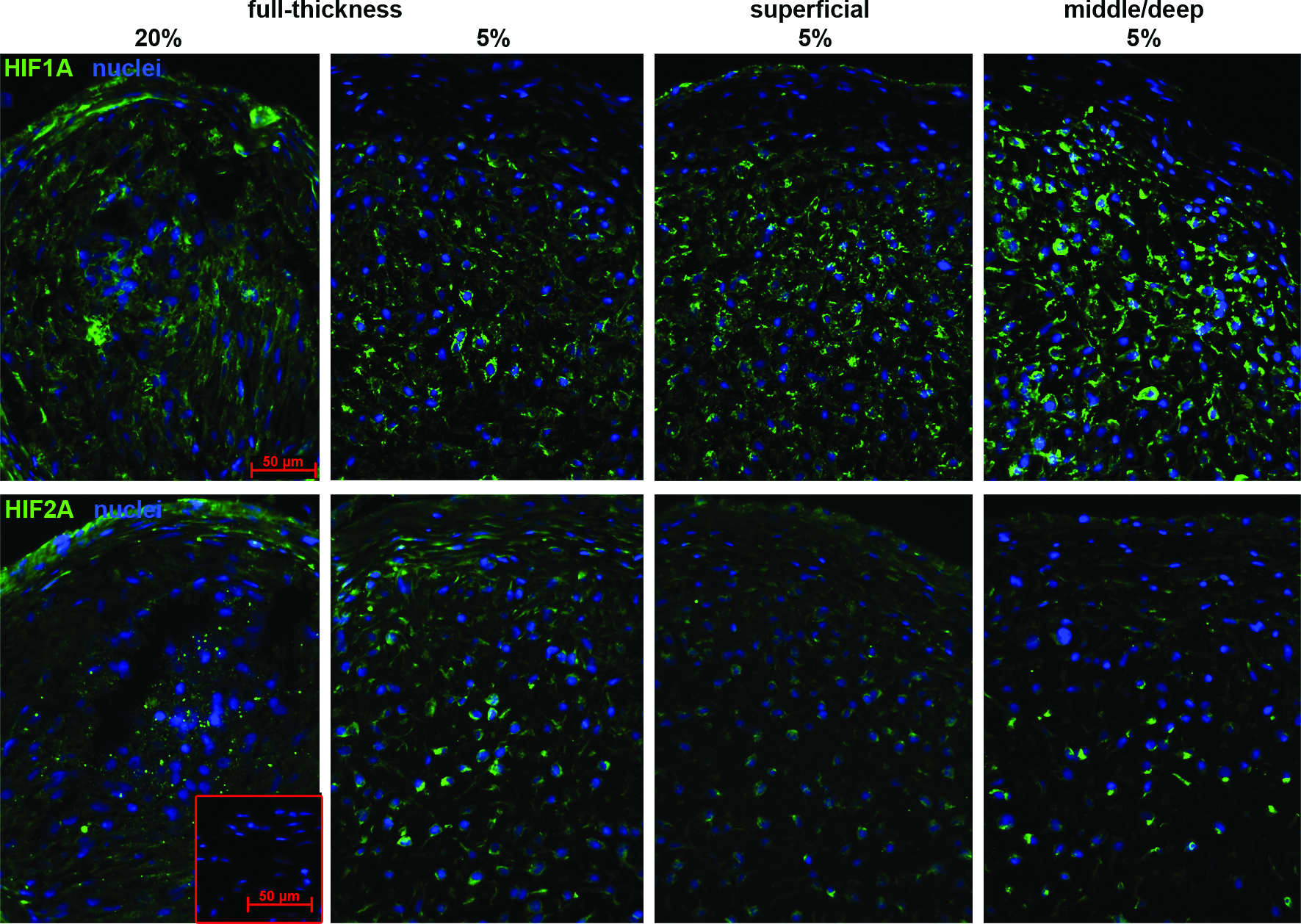
Immunofluorescence for HIF-1α or HIF-2α in pellet cultures of zonal chondrocytes redifferentiated under normoxic (20% O2) or hypoxic (5% O2) conditions. After expansion, micromass pellets of cells (2.5×105) from full-thickness, superficial, and middle/deep zone cartilage were cultured under hypoxic or normoxic conditions, respectively, in serum-free chondrogenic media for up to 28 days. Samples from pellet cultures were processed for fresh-frozen tissue immunofluorescence at day 28. Sections were probed with antibodies specific for HIF-1α or HIF-2α (green) and counterstained with DAPI (blue). Control sections without primary antibody incubation were included (red-framed inset). The figure presents samples representative of the experiments with cells from three patients. DAPI, 4′,6-diamidino-2-phenylindole; HIF-1α, hypoxia-inducible factor 1alpha; HIF-2α, hypoxia-inducible factor 2alpha. Color images available online at www.liebertonline.com/tea
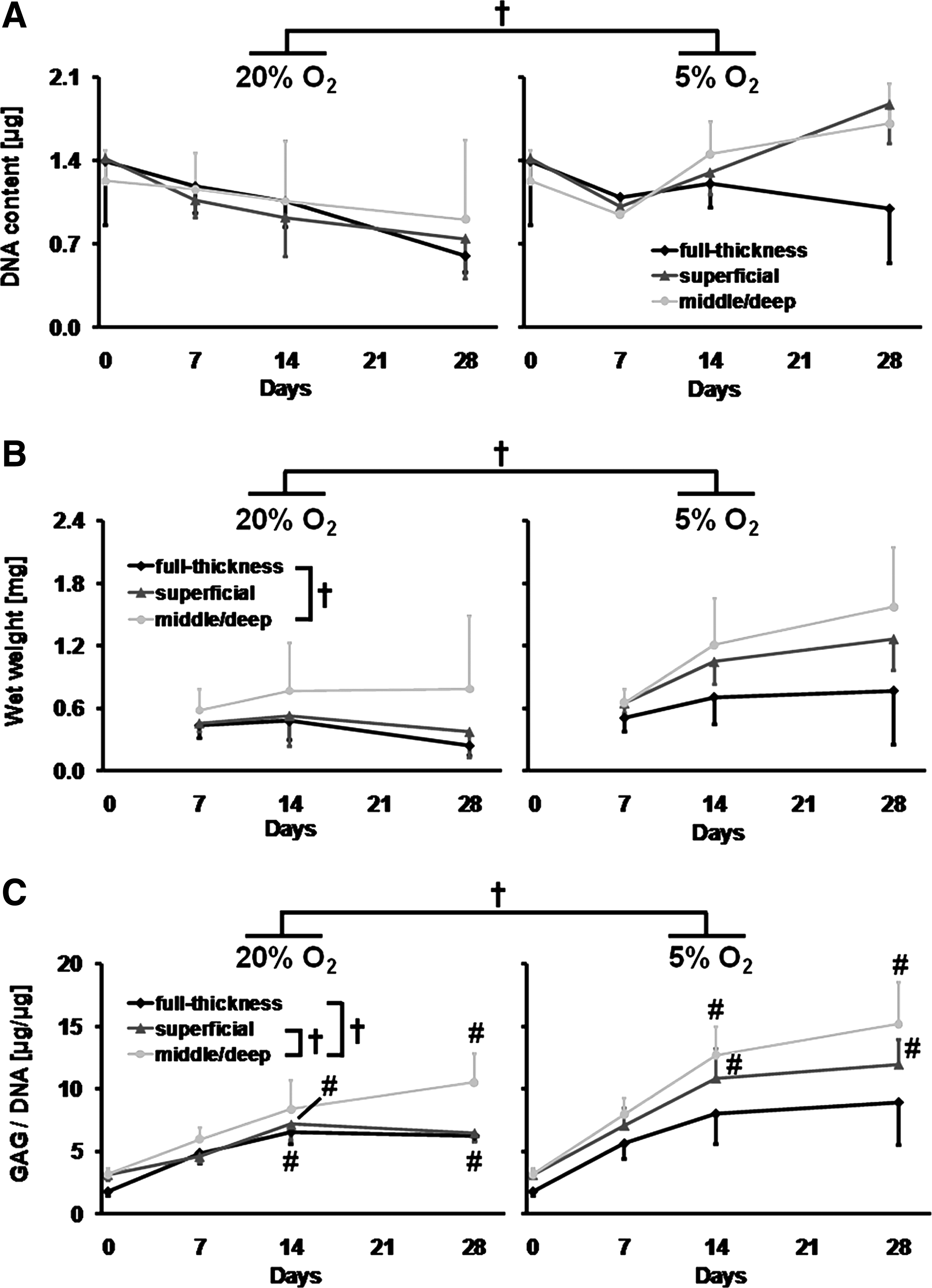
Wet weight, DNA, and GAG content was determined to compare the chondrogenic potential of zonal chondrocytes under various oxygen tensions. Oxygen tension during redifferentiation had a significant effect on pellet wet weight (p<0.001) and DNA content (p<0.01), as well as GAG content normalized to DNA (GAG per DNA; p<0.001; Fig. 2). There were no differences in DNA content (p=0.3) of pellets from the various harvest zones/sites (Fig. 2A). However, chondrocytes harvested from middle/deep zone cartilage showed overall higher pellet wet weights than chondrocytes from full-thickness cartilage (p<0.01). Moreover, middle/deep chondrocytes also produced overall more GAG per DNA compared with superficial (p<0.05) and full-thickness chondrocytes (p<0.001; Fig. 2B, C). DNA content and pellet wet weight remained stable over the differentiation period under normoxic and hypoxic conditions (Fig. 2A, B), yet there was a trend of increasing pellet wet weights under 5% oxygen between day 7 and 28 (Fig. 2B). Under 20% oxygen tension, higher levels of GAG per DNA were present in pellets from full-thickness and superficial chondrocytes at day 14 as well as in full-thickness and middle/deep chondrocytes cultures at day 28 compared with day 0 (p<0.05; Fig. 2C). Increased levels of GAG per DNA were also found under hypoxic conditions in pellets from middle/deep and superficial chondrocytes at day 14 and 28 compared with day 0 (p<0.05).
