
Abstract
The aim of this study was to evaluate whether human Wharton's jelly cells (WJCs) could be differentiated into nucleus pulposus (NP)-like cells by coculturing with NP cells (NPCs) in vitro. WJCs were isolated from the human umbilical cord, and NPCs were isolated from healthy human intervertebral disc. After coculturing WJCs with NPCs in a monolayer environment with or without cell–cell contact for 7 days, the real-time polymerase chain reaction showed the relative gene expressions of NP-marker genes (aggrecan, type II collagens, and SRY-type HMG box-9) were significantly increased (p<0.05) in all groups, and the increase in the group of 25:75/WJCs:NPCs was the largest (p<0.05). The increases of relative gene expression in WJCs cocultured with cell–cell contact were larger than those cocultured without contact in all ratios (p<0.05). WJCs were positive for telomerase expression. Flow cytometry analyses showed that WJCs expressed CD73, CD105, CD90, CD29, CD166, and human leukocyte antigen (HLA)-ABC while being negative for the expression of CD34, CD45, and HLA-DR. The results of this study indicated that the WJCs had the feature of the mesenchymal stem cell and might be induced to differentiate to NP-like cells by coculturing with NPCs.
Introduction
One possibility is the use of tissue engineering or cell transplantation techniques to deliver viable cells to the NP to reverse the degeneration progress, which are capable of surviving within the IVD and producing extracellular matrix with similar or improved biological and biomechanical properties compared with the original.3,4 This strategy was first attempted by reinserting autologous NP into the IVD in 1998. The author found that reinsertion of NP decelerated disc degeneration. 5 Ganey et al. transplanted autologous disc chondrocyte cells into degenerated IVDs, where it was shown that disc chondrocyte cells remained viable after transplantation and produced an extracellular matrix that contained components similar to normal IVD tissue. 6 However, these methods have practical limitations in the clinical, because harvesting of the patient's own cells or NP from a disc can accelerate degeneration within that disc, whereas removal of cells from an adjacent normal disc would induce degeneration at that disc level. In addition, the IVD is relatively acellular tissue with only approximately 4,000 cells per mm3. 7 So, the suitable source of viable cells is one of the key problems for the cell-based biologic repair strategy.
Due to proliferative potential and multidifferentiation capacity, adult mesenchymal stem cells (MSCs) provide an attractive choice for managing IVD degeneration. Previous studies have proved that adult MSCs such as bone marrow-derived MSCs (BMSCs) 8 and adipose-derived stem cells (ADSCs) 9 could differentiate toward a NP-like cell phenotype in vitro. Studies demonstrated that BMSCs could differentiate toward a phenotype consistent with NPCs in the presence of hypoxia and transforming growth factor-β1 (TGF-β1) 10 Richardson showed that cell–cell contact coculture of NPCs and BMSCs caused BMSCs to upregulate the gene expression of NP marker genes: SRY-type HMG box-9 (SOX-9), type II collagen, and aggrecan, and differentiate to an NP-like phenotype. 3 After being transplanted into the NP, BMSCs could survive and proliferate within the discs with 100% viability 11 and increase the amount of proteoglycans in IVDs, 12 and BMSCs differentiated into cells expressing some of the major phenotypic characteristics of NPCs. 13 Lu demonstrated that ADSCs could be induced to differentiate toward the NP cell-like phenotype by coculture with NPCs. 9 Autologous ADSCs could improve the quality of a reconstructed NP tissue in vitro 14 and be effective in promoting disc regeneration. 15
However, the harvest of adult MSCs is a highly invasive procedure. Moreover, BMSCs represent only a small percentage of the total number of cells in bone marrow, and the number of cells useful for regenerative medicine applications is extremely low (0.001% to 0.01%) 16 and yield of MSC from bone marrow also significantly decreases with donor age.17,18 In addition, it has been observed that older individuals have a lower number of ADSCs as compared with younger subjects. 19
In recent years, Wharton's jelly cells (WJCs) have been showed as a new alternative source of MSCs, a population of cells deriving from umbilical cord Wharton's jelly. WJCs are multipotent and can be induced to differentiate to various cell types such as cardiomyocyto, osteogenic, adipogenic cell, 20 neural cell, 21 and myogenic cell 22 under suitable culture conditions. We hypothesize that WJCs have the potential to differentiate into NP-like cells, and might be injected into the IVD by tissue-engineering or cell-transplantation techniques and produce disc extracellular matrix with similar biological and biomechanical properties compared with the original NP. If this is the case, then the possibility exists that WJCs might be used in managing IVD degeneration. It is well known that differentiation of MSCs depends primarily on the specific environment in which they are placed. 23 Previous researches had shown that coculture of NPCs and MSCs could cause MSCs to speed up the conversion to a chondrogenic/discogenic phenotype 24 and differentiate to an NP-like phenotype. 3 In the current study, we investigate the effect of NPCs stimulation on the differentiation of WJCs to NP-like cells in two in vitro coculture systems. In this study, we cocultured human WJCs with NPCs in transwells (with 0.4 μm pore-size filter inserts) without cell–cell contact and in six-well plates with cell–cell contact at a variety of different ratios for 7 days. Gene expressions of the transcription factor SOX-9, aggrecan, versican, the collagens types I, II, and VI were measured by real-time polymerase chain reaction (PCR) to examine the differentiation states of the WJCs.
