
Abstract
The fibroblast growth factor 2 (FGF-2) is known as pleiotropic cytokine with myoblast proliferative properties. In the present study, we tested the hypothesis that gene transfer of human FGF-2 via transplantation of genetically modified L8-myoblast encapsulated in alginate modulates the skeletal muscle recovery after crush injury in Wistar rats. Therefore, we performed a crush injury to the soleus muscle and transplanted alginate spheres containing myoblasts genetically modified to overexpress human FGF-2 (FGF-2) or a luciferase (LUC) cDNA at the site of injury. Animals that underwent muscle injury without transplantation of alginate spheres served as control (control). At day 4 after trauma the FGF-2 group showed significant higher mean values of cell proliferation (bromodeoxyuridine immunohistochemistry) and significant lower values of cell apoptosis (terminal deoxynucleotidyl transferase nick end labeling histology) compared to animals receiving luciferase-overexpressing myoblasts. At the same time point adiponectin expression (ACRP30 immunohistochemistry) was increased in the FGF-2 group exclusively. The p75NTR expression (p75NTR immunohistochemistry) significantly improved in both the FGF-2 and LUC group compared to the control group. Functional analysis of the injured muscle did not reveal a significant increase of the muscle force in the FGF-2 group compared to the control and LUC group 14 days after injury. In vitro analysis for 14 days of the FGF-2-modified spheres demonstrated at day 7 and day 14 a significant increase of the relative cell count as well as of the relative viable cell count in the FGF-2 myoblast spheres compared to luciferase myoblast spheres. Additionally, the expression of FGF-2 (enzyme-linked immunosorbent assay analysis) and luciferase (chemiluminescence analysis) persisted in vitro for 4 and 14 days, respectively. These results demonstrate that FGF-2-overexpressing myoblasts cannot considerably improve muscle strength but are able to modulate the proliferation as well as the apoptosis of injured muscle tissue mainly by conducting adipogenesis.
Introduction
Depending on the mechanism of trauma, muscle injuries can be subdivided in contusions, strains, and muscle ruptures. In particular, the contusion injuries can be caused by direct impact on the muscle usually caused by contact sports. During a contusion injury, the traumatic force damages directly the muscle tissue and presses the muscle fibers on the underlying bone. As a result the muscle fibers, blood vessels and nerves became irreversibly damaged and hematoma formation occurs. 3
Depending on the injury severity, muscle injuries can regenerate spontaneously without intervention or heal incompletely with scar tissue formation. Although significant scientific efforts were made to better understand the underlying mechanisms of skeletal muscle injury and regeneration, only few satisfactory clinical approaches have been proposed to cure traumatic muscular lesions. 4 Implementation of clinical studies for the muscle injury analysis can be very demanding since there is a high heterogeneity regarding the injury severity and the localization of the lesions. 5 Up to now no sufficient treatment after muscle injuries exists and novel therapeutic strategies are required to enhance muscular regeneration after injury. 6
Growth-promoting agents increase the size of existing and newly regenerating muscle fibers and are used to improve muscle function if administered during the repair process after injury. Till now, several animal studies have been performed after muscle injury, investigating the regenerative capacity of various growth-promoting factors. The fibroblast growth factor-2 (FGF-2) is a member of the multifunctional fibroblast growth factor family and a potent mitogen with anabolic actions for several cell types. 7 Further, FGF-2 is a well-known pleiotropic cytokine that modulates numerous cellular functions, like proliferation, differentiation, survival, adhesion, migration, motility, and apoptosis. 8 Although regenerative actions of FGF-2 on muscle tissue are described in vivo and in vitro, continuous application or high doses of the growth factor are required to produce a sufficient biologic response.
It has been suggested that overproduction of pleiotropic cytokines into the injured tissue via genetically modified cells may represent an attractive alternative to conventional therapeutic strategies. An interesting method that has been already used for tissues repair of muscle, skin liver, and cartilage is to transfect specific cells with genome, which enhances the repair processes. 9 For this purpose, transfection of myoblasts may be an attractive option since this cells are used during the repair process of damaged skeletal muscle. 10
For this purpose, we tested the hypothesis that genetically modified human FGF-2-overexpressing L8-myoblasts encapsulated in alginate spheres can express the transfected genome and modulate muscular regeneration after muscle injury.
Material and Methods
Cell transfection and encapsulation in alginate spheres in vitro
L8-myoblasts were transfected using the nonliposomal lipid formulation FuGENE-6 (Roche) as previously described. 11 Plasmid vectors carried either the Photinus pyralis luciferase gene 12 or a human FGF-2 cDNA 13 under the control of the human cytomegalovirus immediate-early promoter/enhancer. Briefly, endotoxin-free plasmid DNA was complexed with FuGENE 6 (lipid/DNA ratio of 2:1) in Opti-MEM and transferred to subconfluent L8-myoblast monolayers. One day after transfection, cells were trypsinized, washed, and suspended in 1.2% alginate in 0.15 M NaCl at 2×106 cells/mL. The cell suspension was then extruded at a rate of 120 mL/h through a 21-gauge needle (Braun) into a 102 mM CaCl2 solution at room temperature under constant shaking. The resulting alginate-L8-myoblast spheres were allowed to polymerize for 10 min and then washed twice in 0.15 M NaCl followed by two consecutive washes in basal medium. Spheres were placed in Dulbecco's modified Eagle's medium (DMEM) containing 100 U/mL penicillin G, 100 mL/mL streptomycin, and 10% fetal bovine serum (five spheres/mL medium) that was changed three times per week and kept at 37°C in a humidified atmosphere of 10% CO2. Cultured spheres were assessed for cell number and viability at 1, 4, 7, and 14 days after transfection. Single spheres were solubilized by individual incubation in 100 μL 55 mM sodium citrate and 90 mM NaCl (pH 6.8) for 20 min at room temperature. The released cells were counted and their viability assessed using a Neubauer chamber and trypan blue exclusion staining based on four counts per sample. From each time point, two spheres per group were frozen and individually stored at −80°C for subsequent assay of luciferase activity.
Spheres containing luciferase-overexpressing myoblasts (LUC group) served as control to those spheres containing FGF-2-overexpressing myoblasts (FGF-2 group).
