
Abstract
Dental follicle cells (DFCs) are a heterogeneous population that exhibit a variety of phenotypes. However, it remains unclear whether DFCs can maintain stem cell characteristics, or mediate tissue-regeneration to form single or complex tissues in the periodontium, after long-term culturing. Therefore, DFCs were isolated from human impacted molars (HIM-DFCs), passaged >30 times, and then evaluated for their heterogeneity and multipotential differentiation. Morphology, proliferation, epitope profile, and mineralization characteristics of clones derived from single HIM-DFCs in vitro were also assayed. HIM-DFCs (passage #30) were found to be positive for the heterogeneous markers, Notch-1, stro-1, alkaline phosphomonoesterase (ALP), type I collagen (COL-I), type III collagen (COL-III), and osteocalcine. Moreover, passage #30 of the HDF1, 2, and 3 subclone classes identified in this study were found to express high levels of the mesenchymal stem cells markers, CD146 and Stro1. HDF3 subclones were also associated with the strongest ALP staining detected, and strongly expressed osteoblast and cementoblast markers, including COL-I, COL-III, bone sialoprotein (BSP), and Runx2. In contrast, HDF1 subclone analyzed strongly expressed COL-I and COL-III, yet weakly expressed BSP and Runx2. The HDF2 subclone was associated with the strongest proliferative capacity. To evaluate differentiation characteristics in vivo, these various cell populations were combined with ceramic bovine bone and implanted into subcutaneous pockets of nude mice. The 30th passage of subclone HDF1 and 3 were observed to contribute to fiber collagens and the mineralized matrix present, respectively, whereas HDF2 subclones were found to have a minimal role in these formations. The formation of a cementum-periodontal ligament (PDL) complex was observed 6 weeks after HIM-DFCs (passage #30) were implanted in vivo, thus suggesting that these cells maintain stem cell characteristics. Therefore, subclone HDF1-3 may be related to the differentiation of fibroblasts in the PDL, undifferentiated cells, and osteoblasts and cementoblasts, respectively. Overall, this study is the first to amplify HIM-DFCs and associated subclones with the goal of reconstructing complex or single periodontium. Moreover, our results demonstrate the potential for this treatment approach to address periodontal defects that result from periodontitis, or for the regeneration of teeth.
Introduction
The dental follicle contains embryogenic tissues that surround the tooth germ during tooth genesis and development. DFCs have been characterized to be a heterogeneous cell population, 1 which may contain precursors of the cellular components involved in periodontal development. Correspondingly, DFCs are hypothesized to have the potential to differentiate and form the PDL, cementum, and alveolar inherent bone. In our previous study, periodontal ligament stem cells and DFCs were shown to contribute to the formation of cementum/PDL-like tissues.2–4
Typically, embryogenic dental follicles cannot be harvested during early tooth development due to ethical limitations. However, dental follicles are also present in impacted teeth. The incidence of impacted or embedded third molars is experienced by ∼98% of all individuals, 5 and impacted teeth are often extracted to avoid the incidence of malocclusion deformity,6,7 periodontitis,8–10 and tumor development. 11 Therefore, impacted teeth represent a valuable resource for harvesting DFCs, and a reliable method for harvesting DFCs has been established. 4 In our previous study, DFCs from human impacted molars (HIM-DFCs) were found to maintain their tissue-regenerative abilities, and were able to produce a typical cementum/PDL-like complex. 4 However, this capacity was limited to early passage DFCs, and implied that there are differences that exist between HIM-DFCs and DFCs from tooth germ. As a result, the mechanisms that are involved in the maintenance of stem cell characteristics, heterogeneity, and even embryonic characteristics are of great interest.
In previous studies,1,12 stem cell growth medium (SCGM) was shown to play an important role in maintaining the stem cell characteristics of DFCs in vitro. Therefore, in the present study, HIM-DFCs were harvested, maintained in SCGM, and subclones were established. After 30 passages, the biological characteristics of the original HIM-DFC population, as well as a few isolated subclones, were evaluated. The ability of these cell types to induce EPT regeneration in vivo was also evaluated and compared.