Materials and Methods
NP cell isolation and culture
Human normal IVD tissue was obtained from disc level T12-L1 of a 25-year-old man, who had undergone a fusion surgery because of thoracolumber burst fracture. The IVD showed no evidence of degeneration based on the system described by Sive et al. 25 Informed consent from the patient and approval was obtained for the use of the sample for research. Cartilaginous endplates and the annulus fibrosus tissue were carefully removed from the specimens, and the NPCs were isolated as previously described. 26 Briefly, NP tissues were digested with 0.4 mg/mL collagenase type II (Sigma) solution in serum-free medium containing 100 U/mL penicillin, 100 mg/mL streptomycin, and 2.5 mg/mL amphotericin B antibiotics for 5 h at 37°C; the digested tissue/cell suspension was passed through a 100 μm cell strainer to remove tissue debris, and cells were then pelleted by centrifugation at 400 g for 5 min. The supernatant was removed, and cells were resuspended and cultured to confluence in a 25-cm2 flask with Dulbecco's modified Eagle's medium/F12 medium (DMEM/F12; Hyclone), supplemented with 10% fetal bovine serum (FBS), 100 U/mL penicillin, 100 mg/mL streptomycin, and 2.5 mg/mL amphotericin B at 37°C in a humid atmosphere containing 5% CO2. Culture medium was changed twice a week.
WJC isolation and culture
About 30 cm long umbilical cord was obtained after normal birth and stored in sterile D-Hanks solution at 2°C–6°C before tissue processing to obtain WJCs. Informed consent from the patient and approval from the hospital was obtained for the use of the sample for research purpose. WJCs were isolated as previously described. 20 Arteries, veins, and epithelium were stripped and discarded from umbilical cord tissue with surgical tweezers. The Wharton's jelly was cut into fragments with a scalpel. Then, the fragments were treated with 0.2 mg/mL collagenase type II (Sigma) solution in serum-free medium containing 100 U/mL penicillin, 100 mg/mL streptomycin, and 2.5 mg/mL amphotericin B antibiotics for 18 h at 37°C; washed; and treated with 2.5% trypsin for 30 min at 37°C with agitation. Finally, the cells were washed and cultured in DMEM/F12 medium supplemented with 20% FBS in 5% CO2 in 37°C incubator. The medium was changed once every 2 days. Once the cells reached 80%–90% confluence, they were ready to be sub-cultured. After first passage, the cells were then seeded at a density of 4×103cells/cm2 in DMEM/F12 medium supplemented with 10% FBS, with the culture medium being replaced every 2–3 days until the cells were ready for sub-culture and seeding was required. The third passage cells were used for coculture.
Flow cytometry analysis
WJCs at passage 3 were taken for flow cytometric analysis. The cells were trypsinized and suspended in phosphate-buffered saline (PBS) at a concentration of 1×106/mL, and washed twice with PBS. Collected cells were then incubated with antibodies against phycoerythrin (PE)-conjugated CD105, CD73, CD45, CD29, CD166, human leukocyte antigen (HLA)-DR, HLA-ABC, and fluorescein isothiocyanate (FITC)-conjugated CD34, CD90 (all eBioscience). All analyses were standardized against negative control cells incubated with isotype-specific IgG1-PE and IgG1-FITC (eBioscience). Cells were incubated with labeled antibody (or isotype control) in the dark for 30 min at 4°C. After incubation, cells were washed with PBS and centrifuged to remove unbound antibody. Cells were resuspended in 1 mL PBS and analyzed by flow cytometry with use of EPICS XL MCL-4 (BECKMAN COULTER).
Telomerase activities detection
The WJCs cultured in six-well plates (3×105cells/well) were pooled to produce 1 μg of protein extract for each assay. Telomerase activity was measured by using the TRAPEZE XL Telomerase detection kit (Millipore) as per manufacturer's instructions. Fluorescence energy transfer primers permitted nonisotopic, quantitative analysis of telomerase activities. The fluorescence was measured with a fluorometer (Berthold Twinkle LB970) in a black 96-well plate (Corning) with excitation/emission parameters for fluorescein (485 nm/535 nm) and sulforhodamine (585 nm/620 nm). The formulae used to calculate relative telomerase activities was: [(ΔFL sample − ΔFL heat inactivated sample) /ΔR)/(ΔFL/ΔR positive control)]×100. We used Hela cells as the positive control, and NPCs as the negative control.
