
Abstract
Embryonic stem cells (ESCs) have the potential to differentiate into all tissues of the adult organism. This, along with the ability for unlimited self-renewal, positions these cells for regenerative medicine approaches based on tissue engineering strategies. With the objective of developing a treatment regime for skeletal injuries and diseases, this study presents a novel protocol that effectively induces ESC differentiation into osteogenic and chondrogenic lineages while concurrently eliminating observed tumorigenicity during the period of observation after transplantation in vivo. Exposure to a collagen I matrix polymerized with beta-glycerol phosphate (BGP) induced the osteogenic differentiation of the ESCs with an efficiency of >80% without purification and/or lineage-specific cell selection. Furthermore, when the collagen I matrix was supplemented with chondroitin sulfate, chondrogenesis was promoted instead of osteogenesis. Interestingly, without purification of the differentiated cells from the collagen I matrix, these constructs did not lead to the formation of teratomas or tumors when implanted subcutaneously in a severe combined immunodeficiency (SCID). Furthermore, if undifferentiated ESCs were mixed with collagen I and then injected immediately (i.e., without previous in vitro differentiation), again, no teratomas or tumors were observed, whereas undifferentiated ESCs without collagen scaffolds all produced teratomas in this bioassay system. These results suggest that collagen I scaffolds not only induce osteogenic differentiation of ESCs, but also prevent ESCs from producing unwanted tumors when injected in vivo.
Introduction
Adult bone marrow-derived mesenchymal stem cells (MSCs) are the most common source of primary cells for orthopedic tissue engineering. Although MSCs have high osteogenic and chondrogenic potential, they are isolated from the marrow in very low numbers, especially in older individuals.1–3 Furthermore, the osteogenic potential of MSCs diminishes with the long-term expansion of MSCs ex vivo, 4 which limits the numbers of cells available for clinical transplantation. Pluripotent stem cells are an alternative cells source for tissue engineering: they expand readily in culture and possess the capacity to generate any cell type. Traditionally, pluripotent embryonic stem cells (ESCs) have been derived from the early embryonic blastocyst; more recently, induced-pluripotent stem (iPS) cells 5 have been derived from adult tissues. Regardless of source, the differentiation of stem cells into a targeted lineage can be driven by exposure to certain soluble bioactive factors (i.e., growth factors, hormones, and oxygen), extracellular matrix (ECM) molecules, specialized cells (i.e., co-culture), and mechanical loading.
Until recently, the differentiation of ESCs into skeletal cell types has typically involved the use of bioactive factors. Specifically, various studies have reported that exposure to ascorbic acid, beta-glycerophosphate, dexamethasone, and vitamin D3 (VD3) can induce osteogenesis in ESC populations, as observed through the expression of lineage-specific genes, proteins, and production of specific ECM constituents.6–10 In a recent study by Evans et al., 11 the osteogenic differentiation of ESCs by humoral factors was enhanced in the presence of a tissue-specific ECM. 11 Collagen matrices have also been shown to have osteo-inductive effects on other types of stem cells, including bone marrow and umbilical cord-derived MSCs.12,13 In another study, the ability of ESCs to form tumors was reduced when loaded onto a collagen I gel before implantation into an osteochondral defect and exposed to natural loading of the joint in vivo. 14 The primary objectives of the current study were to evaluate the efficiency with which fibrillar collagen I matrix induces osteoblastic differentiation of ESCs in vitro and suppresses ESC tumors in vivo. The current study suggests that collagen I matrix alone can effectively be used to induce osteoblastic differentiation and reduce the tumorigenicity of mouse ESCs.
Methods and Materials
Mouse ESC culture
Murine D3 ESCs were maintained in the pluripotent state in high-glucose Dulbecco's modified Eagle's medium (DMEM) supplemented with 15% fetal bovine serum (FBS), 1% nonessential amino acids, 50 U/mL penicillin, 50 μg/mL streptomycin, 0.1 mM β-mercaptoethanol (all Invitrogen), and 1000 U/mL leukemia inhibitory factor. Pluripotent cultures were subcultured every second day on murine embryonic fibroblast feeder cells (MEFs), until one passage before differentiation when MEFs were removed and ESCs were placed on cultures prepared with 0.1% gelatin. D3 cells below passage 10 were used for all experiments. The karyotype of ESC cultures was checked every three to four passages; if a batch of cells fell below 85% diploid, it was discarded.
In vitro differentiation of ESCs
For differentiation protocols driven by VD3 and dexamethasone, embryoid bodies (EBs) were generated as reported previously.7,8,10 EBs formed in hanging drops were differentiated by adding specific medium supplements on the fifth day after EB formation. Specifically, osteoblast differentiation was induced with β-glycerophosphate (10 mM; Sigma), ascorbic acid (50 μg/mL; Sigma), and either 1,25-OH2 VD3 or Dexamethasone (5×10−8 M; Sigma) from day 5 to 30 as previously described.7,8,10
In vitro differentiation of cell-loaded collagen constructs
Bovine fibrillar collagen I (StemCell Technologies) was either polymerized as a two-dimensional (2D) film (∼1 mm in thickness) or 3D gel (∼10–15 mm in thickness) using 10 mM beta-glycerol phosphate (BGP). Specifically, 80% v/v 3 mg/mL-type collagen I solution was mixed with a single ESC suspension (1 million cells/mL, without a prior EB formation step) and 20% v/v BGP dissolved in either phosphate-buffered saline (PBS; for 2D films) or 5× concentrated DMEM (for 3D gels). The 5× concentrated DMEM was generated by dissolving powdered DMEM (Invitrogen) in 1/5 the recommended amount of water (200 mL instead of 1L) and supplemented with 15% FBS (Invitrogen), 1% nonessential amino acids, 50 U/mL penicillin, 50 μg/mL streptomycin, and 0.1 mM β-mercaptoethanol. Collagen I and chondroitin sulfate gels were formed by adding a 40% v/v solution of 1 mg/mL chondroitin sulfate (Sigma) to 40% v/v solution of 3 mg/mL collagen I and a 20% v/v solution of BGP dissolved in 5× concentrated DMEM.