Materials and Methods
Cell culture and phenotype characterization
Cell isolation
Based on panoramic radiographs obtained for each patient, and completion of written informed consent, a single normal impacted third molar was collected from nine individuals ranging in age from 12 to 18 years. Moreover, the experimental protocol for this study was approved by the Committee of Ethics of the Fourth Military Medical University (FMMU), and was conducted in accordance with the Guidelines of FMMU. Human dental follicles were isolated from extracted third impacted molars as previously described. 4 Isolated HIM-DFCs were cultured from these follicles and maintained in SCGM consisting of minimum essential medium (Gibco BRL), 20% heat-inactivated fetal bovine serum (Gibco BRL), 0.292 mg/mL glutamine, 100 U/mL penicillin, 100 μg/mL streptomycin, and 100 μg/mL ascorbic acid (Sigma) in a humidified atmosphere at 37°C and 5% CO2. Medium was changed every 2 days.1,12
Morphological observations of HIM-DFCs
Morphological characteristics of HIM-DFCs were observed using an optical microscope (Olympus Optical), and also by transmission electron microscopy (TEM) (Jeol) as previously described. 12 Alkaline phosphomonoesterase (ALP) activity assays were also performed using an ALP kit according to the manufacturer's protocol (Jiancheng). Briefly, DFCs were fixed with 70% ethanol and incubated with 0.25% naphthol AS-BI phosphate and 0.75% Fast Blue BB dissolved in 0.1 M Tris buffer (pH 9.3). After three washes in phosphate-buffered saline (PBS), cells were monitored and imaged using a phase-contrast inverted microscope (Olympus Optical).
Immunocytochemistry
For immunocytochemistry (ICC) assays that were performed, the following primary antibodies were used as previously described. 12 Primary antibodies included polyclonal rabbit anti-vimentin (1:100), monoclonal mouse anti-cytokeratin (CK14) (1:150), polyclonal rabbit anti-Notch1 (1:100), monoclonal rabbit anti-Stro1 (1:100), polyclonal rabbit anti-ALP (1:80), monoclonal mouse anti-type I collagen (COL-I) (1:100), polyclonal rabbit anti-type III collagen (COL-III) (1:100), and monoclonal mouse anti-osteocalcin (OCN) (1:50). These antibodies were purchased from Abcam Biotechnology.
Flow cytometry
HIM-DFCs (5×105) were incubated with CD105 (1:150; Biolegend), CD90 (1:200; eBioscience), CD29 (1:200; eBioscience), CD44 (1:100; eBioscience), CD14 (1:150; eBioscience), CD45 (1:120; eBioscience), CD34 (1:100; Chemicon), CD146, and Stro-1 (1:100; R&D Systems) according to each manufacturer's protocol and our previous study. 13 FITC-conjugated isotype-matched immunoglobulins were also used to determine nonspecific staining, including goat anti-rat IgG-FITC (Santa Cruz Biotechnology). Cells were analyzed using an FACS Caliber (Becton-Dickinson), and data were analyzed using CellQuest software.
Colony-forming unit assay
HIM-DFCs (5×103) were seeded into 100 mm dish (Costar) and cultured in SCGM. Medium was changed every 2 days, and after 7 days, DFCs were fixed with 4% buffered formaldehyde, stained with 0.1% toluidine blue, and imaged using a phase-contrast inverted microscope. Aggregates with ≥50 cells were scored as a colony. Colony-forming efficiency was calculated according to the number of colonies formed relative to the total number of cells seeded in each culture flask. These experiments were performed in triplicate.
Multipotential differentiation
Adipogenic differentiation
HIM-DFCs (1×105) were seeded into six-well plates. At 80% confluence, HIM-DFCs were cultured in complete Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% newborn bovine serum (NBS; Gibco BRL), 2 μM insulin (Sigma), 0.5 mM isobutyl-methylxanthine (Sigma), and 10 nM dexamethasone (Sigma). 14 The control group was also cultured in DMEM plus 10% NBS, and medium was changed every 2 days. After 15 days, cells were histologically examined, and gene expression profiles were obtained.