Expression of genes induced during dedifferentiation or chondrogenesis
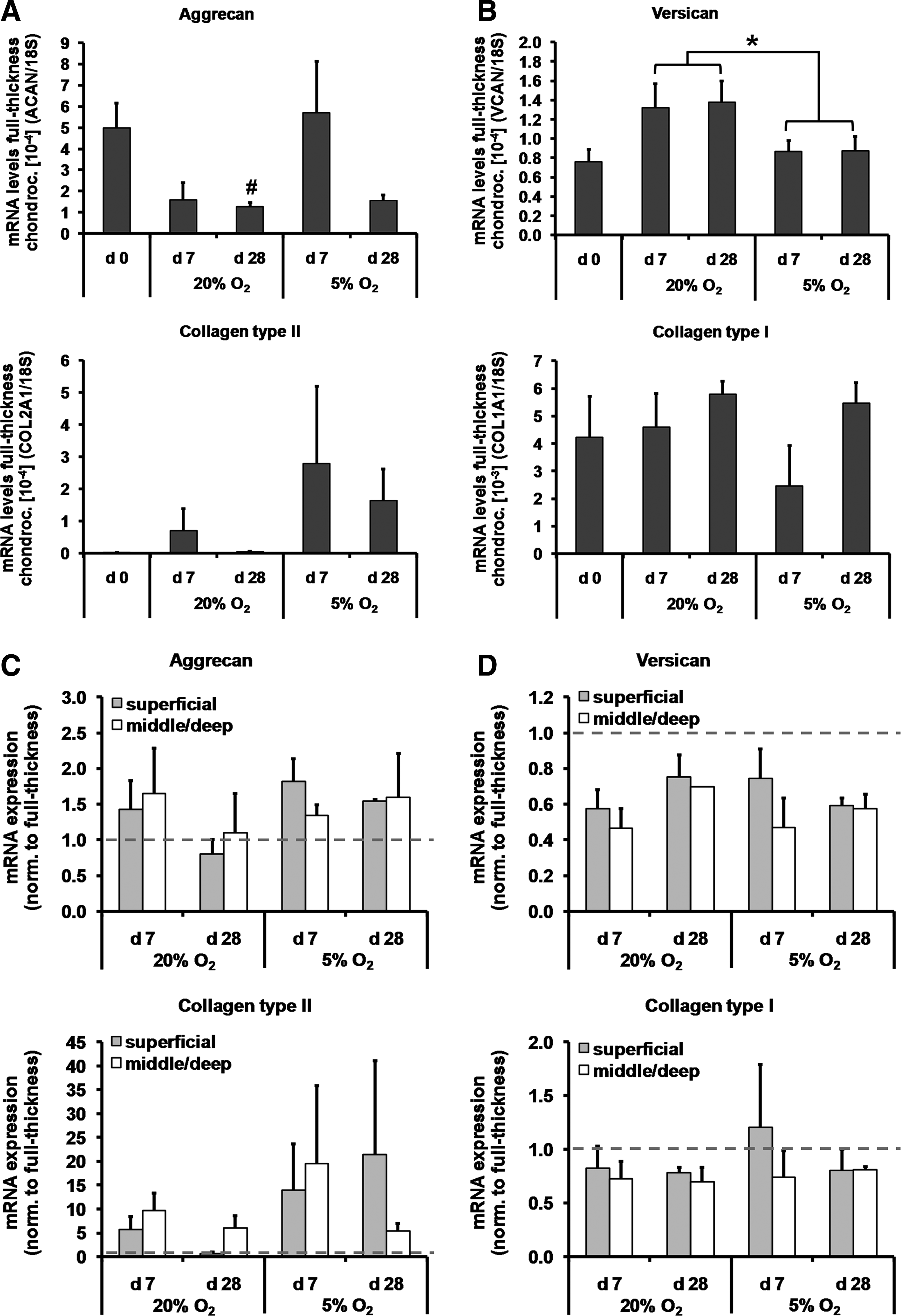
Expression of marker genes of chondrogenesis and dedifferentiation was assessed using qRT-PCR. Reduced oxygen tension tended to increase the overall mRNA expression of the chondrogenic markers aggrecan and collagen type II; however, this was not significant for full-thickness chondrocytes (p=0.1; Fig. 3A). At normoxic conditions, aggrecan expression was lower in full-thickness chondrocytes at day 28 compared with day 0 (p<0.01), but remained unaltered under low oxygen. Transcript levels of the dedifferentiation marker versican were higher at normoxic compared with hypoxic conditions (p<0.05; Fig. 3B). However, mRNA expression of collagen type I, another dedifferentiation marker, was not affected by the oxygen environment (p=0.2).