Fluorescent labeling of WJCs
WJCs were trypsinized, counted, and resuspended at a concentration of 1×106 in DMEM/F12, washed twice with PBS. Then, 5, 6-carboxyfluorescein diacetate, succinimidyl ester (CFSE; sigma) was added. After being incubated for 30 min at 37°C, the reaction was stopped by adding serum with a final concentration of 40% of the volume for 10 min, centrifuging at 400 g for 5 min, and resuspending in DMEM/F12 to a concentration suitable for seeding.
Coculture of WJCs and NPCs
Transwell six-well plates with 0.4 μm pore size, high-pore density, polyethylene terephthalate (PET) track-etched tissue culture inserts were used for coculture without contact. For coculturing without contact, CFSE-labeled WJCs were seeded on the base of the six-well tissue culture plate, whereas NPCs were seeded on to the upper surface of the membrane of a tissue culture insert. For coculturing with direct cell–cell contact, both NPCs and CFSE-labeled WJCs were mixed in different ratios and seeded on the base of the six-well plate. Cells were seeded at 3 different ratios: 25:75%, 50:50%, and 75:25% (WJCs/NPCs). The actual number of cells seeded in each well is summarized in Table 1. Cocultured cells were maintained for 7 days in DMEM/F12 at 37 °C and 5% CO2 in a humidified atmosphere with the medium being changed every 2 days.
WJCs, umbilical cord Wharton's jelly cells; NPCs, nucleus pulposus cells.
Separation of WJCs and NPCs cocultured with cell–cell contact
Seven days after coculture with cell–cell contact, MoFlo high-speed cell sorter (Dakocytomation) was used for cell sorting. Cocultured cells were trypsinized, resuspended in DMEM/F12, and pelleted by centrifugation at 400 g for 5 min. The pellet was resuspended in 200 μL of DMEM/F12 and passed through a 30 μm sterile filter to remove cell clumps. The samples were then analyzed and sorted by using a MoFlo high-speed cell sorter. Excitation wave length was 490 nm, whereas emission wave length was 530 nm. Fluorescence-positive cells were WJCs, and negative cells were NPCs. Cells were collected in a centrifuge tube containing PBS solution.
Real-time PCR analysis of gene expression
Total RNA was extracted from cells by using Trizol reagent (Invitrogen) according to the manufacturer's instructions. For cells cocultured without contact, the inserts were removed, and cells were rinsed in PBS, and Trizol reagent was added to both the original wells (WJCs) and the inserts (NPCs) to lyse the cells at room temperature for 5 min. Then, chloroform was added to each sample at room temperature for 5 min. All sample tubes were centrifuged at 400 g for 5 min at 4°C to enable phase separation. RNA was precipitated by addition of isopropanol to the aqueousphase, followed by centrifugation. Precipitated RNA pellets were washed in 75% ethanol and then resuspended in distilled RNAse-free water. RNA was reverse transcribed into cDNA by using AMV reverse transcriptase (Takara Biotechnology). 1 μL RNA was mixed with 50 pmol random primer, 10U AMV reverse transcriptase, 2 μL dNTPs, 4 μL 5× buffer, and 20 U RNase Inhibitor, and DEPC water was added, thus giving a final volume of 20 μL. Samples were then incubated at 42°C for 60 min after 10 min at home temperature. Then, the cDNA were obtained from RNA.
After the cDNA had been obtained by reverse transcription, relative gene expressions of aggrecan, type I collagen, type II collagen, type VI collagen, versican, and SOX-9 were determined by real-time PCR and normalized to the glyceraldehyde-3-phosphate dehydrogenase (GAPDH) housekeeping gene. Sequences of primers for human aggrecan, type I collagen, type II collagen, type VI collagen, versican, and SOX-9 genes used have been previously reported.3,27 GAPDH primers were designed by using the Premier 5.0 software as follows: human GAPDH (forward: GAA GGT CGG AGT CAA CGG; reverse: GGA AGA TGG TGA TGG GAT T) (NM_000576.2). The Mini OpticonTM Detector System (Bio-Rad) and SYBR Green PCR kit (Takara Biotechnology) were used for real-time PCR analysis.
The real-time PCR consisted of an initial enzyme activation step at 95°C for 20 s, followed by 40 cycles of 95°C for 5 seconds and 60°C for 20 s. A cycle threshold (Ct) value was obtained for each sample, and triplicate sample values were averaged. The 2−ΔΔCt value was then used to calculate relative expression of each target gene. 28 Data presented (mean) were from three independent experiments in which both sample sets were analyzed in triplicate.
Statistical analysis
The SPSS version 15.0 software was used for statistical analysis. For statistical analysis, results were presented as mean±standard deviation of coculture-over-control ratios of real-time PCR data (n=3). Significances were tested by using the Student's t-test, in which p<0.05 was considered significant.