Cell counting and viability assessment
Two-dimensional cell-collagen constructs were dissociated using 1 mg/mL collagenase at 37°C for 5 min. The cells were counted using a Bio-Rad TC-10 automated cell counter with trypan blue staining to quantify cell viability.
Immunofluorescence
Cell-collagen-BGP constructs were washed in PBS and fixed in 4% paraformaldehyde (PFA) in PBS at 4°C overnight. Constructs were then permeabilized in 0.5% saponin in PBS at 4°C overnight, rinsed once in PBS, and then blocked in 3% bovine serum albumin (BSA) again at 4°C overnight. Primary antibodies (osteopontin, alkaline phosphatase, osteonectin, and collagen II [Developmental Studies Hybridoma Bank] or osteocalcin, osterix, and aggrecan [Santa Cruz]) were diluted 1:50 in 3% BSA, added to the cell samples, and incubated overnight at 4°C. The constructs were then washed three times with PBS and blocked again overnight at 4°C. Following the block, the constructs were incubated with an appropriate Alexa-fluor 488 secondary antibody (Molecular Probes) and the nuclear counterstain Toto-3 (Molecular Probes) overnight at 4°C. Negative methodological controls included the absence of primary antibody (secondary-alone) to assess auto-fluorescence and nonspecific staining. After incubation, the constructs were washed thrice with PBS and observed using a Zeiss Microscope (Zeiss Axiovert 200 MOT inverted fluorescence microscope with deconvolution software).
Quantitative polymerase chain reaction
ESC differentiation was assessed using markers of osteoblastic differentiation by both quantitative reverse transcription–polymerase chain reaction (qRT-PCR) and immunofluorescence. For qRT-PCR analysis, RNA was collected using Trizol (Invitrogen) and converted to cDNA using a high-capacity reverse transcription kit (Applied Biosystems [ABI]). The cDNA was probed using validated TaqMan primer/probe sets for mouse and human Sox9, Col2A1 (collagen II), ACAN (aggrecan), Sp7 (osterix), OCN (osteocalcin), Oct-4, and Nanog on an ABI 7900HT using 18S as the internal control and the ddCT method included within ABI software (rq manager version 1.1) to analyze results.
von Kossa assay
Cell cultures were fixed in 4% paraformaldehyde for 30 min, washed twice with PBS, and then stained. Fixed cells were incubated for 1 h in 5% AgNO3 solution and exposed to light for at least 30 min.
In vivo tumor formation assay
Undifferentiated ESCs transplanted into immunocompromised hosts can form teratomas. Briefly, Fox Chase CB-17 severe combined immunodeficiency (SCID) mice were obtained from Charles River and housed in the single-barrier animal facility of the Faculty of Medicine, University of Calgary. Animal protocols were carried out as approved by the Animal Care Committee at the University of Calgary according to the standards of the Canadian Council of Animal Care. Mice were fed ad libitum with a standard diet and water. One million viable cells were injected into the skin fold of the inner thigh of six mice per group. Undifferentiated cells were injected as single cells, whereas cell-loaded collagen constructs where injected intact. The animals were sacrificed at different time points, as outlined in the Results section, and emerging tissue material was dissected. Excised tissues were fixed overnight in 4% PFA at 4°C and then embedded in paraffin. Sections were stained in hematoxylin/eosin according to standard procedures.
In vivo imaging
Before injection of cells into SCID mice or incorporation of cells into the collagen scaffolds, the cells were incubated with 1μM IR-786 (Sigma) in serum-free media for 30 min at 37°C at 5% CO2. The cells were rinsed three times with Dulbecco's Phosphate-Buffered Saline (DPBS) and returned to serum-containing medium. After cell injection into a single thigh of a SCID mouse, the mice were returned to the animal facility until 21 days or 3 months after cell injection. The mice were sacrificed and scanned using a LICOR Odyssey near-infrared (IR) scanner.
Flow cytometry
Cells were dissociated into single-cell suspensions and subjected to fluorescence-activated cell sorting (FACS) to identify bone cell differentiation markers using a FACS Calibur instrument and the CellQuest software from Becton Dickinson. Ten thousand events were registered per sample, and analysis of whole cells was performed using appropriate scatter gates to avoid cellular debris and aggregates. Cells were stained using the following antibodies: bone sialoprotein, osterix, osteonectin, osteocalcin, collagen II, and Aggrecan. All primary antibodies were directly conjugated to R-Phycoerythrin (R-PE) or Alexa Fluor 488 using Zenon conjugation kits (Invitrogen) following the manufacturer's method. Anti-mouse or anti-goat IgG R-PE or fluorescein isothiocyanate were used as isotope controls depending on the primary conjugation molecule used.
Statistical analysis
Data from the FACS analysis and qRT-PCR were performed in triplicate. Statistical analysis (analysis of variance) was performed using GraphPad Prism4 (GraphPad Software) and significance was set at p<0.05.
Results
ESC differentiation using 2D gels
A series of experiments was initially carried out to observe the effects of collagen I films polymerized with BGP on undifferentiated mouse ESCs. A single-cell suspension was seeded on a polymerized collagen I film and changes in cell morphology were observed over 15 days in culture. The ESCs took up residence within the fibers of the collagen film; by 15 days, evidence of mineralization was observed within the films through light microscopy. Therefore, a von Kossa reaction was performed to detect mineralization, and nodules of mineralized cells were observed by day 15 (Fig. 1A). To determine if the mineralization was produced as a result of osteoblast differentiation and not spontaneous mineralization, undifferentiated cells (Fig. 1B–D) and cells from day 15 collagen films (Fig. 1E–G) were stained with osteopontin (Fig. 1B, E), alkaline phosphatase (Fig. 1C, F), and osteonectin (Fig. 1D, G). As expected, undifferentiated ESCs expressed osteopontin (bone sialoprotein 1) and alkaline phosphatase. However, by day 15 of differentiation, osteopontin and alkaline phosphatase were still expressed along with osteocalcin (which is not expressed in undifferentiated ESCs). The presence of osteogenic cells in the differentiated cultures was further confirmed with qPCR (Fig. 1H). RNA was collected from cells on 2D collagen films on days 0, 5, 10, and 15 of differentiation and probed with primer sets specific to osterix (Sp7), osteocalcin, and 18S as an internal control. Osteopontin was not used for qRT-PCR analysis since it has very high expression in day 0 controls. Significant increases in osterix and osteocalcin mRNA was observed at days 5, 10, and 15 compared to day 0 controls, which is consistent with osteoblast differentiation.