In vitro analysis: Cell viability, relative cell count, relative viable cell count, and gene transfer analysis
To elucidate the increase of the initial cell count, the relative cell count per sphere was calculated by dividing for each day the total cell count of each sphere by the average cell count of day 1. The same calculation was performed to determine the relative viable cell count. Relative cell count and relative viable cell count are given in percent.
For analysis of enzymatic activity, luciferase protein was extracted from alginate spheres cultured in vitro or retrieved from the muscle tissue in vivo as described by the manufacturer (Promega). Frozen tissue was thawed, suspended in 100 μL Promega lysis buffer, homogenized, frozen and thawed three times, and centrifuged for 5 min at 10,000 g. The supernatant was transferred to a new microfuge tube and luciferase activity was assayed by adding 5 μL of tissue extract to 25 μL of luciferase substrate from one single lot using a Berthold Lumat LB 9507 luminometer (Berthold Analytical). Sample light units (relative light units, RLU) were recorded in duplicates within 5 s after substrate addition. For the in vitro FGF-2 protein analysis cell layers were washed twice, and the medium was replaced with serum-free DMEM. After 48 h, the conditioned medium was collected, centrifuged to remove cell debris, and stored at −80°C. Recombinant hFGF-2 production was measured by ELISA (R&D Systems) with a detection limit of 3 pg/mL. Data were given for the luciferase transgene expression as RLU per μg total protein and for the FGF-2 transgene expression as pg per 104 viable cells.
Animal model and experimental groups
For the in vivo experiments 42 male Wistar rats (300–325 g body weight [bw]; Charles River Laboratories) were used and were kept on water and standard laboratory chow ad libitum. The experiments were conducted in accordance with the German legislation on protection of animals and the National Institutes of Health Guide for the Care and Use of Laboratory Animals (Institute of Laboratory Animal Resources, National Research Council).
Under pentobarbital sodium anesthesia (55 mg/kg bw i.p., Narcoren; Merial) the right and left lower limb was shaved and disinfected with povidone-iodine. Through a 2 cm posterolateral longitudinal incision of the skin and the underlying fascia from the lateral gastrocnemius head to the Achilles tendon, the soleus muscle was mobilized on both lower extremities. 14 The blunt injury was induced via an instrumented clamp, which allowed a standardized force application (crush force between forceps and muscle tissue in the 2×lock position 14 : 25 N) for 10 s (DMC PLUS; HBM Hottinger Baldwin Messtechnik GmbH). The muscle was manually clamped seven times over its complete length with exception of the entry point of the supplying neurovascular structures, which arise from the mid part of the medial gastrocnemius. After injury induction animals were transplanted with alginate spheres on the site of injury containing genetically modified myoblasts overexpressing luciferase. The transfected cells were encapsulated in alginate spheres and transplanted into the site of injury 24 h after encapsulation. The spheres on each leg were placed between the gastrocnemius and the soleus muscle as well as between the extensor digitorum longus and the soleus muscle. Same procedure was repeated for animals using FGF-2-overexpressing alginate spheres. Next, the superficial muscle and the skin were closed using 4-0 vicryl sutures (Ethicon GmbH). During the procedure body temperature was maintained at 36°C–37°C by means of a heating plate. Upon transplantation animals were allowed to awake from anesthesia and remained in single cages with free access to water and laboratory chow. For analysis of muscle cell proliferation animals received at day 2 a single intraperitoneal injection of 50 mg/kg bromodeoxyuridine (BrdU). At day 4 after injury induction all rats were euthanized by pentobarbital sodium overdose and muscle specimens of the right and left lower extremity (containing the injured soleus muscle as well as parts of the noninjured gastrocnemius muscle and extensor digitorum longus muscle) were harvested for chemiluminescence analysis or histology/immunohistochemistry respectively.
To elucidate to impact of FGF-2 myoblast spheres in the muscle regeneration, we used two in vivo approaches. In the first approach we elucidated the muscle regeneration using histology, immunohistochemistry, and chemiluminescence analysis in 24 animals. For this purpose the soleus muscle was bilaterally traumatized to gain enough tissue for the previously mentioned methods. Subsequent analysis was performed at day 4 after injury. Three experimental groups of animals were included in this set of experiments: control group, that is, animals that underwent bilateral crush injury of the soleus muscle (n=8); LUC group, that is, animals that underwent crush injury of the soleus muscle bilaterally and subsequent transplantation of four alginate spheres containing luciferase-overexpressing myoblasts into each lower extremity (n=8); FGF-2 group, that is, animals that underwent crush injury of the soleus muscle bilaterally and subsequent transplantation of four alginate spheres containing FGF-2-overexpressing myoblasts into each lower extremity (n=8).
In the second approach of our in vivo analysis we evaluated the functional restoration of the muscle and measured the muscle strength in additional 21 animals. For this purpose only the left soleus muscle was traumatized and the subsequent muscle strength measurement was performed at day 14 after muscle injury. The right soleus muscle was intentionally not manipulated in order to express the muscle force of the injured left soleus muscle in percentage of the corresponding values of the contralateral noninjured muscle. Three experimental groups of animals were included in this part of our study: control group, that is, animals that underwent crush injury of the left soleus muscle (n=7); LUC group, that is, animals that underwent crush injury of the left soleus muscle and subsequent transplantation of four alginate spheres containing luciferase-overexpressing myoblasts into the traumatized lower extremity (n=7); FGF-2 group, that is, animals that underwent crush injury of the left soleus muscle and subsequent transplantation of four alginate spheres containing FGF-2-overexpressing myoblasts into the traumatized lower extremity (n=7).
In vivo analysis: Histology, immunohistochemistry, chemiluminescence analysis, and muscle strength measurement
To elucidate the impact of the transplanted myoblasts spheres we analyzed histologically the injured muscle (i.e., soleus muscle) as well as the noninjured muscle (i.e., gastrocnemius muscle and extensor digitorum longus muscle). For this purpose the dorsal muscle compartment of the left hind limb, containing the gastrocnemius, extensor digitorum longus, and soleus muscle was surgically excided. Muscle tissue was fixed in 4% phosphate buffered formalin for 2–3 days, straight embedded in paraffin, and strictly cut in a longitudinal fashion from proximal to distal. By that procedure we were able to detect part of the noninjured muscle tissue (gastrocnemius and extensor digitorum longus muscle) and part of the injured muscle tissue (soleus muscle) in the histological/immunohistochemical sections. Approximately 30 consecutive observation fields were counted in the noninjured and injured muscle tissue, respectively, and only one investigator (I.S.) evaluated the histology and immunohistochemistry.