Expression of
To account for donor-related differences in chondrogenic potential of the samples, the mRNA data from superficial and middle/deep chondrocytes were normalized to mRNA levels from full-thickness chondrocytes. Regardless of oxygen tension and culture times, no differences in the mRNA expression levels of chondrogenic and dedifferentiation markers were detected between cells from the middle/deep and superficial zones (Fig. 3C, D).
Matrix synthesis during redifferentiation
The synthesis of cartilaginous matrix in pellet cultures was assessed by staining with safranin O (GAG) and by probing with antibodies specific for collagen type I and collagen type II. Pellets cultured under ambient oxygen exhibited weak staining for GAG and immunoreactivity for collagen type II, in contrast to pellets cultured under reduced oxygen (Fig. 4). Cells cultured at normoxic conditions, particularly from full-thickness and superficial samples, appeared to be more densely packed in the center of the pellets and were surrounded by minimal extracellular matrix, which immuno-reacted predominantly for collagen type I. At hypoxic conditions, the extracellular matrix of samples from middle/deep zone chondrocytes yielded the most intense immuno-reactivity staining for GAG and collagen type II. All pellets exhibited a superficial rim of multiple layers of flattened, elongated cells. The tissue in the outer layers stained weakly with safranin O and immuno-reacted nonuniformly for collagen type I. In general, samples from all conditions immuno-reacted positively for collagen type I. Interestingly, the matrix of osteoarthritic cartilage immuno-reacted not only for collagen type II but also for collagen type I, localized primarily to the superficial zone.

Histology and immunohistochemistry on pellet cultures of zonal chondrocytes redifferentiated under normoxic (20% O2) or hypoxic (5% O2) conditions. Samples from redifferentiation pellet cultures (full-thickness, superficial, and middle/deep zone cartilage) were processed for fresh-frozen tissue (immuno) histology at day 28. Sections were either stained with safranin O (red) and Fast Green or probed with specific antibodies against collagen type II (COLII, brown) or collagen type I (COLI, brown) and counterstained with hematoxylin. Sections from monensin-treated native cartilage (macroscopically normal site of osteoarthritic [OA] tissue) were included for comparison. The figure presents samples representative of the experiments with cells from three patients. Scale bars=50 μm. Color images available online at www.liebertonline.com/tea