For histological examinations, induced HIM-DFCs were fixed in 4% paraformaldehyde for 10 min, and incubated in 0.3% Oil Red O (Sigma) for 15 min. After three washes in PBS, cells were monitored and imaged using a phase-contrast inverted microscope.
For gene profiling, RNA of HIM-DFCs was extracted using Trizol Reagent (Invitrogen), according to the manufacturer's protocol. cDNA synthesis and polymerase chain reactions (PCRs) were performed as previously described. 15 Primer sequences used to detect peroxisome proliferator-activated receptor gamma (PPARγ) and CCAAT/enhancer binding protein α (C/EBPα) are listed in Table 1. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as an internal control. The PCR program used included the following cycles: 94°C for 5 min, 35 cycles of 94°C for 45 s, 57°C for 45 s, and 72°C for 1 min, followed by 72°C for 1 min. These assays were performed in triplicate, and the PCR products obtained were confirmed by sequencing (Sangon Biotechnology Co.).
Osteogenic differentiation
HIM-DFCs (1×105) were seeded into six-well plates. At 80% confluence, HIM-DFCs were cultured in osteogenic inducing medium containing 10% NBS, 5 mM L-glycerophosphate (Sigma), 100 nM dexamethasone (Sigma), and 50 μg/mL ascorbic acid for 15 days. The control group was cultured in DMEM plus 10% NBS. Medium was changed every 2 days. After 15 days, cells were histologically examined, and the expression of certain genes was profiled.
For histological examinations, induced HIM-DFCs were fixed in 4% paraformaldehyde for 10 min, then incubated in 0.1% alizarin red solution (Sigma) in Tris-HCl (pH 8.3) at 37°C for 30 min. After two washes with PBS, cells were imaged using an inverted microscope.
Gene expression of Runx2 and OCN was performed according to Adipogenic differentiation section, and primer sequences used are listed in Table 1.
Selecting HIM-DFCs subpopulation clones
To grow subclones of the HIM-DFCs isolated, limited dilution and amplification was performed. Briefly, isolated HIM-DFCs in a logarithmic growth phase were resuspended in SCGM to a density of 10 cells/mL. In a 96-well plate, 100 μL aliquots were plated, and after 24 h, wells containing only a single cell were considered as valid groups. Medium was subsequently changed every 2 days, and at ∼70% confluence, amplified cells were successively passaged into larger plates (Costar), up to a T-75 cell culture flask. DFx clones were passaged until proliferation was no longer observed.
Biological characteristics of HIM-DFCs subpopulation clones
To characterize the biological characteristics of HIM-DFCs clones, those that completed 30 passages were assayed for cell proliferation, ALP activity, and expression of genes relative to mineralization.
Cell proliferation
HIM-DFCs clones (1×104 cells/mL) were seeded onto 0.8×0.8 cm coverslips in 24-well plates. After 2 days, cells were stained for Ki67. Briefly, cells were incubated with a monoclonal human anti-Ki67 antibody (1:200) (Millipore), and then a rhodamine-conjugated secondary antibody (1:200; Chemicon AP182F). Cells were also counterstained with Hoechst. All samples were examined under a fluorescence microscope (Olympus Optical), and the number of cells positive for Ki67 per visual field (e.g., at 10× and 20×) were counted and recorded. These experiments were performed in triplicate.
ALP activity
ALP activity was assayed as described in the Morphological observations of HIM-DFCs section.
Gene detection
Total RNA was extracted from each HIM-DFC clone using TRIzol reagents (Invitrogen Life Technologies). First-strand cDNA synthesis was performed using the PrimeScript® RT reagent kit (Takara). For Real-Time PCR, the Quantitect Sybr Green Kit (Toyobo) and an ABI Prism 7700 Sequence Detection System (Applied Biosystems) were used. Primer sequences used to detect COL-I, COL-III, BSP, and Runx2 are listed in Table 2. PCR assays included the following cycles: 95°C for 10 min, then 45 cycles of 95°C for 15 s followed by 60°C for 1 min. These assays were performed in triplicate, and gene expression levels were normalized to GAPDH.
