
Abstract
In pediatric patients requiring vascular reconstruction, the development of a cell-based tissue-engineered vascular patch (TEVP) has great potential to overcome current issues with nonliving graft materials. Determining the optimal cell source is especially critical to TEVP success. In this study, we compared the ability of human aortic smooth muscle cells (HuAoSMCs) and human mesenchymal stem cells (hMSCs) to form cell sheets on thermoresponsive poly(N-isopropylacrylamide) (PIPAAm) substrates. hMSCs treated with transforming growth factor beta 1 (TGFβ1) and ascorbic acid (AA) had higher expression of SMC-specific proteins compared to HuAoSMCs. hMSCs also had larger cell area and grew to confluence more quickly on PIPAAm than did HuAoSMCs. hMSCs typically formed cell sheets in 2–3 weeks and had greater wet tissue weight and collagen content compared with HuAoSMC sheets, which generally required growth for up to 5 weeks. Assays for calcification and alkaline phosphatase activity revealed that the osteogenic potential of TGFβ1+AA-treated hMSCs was low; however, Alcian Blue staining suggested high chondrogenic behavior of TGFβ1+AA-treated hMSCs. Although hMSCs are promising for cell-based TEVPs in their ability to form robust tissue with significant extracellular matrix content, improved control over hMSC behavior will be required for long-term TEVP success.
Introduction
A cell-based tissue-engineered vascular patch (TEVP) may overcome current challenges with using synthetic materials or decellularized tissues in pediatric patients. Cell sheet engineering avoids the use of scaffolds and relies on cells for the synthesis and remodeling of extracellular matrix (ECM), resulting in a completely biological, living tissue. One of the earliest successes of cell sheet-based vascular grafts involved the use of vascular smooth muscle cell (VSMC) and fibroblast sheets wrapped around a mandrel; after maturation, the grafts were seeded with endothelial cells. 6 This approach has demonstrated clinical success in adults, 7 but currently would be challenging to apply in pediatric patients. A major drawback is the time required to mature the cell sheet graft, reaching up to 6 months or longer.6,8,9 Additionally, this method requires peeling the tissue from the culture substrate surface, 6 which may lead to mechanical trauma, tissue damage, and/or injury responses in the cells. An alternative approach using the thermoresponsive polymer poly(N-isopropylacrylamide) (PIPAAm) as a smart biomaterial substrate allows for noninvasive harvest of cell sheets with a temperature change. 10 PIPAAm has been applied to the development of a variety of engineered tissues, including skin, 11 heart,12–14 cornea, 15 and liver. 16 Cell sheets can be grown in a matter of days and then stacked in multiple layers13,17 or rolled into tubes 18 to form three-dimensional, completely biological tissue constructs. Although it has yet to be applied to TEVPs, PIPAAm is a promising solution to the issues of lengthy culture times and invasive harvest encountered with other cell sheet methods.
In the cell sheet engineering approach, optimal cell source and ECM characteristics are critical to the successful development of a TEVP. VSMCs have been considered as a natural first choice in recapitulating the mechanical strength and vasoactive function of the native media in tissue-engineered vessels. However, VSMCs may not be the best cell source option for TEVPs due to poor ECM deposition and proliferation characteristics; alternative cell sources should therefore be explored. Recently, mesenchymal stem cells (MSCs) have gained much attention in the field of tissue engineering.3,19–26 MSCs can be harvested from bone marrow using a simple aspiration technique27,28 that is less invasive and leads to less morbidity than the vessel harvest 29 required for isolation of VSMCs. MSCs have a high proliferative capacity28,30 compared to VSMCs; thus, potentially more tissue can be made with MSCs in a shorter period. MSCs are multipotent and capable of differentiating toward a variety of cell lineages,20,30–39 including VSMCs.40–43 In this study, we investigated primary human aortic SMCs (HuAoSMCs) and human MSCs (hMSCs) as potential cell sources for cell sheet-based TEVPs using PIPAAm substrates; we have previously used these cell types in patterned cell sheets grown on PIPAAm.44,45 As our interest is to mimic the medial layer of the artery, which contains VSMCs, we used transforming growth factor beta 1 (TGFβ1) and ascorbic acid (AA) to induce SMC-specific protein expression in MSCs.40,42 HuAoSMCs and TGFβ1+AA-treated hMSCs were cultured in their respective optimal growth media on PIPAAm; cell sheet growth, ECM synthesis, and cell differentiation were assayed.
Materials and Methods
Cell culture and treatments
Primary HuAoSMCs and primary hMSCs were purchased from Lonza (Basel, Switzerland). Cells were from healthy young adults (male and female, ages 22–33) and at least three different age-matched donors for both cell types were used in these studies. Cells were cultured in a humidified incubator with 5% CO2 at 37°C. Note that different growth media were used for HuAoSMCs versus hMSCs because preliminary studies that attempted to culture HuAoSMCs in the same growth medium as hMSCs resulted in poor HuAoSMC proliferation. Thus, each cell type was maintained in its optimal growth medium for standard culture and experiments. HuAoSMCs were cultured in smooth muscle cell growth medium (SmGM) consisting of smooth muscle cell basal medium (Clonetics, Walkersville, MD) supplemented with growth factors (SmGM-2 SingleQuots; Clonetics), 5% fetal bovine serum (FBS; Clonetics), and 1% penicillin–streptomycin (Invitrogen, Carlsbad, CA). Primary hMSCs were cultured in low-glucose Dulbecco's modified Eagle's medium (DMEM; Invitrogen) supplemented with 10% FBS (Invitrogen), 1× antibiotic–antimycotic solution (Invitrogen), and 5 mM HEPES (Sigma, St. Louis, MO) and passaged at 80% confluence to maintain multipotency. For all experiments, both cell types were used between passages 5 and 8.