Expression of zonal marker genes during redifferentiation
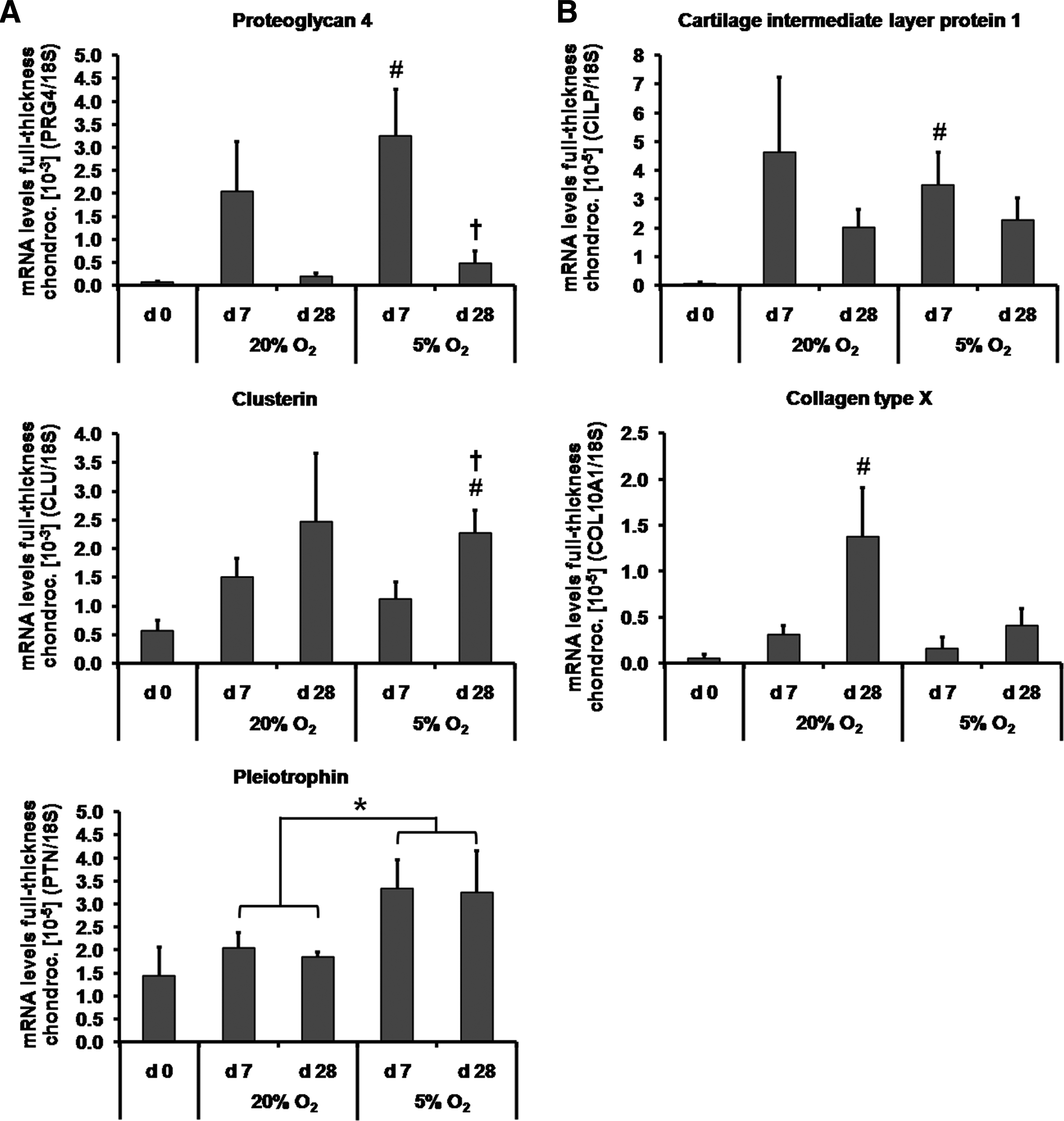
The expression of putative zone-specific genes of the superficial zone (PRG4, clusterin, PTN) was assessed by qRT-PCR. For full-thickness chondrocytes, PRG4 mRNA was undetectable after 4 weeks of monolayer propagation (Fig. 5A). It was re-expressed after 7 days of pellet culture at 5% oxygen tension (p<0.05). However, after 28 days, PRG4 expression decreased again (p<0.05). Clusterin levels were only significantly higher at day 28 compared with day 0 (p<0.01) and day 7 (p<0.05) under hypoxic conditions. PTN expression remained unchanged over the entire culture period. However, transcript levels of this molecule were overall higher at hypoxic compared with normoxic conditions (p<0.05).
Expression of genes of
PRG4 expression was higher in superficial chondrocyte pellets compared with that from middle/deep cells at day 7 (p<0.05) and at day 28 (p<0.01) under 5% oxygen tension (Fig. 5C). Expression levels for clusterin were also elevated in superficial chondrocytes compared with middle/deep cells under hypoxic conditions, however, only at day 7 (p<0.05). There was no difference in transcript levels between superficial and middle/deep cells for PTN at any time point or oxygen tension.
The middle zone marker CILP and deep zone marker COL10A1 were not detected at the start of the pellet culture (Fig. 5B). CILP was re-expressed after 7 days of full-thickness chondrocyte pellet culture at 5% oxygen tension (p<0.05). However, COL10A1 transcript levels increased only after 28 days under normoxic conditions compared with day 0 (p<0.05). Oxygen tension did not influence CILP (p=0.4) or COL10A1 (p=0.1) transcription. Irrespective of culture time, no differences in the mRNA expression levels of middle/deep markers were detected between cells from the middle/deep and superficial zones in response to different oxygen tensions (Fig. 5D).