For immunohistochemical demonstration of BrdU, tissue specimens collected on poly-L-lysine-coated glass slides were treated by microwave for antigen unmasking. Monoclonal mouse anti-BrdU (1:50; Dako Cytomation) was used as primary antibody and incubated for 18 h at 4°C. After equilibrating to room temperature sections were incubated with a horseradish peroxidase-conjugated secondary antibody for 30 min according to the manufacturer's instructions (Dako Cytomation). 3,3′-Diaminobenzidine was used as chromogen. Sections were counterstained with hemalaun and examined by light microscopy (BX 51; Olympus). Using a ×40 objective (numerical aperture 0.65), BrdU-positive cells were counted within the injured and noninjured muscle tissue (∼30 consecutive observation fields). BrdU-positive cells were given as cells per mm2 muscle tissue.
In addition, by performing a double immunohistochemistry for laminin and BrdU we differentiated in the penumbra area of the soleus muscle (i.e., the area of the muscle that still reveals viable myocytes) the proliferating cell types. Thereby, we identified proliferating satellite cells due to their localization beneath the laminin-positive basal lamina. BrdU-positive cells, which were found localized outside the basal lamina, were identified as interstitial cells and BrdU-positive cells in direct vicinity to blood cell-containing lumen were estimated as proliferating endothelial cells. Using a ×40 objective (numerical aperture 0.65) individual types of cells were counted and given as number of cells per 10 myocytes (∼6 consecutive observation fields). Herein, a rabbit polyclonal laminin antibody (1:50; 1 h incubation at 37°C; Dako Cytomation) was used with 3,3′-diaminobenzidine and fuchsin as chromogens.
To determine apoptotic cell death, terminal deoxynucleotidyl transferase nick end labeling (TUNEL) analysis was performed using a commercially available in situ apoptosis detection kit (ApopTag, S7100; Chemicon). Using a ×40 objective TUNEL-positive cells were counted within the injured and noninjured muscle tissue (∼30 consecutive observation fields), as described above. TUNEL-positive cells were given as cells per mm2 muscle tissue.
To look in more detail on the effect of muscle injury on angiogenesis, we performed a CD31 immunohistochemistry. For demonstration of endothelial cell marker, CD31 tissue specimens collected on poly-
For immunohistochemical demonstration of the satellite cell associated p75 neurotrophin receptor (p75NTR) expression in the muscle tissue was identified using an anti-p75NTR (1:100, ab-8874; Ab-cam) antibody. 15 3-amino-9-ethylcarbazole was used as chromogen. Using a ×40 objective, p75NTR-positive cells were counted within the injured and noninjured muscle tissue (∼30 consecutive observation fields) and data were given as cells/mm2.
Immunohistochemical staining was performed for identification of adiponectin (ACRP30) as an intermediate marker for adipogenesis. ACRP30 expression in the muscle tissue was identified using an anti-ACRP30 (1:200, AF-3100; R&D Systems) antibody. 3,3′-Diaminobenzidine was used as chromogen. Using a ×40 objective ACRP30-positive cells were counted within the injured and noninjured muscle tissue (∼30 consecutive observation fields) and data were given as cells/mm2.
The immunohistochemical demonstration of FGF receptor 1 (FGFR1) expression in the muscle tissue was identified using an anti-FGFR1 (1:200, F4305-02K; Biomol) antibody. 3,3′-Diaminobenzidine was used as chromogen. Using a ×40 objective, FGFR1-positive cells were counted within the injured and noninjured muscle tissue (∼30 consecutive observation fields) and data were given as cells/mm2.
For in vivo analysis of the luciferase transgene expression, the luciferase activity was measured by chemiluminescence analysis. For this purpose we used the muscle specimens from the right hind limb and the tissue was processed as described in the previous section In vitro analysis. Data were given as RLU per mg total protein.
For in vivo assessment of muscle strength animals were re-anesthetized at day 14 postinjury. After bilateral exposure of the sciatic nerve and the soleus muscle the Achilles tendon was cut and the lower extremity was fixed into the muscle force-measuring device (Experimetria). The sciatic nerve was subsequently stimulated with 9 mA/75 Hz bipolar five times, 0.1 s (eight periods) with 5 s interval between the pulses. After this fast twitch stimulation protocol maximal strength was measured by application of 9 mA/75 Hz pulses for five times, 3 s each with 5 s intervals, reaching tetany in all cases. Contraction forces under fast twitch and tetanic stimulation were analyzed by calculating the mean of the maximal values from first five consecutive contractions and given as percentage of the corresponding values of the contralateral noninjured muscle.14,16,17 After completion of muscle strength measurements experiments were terminated.
Statistical analysis
All data are expressed as means±standard error of the mean. Before statistical analysis the population normality of each group was tested. Upon passed normality test, differences between groups were assessed using one-way analysis of variance (ANOVA), followed by the Holm-Sidak method for pairwise multiple comparison. By failed normality test Kruskal–Wallis one-way ANOVA on ranks was performed followed by the Tukey test for pairwise multiple comparison. Statistical significance was set at p<0.05. Statistics were performed using the software package SigmaStat (Jandel Corporation).
Results
In vitro results
Gene transfer of a human FGF-2 cDNA in myoblasts encapsulated in alginate resulted in efficient and sustained production of FGF-2 for at least 4 days in vitro. Alginate spheres containing LUC-transfected myoblasts demonstrated luciferase activity above the background activity for at least for 14 days in vitro (Table 1).
Spheres containing luciferase-overexpressing myoblasts (LUC) served as control to those spheres containing FGF-2-overexpressing myoblasts (FGF-2). The spheres were placed in well plates containing physiological medium and subsequent assessments were performed on days 1, 4, 7, and 14 after transfection (n=2 myoblast spheres per time point and group). Data are given as means±SEM; t-test.
p<0.05 versus LUC.
FGF-2, fibroblast growth factor-2; RLU, relative light units; SEM, standard error of the mean.