Results
Characters of WJCs
WJCs were isolated from human umbilical cord explants and expanded as primary cultures. After attachment, the cells gradually spread out and were shown as fibroblast-like morphology after treatment with 10% FBS-DMEM/F12 for 3 days. With further culturing, the WJCs mostly appeared to be flat, wide, and polygonal after passaging, and were closely opposed to each other due to proliferation (Fig. 1A). To determine whether WJCs could have multipotent characteristics, we assessed the expression of a number of markers associated with MSCs by flow cytometry of cultured WJCs at passage 3. According to the immunophenotypic analysis of flow cytometry, the WJCs we had isolated from the umbilical cord Wharton's jelly expressed MSC markers CD73\CD105\CD90\CD29\CD166 and HLA-ABC, but did not express the hematopoietic lineage markers CD34\CD45 and were negative for human leukocyte antigen HLA-DR (Fig. 2). WJCs were positive for telomerase expression. Relative telomerase activity was higher than the controls (Fig. 3).

Characteristics of WJCs and NPCs.
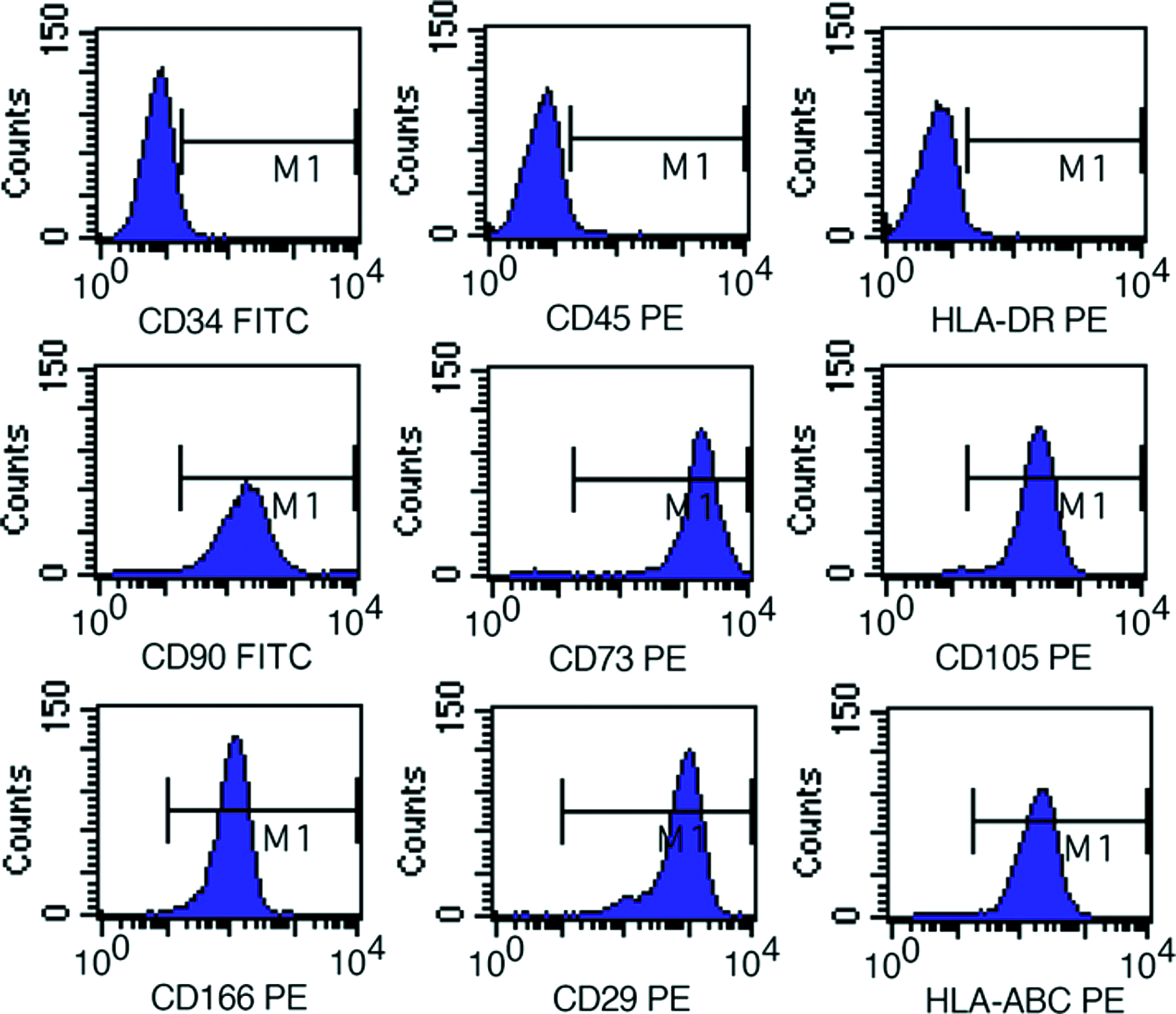
Flow cytometry analysis of surface-marker expression on WJCs. Phenotypes of WJCs at passage 3. Cells were labeled with PE- or FITC-conjugated antibodies and examined by flow cytometry. Cells were positive for CD73, CD90, CD105, CD29, CD166, HLA-ABC, and negative for CD45, CD34, and HLA-DR. Data are representative of three independent experiments. PE, phycoerythrin; FITC, fluorescein isothiocyanate; HLA, human leukocyte antigen. Color images available online at www.liebertonline.com/tea

The result of the telomerase activity. WJCs and control cells (hela cells) are positive. Negative control cells are nucleus pulposus cells. Color images available online at www.liebertonline.com/tea
High-speed cell sorting of NPCs and CFSE-labeled WJCs
The CFSE molecule was evenly divided when cells proliferate, which was important to discriminate between the two kinds of cells after coculture (Fig. 1C, D). The MoFlo high-speed cell sorter was used for analysis and sorting of the cells after coculture with cell–cell contact (Fig. 4).