To determine the effects of TGFβ1+AA treatment on hMSC differentiation, hMSCs and HuAoSMCs were seeded into six-well tissue culture polystyrene (TCPS) plates (Corning, Lowell, MA). hMSCs received 5% FBS-containing DMEM supplemented with 1 ng/mL TGFβ1 (R & D Systems, Minneapolis, MN) and 5 μg/mL AA (Sigma) at every feeding for 2 weeks. Nontreated HuAoSMCs cultured with SmGM and undifferentiated (nontreated) hMSCs cultured in 5% FBS-containing DMEM served as controls. For cell sheet studies, cells were cultured on 60-mm-diameter PIPAAm-grafted TCPS dishes (Cell Seed, Tokyo, Japan). hMSCs received TGFβ1+AA, while HuAoSMC sheets received 50 μg/mL AA at every feeding in their respective growth media to promote collagen synthesis. 46 At various time points, samples were assayed, as described below.
Immunofluorescent staining for smooth muscle cell marker proteins
Samples were fixed in cold 4% paraformaldehyde (Electron Microscopy Sciences, Hatfield, PA) for 15 min and washed three times with phosphate-buffered saline (PBS; Invitrogen). Samples were then permeabilized with 0.5% Triton X-100 (Sigma) for 10 min at room temperature (RT) and again washed three times with PBS. Nonspecific binding was blocked with 0.1% bovine serum albumin (BSA; Sigma) and goat serum (Invitrogen) dissolved in PBS for 1 h at 37°C. Next, samples were incubated with monoclonal mouse primary antibodies against smooth muscle myosin heavy chain (SM-MHC; clone SMMS-1; Dako, Glostrup, Denmark), α-actin (clone 1A4; Sigma), basic calponin (clone hCP; Sigma), or SM22α (clone 1B8; Abcam, Cambridge, MA) at 1:200 dilution for 2 h at 37°C. Samples were rinsed three times with 0.1% BSA solution and then incubated with Alexa Fluor 488–conjugated goat anti-mouse IgG (Invitrogen) at 1:200 dilution and Hoechst 33342 (Invitrogen) for 1 h at RT. Samples were washed three times with 0.1% BSA, covered with PBS, and imaged. Phase-contrast and fluorescent images were acquired using an Axiovert S-100 microscope system (Carl Zeiss, Inc., Thornwood, NY) equipped with Metamorph imaging software (Molecular Devices, Sunnyvale, CA).
Western blot for smooth muscle cell marker proteins
Cells were collected in cold lysis buffer (20 mM Tris, 5 mM EDTA, 1% Triton X-100, 150 mM NaCl, and 5 mM EETA, pH 7.4) with 1 mM phenylmethanesulfonyl fluoride (Sigma), 0.2 mM Na3VO4, and protease inhibitor cocktail (Sigma) at 1:500 dilution added fresh before use. Samples were vortexed, incubated for 5 min on ice, and spun at 15,000 rpm for 15 min at 4°C. The supernatant from each sample was stored at −20°C until use. Total protein was determined using a bicinchoninic acid assay (Pierce, Rockford, IL) with each sample run in triplicate. Equal amounts of protein for each sample were separated by SDS-PAGE using 7.5% polyacrylamide gels. Proteins were transferred to polyvinylidene difluoride membranes (Millipore, Billerica, MA) and blocked with 5% Carnation nonfat dry milk dissolved in Tris buffered saline with Tween 20 (TBST) for 1 h at RT. The same primary antibodies described above, as well as mouse monoclonal β-actin antibody (Abcam) for a load control, were incubated with membranes using optimized dilutions at 4°C overnight. After washing with TBST, membranes were incubated with optimized dilutions of horse radish peroxidase-conjugated goat anti-mouse IgG (Zymed, S. San Francisco, CA) for 30 min at RT. The membranes were again washed, and then reacted with SuperSignal West Pico Chemiluminescent Substrate (Pierce) for 5 min. Signals were captured on film (GE Life Sciences, Piscataway, NJ), developed, and analyzed using ImageJ's gel plug-in (1.37v; NIH, Bethesda, MD). The raw data were normalized such that HuAoSMC expression levels were adjusted to unity; that is, hMSC expression levels represent relative change compared to HuAoSMCs.
Cell morphology and growth to confluence on PIPAAm substrates
HuAoSMCs and hMSCs were seeded at 200,000 cells per PIPAAm or TCPS dish (60 mm diameter) in their respective growth media. After 24 h, samples were imaged and cell outlines were traced to determine projected cell area (A) and perimeter (P) using ImageJ. Cell shape index (CSI) was calculated according to the following formula: CSI=4πA/P 2 ; as CSI→0, cells become infinitely elongated and as CSI→1, cells become perfectly circular. 47 To determine growth over time in culture, samples were imaged at 1, 7, and 14 days. Confluence was determined as the total cell area measured in an image divided by the total image area multiplied by 100%. At least five images of random fields were used per substrate per time point for analysis.