In vitro analysis of alginate spheres at day 7 post-transfection revealed that overexpression of FGF-2 stimulated myoblast proliferation in alginate spheres as indicated by the 2.9-fold amplification of the relative cell count in FGF-2 spheres compared to LUC spheres. At the same time point the relative viable cell count per sphere was increased by over 413% in the FGF-2 group compared to a descent increase in the control group. After 14 days in vitro, proliferation of myoblasts in alginate spheres persisted in FGF-2 spheres as reflected in the 2.7-fold increase of the relative cell count and the 3.4-fold increase of the relative viable cell count compared to a 0.8-fold increase in LUC spheres (p<0.05). The cell viability was not influenced by the FGF-2 overexpression as no differences between the groups and time points were noted (Table 1).
In vivo results
All animals awoke from anesthesia without complications. There were no signs of uncomfortableness or illness. In addition, injured hind limbs did not show macroscopic signs of inflammation, such as swelling, skin redness, hyperthermia, or wound secretion.
Luciferase activity was present in muscle tissue retrieved from animals receiving LUC spheres at 4 days after transplantation, compared to the background activity in muscle tissue retrieved from animals receiving FGF-2 spheres (LUC: 187.4±101.9 vs. FGF-2: 8.0±2.7 RLU per mg total protein; n=5 per group).
We checked all tissue specimens for microscopic signs of inflammation (e.g., inflammatory cell accumulation, foreign body reactions, and edema formation) and could confirm the absence of an alginate-induced inflammatory process.
Using TUNEL staining of muscle tissue specimens, we verified apoptotic cell death upon muscle injury. The quantitative analysis revealed very low values of cell apoptosis on the noninjured muscles reaching values between 0.5 and 1.1 apoptotic cells per mm2. In contrast, muscle injury caused a 40-fold increase of apoptotic cells 4 days after injury. This massive increase of apoptotic cell death after injury could significantly be reduced upon transplantation of FGF-2-overexpressing myoblast spheres to less than 65% (Fig. 1). Transplantation of luciferase-overexpressing myoblast spheres could also reduced the cell apoptosis of the injured muscle, but in a less effective manner than FGF-2 myoblast spheres (25% reduction compared to control animals).
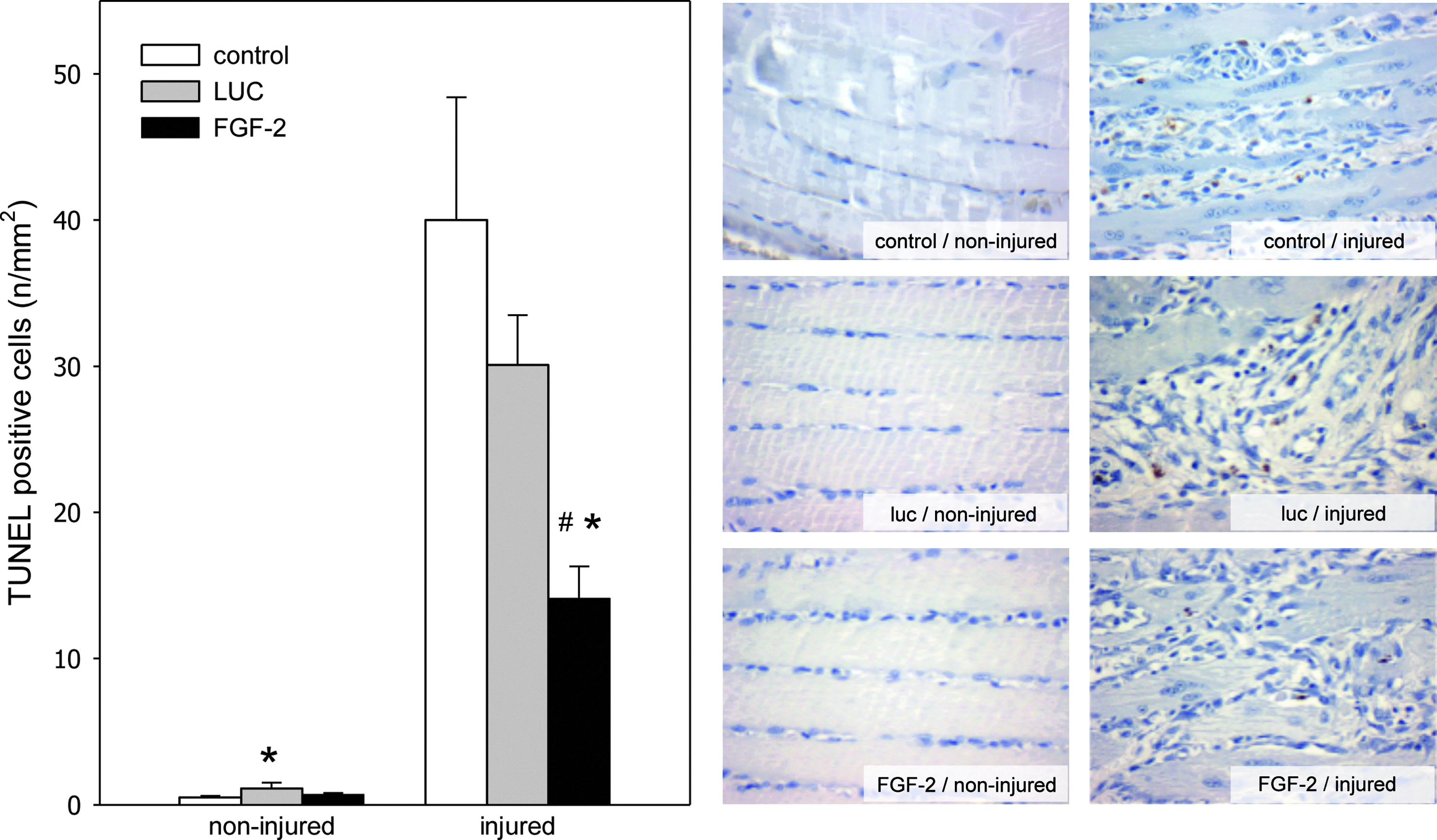
Quantitative analysis of terminal deoxynucleotidyl transferase nick end labeling staining (left panel) as well as representative light microscopic images (right panel; ×400 magnification) of injured and noninjured muscle tissue. Animals underwent a crush injury to the soleus muscle and subsequent transplantation of luciferase-overexpressing myoblast spheres (LUC; gray bars; n=8 animals) or FGF-2-overexpressing myoblast spheres (FGF-2; black bars; n=8 animals). Control animals (control; white bars; n=8 animals) underwent injury to the soleus muscle without transplantation of myoblast spheres. Analysis was performed at 4 day after injury and data are given as means±SEM; one-way ANOVA: *p<0.05 versus control, #p<0.05 versus LUC. FGF-2, fibroblast growth factor-2; SEM, standard error of the mean; ANOVA, analysis of variance.