MoFlo data showing parameters for separation of CFSE-labeled WJCs and unlabeled NPCs after coculture with cell–cell contact.
Effect of CFSE labeling on WJCs gene expression
To ensure that CFSE labeling had no effect on WJCs gene expression or phenotype, the expressions of SOX-9; collagen types I, II, and VI; aggrecan and versican genes were measured by real-time PCR after 7 days. All the genes showed no significant change in expression (Fig. 5).
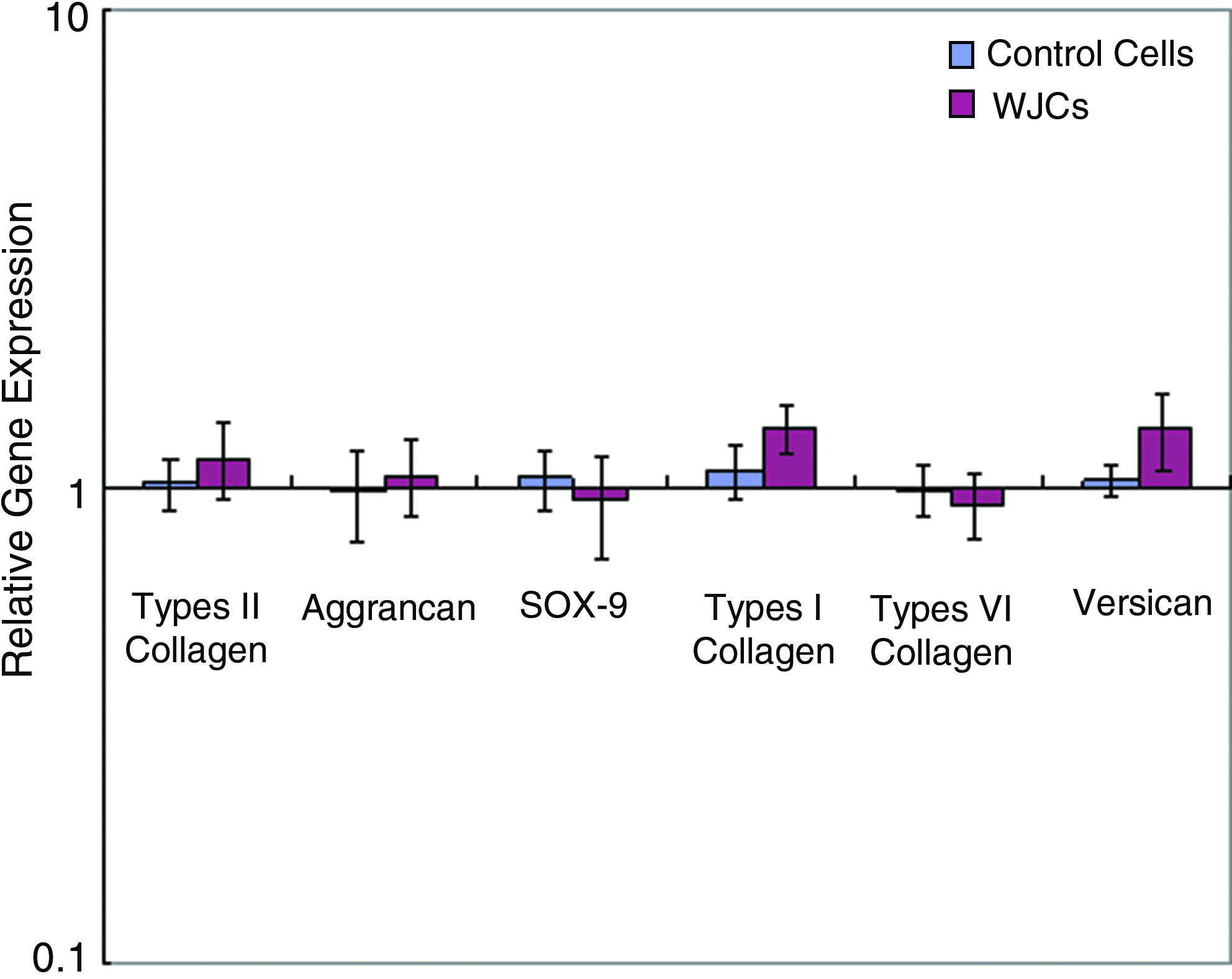
Relative expression of SRY-type HMG box-9 (SOX-9), aggrecan, types I, II, and VI collagen, and versican mRNA by CFSE-labeled and unlabeled WJCs after 7 days in culture. Gene expression in CFSE-labeled cells normalized to average of housekeeping genes (glyceraldehyde-3-phosphate dehydrogenase [GAPDH]) and unlabeled cells. Color images available online at www.liebertonline.com/tea
Gene expression of WJCs after coculture
After 7 days of coculturing, we used real-time PCR to determine the changes in gene expression. The results were expressed as relative gene expression. Although cells had been cocultured with direct contact, mRNA was significantly increased in WJCs in all three cell ratios (Fig. 6). After 7 days in coculture with contact, SOX-9 mRNA was significantly (p<0.05) increased in WJCs in all three cell ratios (Fig. 6A), with the largest increase being seen at a ratio of 25%:75% WJCs/NPCs (2429-fold over controls compared with 153-fold and 109-fold in 50%:50% WJCs/NPCs and 75%:25% WJCs/NPCs, respectively). Type II collagen expression (Fig. 6C) was also significantly increased in WJCs at all ratios, with the largest increase of almost 9463-fold again being seen at the ratio of 25%:75% WJCs/NPCs. Aggrecan expression in WJCs showed the largest increase in samples with the highest number of NPCs (5974-fold) and decreased as NPC number decreased (640-fold at 50% and 13.7-fold at 25%; Fig. 6B). While coculturing without contact, gene expression levels of aggrecan, type II collagen, and