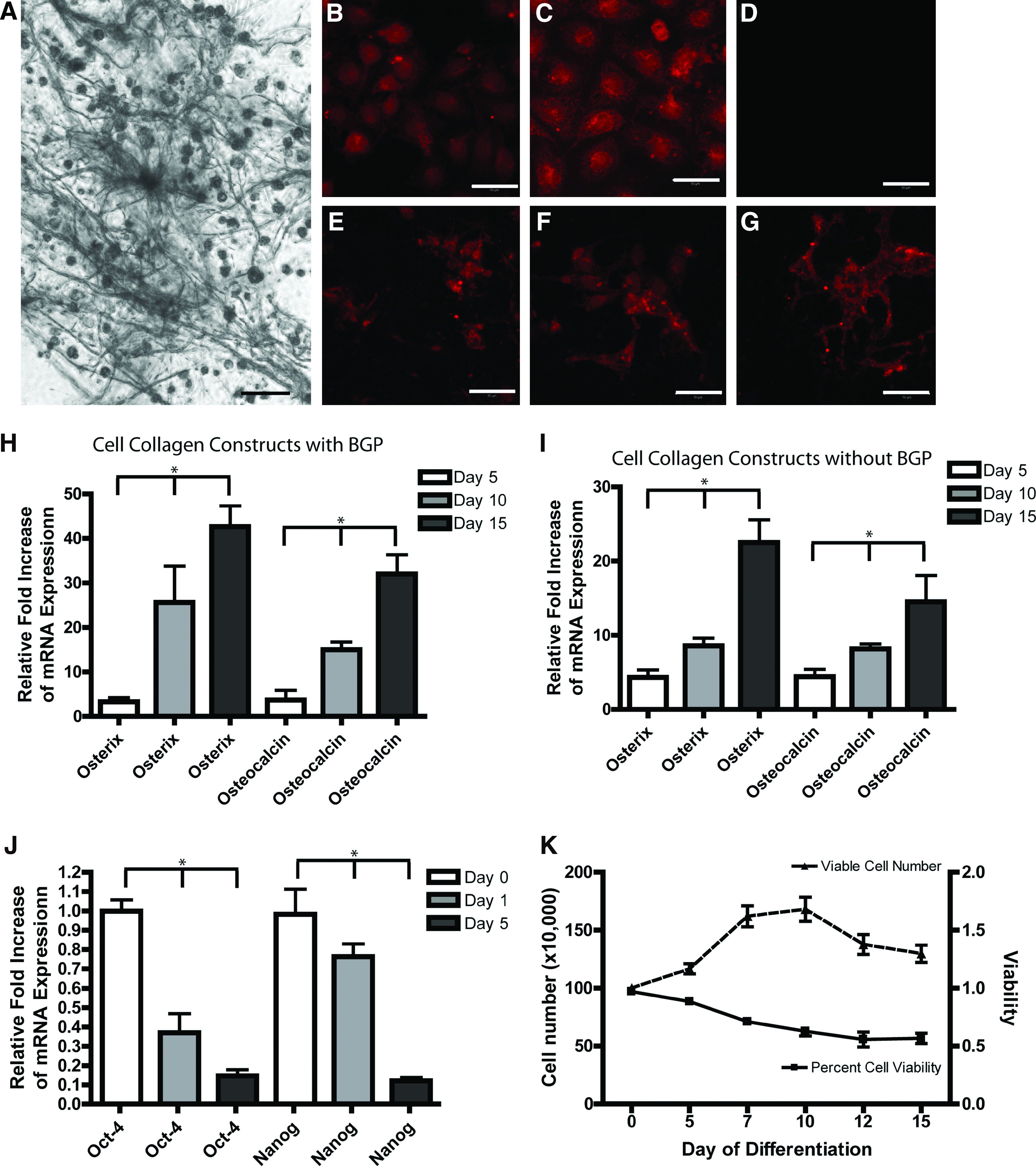
Osteoblast differentiation on a 2D collagen film. ESCs were cultured on a collagen-BGP film and stained with von Kossa to assay for mineralization at day 15
To examine the effect of BGP on ES differentiation, collagen I films were prepared without BGP and analyzed using qRT-PCR (Fig. 1I). Collagen I alone was able to induce the expression of osteoblast markers osterix and osteocalcin; however, expression of these markers was enhanced when the collagen I films were combined with BGP (Fig. 1H).
Expression of the pluripotency markers Oct-4 and Nanog was significantly decreased in ESCs that were exposed to the collagen-BGP scaffold by 24 h (Fig. 1J). Expression of both Oct-4 and Nanog further decreased between days 1 and 5 of differentiation, after which expression of these markers was not detected.
To examine the viability and proliferation of cells within the collagen-BGP films, the films were digested with collagenase at days 0, 5, 7, 10, 12, and 15 of differentiation, and cell counting was performed with a trypan blue exclusion assay (Fig. 1K). Overall, the viability of the cells within the films decreased over the differentiation period, leveling off at ∼60%. Further, an increase in viable cell density was observed until day 7, after which the number of cells decreased for the remainder of the differentiation period (Fig. 1K).