To further analyze the muscle repair we determined angiogenesis in muscle tissue. Staining of expression of the endothelial cell marker CD31 served as a read out for angiogenesis at the healthy muscle as well as at the site of injury. Quantitative analysis of the noninjured muscle tissue displays no difference of microvessel density between control, FGF-2 group, and LUC group (between 1.3 and 2.1 CD31-positive microvessels/mm2). A high density of microvessels after muscle injury, presenting with CD31-positive endothelial lining, was observed after muscle injury. Implantation of luciferase-overexpressing myoblast spheres did not influence the microvessel density. Almost equal levels of CD31-positive microvessels were observed between the control and LUC group 4 days after injury. In contrast, FGF-2-overexpressing myoblast spheres induced an additional twofold increase of microvessels, though the differences among the groups were not large enough to reach statistical significance (p=0.053) (Fig. 2).
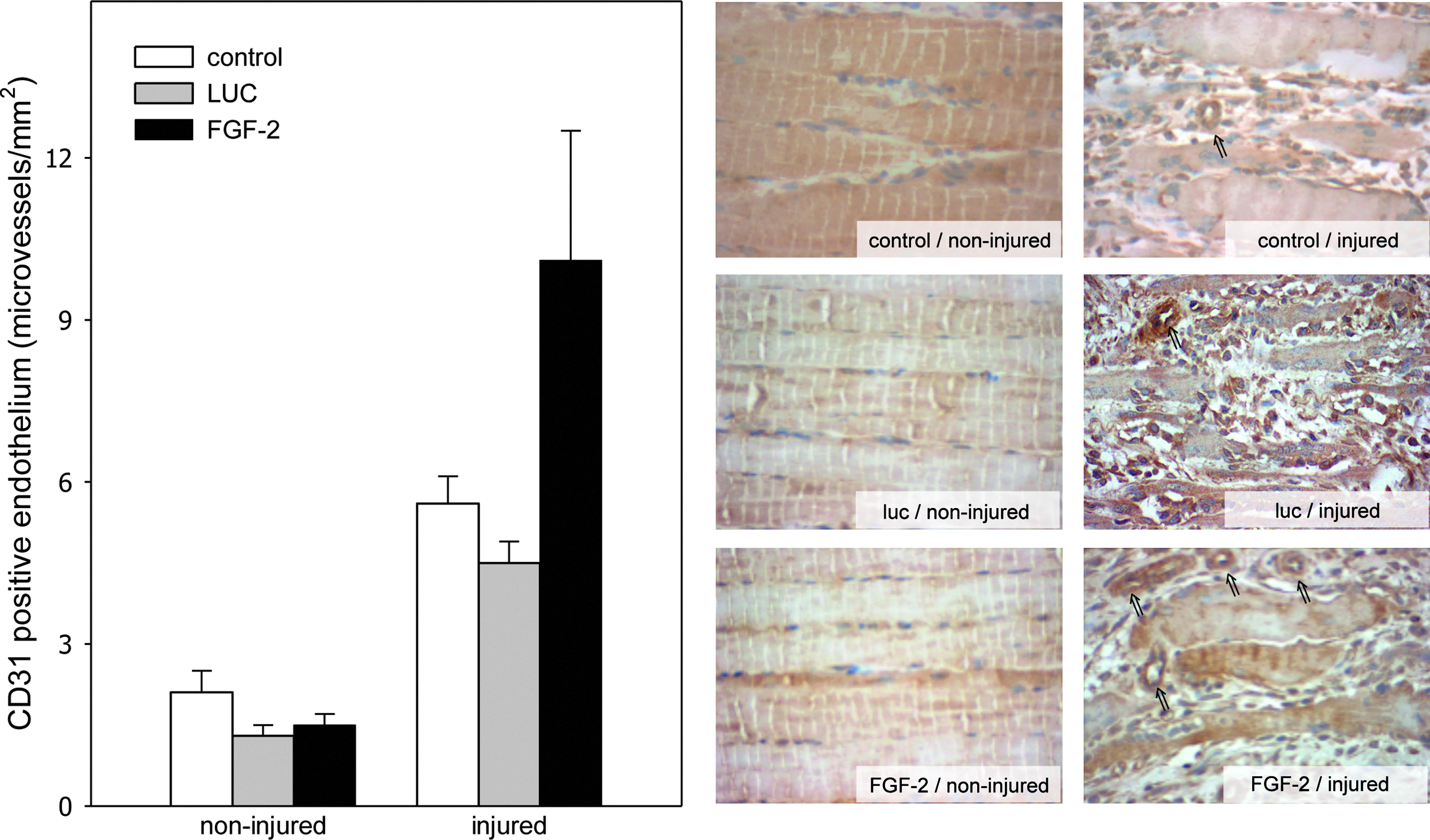
Quantitative analysis of CD31-positive microvessels (left panel) as well as representative light microscopic images for CD31-positive microvessels (right panel; arrows; ×400 magnification) of injured and noninjured muscle tissue. Animals underwent a crush injury to the soleus muscle and subsequent transplantation of luciferase-overexpressing myoblast spheres (LUC; gray bars; n=8 animals) or FGF-2-overexpressing myoblast spheres (FGF-2; black bars; n=8 animals). Control animals (control; white bars; n=8 animals) underwent injury to the soleus muscle without transplantation of myoblast spheres. Analysis was performed at 4 day after injury and data are given as means±SEM; one-way ANOVA.
Muscle injury is followed by a massive increase of cell proliferation. To assess this process, the proliferating cells were identified via BrdU incorporation into the DNA. The subsequent quantitative immunochistochemical analysis between control, FGF-2, and LUC group revealed normal cell proliferation and no difference regarding the BrdU-positive cells in the noninjured muscles. On the contrary, muscle injury resulted in a notable increase of cell proliferation (74 BrdU-positive cells/mm2 in the control group). Transplantation of luciferase-overexpressing myoblast spheres does not influence the proliferative activity of the injured muscle (p>0.05 between the control and LUC group). Traumatized muscles majorly benefit from the FGF-2 sphere transplantation by a significant 1.8-fold increase of skeletal muscle cell proliferation at day 4 after trauma when compared to animals with luciferase myoblast spheres (Fig. 3).