SOX-9 also had significantly increased in all groups (p<0.05) in WJCs with the largest increase being seen at a ratio of 25%:75% WJCs/NPCs (67-fold for aggrecan, 57-fold for type II collagen, and 114-fold for SOX-9), but the increase was smaller than that of cocultures with cell–cell contact (Fig. 6A–C). There were no significant changes in type VI collagen, type I collagen, and versican mRNA expression in WJCs at all ratios (Fig. 6D–F); and gene expressions for type I collagen, type VI collagen, and versican were not significantly up-regulated in WJCs after 7 days in coculture with or without direct contact. It was also evident from the data that cell ratio had effect on gene expression, and the WJC cells had the largest increase of expression of NP marker gene at a ratio of 25%:75% WJCs/NPCs (Fig. 6A–C).
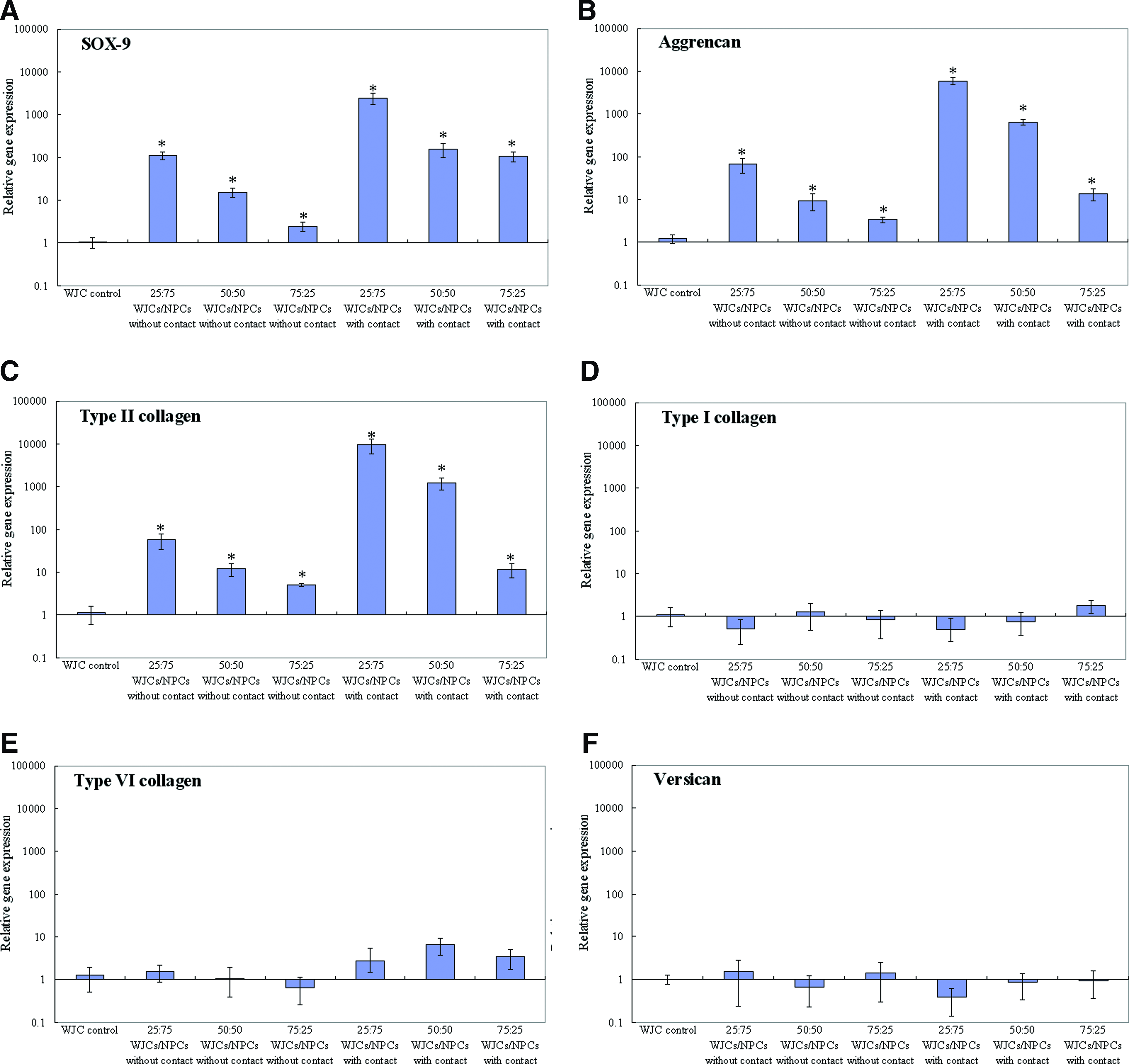
Relative gene expression in WJCs after coculture with or without contact for 7 days. Expression normalized to average of housekeeping genes (GAPDH) and control cells cultured alone.
Discussion
In this study, WJCs were isolated from human umbilical cord Wharton's jelly. WJCs were found to express the surface markers of MSCs and have the activity of telomerase. Further, it was found that coculture of WJCs and NPCs with or without cell–cell contact could enhance NP marker genes expression in WJCs, and cell–cell contact and cell ratios had a positive effect on the differentiation of WJCs. This study suggested that WJCs might differentiate into NP-like cells after being delivered into the NP for managing the IVD degeneration, and could restore extracellular matrix. To the best of our knowledge, this is the first report indicating the feasibility of using WJCs as a potential source of stem cells toward managing IVD degeneration in future.