To compare the overall efficiency of differentiation between the collagen film osteogenic induction method and methods using EBs with media supplementation,7,10 we used FACS analysis to quantify the number of cells expressing osteogenic markers. Undifferentiated mouse ESCs expressed only osteopontin as expected (Fig. 2A); however, by day 15 of differentiation on collagen I it was observed that at least 75% of the cells were positive for osteopontin, osteocalcin, and osterix (Fig. 2A). Fewer than 25% of the cells were positive for aggrecan and collagen II, both markers of chondrogenesis (Fig. 2A). By comparison, it was found that exposure to dexamethasone induced osteogenic differentiation of the cells at an efficiency of ∼25% (Fig. 2B), while exposure to VD3 resulted in an efficiency of ∼15% (Fig. 2C). Furthermore, under routine microscopic analysis of VD3-induced cultures, multi-nucleated cells were observed, suggesting the presence of osteoclasts. VD3-differentiated cells were then stained with tartrate-resistant acid phosphatase and ∼40% of VD3 cells stained positive for this marker (Fig. 2C).
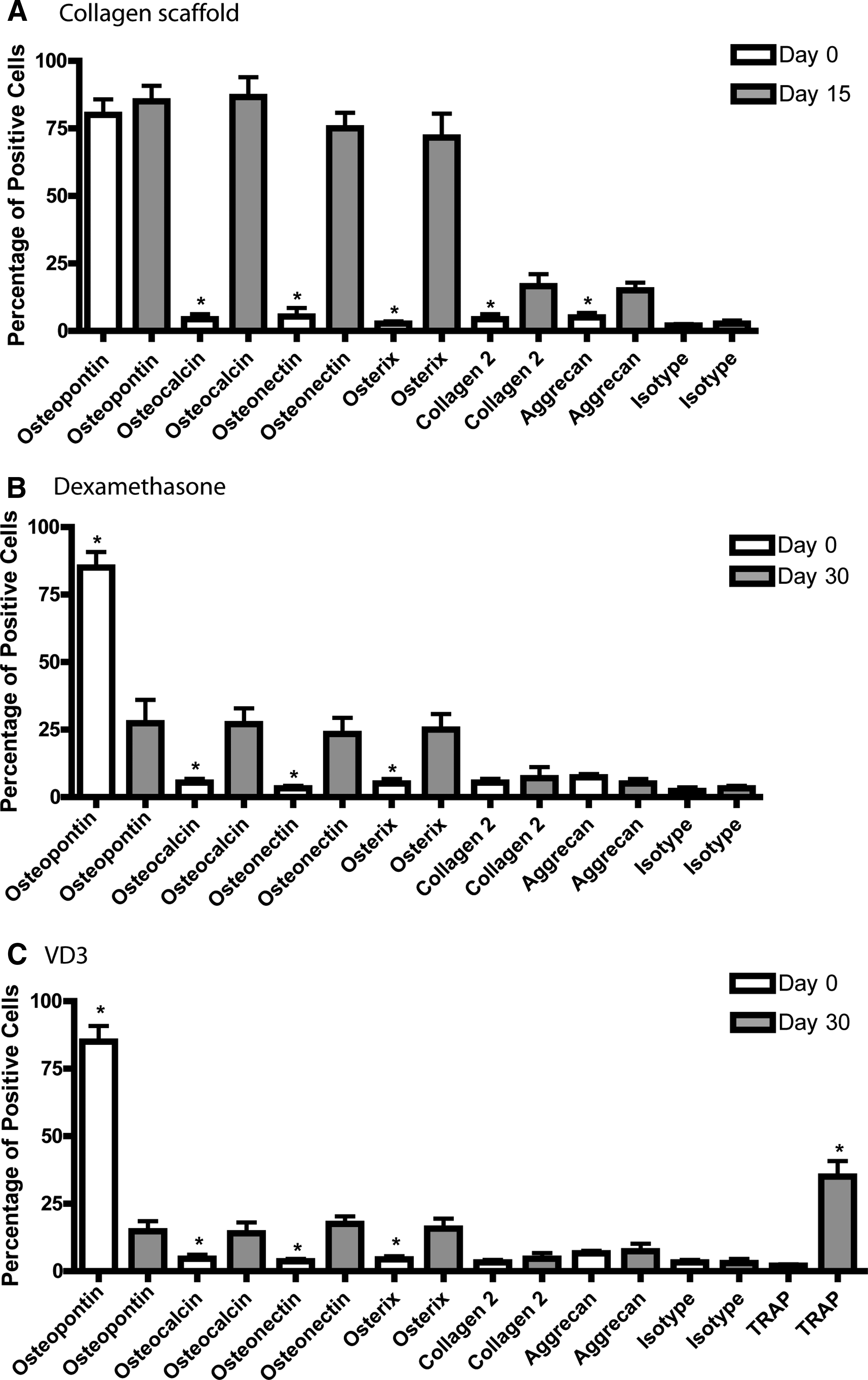
FACS analysis of multiple osteogenic induction methods. Cells were assayed using flow cytometry on days 0 and 15 of collagen-BGP-induced ES differentiation
To further test the efficacy of the collagen matrix to drive osteogenic differentiation, day 15 films loaded with cells were injected subcutaneously into the thigh of 5 SCID mice. After 20 days, hard white nodules were observed at the site of implantation (Fig. 3A). Upon X-ray visualization, these tissues were radio-opaque (Fig. 3A inset), and after histological analysis tissues resembling mineralized bone were observed (Fig. 3B, C). When the mice were imaged with a near-IR scanner, the IR membrane dye conjugated to the injected cells was observed only within the area where the cells were originally injected (Fig. 3D), suggesting that limited/no migration of the cells had occurred. Collagen films alone (without ESCs) produced no identifiable tissues in vivo (Fig. 3E–G), and as expected no cells were observed at the site of injection after 21 days in vivo (Fig. 3H).