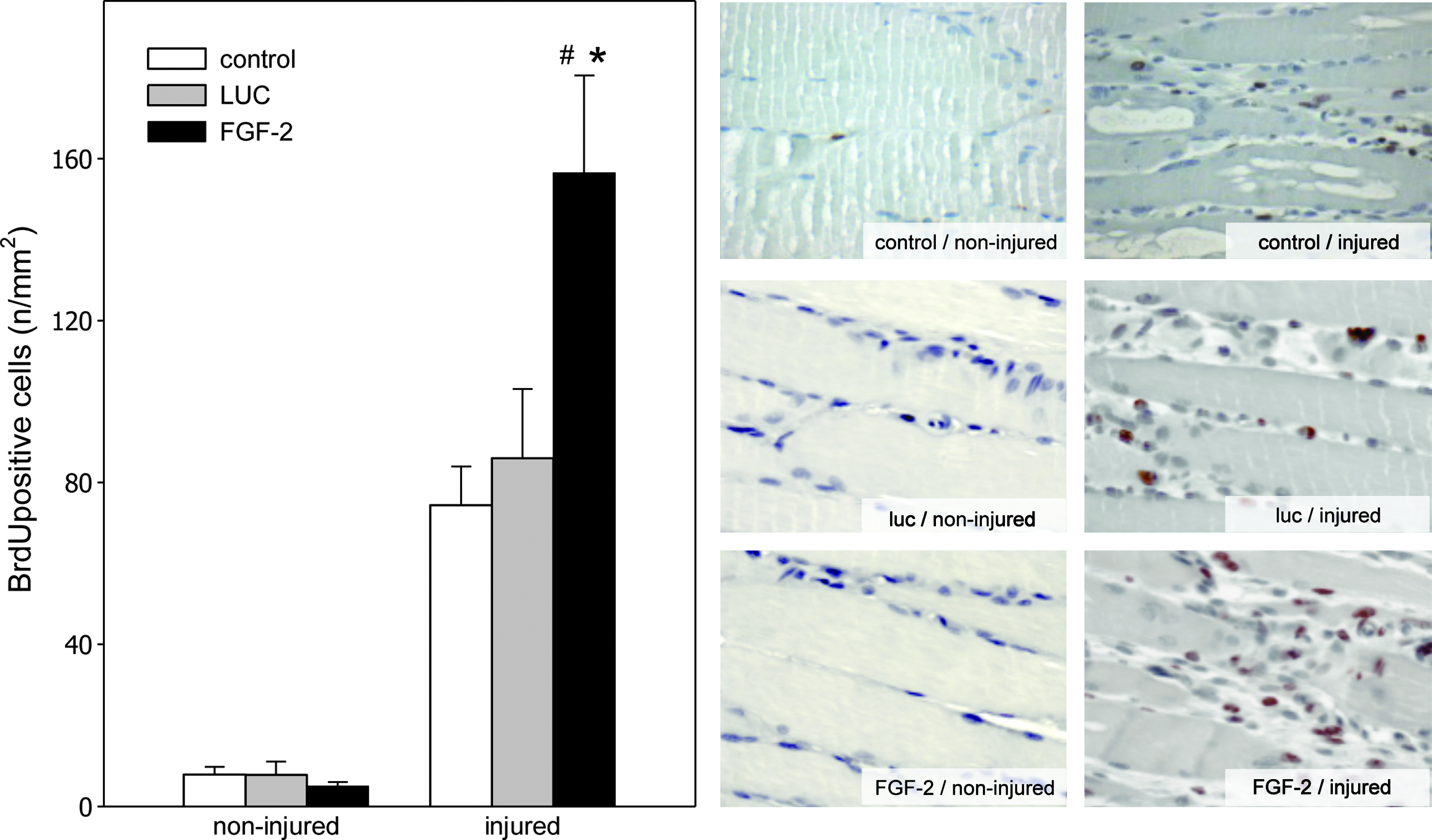
Quantitative analysis of BrdU staining (left panel) as well as representative light microscopic images (right panel; ×400 magnification) of injured and noninjured muscle tissue. Animals underwent a crush injury to the soleus muscle and subsequent transplantation of luciferase-overexpressing myoblast spheres (LUC; gray bars; n=8 animals) or FGF-2-overexpressing myoblast spheres (FGF-2; black bars; n=8 animals). Control animals (control; white bars; n=8 animals) underwent injury to the soleus muscle without transplantation of myoblast spheres. Analysis was performed at 4 day after injury and data are given as means±SEM; one-way ANOVA: *p<0.05 versus control, #p<0.05 versus LUC. BrdU, bromodesoxyuridine.
In addition, we differentiated the proliferating cells in injured muscle tissue using the BrdU/laminin double immunochistochemistry. Qualitative analysis on day 4 after injury revealed in the FGF-2-overexpressing myoblast spheres transplanted animals a higher ratio of proliferating satellite cells and endothelial cells compared to the luciferase-overexpressing myoblast sphere–transplanted animals. In addition, a similar increase of interstitial cells proliferation was observed 4 days after trauma (Table 2).
Animals underwent a crush injury to the soleus muscle and subsequent transplantation of luciferase-overexpressing myoblast spheres (LUC; n=8 animals) or FGF-2-overexpressing myoblast spheres (FGF-2; n=8 animals). Control animals (control; n=8 animals) underwent injury to the soleus muscle without transplantation of myoblast spheres. Analysis was performed at 4 day after injury. Data are given as means±SEM; one-way analysis of variance.
Analysis of the FGFR1 showed low receptor expression in noninjured tissue. Trauma of the soleus muscle caused a massive upregulation of the FGFR1 receptor as shown in the control group. Remarkably, the receptors' expression was decreased after myoblast transplantation not only in the FGF-2 group but also in the LUC group (Fig. 4a). Additionally, expression of FGFR1 was not influenced by FGF-2-overexpressing myoblasts upon injury. Thus, both muscle injury and transplantation of myoblast spheres seem to influences the expression FGFR1.