Characterization of epitope profiles
The methods used were described in the Flow cytometry section: Passage #30 HIM-DFCs and derived clones were analyzed.
Generation of a cementum-PDL complex in vivo
To evaluate HIM-DFC differentiation in vivo, HIM-DFCs and HIM-DFC clones were combined with ceramic bovine bone (CBB; Research and Development Center for Tissue Engineering, FMMU) and implanted into the dorsum of immunodeficient mice. Briefly, for a single transplantation, 3×106 cells were mixed with 40 mg CBB and centrifuged at 800 g for 5 min. For each sample, the supernatant was aspirated so that the remaining precipitate could be resuspended in 50 μL of medium. These aliquots were then transferred to culture dishes and incubated at 37°C, and 5% CO2 for 24 h before being implanted into the dorsum of 6-week-old severe combined immunodeficiency mice (Laboratory Animal Research Centre of FMMU). Six mice were used in each group. Our animal use protocols were reviewed and approved by the Animal Care Committee of FMMU. Six weeks later, implants were resected under deep anesthesia. Implants were subsequently fixed with 4% paraformaldehyde overnight at 4°C, demineralized with 10% EDTA (pH 8.0), and then embedded in paraffin. Paraffin sections were stained with hematoxylin and eosin and Mason stain.
For immunohistochemical staining, sections were incubated with primary antibodies of monoclonal mouse anti-COL-I (1:100), polyclonal rabbit anti-COL-III (1:100), monoclonal mouse anti-OCN (1:50), polyclonal rabbit anti-bone sialoprotein (BSP) (1:80), Polyclonal rabbit anti-Periostin (1:500), and monoclonal mouse anti-cementum attachment protein (1:200). These antibodies were purchased from Abcam Biotechnology except that CAP were obtained from Santa Cruz Biotechnology. Biotinylated secondary antibodies (1:1000) were purchased from Dako. All the samples were examined under a compound microscope (Olympus Optical). The experiment was repeated at least thrice.