Cell sheet detachment and measurement of wet and dry weight
Once HuAoSMC and hMSC sheets were ready for harvest (defined as the point in culture when cell sheets began to spontaneously pull away from PIPAAm dish edges), samples were detached from PIPAAm with temperature change from 37°C to RT (∼22°C) in physiological salt solution (119 mM NaCl, 4.7 mM KCl, 1.2 mM KH2PO4, 25 mM NaHCO3, 1.2 mM MgSO4, 2.5 mM CaCl2, 10 mM glucose, and 10 mM HEPES, pH 7.4). Before and after detachment, macroscopic images of cell sheets were acquired using a Canon Powershot A2000 digital camera. Detachment characteristics were qualitatively noted. After harvest, cell sheet wet weight was measured. To determine dry weight, harvested HuAoSMC and hMSC sheets were washed with diH2O, transferred to microcentrifuge tubes, and frozen in diH2O at −80°C until use (≤2 weeks). Frozen sheets were lyophilized using a VirTis Benchtop 4k Series Freeze Dryer (Model BT4KZL-105) for 4.5 h at −105°C. Dry weight was then determined on a microbalance.
Collagen and elastin assays
Collagen and elastin content in cell sheets were determined using the Biocolor Sircol™ collagen and Fastin™ elastin assay kits (Accurate Chemical, Westbury, NY) according to the manufacturer's protocols. Absorbance was read at 540 and 513 nm for collagen and elastin, respectively. Raw data were converted to μg of collagen or elastin per mg of wet tissue weight based on standard curves generated from collagen and elastin samples of known amounts.
Rabbit aorta tissue was analyzed in order to compare cell sheet matrix content to that of a native artery. All procedures were performed in accordance with the Institutional Animal Care and Use Committee at Boston University and the NIH Guide for the Care and Use of Laboratory Animals. Healthy male New Zealand white rabbits (2.5 kg; Pine Acres Rabbitry, Brattleboro, VT) were euthanized with an overdose of sodium pentobarbital (>120 mg/kg) administered intravenously to the ear. Left and right common carotid arteries were harvested using sterile tools. Vessels were immediately placed in cold Hank's balanced salt solution (HBSS: 137 mM NaCl, 5.4 mM KCl, 0.42 mM Na2HPO4, 0.44 mM KH2PO4, 4.17 mM NaHCO3, 10 mM HEPES, 5.55 mM glucose; 1% penicillin–streptomycin; pH 7.4) and stored on ice until use (<1 h). Vessels were cleaned of excess tissue, and then assayed for collagen and elastin, as described above.
Osteogenic and chondrogenic differentiation assays
Osteogenic behavior of hMSCs and HuAoSMCs was assayed using Alizarin Red staining and alkaline phosphatase (ALP) activity. Alizarin Red staining and quantification was performed using the Osteogenesis Assay Kit (Millipore) according to the manufacturer's instructions. ALP activity was determined using the Sensolyte pNPP Alkaline Phosphatase Assay Kit (AnaSpec, Fremont, CA) according to the manufacturer's instructions. For these assays, hMSCs cultured with osteogenic differentiation medium (Basal Medium+Osteogenic SingleQuot Kit; Lonza) were used as positive controls. Osteoblast-differentiated hMSC data were normalized to 100% (i.e., assumed to be fully differentiated toward the osteoblast lineage), and all other conditions were expressed as a percentage of the osteoblast-differentiated hMSCs.
Alcian Blue was used to stain glycosaminoglycans as a measure of chondrogenesis using the Chondrogenesis Assay Kit (Biomedical Research Service Center, Buffalo, NY) according to the manufacturer's instructions. hMSCs cultured with StemPro Chondrogenesis Differentiation Medium (Basal Medium+Chondrogenesis Supplement; Invitrogen) were used as positive controls. Chondrocyte-differentiated hMSC data were normalized to 100% and all other conditions were expressed as a percentage of the chondrocyte-differentiated hMSCs.
Statistical analysis
In the case where only two conditions were compared, the two-tailed Student's t-test was used to determine statistical significance. For comparison of three or more groups, statistical significance was determined using one-way analysis of variance and Tukey's post hoc test. A significant difference was considered to exist for p-values less than 0.05.
Results
SMC protein expression in TGFβ1+AA-treated hMSCs
TGFβ1 stimulates increased expression of SMC contractile proteins in cultured SMCs and other cell types.42,48–51 To determine whether hMSCs could be induced to express SMC-specific proteins, hMSCs were treated with TGFβ1+AA at every feeding for 2 weeks. Immunofluorescent staining for the SMC marker proteins α-actin, basic calponin, and SM22α showed that TGFβ1+AA treatment led to apparent upregulation in hMSCs (Fig. 1), as others have found.40,42,52 To determine whether SMC proteins were significantly higher in TGFβ1+AA-treated hMSCs compared to nontreated hMSCs and HuAoSMCs, protein expression was quantified by Western blot. Both α-actin and SM22α were consistently significantly upregulated in TGFβ1+AA-treated hMSCs compared to undifferentiated (nontreated) hMSCs and HuAoSMCs; the effect on basic calponin was more variable and did not always result in a significant difference (Fig. 2). We also investigated SM-MHC expression; however, we did not detect SM-MHC by immunofluorescent staining or Western blot in any condition (data not shown).