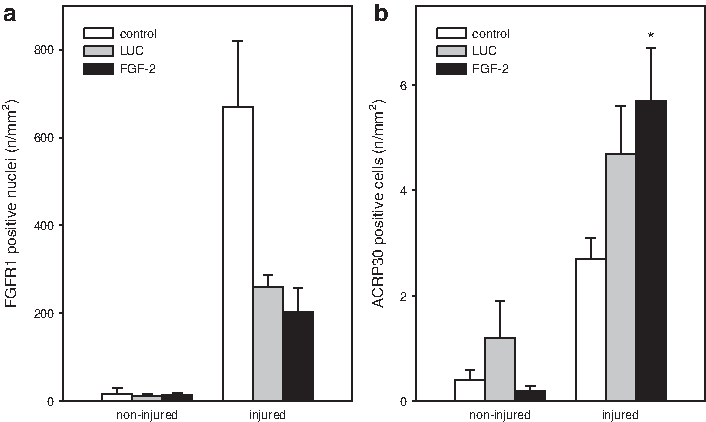
Quantitative analysis of FGFR1
The intermediate marker for adipogenesis ACRP30 was found increased after muscle injury and transplantation of luciferase-overexpressing myoblast spheres, with the vast majority of the cells being interstitially localized. Additional expression of ACRP30 was evident in the FGF-2 group that resulted into a significant difference in extent when compared to the control group (Fig. 4b). This observation is indicative for a post-traumatic adipogenic differentiation of the injured tissue, which is mediated by FGF-2 overexpression.
To further characterize the satellite cells we quantified p75NTR-expressing cells. Whilst low values of p75NTR-positive cells were identified between the groups in the noninjured muscle, increased levels of p75NTR receptor-expressing cells were observed after injury. The control group expressed on average the p75NTR receptor on 70 cells/mm2. Transplantation of FGF-2 and luciferase transfected myoblasts significantly stimulated the proliferation of satellite cells, thereby increasing the ratio of p75NTR-positive cells compared to the corresponding values of the control group (Fig. 5a).
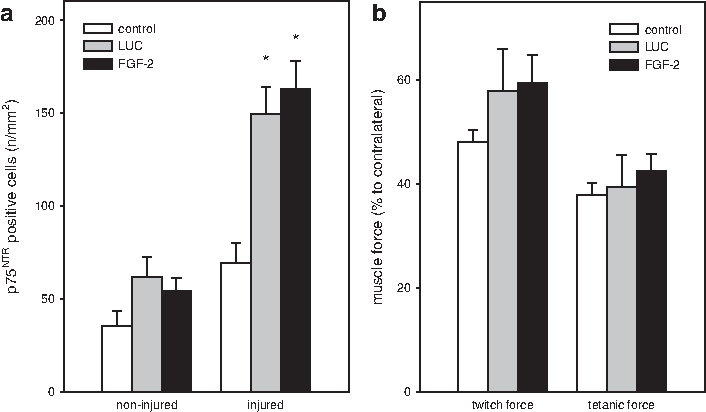
Quantitative analysis of p75NTR staining at day 4 after injury (n=8 animals per group) of injured and noninjured muscle tissue
Functional analysis of muscle strength revealed 14 days after muscle injury a 1.2-fold increase of twitch force in the FGF-2 group compared to the control group. Measurement of the tetanic muscle force in the FGF-2 and LUC group reached values similar to that of the control group (Fig. 5b). Interestingly, the strength of the injured muscle was not different upon transplantation of FGF-2 or luciferase transfected myoblasts spheres 14 days after injury.
Discussion
The data indicate that FGF-2-transfected myoblasts embedded in alginate spheres release functionally active FGF-2 protein for at least 4 days and that the secreted factor selectively stimulates myoblast proliferation in vitro. Luciferase-transfected myoblasts demonstrate no impaired viability when encapsulated in alginate spheres over a 14-day time period in vitro. The data also show that luciferase activity in LUC-transfected myoblasts encapsulated in alginate spheres is present for at least 14 days in vitro and 4 days in vivo. This suggests that the transplanted myoblasts survive for at least 4 days after intramuscular transplantation in vivo. Present in vivo data demonstrate that transplantation of myoblast spheres increases the number of p75NTR-positive cells and downregulate expression of FGFR1. FGF-2-overexpressing myoblast spheres induce an adipogenic differentiation, promote muscle proliferation, and reduce muscle apoptosis of muscle cells in an FGFR1-independent way. Finally, this increased cellular turnover does not majorly improve the mechanical characteristics of the injured muscle tissue and is not accompanied by a significant increase of muscle strength 14 days after injury.
Methodological considerations
The systemic delivery of recombinant gene products using cells encapsulated in alginate spheres has been shown to be feasible in various animal models. 18 Alginate hydrogels have a wide applicability as biomaterials and are used extensively for cell encapsulation, cell transplantation, and tissue engineering applications. 19 In vivo experiments have shown that alginate sphere cell transplantation ensures the biocompatibility, the long-term cell integrity, and the cell function. 20 Furthermore, natural hydrogels like alginate gel are commonly applied as cell-entrapping materials due to their outstanding biocompatibility and mild gelling conditions. 21 Current literature demonstrates that alginate spheres have excellent transplantation potential when used with growth factor-producing cells22–25 or cells with enhanced drug metabolism activity. 26 In line with this, alginate spheres have further been used as scaffolds for tissue engineering or as delivery vehicles for drugs and provide thereby a wide applicability as biomaterials. 27 Therefore, cell encapsulation technology represents an attractive approach to produce and deliver biologic active compounds to injured tissues and pathologic conditions gene transfer approaches offer a potential treatment to a variety of musculoskeletal diseases. Although Wolff et al. 28 reported that injection of naked plasmid DNA into myofibers is possible, the therapeutic use of naked plasmid DNA is impeded due to its high extracellular instability, poor cellular entry, intracellular degradation in endolysosomes, and reduced effective transport into the nucleus. 29 High levels of transgene expression in myoblasts can be achieved by using retroviral vectors, 30 which are efficient vehicles to deliver genes into skeletal muscles. 31 Although adenovirus-mediated gene transfer into adult skeletal muscle is difficult due to low transfection efficiency, 32 various studies demonstrated that adenoviral vector delivery through FGF-2 receptors not only enhances cellular transduction but also increases the vivo efficacy.33–35 Furthermore, adenoviral-mediated gene transfer using the FGF-2/FGFR1 system is an effective method for gene delivery to increase the potency of gene expression.36,37 However, the use of viral vectors is complicated, as adenoviral vectors do not allow sustained transgene expression and are most likely to activate host immune responses in vivo, retroviral vectors carry the risk of insertional mutagenesis, whereas the major issue with lentiviral vectors (a subfamily of retroviral vectors derived from the human immunodeficiency virus) is their potential to generate replication-competent viruses and to induce oncogenesis. Nonviral vectors are an alternative because they do not elicit a detectable immune response, are less expensive to manufacture, and can carry large DNA plasmids.31,38 In particular, the FuGene6 transfection system efficiently transfects L8 cells11,39 with minimal cell toxicity, is relatively easy to use, 40 and mediates levels of transgene expression comparable to those resulting from high-titer retroviral gene transfer. 11
The optimal time point for muscle strength analysis was chosen based on our experience from previous studies using the identical injury model,14,16 where we measured the muscle force development and analyzed histologically the muscle regeneration over a total observation time of 42 days. Thereby, we could observe that the muscle force did not drastically improve beyond 14 days. Thus, we refrained in the present study to perform additional experiments in animals at day 42 and finalized the observation period at day 14, representing the ideal time point for muscle strength analysis.