Statistical analysis
All data are expressed as the mean±SD. Statistical significance was analyzed using SPSS 11.5 software (SPSS), and analysis of variance was according to the Bonferroni method. A value of p<0.05 was considered statistically significant.
Results
Characterization of isolated HIM-DFCs
Human dental follicles were obtained from extracted third impacted molars, and isolated cells were cultured in SCGM. Expression of vimentin was detected (Fig. 1D), whereas CK14 remained undetectable (Fig. 1E). These results indicated that mesenchymal-type cells had been isolated, and these cells were not contaminated with epithelial cells. TEM examination also confirmed that homogeneous electron-dense granules present in the cultured HIM-DFCs represented characteristic markers of DFCs (Fig. 1F). In combination, these results confirmed that the dental follicles isolated were DFCs.
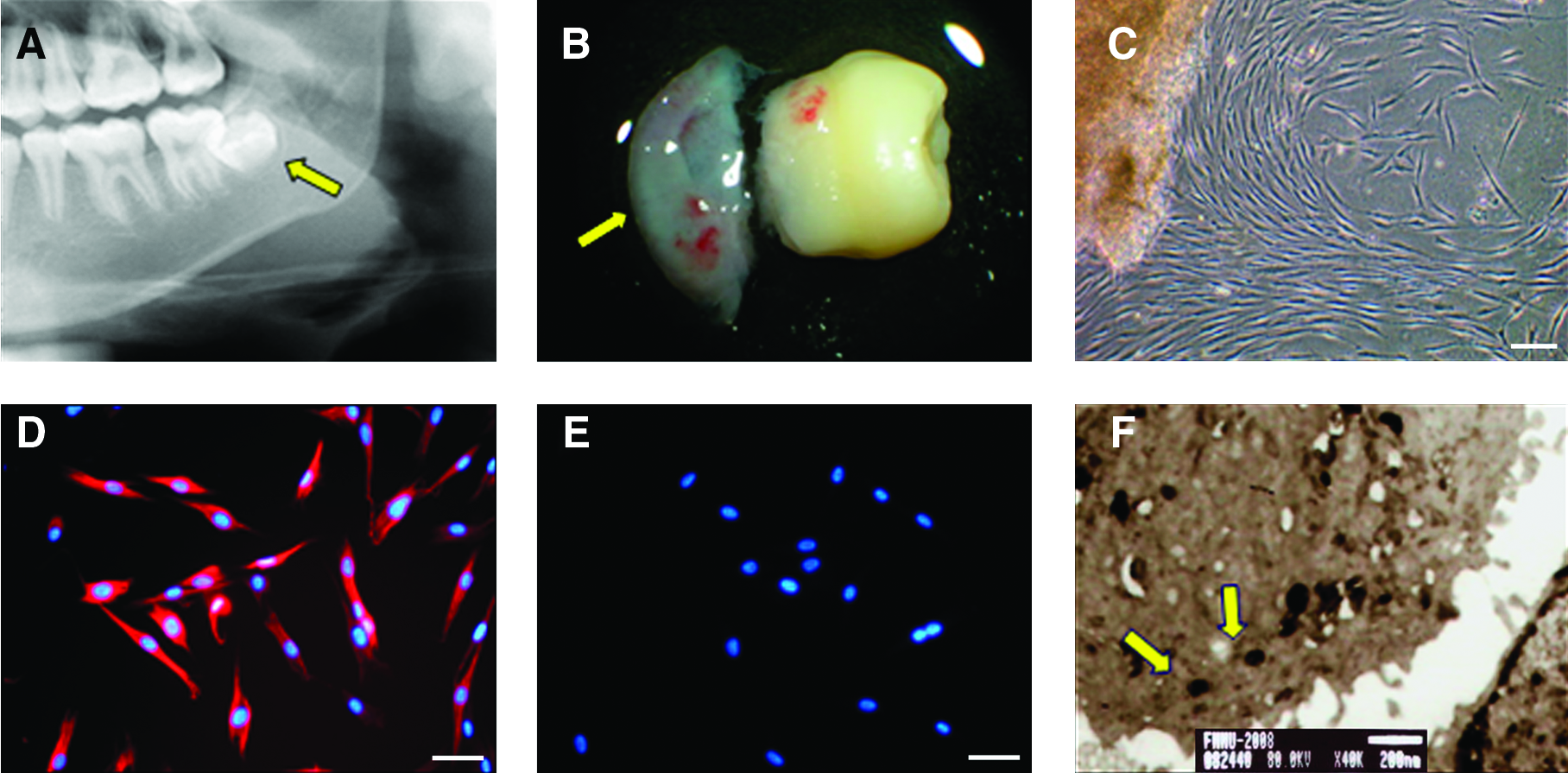
Culture and identification of HIM-DFCs.
Detection of HIM-DFC heterogeneity
Using a light microscope, both long and short spindles, as well as polygons, were observed (a in Fig. 2A). In ALP staining assays, HIM-DFCs appeared to exhibit colony-like growth, and a subset of colonies were positive for ALP. Depending on the extent of ALP staining observed, different grades of HIM-DFC colonies could be observed (b, c in Fig. 2A). In addition, organelles at different developmental stages were also detected in an analysis of HIM-DFCs ultrastructure (d, e in Fig. 2A).