TGFβ1+AA-treated hMSCs express SMC marker proteins. Phase-contrast and fluorescent images of HuAoSMCs, undifferentiated (nontreated) hMSCs, and TGFβ1+AA-treated hMSCs stained for SMC-specific proteins after 2 weeks culture.
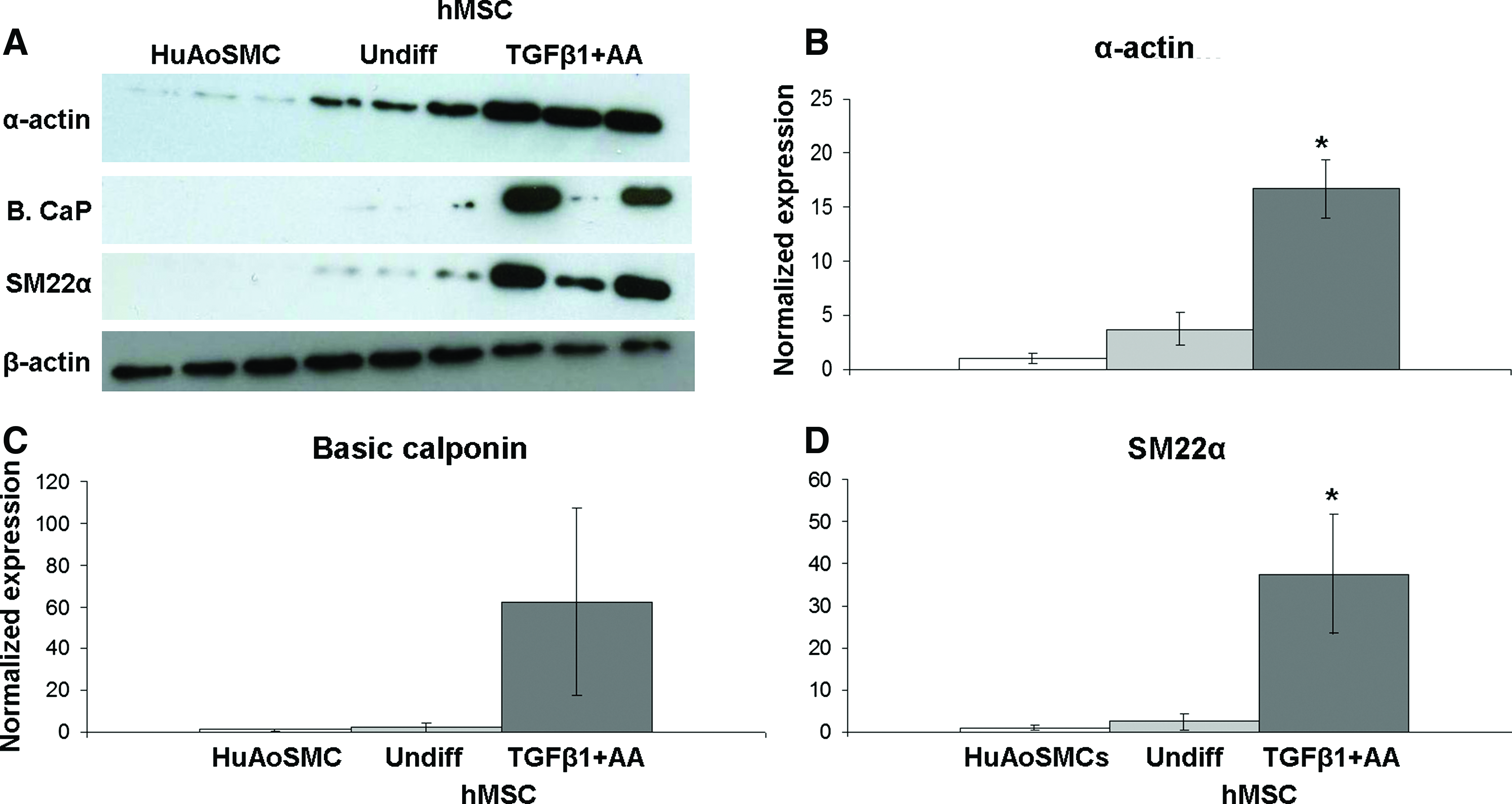
TGFβ1+AA-treated hMSCs express significantly more α-actin and SM22α compared to nontreated hMSCs and HuAoSMCs.
Projected cell area and growth on PIPAAm
Cell morphology and growth on PIPAAm substrates were compared between HuAoSMCs and hMSCs at several time points. At 24 h, hMSCs had significantly larger cell area and more spread cell shape on PIPAAm and TCPS substrates compared to HuAoSMCs (Fig. 3). Cell area and CSI were not statistically different between HuAoSMCs cultured on TCPS vs. PIPAAm substrates; the same was true for hMSCs. Thus, PIPAAm did not initially affect hMSC or HuAoSMC adhesion and spreading compared to standard TCPS substrates.

hMSCs are larger and more spread on PIPAAm compared to HuAoSMCs. Phase-contrast images showing
We assessed growth of HuAoSMCs and hMSCs on PIPAAm substrates compared to TCPS over a 2-week period. All substrates were seeded with 200,000 cells and imaged to assess changes in confluence. At 1 day in culture, confluence was greater for hMSCs compared to HuAoSMCs; this was likely because hMSCs tended to have larger cell area than HuAoSMCs, as described above. By 14 days, hMSCs on both PIPAAm and TCPS substrates, as well as HuAoSMCs on TCPS, were 100% confluent; however, HuAoSMCs on PIPAAm reached only 75%–80% confluence (Fig. 4).