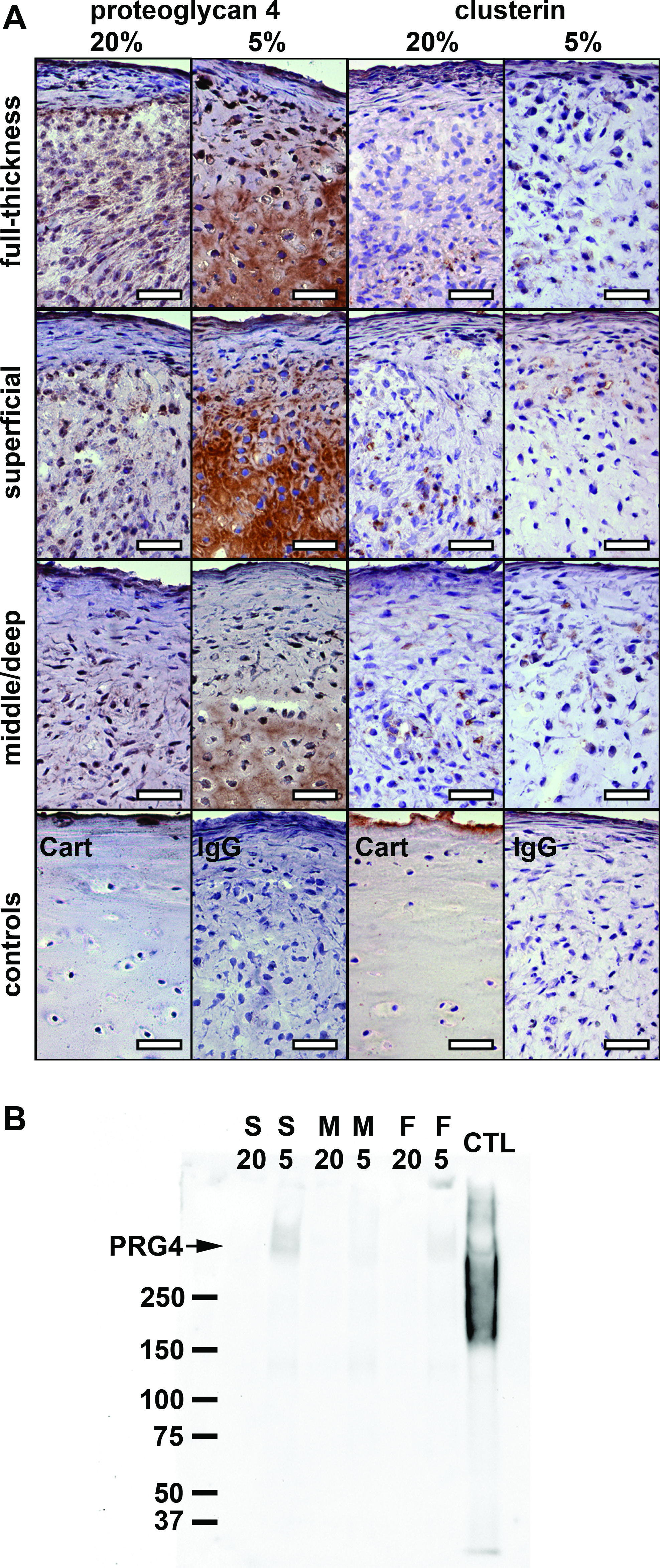
PRG4 and clusterin synthesis during redifferentiation
To confirm protein synthesis of PRG4 and clusterin, sections of pellets after 28 days in culture and conditioned media of week 4 of culture were analyzed by immunohistochemistry and western blotting, respectively (Fig. 6). Pellets redifferentiated at normoxic conditions exhibited only faint or no immuno-reactivity for PRG4 (Fig. 6A). In contrast, pellets cultured at hypoxic conditions with superficial cells exhibited intense immuno-reactivity for PRG4. Pellets from full-thickness and middle/deep chondrocytes cultured under hypoxia also immuno-reacted positively for PRG4, but less intensely than the pellets from superficial cells. Surprisingly, immuno-reactivity in these tissues appeared to be confined to the extracellular matrix near the center of the pellet. Conversely, PRG4 in monensin-treated osteoarthritic cartilage was localized to the outermost boundary of the superficial zone and to cells within this layer. Faint positive immuno-reactivity for clusterin was detected in all sections without clear differences between samples from the various oxygen conditions or tissue origins. Immuno-reactivity was confined to intra-/pericellular areas of cells scattered throughout the sections. Immunoblotting of conditioned pellet media revealed higher levels of PRG4 secreted by pellets derived from superficial chondrocytes under 5% oxygen in contrast to middle/deep or full-thickness cells. PRG4 was undetectable in all in media of cells cultured under ambient oxygen. (Fig. 6B). Recombinant lubricin appeared as a broad band around 250 kDA and showed a second narrow band below 37 kDa as well as an additional band in the higher molecular-weight range, which was described as the dimer of the 240 kDa lubricin monomer by others. 13
Discussion
The aim of this study was to evaluate the effect of oxygen tension on chondrocytes from the different zones (“normal” full-thickness, superficial, and middle/deep) of human articular cartilage. We observed that zonal chondrocytes retain phenotypic differences during in vitro cultivation. Of the putative zonal markers investigated here, only the expression of PRG4 appeared to be zone specific, which was favored by low oxygen tension. We generally found that moderate hypoxia enhanced the expression of HIFs along with the synthesis of extracellular matrix and chondrogenic marker proteins in all chondrocytes during in vitro redifferentiation. However, the positive effect of hypoxia on chondrogenesis was superseded by donor-specific limited redifferentiation capacity, as reported by others. 26 In agreement with several other studies, chondrocytes isolated from the middle/deep zone synthesized the most matrix compared with cells from other harvest sites.24,27–30 Immunostaining for HIF-1α and HIF-2α showed a clear oxygen-dependent expression pattern in all pellets at day 28 with both transcription factors only localized peripherally to the nuclei under hypoxic conditions. Moreover, HIF staining in these cultures appeared stronger toward the center of the pellets, which is consistent with decreasing oxygen concentrations as confirmed for cartilaginous tissue-engineered constructs elsewhere. 31