Human umbilical cord is a postnatal organ discarded after birth, the collection of cells does not require an invasive procedure with mild ethical concerns. Recent year many studies have indicated the pluripotency of MSCs isolated from the human umbilical cord tissue. MSCs have been isolated from several compartments of the umbilical cord such as umbilical cord blood, umbilical vein subendothelium, and Wharton's jelly. MSCs isolated from Wharton's jelly have been induced to form bone, cartilage, and adipose cells,21–24 Compared with adult MSCs such as BMSCs and ADSCs, the WJCs have telomerase activity29,30 and greater ex vivo proliferation capacity and numbers of passages to senescence. 20 The WJCs have higher frequency of colony-forming unit-fibroblast31,32 than BMSCs. 20 Unlike ADSCs, a large numbers of cells could be rapidly initially isolated from Wharton's jelly, thus avoiding the necessity of extensive multiplication and potential epigenetic damage. 33 In our study, we isolate the cells from Wharton's jelly. According to the analysis of flow cytometry, the WJCs express the surface markers CD73\CD105\CD90\CD29\CD166 and HLA-ABC, and do not express the surface markers CD34\CD45 and HLA-DR, which indicate that WJCs display MSC surface markers. CD105, CD73, CD90, CD29, and CD166 are known to characterize MSCs. They do not express hematopoietic stem cell markers such as CD45, CD34, and HLA-DR, which are also lacking in MSCs. 34 These results were supported by Wang et al.'s study, 20 who had proved that the WJCs express significant amounts of MSC markers (CD73/CD105) and could be induced to differentiate to cells of the adipogenic and osteogenic lineages and cardiomyocytes. Our results also showed that WJCs were positive for telomerase expression, which was in agreement with that reported by Mitchell et al. 29 As we know, telomerase is a ribonucleoprotein that replicates telomeres during the S phase of mitosis. 35 Telomerase activity is found in human germ, tumor, and embryonic cell lines and is thought to be responsible for the unlimited capacity for self-renewal of these cell types.36,37 Thus, telomerase activity is often used as a marker for stem cells.