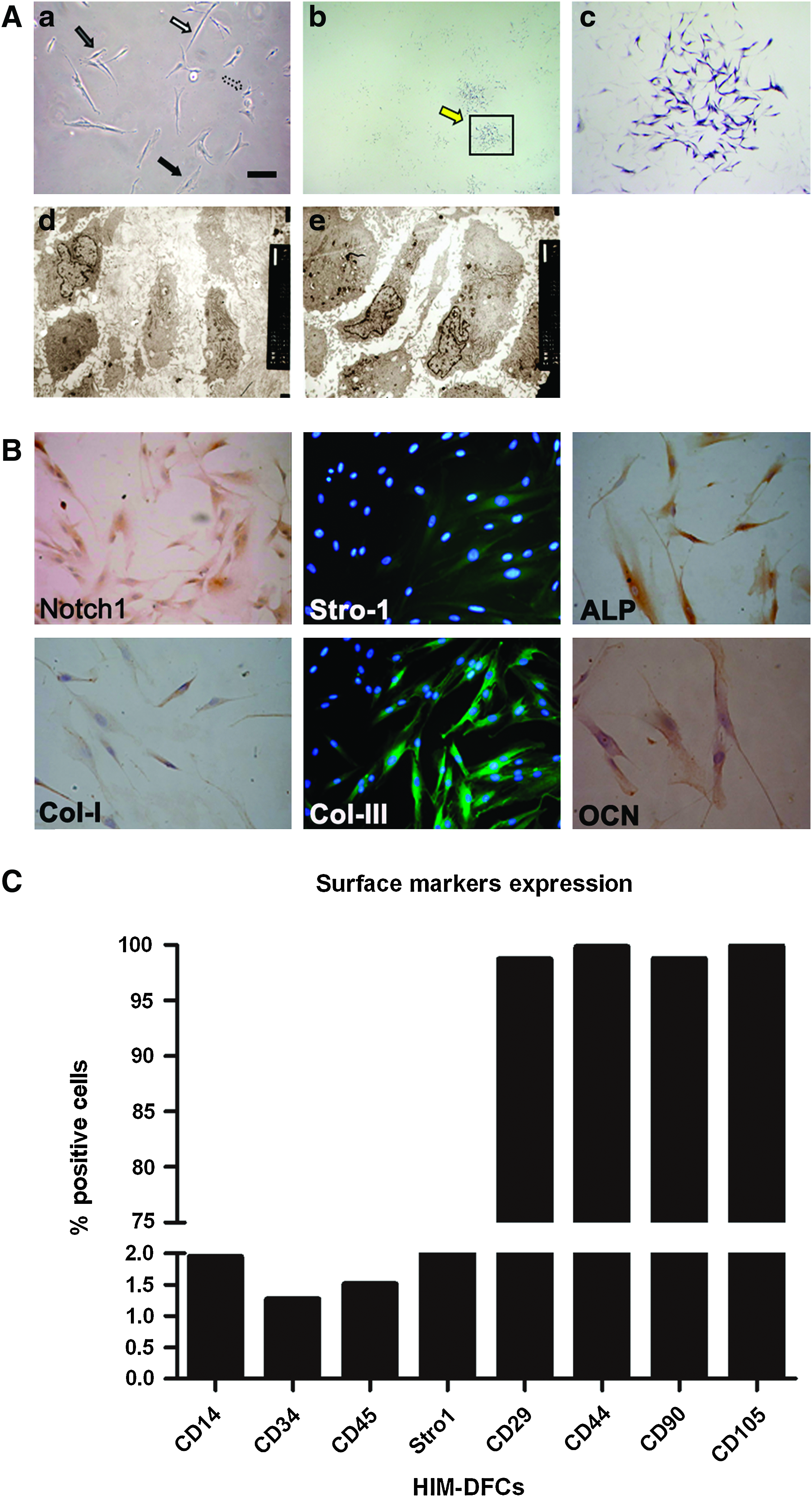
Detection of heterogeneity.
ICC analysis further demonstrated that HIM-DFCs were positive for expression of Notch-1, stro-1, ALP, COL-I, COL-III, and OCN (Fig. 2B), which are regarded as markers of heterogeneity. In addition, flow cytometry (FCM) assays detected the expression of CD105, CD90, stro-1, CD29, and CD44 by HIM-DFCs (Fig. 2C and Supplementary Fig. S1; Supplementary Data are available online at www.liebertonline.com/tea). However, expression of CD14, CD34, and CD45 was not detected. In combination, these results demonstrate that a heterogeneous population of cells was isolated from impacted molars.
When colony-forming assays were performed, colonies were clearly observed after HIM-DFCs were incubated with SCGM for 7 days (Fig. 3A–D). Further, when HIM-DFCs were cultured in osteogenesis medium, or adipogenesis-inducing medium, for 15 days, mineralized nodules (Fig. 3F) and lipid droplets (Fig. 3H) were detected with alizarin red staining and oil red staining, respectively.