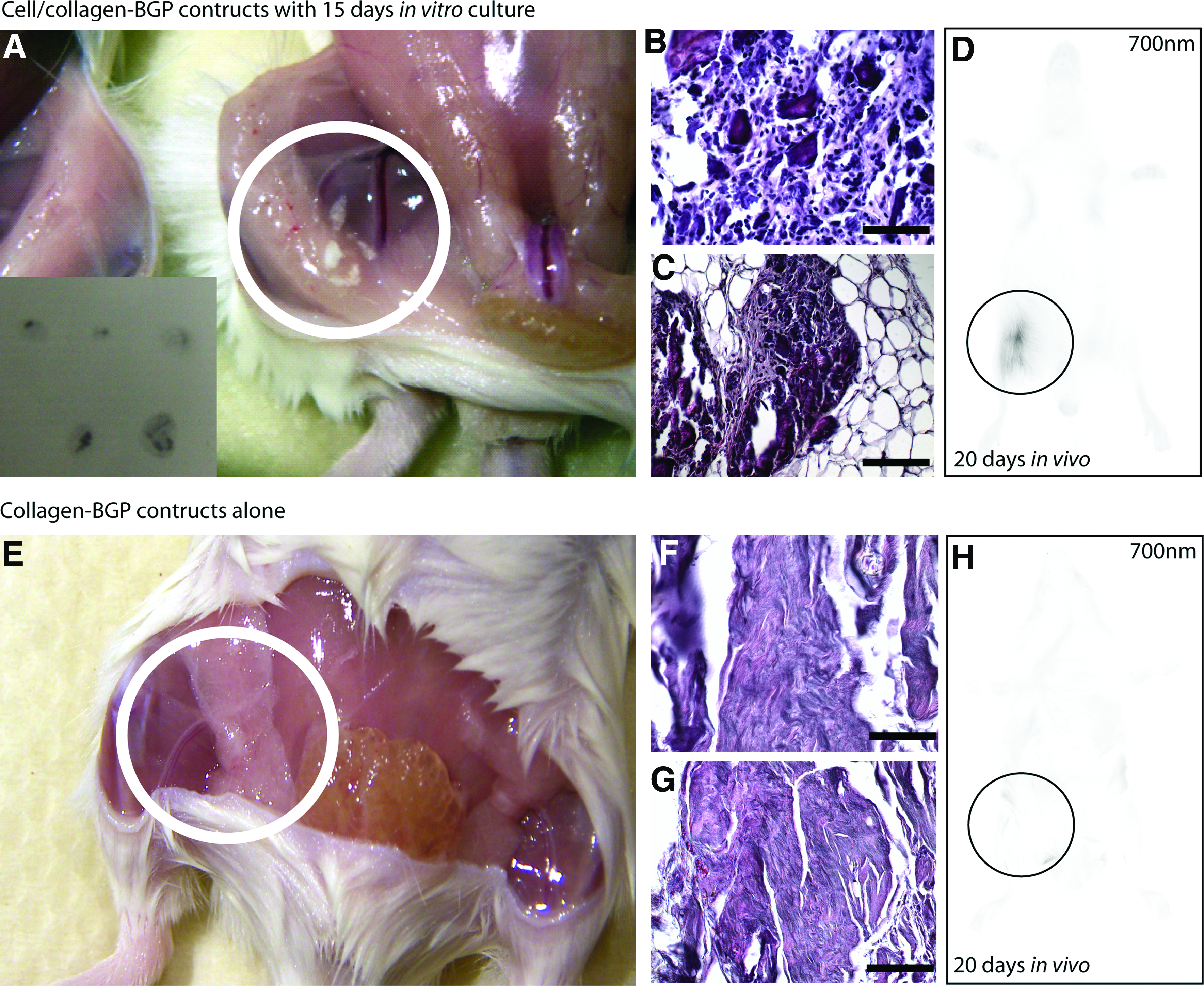
Injection of collagen-induced ESCs into a SCID mouse. ESCs were allowed to differentiate the collagen-BGP films for 15 days in vitro and then injected into a SCID mouse and allowed to differentiate in vivo for an additional 20 days. The resultant tissue was hard and white (
3D collagen gel induced differentiation
Since it was observed that collagen polymerized with BGP could induce osteoblast differentiation when cultured as a 2D film, the method was extended to differentiate ESCs into osteoblasts using a 3D collagen gel. The development of the 3D gel was undertaken for two reasons. First, although the collagen films induced efficient differentiation of ESCs, media changes where quite difficult, as removal and replacement of media usually caused the collagen film to detach from the culture dish. Second, the development of a 3D construct with the necessary media components built into the gel would create a self-sustaining culture environment that would maintain the viability of the cells while providing a transplantable cell-loaded construct.
As before, the expression pattern of a number of osteoblast and chondrocyte markers was examined with qRT-PCR and immunofluorescence over the course of ESC differentiation within the 3D collagen gels. mRNA collected from 3D collagen cell/gel constructs was probed with primers against osterix and osteocalcin on days 0, 5, 10, and 15 of differentiation. Significant increases in mRNA expression of both genes was observed on days 10 and 15 (Fig. 4A). Because of the thickness (10–15 mm) of the 3D collagen gels, high magnification imaging was not possible without removing the gels from the culture dishes. This was found to disrupt the overall uniformity and consistency of the matrix resulting in poor quality images. Therefore, low-power fluorescent images were taken to show the global changes in osteogenic markers at the protein level. Consistent with the 2D collagen I film it was observed that osteopontin was expressed in undifferentiated cells (Fig. 4B), but was not present on day 10 of differentiation (Fig. 4C); however, osteopontin was present by day 15 (Fig. 4D). As expected, osteocalcin (Fig. 4E) and osterix (Fig. 4F; both of which are not expressed in undifferentiated ESCs) were also observed by day 15. Control samples were used to examine unspecific binding of the secondary antibodies (anti-mouse and anti-goat 488) and these depicted minimal staining (Fig. 4G, anti-mouse shown).