To characterize the phenotypic differences of the in vitro cultivated zonal chondrocytes in greater detail, we investigated the expression of proteins that are specifically synthesized in different zones of articular cartilage. Only chondrocytes in the superficial layer of normal cartilage express PRG4. 5 PRG4 expression is lost together with the chondrocyte phenotype during prolonged monolayer cultivation, as observed in our study and elsewhere. 32 However, PRG4 was re-expressed in pellet culture, particularly at reduced oxygen tension. Interestingly, in spite of equivalent or slightly reduced levels of HIF-1α and HIF-2α in superficial cells, they exhibited higher levels of PRG4 transcripts than middle/deep cells, which argues against a direct involvement of the HIFs and, hence, of hypoxia in the expression of this zonal marker. The presence of PRG4 in the middle/deep cells may be due to induction of this gene in cells that were originally PRG4-negative. 33 There is evidence that certain cytokines, such as TGF-β, interleukin 1α/β, and potentially insulin-like growth factor 1, modulate its metabolism.34–36 In our study, PRG4 transcript levels reflected the general expression trends of chondrogenic markers, particularly Collagen II with slightly decreased transcription after 4 weeks even when cultivated at low oxygen. This suggests that conditions which generally favor chondrogenesis influence PRG4 synthesis only in chondrocytes from the superficial zone, and that prolonged exposure to TGF-β and hypoxia is not sufficient to maintain either the expression of chondrogenic markers or PRG4 synthesis in vitro. Surprisingly, cells in the center of the pellets immuno-reacted more intensely for PRG4 than in the periphery. Since it is unlikely that oxygen and nutrient gradients in pellets act contrarily to the situation in vivo, they may be less important to PRG4 metabolism within pellets. This is also supported by the observation of PRG4-positive, ectopic chondrocyte aggregates in exposed subchondral bone of osteoarthritic human knees. 37 The gradient in immuno-reactivity intensity in the pellets could be the result of diffusion of PRG4 from outer layers of pellets into the surrounding medium, which unlike synovial fluid has a low PRG4 concentration. Testing the conditioned culture media for PRG4 protein by western blotting confirmed, indeed, higher levels of PRG4 secreted by pellets derived from superficial chondrocytes under 5% oxygen in contrast to middle/deep cells or the same cells cultured under ambient oxygen (Fig. 6B).
Another putative marker protein for the superficial zone of cartilage is clusterin.4,38 Clusterin is often found at fluid-tissue interfaces, suggesting a potential role in cell membrane protection from the damaging effects of various secretions. 39 Consequentially, it is confined to the superficial layer of cartilage.4,38 However, clusterin is up-regulated in early osteoarthritis, not only in the superficial layers. 40 In this study, clusterin synthesis appeared to increase over pellet culture time, and superficial chondrocytes expressed higher levels of this molecule at day 7 under 5% oxygen compared with middle/deep cells. However, this zone-specific difference was lost at day 28, as shown in mRNA and protein level. Elevated levels in middle/deep chondrocytes at later time points could be attributed to the osteoarthritic nature of the cells, but may also be related to the increasing stress, which cells certainly experience ex vivo in the serum-free pellet culture model. Interestingly, Schuurman et al. 30 found zone-specific synthesis of clusterin in equine articular chondrocytes cultured in alginate cultures but no zonal differences when using pellet cultures. Moreover, our use of TGF-β1 could have played a role in the up-regulation of clusterin, as this growth factor is known to induce clusterin expression in epithelial, endothelial, and fibroblast cell lines.41,42 Given that clusterin is involved in many cellular processes and that its zone-specific expression by chondrocytes appears to be dependent on the in vitro culture system, this protein might not be a suitable marker of zonal stratification in tissue-engineered cartilage.