hMSCs have greater proliferative capacity than HuAoSMCs on PIPAAm.
Tissue weight and ECM content of cell sheets
We allowed hMSCs and HuAoSMCs to form cell sheets on PIPAAm substrates until ready for harvest and analysis. The cell sheet formation process was considered complete when the tissue began to spontaneously separate from the PIPAAm dish edge; in general, this took 2–3 weeks for hMSCs and 3–5 weeks or more for HuAoSMCs. hMSCs received TGFβ1+AA, while HuAoSMCs were given 50 μg/mL AA at every feeding to boost collagen production46,53,54 and thus improve cell sheet integrity. We used 50 μg/mL AA for HuAoSMCs because it has been used previously in VSMC sheet production. 6 Furthermore, our preliminary studies revealed that TGFβ1+AA treatment appeared to halt proliferation and induce death in HuAoSMCs, as others have found in cultured human aortic VSMCs,55,56 thus preventing formation of cell sheets (data not shown). After 5 weeks of culture, HuAoSMC sheets remained thin and barely discernable by eye on PIPAAm. On the other hand, hMSCs formed thicker sheets that were macroscopically visible and started to spontaneously detach in culture near PIPAAm dish edges within 2–3 weeks (Fig. 5A). When cell sheets were harvested from PIPAAm and weighed, hMSC sheets had significantly greater wet weight compared to HuAoSMC sheets, despite typically less time in culture (Fig. 5B). However, hMSC and HuAoSMC sheets had similar dry weights (Fig. 5C).
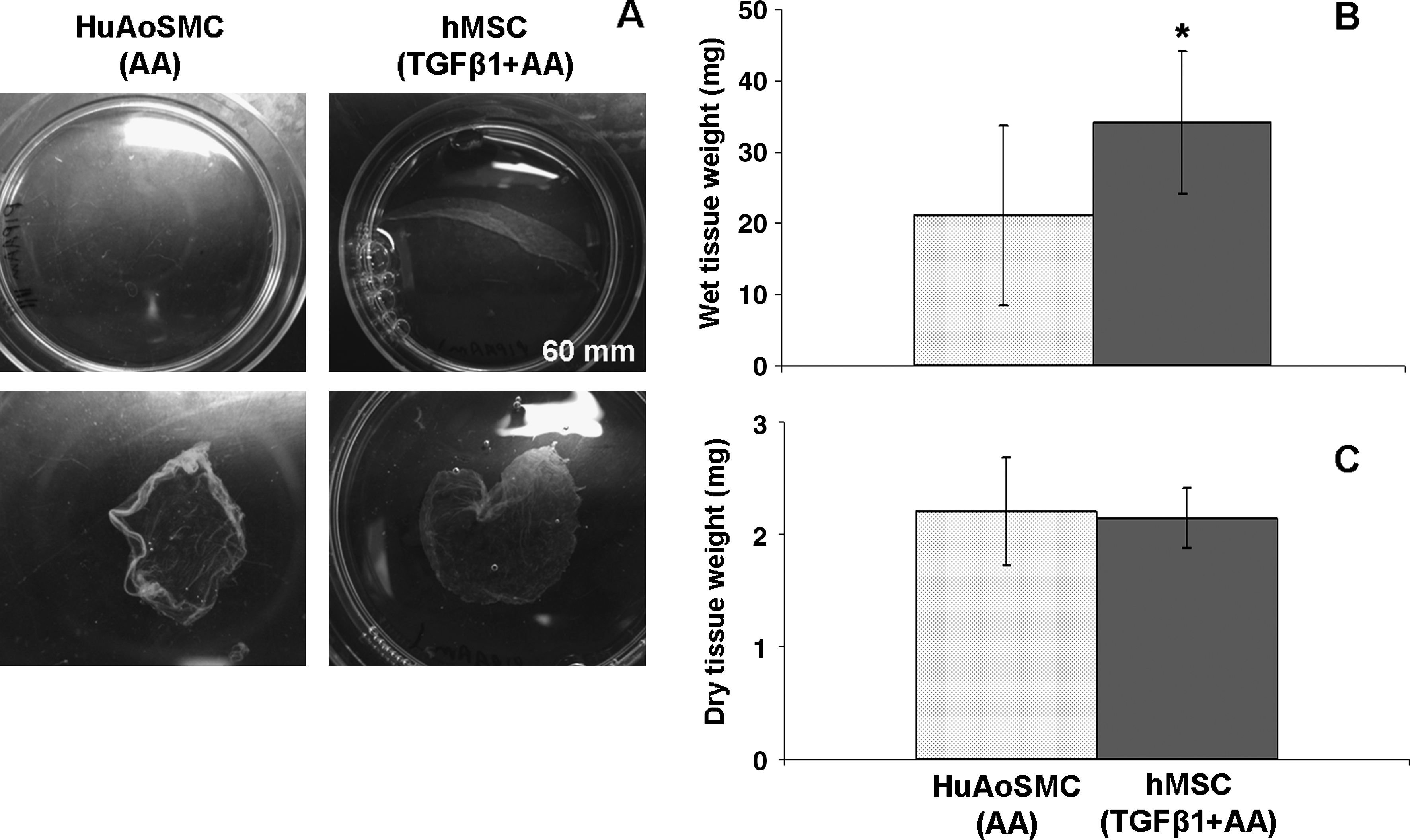
hMSCs form robust cell sheets with greater mass compared to HuAoSMCs.