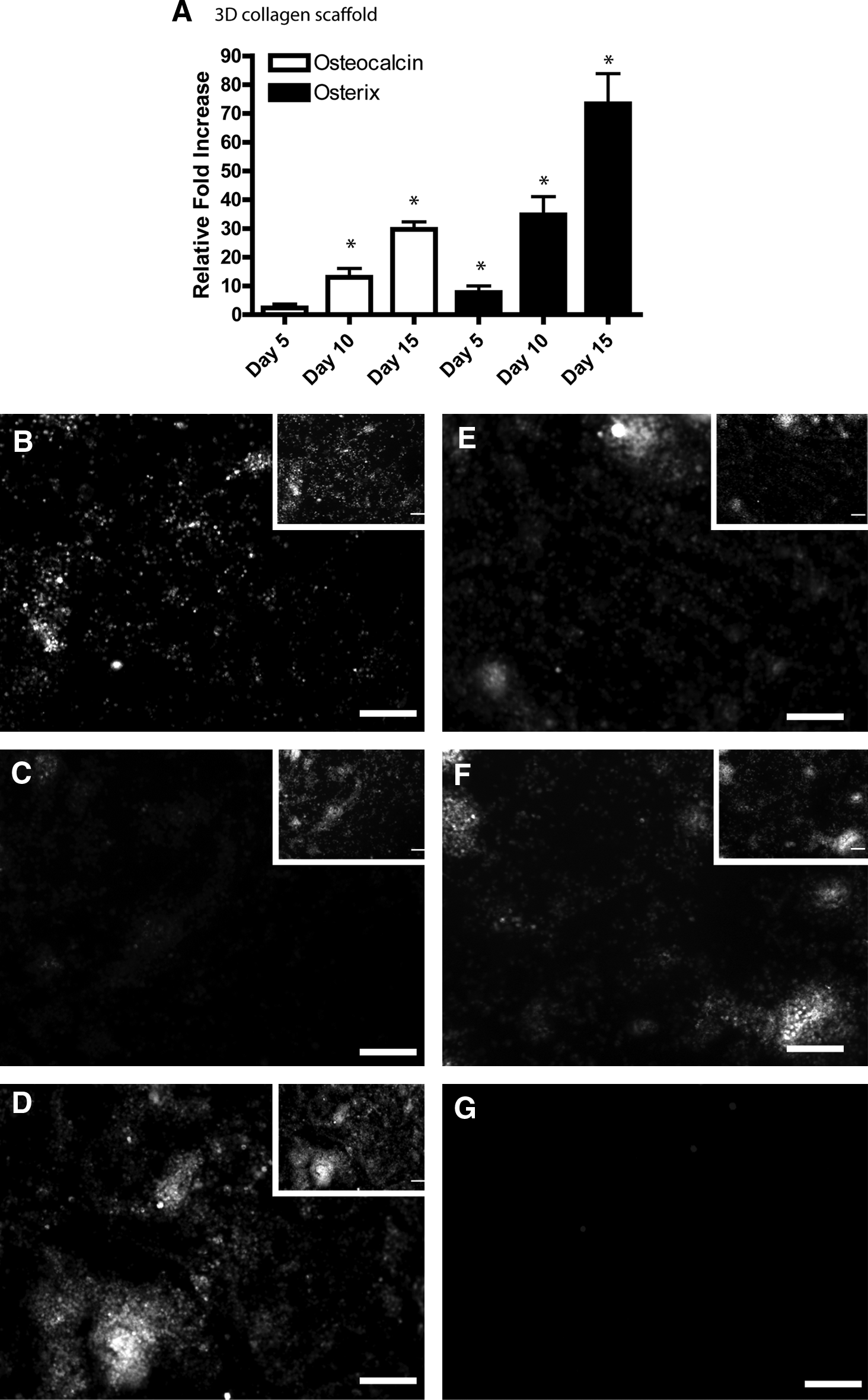
Characterization of osteogenic differentiation of ESCs in 3D collagen I gels. Cells were allowed to differentiate for 15 days in vitro and then assayed for marker expression using qRT-PCR and immunofluorescence. qRT-PCR analysis demonstrated an increase of osteocalcin and osterix mRNA during differentiation
As chondrogenic markers were observed in the 2D collagen films by FACS analysis (Fig. 2A), expression of the aggrecan and collagen II markers in cells differentiated in 3D collagen gels was examined using qRT-PCR and immunofluorescence. Using immunofluorescence, expression of collagen II and aggrecan was observed in a subset of cells by day 15 (Fig. 5A, B). By comparison, when the collagen was supplemented with chondroitin sulfate a dramatic increase in the number of aggrecan (Fig. 5C)- and collagen II (Fig. 5D)-expressing cells was observed by day 15 of differentiation. Secondary antibody controls displayed minimal staining (Fig. 5E, anti-mouse shown). Again, these results were verified using qPCR, which demonstrated increased transcription of aggrecan (ACAN), collagen 2 (col2A1), and Sox 9 when chondroitin sulfate was combined with the collagen (Fig. 5F).