Cellular events after muscle injury and transplantation of FGF-2-overexpressing myoblasts
According to current literature exogenous administration of FGF-2 does directly stimulate the muscle regeneration after injury. FGF-2 enhances the number of proliferating cells by facilitating the recruitment of additional satellite cells from the quiescent state. 41 Furthermore, it appears that FGF genes trigger proliferative signals, anti-apoptotic pathways, as well as signals required for angiogenesis that other locally applied growth factors could not provide. Additionally, it is known that FGF-2 significantly improves muscle recovery and increased muscle mass after denervation injury. 42 Iwai-Kanai et al. reported for cardiac myocytes that the anti-apoptotic action of FGF-2 is mediated by the mitogen-activated protein kinase-1-extracellular signal-regulated kinases (MEK-1-ERK) pathway 43 and couples intracellular responses to the binding of growth factors to cell surface receptors. Especially, satellite cell proliferation seems to be mediated via the MEK1/2–ERK1/2 signaling pathways. 44 It has been shown that FGF-2 is an important factor for limb development and boosts muscle regeneration after injury. 45 FGF-2-expressing fibroblasts were able to induce complex regenerative procedures like arteriogenesis or myogenesis in injured ischemic muscles. 46 Angiogenic actions of FGF-2 were also described, 7 and furthermore it is known that FGF-2 is normally expressed in muscle during limb ischemia. 47 In this context it has been implicated that p38 is an essential regulator of FGF-2-driven angiogenesis 48 and that angiogenesis is mediated via the mitogen-activated protein kinase pathway. 49 FGF-2 not only appears to act at different stages of the satellite cell cycle, but also modulates the proliferation of satellite cells and enhances the differentiation of precursor cells toward a myogenic determination.50,51
Various cell types like endothelial cells and fibroblasts contribute to the regeneration of the damaged muscle tissue comparably as cell proliferation does. 52 Especially proliferation of satellite cells initiates the regeneration of muscle tissue and forms new multinucleated myofibers that fuse with the damaged myofibers. 53 In previous studies we could observe that increased cell proliferation in combination with decreased cellular apoptosis maintains the cellular balance in the muscle causing over a longer period a recovery of muscle function and results into an enhanced muscle strength.16,17 In accordance with current literature we know that p75NTR is downregulated during myogenic differentiation 15 and that the FGF as well as his receptor-signaling pathway is an important upstreaming regulator for the p75NTR system. 54 Furthermore, a reverse correlation between myoblast fusion into myotubes and p75NTR expression is observed in myoblast cell lines. 55 Correspondingly, we assume that the increased levels of p75NTR expression we observed were associated with reduced myogenic differentiation and thus absence of significant functional restoration of the injured muscle. Consecutively, the increased p75NTR expression may have downregulated the FGFR1 expression. In addition, the cellular events upon FGF-2 and luciferase transplantation of myoblasts spheres were independent from FGFR1 expression.
While adipogenesis occurs in muscle tissue during the first days after injury, 56 there is evidence that fibrogenic and adipogenic progenitor muscle cells expand upon damage, providing a source of pro-differentiation signals for proliferating myogenic progenitors. 57 In support to our data and in accordance to previous reports, it is known that stem cells differentiate after FGF-2 exposure toward an adipogenic lineage. 58 Although we did not differentiate the ACRP30-positive cells it is possible, though very speculative, to assume that the ACRP30-positive cells we observed were of stem cell origin. Two aspects, namely, (1) the interstitial localization and (2) the small total cell number, are in support of this assumption.
A matter of debate might be the potential development of an alginate-induced inflammation. While transplantation of alginate spheres induces local leukocyte infiltration, the extent of the inflammation is influenced by the alginate composition. 59 Studies have shown that FGF-2 potentiates leukocyte recruitment 60 and that high transfection efficacy of FGF-2 augments this inflammatory process. Although leukocytes can release high amount of cytolytic and cytotoxic substances, 61 they coincidentally seem to be essential for muscle repair by removing tissue debris and by activation of satellite cells. 62 Due to the absence of signs of inflammation, an alginate-induced inflammatory process seems to be of minor or even no importance. This interpretation is further underscored by previous findings of our group, where FGF-2-overexpressing chondrocytes encapsulated in alginate spheres did not induce synovial inflammatory response when transplanted to cartilage defects of rabbit knee joints. 24
The efficiency of transgene delivery and expression by nonviral delivery systems in muscle has been improved significantly. FGF-2-transfected myoblast encapsulated in alginate spheres show a significant biologic action by promoting the proliferation reducing apoptosis and enhancing the adipogenic differentiation. These actions resulted in increased musclular regeneration but did not improve the muscle strength. Nevertheless, additional studies are needed to elucidate further effects of myoblast spheres on the injured muscle and to compare these results with conventional muscle trauma therapies.
Footnotes
Acknowledgments
The authors thank Berit Blendow, Doris Butzlaff, Dorothea Frenz, and Maren Nerowski (Institute for Experimental Surgery, University of Rostock) for excellent technical assistance.
Disclosure Statement
No competing financial interests exist.