Total collagen and elastin content was determined in native rabbit aorta, HuAoSMC, and hMSC sheets using colorimetric assays. Although HuAoSMCs received 10 times more AA than hMSCs, hMSC sheets contained twice as much collagen, with content similar to the native aorta (Fig. 6A). hMSC and HuAoSMC sheets had similar elastin content, but levels were significantly less compared to the native aorta (Fig. 6B). The average collagen:elastin ratios of the aorta, HuAoSMC sheets, and hMSC sheets were 0.4, 0.5, and 1.5, respectively. Although ECM composition was more similar between the native aorta and HuAoSMC sheets, TGFβ1+AA-treated hMSCs were able to form more robust sheets with greater wet weight and collagen content than HuAoSMCs.

hMSCs produce more collagen than HuAoSMCs in cell sheets. Total collagen and elastin content was determined in native rabbit aorta and cell sheets.
Osteogenic and chondrogenic potential of TGFβ1+AA-treated hMSCs
Because calcification is a major challenge in current TEVP approaches and MSCs differentiate readily toward the osteoblast lineage, we investigated the osteogenic potential of TGFβ1+AA-treated hMSCs. Alizarin Red staining was compared between TGFβ1+AA-treated hMSCs, undifferentiated hMSCs, and hMSCs cultured with osteogenic medium (positive control). Although TGFβ1+AA-treated hMSC samples had significantly more calcification than undifferentiated hMSCs and HuAoSMCs after 2 weeks of culture, calcification was low in comparison to osteoblast-differentiated hMSCs (Fig. 7A–C). ALP was also determined as a measure of osteoblast differentiation. TGFβ1+AA-treated hMSCs and HuAoSMCs had the lowest ALP activity among the 4 conditions (Fig. 7D). These results demonstrate that the osteogenic potential of TGFβ1+AA-treated hMSCs is relatively low.
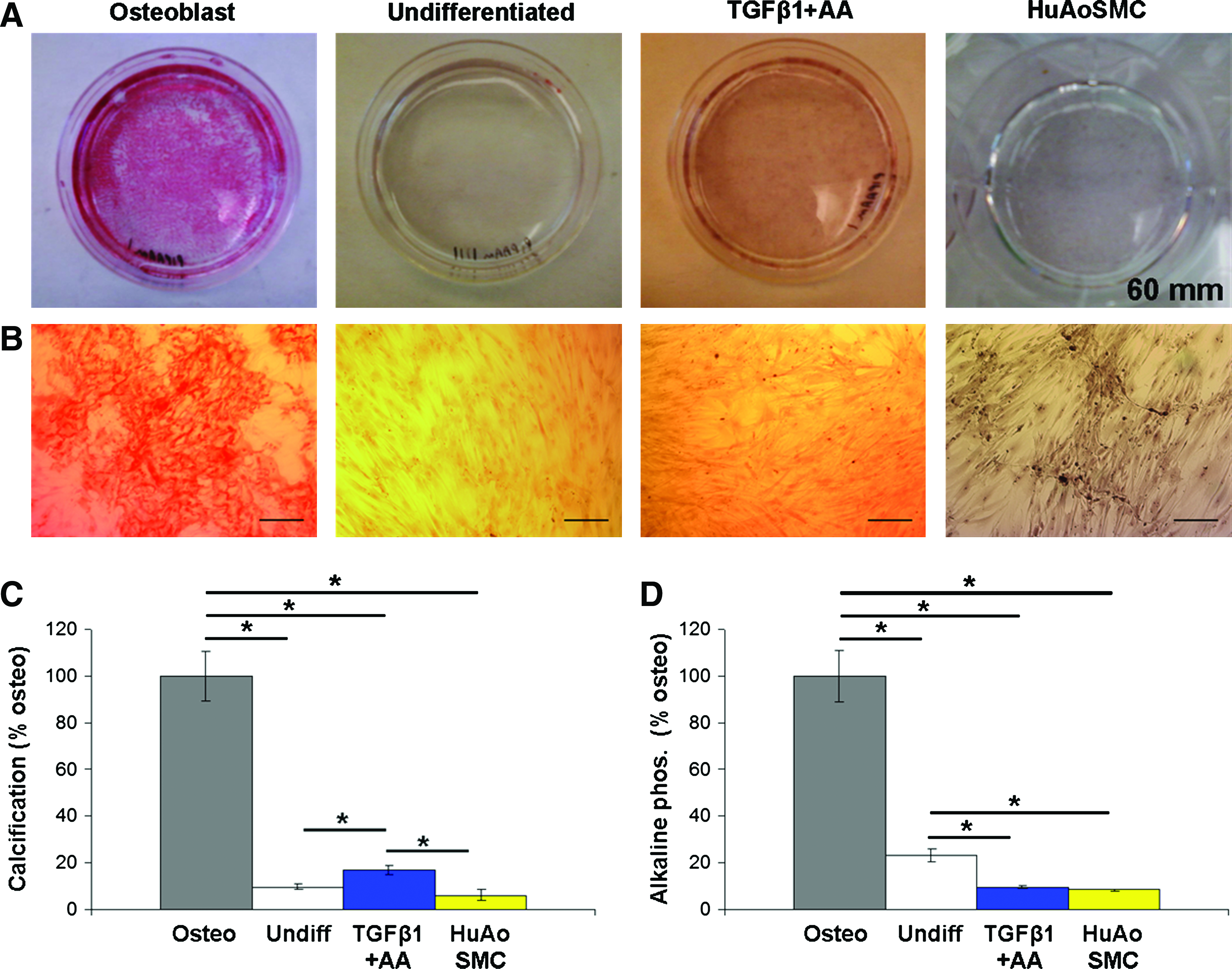
TGFβ1+AA-treated hMSCs have low osteogenic potential after 2 weeks of culture. Alizarin Red staining was used as a measure of calcification in hMSCs and HuAoSMCs. Macroscopic
In addition, we investigated chondrocyte differentiation of TGFβ1+AA-treated hMSCs using Alcian Blue staining. TGFβ1+AA-treated hMSCs had similar staining compared to chondrocyte-differentiated hMSCs. Both undifferentiated hMSCs and HuAoSMCs showed significantly less Alcian Blue staining (Fig. 8). These data suggest that although TGFβ1+AA-treated hMSCs express SMC-specific proteins and have low osteogenic potential at 2 weeks, their chondrogenic behavior may be high.