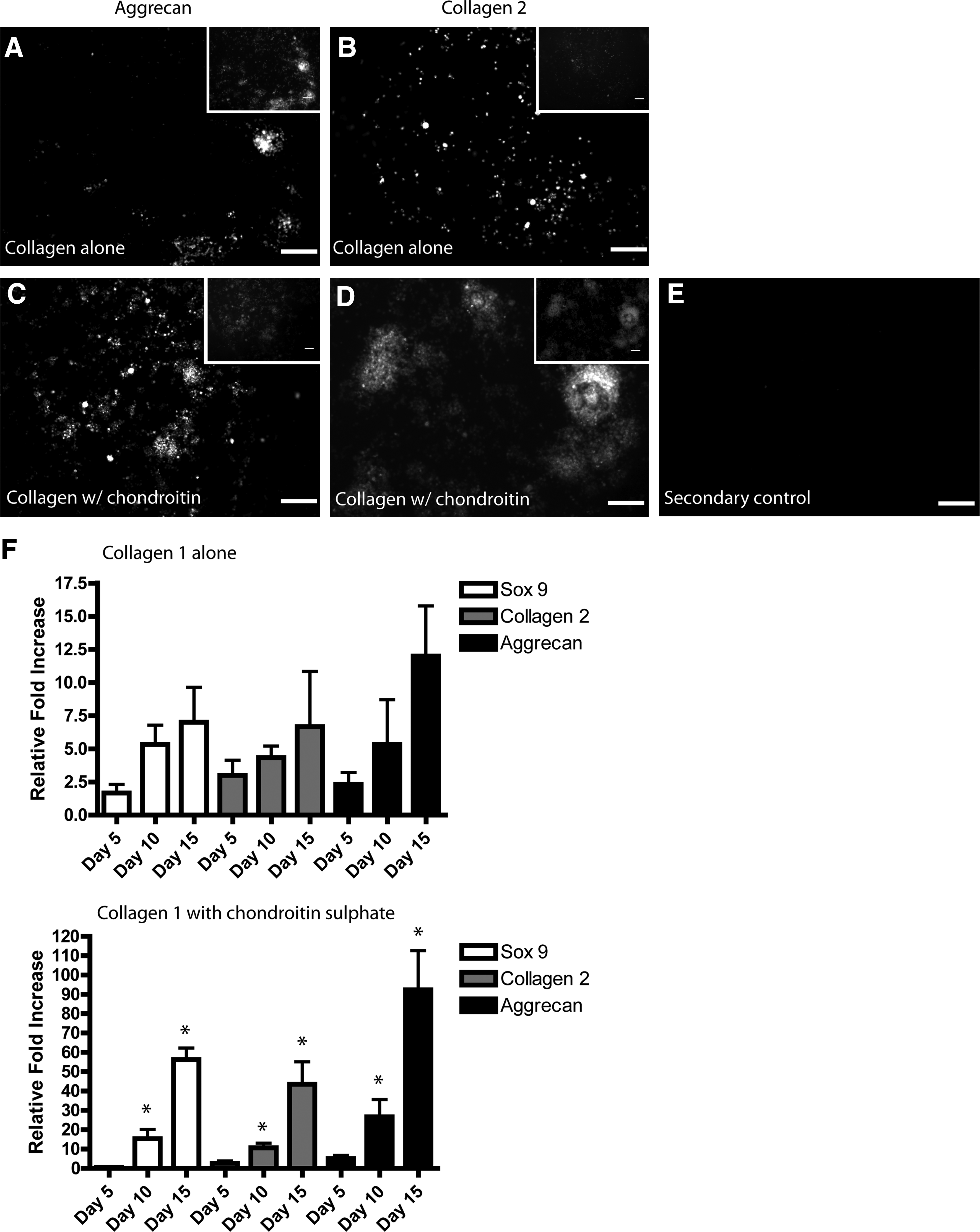
Characterization of chondrogenic differentiation of ESCs in 3D collagen I gels. Cells in 3D collagen gels were stained with aggrecan
Exposure to collagen inhibits tumorigenicity of ESCs in vivo
Undifferentiated ESCs implanted subcutaneously generated teratomas within 18–20 days after injection into the thigh of a SCID mouse (Fig. 6A–C). No tumors or teratomas were generated when ESC-loaded collagen gels were implanted subcutaneously into SCID mice, even without the use of cell purification and/or selection. When undifferentiated ESCs were cultured on collagen I matrix and allowed to incubate in vitro for 15 days and then injected into a SCID mouse, bone-like tissue was excised from the mouse after 1 month (Fig. 6D, E). Furthermore, when these constructs were allowed to remain in vivo for 6 (Fig. 6F) or 8 (Fig. 6G) months, woven bone-like tissue was generated and no tumors were observed. Similar results were achieved when the ESCs were differentiated on chondrotin sulfate-supplemented collagen-BGP gels before implantation: no tumors were detected (Fig. 6H); however, mature articular cartilage tissue was not observed, but a dense fibrous tissue was detected (Fig. 6I).
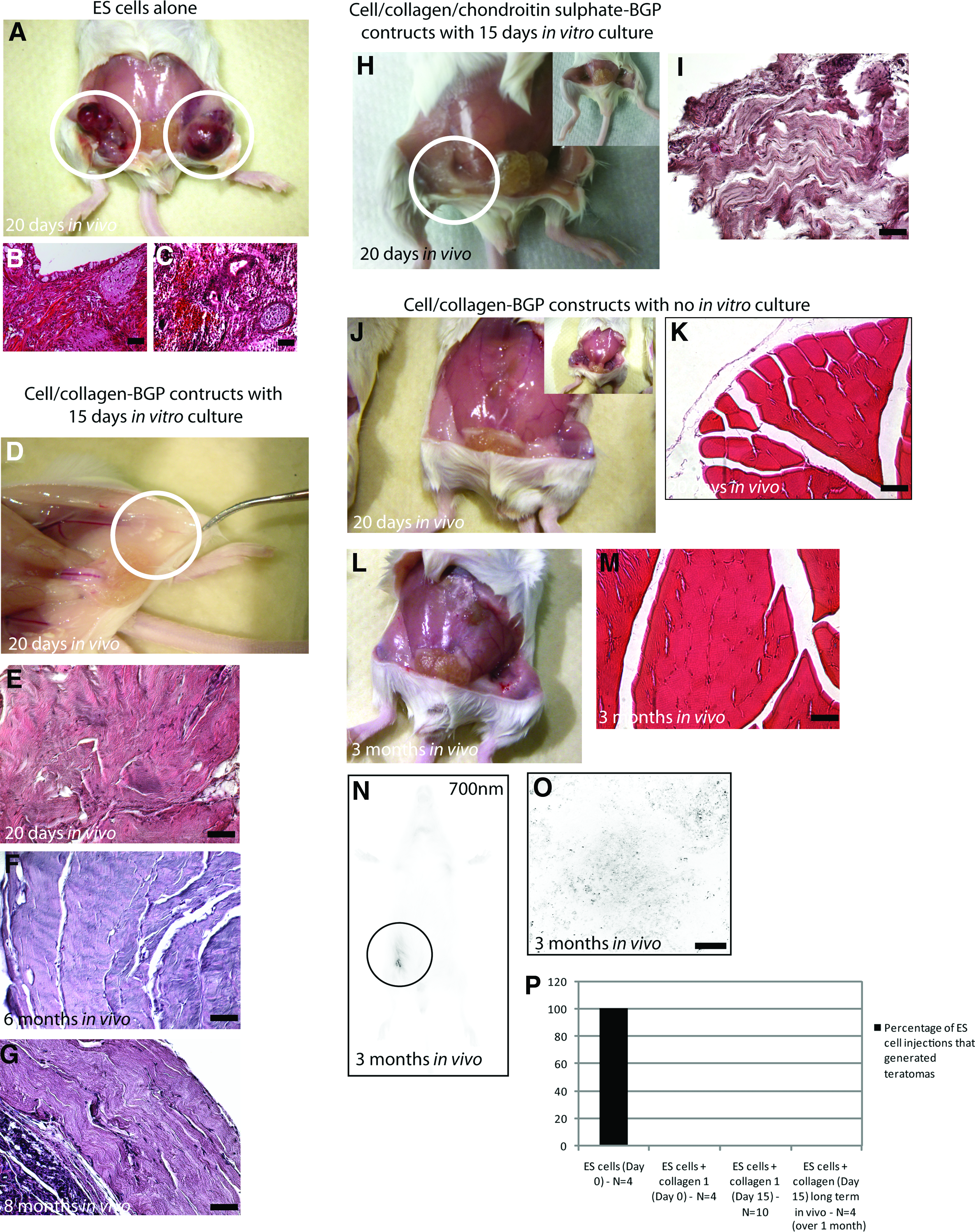
The risk of tumor (teratoma) formation is reduced when ESCs are cultured within collagen. Control undifferentiated mouse ESCs normally produce large teratomas after 20 days in vivo (
When undifferentiated ESCs were mixed with collagen-BGP and directly injected into SCID mice, no tumors or teratomas and no mineralized tissue was observed for up to 3 months in vivo (Fig. 6J, L), and when the muscle was biopsied for histology, no tumors or cysts were observed (Fig. 6K, M). Implanted cells were labeled with an IR marker, permitting long-term in vivo cell tracking. In vivo imaging of the mice at 3 months demonstrated a strong signal only at the area of initial injection (Fig. 6N), and when the tissue was biopsied and scanned at the highest resolution possible, individual cells (black spots) were observed within the endogenous tissue (Fig. 6O). Furthermore, no tumors were ever observed when ESCs were injected in vivo with collagen over the time period examined (Fig. 6P).
Discussion
ESCs and iPS cells hold great promise within regenerative medicine, yet before these cells can be safely utilized as therapeutics, reproducible and efficient differentiation methods need to be developed. 15 Furthermore, the risk of tumor formation needs to be completely controlled and eliminated before ESCs can be used in patients.16–18 Over the last two decades a number of protocols have been developed to differentiate ESCs into osteoblasts and chondrocytes.6–11,19–21 These studies have mainly involved the use of specific growth factors and chemicals that are added to the differentiation cultures after an EB formation step.6–11 However, one of the main problems with osteoblast differentiation of ESCs is selection of cells after mineralization. If immature osteoblasts are selected for transplantation, there are no definite markers available to separate the preosteoblasts from ESCs since pluripotent ESCs express a number of osteoblast markers, including alkaline phosphatase and osteopontin, and preosteoblasts do not express osteocalcin and other mature markers. 