Along with the above mentioned “classic” markers of the superficial zone of cartilage, we also determined the expression of pleiotrophin, an embryonic growth and differentiation factor, which is re-expressed in early osteoarthritis, mainly in the superficial zone. 6 Our data failed to indicate a zone-dependent expression of the molecule. However, we observed elevated transcription of pleiotrophin under moderate hypoxia, which is consistent with observations made in other cell types (e.g., ref. 43 ). A low oxygen environment could also indirectly affect pleiotrophin expression through the promotion of chondrogenesis. In fact, pleiotrophin itself can enhance chondrogenesis and proteoglycan synthesis by chondrogenic cells in vitro in a paracrine/autocrine manner. 44 On the other hand, in mouse growth plates, pleiotrophin is primarily localized in hypertrophic chondrocytes, pointing to its known role as an pro-angiogenic agent. 45 However, there is no apparent link between pleiotrophin expression and hypertrophy, when comparing the expression profiles of pleiotrophin and hypertrophy-associated collagen type X. In general, pleiotrophin appears more likely to be involved in repair and protection processes related to osteoarthritis, 6 where it reportedly reduces expression of catabolic factors. 46 Pleiotrophin's precise role in chondral ossification and the course of osteoarthritis requires further investigation.
CILP is a protein thought to be specifically expressed by middle and deep zone chondrocytes.7,47 It was proposed that CILP is involved in regulating extracellular levels of inorganic pyrophosphate and the deposition of calcium pyrophosphate dehydrate crystals in articular cartilage, which takes place predominantly in the middle/deep zone of aging tissue. 48 Although depth-dependent expression of CILP appeared to be conserved in cartilage affected by osteoarthritis, 49 we did not observe zone-specific CILP expression in our study. However, we did find that CILP expression was culture model-dependent, with major up-regulation during pellet culture. Indeed, there is evidence that CILP protein expression in chondrocytes is induced by TGF-β1.48,50 The increased expression of CILP in our model raises the question of whether calcium pyrophosphate dehydrate crystal deposition was also elevated, which would potentially have adverse effects on the quality of engineered cartilage.
In conclusion, in vitro cultured, dedifferentiated zonal chondrocytes re-express phenotypic differences during redifferentiation. Low oxygen tension favors re-expression of PRG4 in superficial cells, most likely through promoting overall chondrogenesis. This underlines the importance of considering zonal differences when creating native-like cartilaginous constructs. Of course, the zonal harvest of autologous chondrocytes poses some practical challenges that are still to be overcome. In this respect, zonal cartilage tissue engineering could be even more applicable to nonosteoarthritic joints where more tissue is available and zonal harvest is potentially easier. An additional challenge is that, dependent on the specific culture conditions, some putative zonal markers, especially those involved in various cellular processes, might not be expressed exclusively by specific subpopulations of chondrocytes ex vivo, and, thus, are not reliable predictors of in vitro stratification. Therefore, more robust zonal markers, in particular for middle/deep chondrocytes, are needed before we can attune in vitro culture conditions for zonal cartilage regeneration.
Footnotes
Acknowledgments
The authors would like to thank Prof. Dietmar Hutmacher (Queensland University of Technology [QUT]) for his expert opinion, Mr. Donald Geyer (QUT) for his assistance with the histology, as well as QUT and the Australian Research Council for funding the research described. Dr. Schrobback was supported by a PhD scholarship from QUT. Dr. Klein was supported by an Australian Postdoctoral Fellowship from the Australian Research Council, and Dr. Malda was supported by VENI Fellowship from the Dutch Technology Foundation STW, Applied Science Division of now, and the Technology Program of the Ministry of Economic Affairs. The antibody against collagen type II (II-II6B3), developed by T.F Linsenmayer, was obtained from the DSHB developed under the auspices of the NICHD and maintained by The University of Iowa, Department of Biology, Iowa City, IA 52242.
Disclosure Statement
The authors have no conflicts of interest.