Chondrogenic potential of TGFβ1+AA-treated hMSCs is high.
Discussion
As methods for the scaffold-free PIPAAm approach to cell sheet fabrication have been refined, appropriate cell source and methods to optimize ECM production must be considered for specific tissue engineering applications. With regard to TEVPs, there are theoretical advantages to the use of VSMCs, but a practical comparison with other cell types has not yet been previously described. This study demonstrates that cell source significantly affects cell sheet properties that are critical for TEVP development; specifically, we found that the use of TGFβ1+AA-treatedhMSCs was associated with significantly higher SMC protein expression, improved cell sheet formation, and greater collagen deposition when compared to HuAoSMCs (refer to summary in Table 1).
Significant difference for p<0.05.
Cell proliferation, cell adhesion, and ECM deposition are critical for developing tissue strength in scaffold-free technologies, as structural integrity must ultimately derive from the resulting biological tissue. TGFβ1+AA-treated hMSCs formed more robust cell sheets with greater wet weight and twice as much collagen than HuAoSMCs with only 2–3 weeks of culture. More robust cell sheet formation using hMSCs can be attributed, at least partially, to the finding that HuAoSMC proliferation was limited on PIPAAm, while hMSC proliferation was not affected compared to standard culture on TCPS. It is also possible that HuAoSMC adhesion on PIPAAm was poor, and cells may have detached from the substrate over time; however, we did not investigate this further. Regardless, the decreased ability of HuAoSMCs to form a confluent layer on PIPAAm likely hinders their formation of intact, robust cell sheets. Another potential explanation for the difference in cell sheet formation is the difference in growth medium composition for the two cell types. We attempted to optimize the growth media for cell sheet formation for each cell type. Both media contained 5% FBS, and although different growth factors were present in each medium, ECM deposition—particularly, collagen synthesis—is likely due to AA treatment. 53 HuAoSMC sheets received 10 times as much AA as hMSC sheets, but contained only half the amount of collagen, suggesting an intrinsic difference in each cell type's ability to synthesize ECM. The quality and composition of the ECM produced by cells is critical to the mechanical properties of the tissue. Collagen content in hMSC sheets was comparable to native rabbit aorta, suggesting that hMSC sheets may have improved mechanical strength compared to HuAoSMC sheets. However, the collagen:elastin ratio in HuAoSMC sheets and rabbit aorta was more similar. Ideally, the ECM composition and mechanical properties of TEVPs should match the native vessel. Increasing elastin synthesis in hMSC sheets to obtain a collagen:elastin ratio similar to the native artery and assessing the mechanical strength of cell sheets will be of interest in future work.
Control of cell phenotype is another critical factor for successful TEVP development and growth potential. Our result that TGFβ1+AA-treated hMSCs demonstrated significantly higher expression of SMC-specific proteins compared to nontreated hMSCs agrees with previous reports. 42 Surprisingly, TGFβ1+AA-treated hMSCs also expressed more SMC-specific proteins compared to nontreated HuAoSMCs. We chose nontreated HuAoSMCs because we were interested in comparing the differentiation of treated hMSCs to that of VSMCs under standard culture. Indeed, cultured SMCs upregulate contractile proteins in response to TGFβ151; however, TGFβ1 also significantly inhibits SMC proliferation, 56 and could not be used in our cell sheet studies. HuAoSMCs expressed low amounts of α-actin, basic calponin, or SM22α, which is likely explained by the well-known phenomenon of phenotypic modulation. VSMCs are known to de-differentiate from their native contractile phenotype to a synthetic phenotype in culture, which is characterized by decreased expression of SMC-specific proteins associated with contraction. 57 Although the upregulation of SMC proteins in TGFβ1+AA-treated hMSCs was promising, the lack of SM-MHC, a strict marker of the differentiated contractile VSMC, 58 in addition to evidence of chondrogenic behavior via Alcian Blue staining suggests that TGFβ1+AA-treated hMSCs were not completely driven toward the SMC lineage. Factors that result in improved control over hMSC differentiation toward desired cell lineages must be determined in future studies; potential strategies include stimulation with different growth factors (in addition to or instead of TGFβ1+AA), coculture of MSCs with vascular cells, conditioning in bioreactors that mimic the native vessel's mechanical environment, and/or culturing hMSCs on ECM that mimics the native composition of the artery.