20 By contrast, if mature osteoblasts are selected for transplantation, it is quite difficult to separate them from the mineralized matrix in the culture dish, and most of the harvested cells do not thrive in vivo. 20 This study demonstrates the development of a system in which ESCs are cultured in a collagen microenvironment that fosters osteoblast differentiation (or chondrocyte differentiation, with the addition of chondroitin sulfate) and inhibits tumor formation in vivo. These cell-loaded collagen constructs are able to generate mineralized bone tissue after implantation in vivo without the necessity for purification and/or selection. In concurrent studies within our lab, these constructs have been implanted into a mouse fracture model and increased bone formation was observed at the implantation site. Furthermore, no tumors were observed in this study (article in preparation). These findings are in contrast with other mouse ESCs studies where tumors were found to destroy the native tissue after implantation. 22 It is also important to note that no growth factors or supplemental chemicals (except BGP) were necessary to drive ESCs to osteoblasts or chondrocytes in this study. Through modification of the ECM alone, we were able efficiently generate osteoblasts and chondrocytes. Recent studies have demonstrated the MSCs react in a similar fashion in regard to osteogenesis when cultured within a collagen I scaffold, 14 signifying that the effect of collagen I exposure on stem cell differentiation is conserved from an early progenitor stage. This is particularly interesting since collagen I is expressed in almost every tissue, and one could make a convincing argument for MSC specificity for collagen I because of bone composition; however, ESCs would interact with collagen I in vivo yet still are able to produce tissues other than bone or cartilage. Therefore, it may be the mixture of collagen I with free phosphate that drives the ESCs down a mesenchymal pathway, or that collagen I does indeed default for bone and that the in vivo ECM composition provides additional signals that drives the cells down other lineage pathways. In our study it appears that although collagen I alone can direct ESCs down an osteogenic lineage, the effect is amplified when collagen I is cross-linked with BGP. This collagen/BGP synergy may add a level of specificity for differentiation into bone and cartilage tissues that would normally be augmented with growth factors (BMPs, etc.) in vivo. It is also important to note the differential effects on cell differentiation when chondroitin sulfate was supplemented into the collagen I/BGP scaffolds. Under these conditions we observed a strong shift from osteogenesis to chondrogenesis. This is particularly interesting since other groups have demonstrated increased chondrogenesis in both adult stem cells and ESCs with the addition of glucosamine and/or chondroitin sulfate in hydrogels.23,24 These results, taken together, suggest that proteoglycans within joint tissues may play pivotal roles in the differentiation and maturation of progenitor cell populations within that environment.
This study demonstrates that a collagen I construct polymerized with BGP can induce mouse ESCs down an osteogenic pathway. Furthermore, with the addition of chondroitin sulfate this pathway can be altered to favor a chondrogenic fate. These cell-loaded collagen constructs are not only useful for studying the pathways involved in ESC differentiation, but also valuable for skeletal tissue engineering, since the transplanted constructs will not develop into a tumor, even without an in vitro incubation period. This signifies that collagen I is not only implicated in ESC differentiation, but also the regulation of the tumorigenic potential of the cells in vivo.
Footnotes
Disclosure Statement
No competing financial interests exist.