The NP is a semifluid tissue (in early life) that is distinctly different from fibrocartilage tissues or hyaline cartilage. NP contains more type II collagen and proteoglycans and less type I collagen than fibrocartilage tissues or hyaline cartilage. 38 The cells in the collagen fibrous ring produce type I collagen and type II collagen, whereas the NP cells seem to produce only type II collagen.39,40 Proteoglycans are present in a higher amount in the NP than in the fibrous ring, constituting about half the dry weight the NP. It was reported that the proteoglycans to collagens ratio within the NP of young adults was approximately 27:1, whereas the ratio within the hyaline cartilage endplate of the same-aged individuals was about 2:1. 38 Currently, there are no specific markers for NPCs, and the NPCs have often been considered as chondrocyte-like cells, because NP tissue and hyaline cartilage possess similar macromolecules in their extracellular matrix markers. SOX-9 is one of the key regulators of MSC differentiation to chondrocyte-like cells and regulates expression of the type II collagen gene (Col2a1). 41 Aggrecan and type II collagen are the main ECM in NP and hyaline cartilage tissue.
In this study, coculture of human WJCs with NPCs in transwells (with a 0.4-μm pore-size filter insert) without cell–cell contact, and in six-well plates with cell–cell contact were carried out at a variety of different ratios. Gene expressions of the transcription factor SOX-9, aggrecan, versican, the collagen types I, II, and VI showed a significant increase measured by real-time PCR. Our study design is based on previous investigation demonstrating that BMSC differentiation toward NP-like cells in vitro could be induced by cell–cell contact in coculture. 3 What is more in our results is that gene expression significantly increased even without cell–cell contact. However, cells cocultured with contact showed larger increases in gene expression of aggrecan and type II collagen, SOX-9 than cells cocultured without contact, which indicates that direct cell–cell contact is important for WJCs differentiation. The explanation may be that direct cellular contact may enhance cell signaling pathways and expression of specific adhesion molecules controlling differentiation and phenotypic expression of cells. Other possible explanations might be that coculture with contact may induce growth factor expression, in particular TGF-β, insulin-like growth factor-1 (IGF-1), epidermal growth factor (EGF), and platelet-derived growth factor (PDGF), 42 and these growth factors are implicated in matrix production by NPCs. 43 The culture insert we used in the study consists of a PET track-etched membrane with 0.4 μm pores at the bottom, which allow the growth factor secreted by NPCs exchange such as TGF-β, IGF-1, EGF, and PDGF. We have also shown that the cell ratio in coculture affects cell differentiation. In most cases, a ratio of 25:75 WJCs/NPCs was the optimal ratio for stimulating WJC differentiation, possibly because the WJCs received more cellular signals from the greater NPC population.
In the current study, we also took into consideration the influence of CFSE on the gene expression of WJCs. The dye–protein incorporate in labeled cells was retained by the cells throughout development and meiosis. The label was evenly shared between daughter cells after either cell division or cell fusion, and is not transferred to adjacent cells in a population (Fig. 1). CFSE has been shown not to affect cell phenotype, function, or proliferation rate.44,45 Results showed no significant change in gene expression after culturing for 7 days (Fig. 5), thus suggesting that CFSE does not influence the gene expression of WJCs and is a safe and reliable molecule for labeling cells.
Using WJCs for customized cell-based therapies has advantages: adequate cells available from the umbilical cord, no ethical and religious controversies, and avoiding invasive procedure. The most important thing is that human WJCs have low immunogenicity 46 and lower levels of expression of HLA-ABC in addition to the absence of HLA-DR expression 47 that suggests that WJCs might be tolerated in allogenic transplantation which may favor the use of WJCs for allogeneic cell therapy in future. A limitation of the current study was that we did not further confirm NPCs differentiation of WJCs by other methods suggested in the literature. Mwale suggested that different proteoglycans to collagen ratio could be used to distinguish NP cells from chondrocytes. 38 In our study, we only detected the upregulation of gene expression of aggrecan, type II collagen, and SOX-9 in WJCs. Even though this is not enough to confirm whether WJCs were indeed induced to differentiate to NP cells or perhaps to chondrocytes, WJCs showed enhanced ability to produce aggrecan and type II collagen.
In conclusion, it is found that coculturing of WJCs and NPCs could lead to the NP-like cell differentiation of WJCs, which suggests that WJCs might be promising seed cells for cell-based treatment in DDD in future.
Footnotes
Acknowledgments
The authors wish to thank Dr. Li HS for his invaluable help in the English edition of the article and helpful suggestions for the discussion. Thanks are also due to Prof. Gao LR of Navy General Hospital for her invaluable help in isolating WJCs from human umbilical cords, Dr. Yan J and Dr. Yang YF of the Academy of Military Medical Sciences of China for providing real-time PCR, Dr. Yang M of Navy General Hospital for providing access to the flow cytometry, and Dr. Du LY of The School of Life Sciences at Peking University for providing access to the MoFlo high-performance cell sorter.
Disclosure Statement
No competing financial interests exist.