MSCs readily differentiate toward bone and cartilage,31,59–62 outcomes which must be avoided for long-term TEVP success. Calcification and ALP activity of TGFβ1+AA-treated hMSCs was relatively low, suggesting low osteogenic potential with TGFβ1+AA treatment. As calcification is a major issue in current materials used for vascular reconstruction in pediatric patients, our finding is desirable for TEVP development; however, studies beyond 2 weeks will be necessary in the future. While we found that TGFβ1+AA-treated hMSCs had significant Alcian Blue staining, Alcian Blue stains proteoglycans and does not indicate other specific measures of chondrogenesis; indeed, proteoglycans are also present in the artery. 63 High proteoglycan synthesis was also indicated by similar dry weights in hMSC and HuAoSMC sheets, despite hMSC sheets having higher wet weights, as proteoglycans can hold significant amounts of water in tissue. 64 The apparent chondrogenic behavior of TGFβ1+AA-treated hMSCs may only be a temporary result of high proteoglycan production, and thus not a long-term issue for TEVPs once implanted. We further note that although TGFβ1+AA treatment is useful for ECM synthesis by hMSCs, it may not be the optimal treatment for creating cell sheets for TEVPs. TGFβ1 has been used to induce chondrogenic differentiation in hMSCs63,64 and, as it is known to act on a variety of cells and tissues,49,51,65–73 TGFβ1 is likely too nonspecific on its own to induce controlled differentiation toward a single-cell lineage. hMSC behavior may require redirection with other factors once sufficient ECM deposition has occurred in cell sheets.
A limitation of our study was the need to grow HuAoSMCs and hMSCs in different culture media in order to form cell sheets (SmGM supplemented with 50 μg/mL AA vs. 5% FBS in DMEM supplemented with 1 ng/mL TGFβ1 and 5 μg/mL AA). However, different cell types generally require different culture media for optimal growth. Our interest was in comparing cell sheets of different cell types grown under their respective optimal conditions, as determined at the time, rather than investigate the effects of a particular growth medium on cell sheet formation. We initially tried culturing cells in the same growth medium (5% FBS in DMEM); however, HuAoSMC morphology changed and the cells did not grow well in DMEM (data not shown). Additionally, we attempted to culture HuAoSMC sheets with TGFβ1+AA. The cells appeared to stop proliferating and did not form a cell sheet, even after weeks of culture, as described in the Results section (data not shown). Different AA concentrations were used because they had been previously reported in the literature for MSCs and VSMCs.6,42 We acknowledge that we did not perform an exhaustive study to fully optimize cell sheet culture conditions, and indeed our findings suggest that further optimization will be necessary in the future for MSC or VSMC sheet-derived vascular constructs.
Balancing ECM production and cell differentiation is a delicate and complex process. Less differentiated cell types tend to deposit more ECM than their differentiated counterparts, such as the de-differentiated, synthetic VSMC versus the differentiated, contractile VSMC. 57 ECM deposition is important for strength and durability of engineered tissues, especially in scaffold-free cell-based approaches; however, inappropriate cell differentiation can lead to undesirable responses, such as inflammation, uncontrolled proliferation, stenosis, or calcification. The apparent chondrogenic behavior of TGFβ1+AA-treated hMSCs is not necessarily unfavorable in the short-term if it leads to synthesis of ECM, but this behavior will need to be tightly controlled to prevent ectopic cartilage formation in TEVPs. Differentiation of hMSCs must be studied in greater depth, including additional assays for cell differentiation and function and microarray analysis of gene expression. Such studies will provide insight into controlling hMSC behavior in future work, and a variety of techniques to controlling hMSC differentiation, such as gene delivery or developmental biology approaches, should be explored. We also note that hMSCs are one example of a potential cell source for TEVPs, and while our studies find them superior to HuAoSMCs for cell sheet development, hMSCs may not be the optimal cell source. Other cell sources should be identified and investigated for their potential use in cell sheet-based TEVP applications.
Conclusions
This study revealed that cell source is critical for the formation of cell sheets that can potentially be used to assemble TEVPs for pediatric patients with CHD. We found hMSCs to be superior to HuAoSMCs in forming cell sheets on PIPAAm substrates: hMSC sheets had greater wet tissue weight and contained more collagen with typically half the time in culture compared to HuAoSMC sheets. Thus, a major advantage to using hMSCs in cell-based approaches to tissue engineering is their ability to form significant amounts of robust tissue in a short period. However, controlling hMSC differentiation remains a challenge, as our studies revealed characteristics suggestive of both SMC and chondrocyte lineages in TGFβ1+AA-treated hMSCs. While hMSCs are still a promising cell source for TEVPs, an important future goal will be developing methods to better control hMSC differentiation toward specific cell lineages while maintaining the benefits of rapid tissue formation and ECM synthesis.
Footnotes
Acknowledgments
This work was supported by an American Heart Association predoctoral fellowship (C.W.), the BU ENG Scholars and BU Undergraduate Research Opportunity Program (A.W.X.), and The Hartwell Foundation (J.Y.W. and S.M.E.).
Disclosure Statement
Masayuki Yamato is a consultant for CellSeed, Inc. Teruo Okano is an investor in CellSeed, Inc and is an inventor on the patent for temperature-responsive tissue culture substrates.