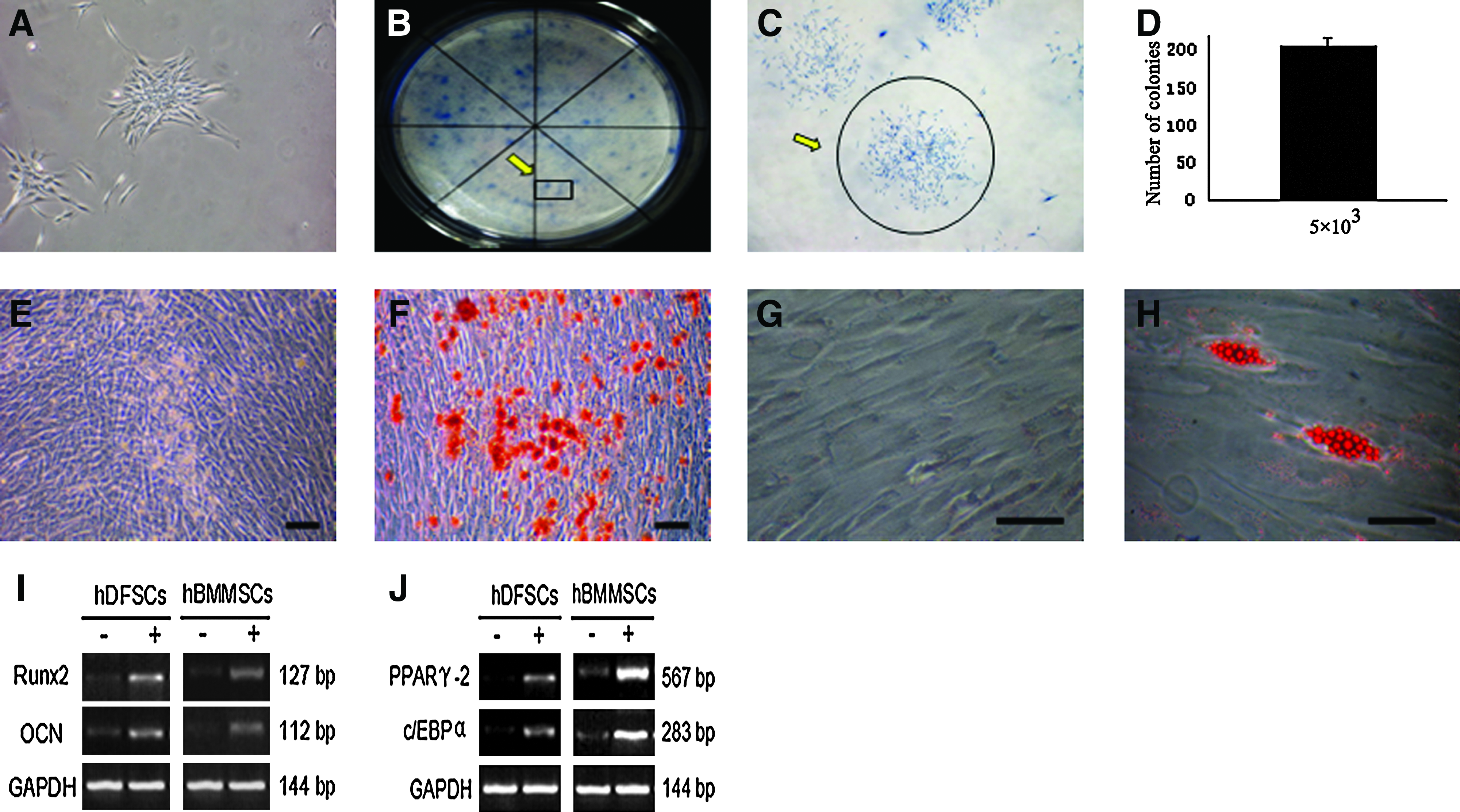
Characteristics of the stem cells present in the HIM-DFCs analyzed.
Reverse transcription-polymerase chain reaction (RT-PCR) analyses also demonstrated that when HIM-DFCs were grown in osteogenic or adipogenic media, mRNA levels of the osteogenic markers, Runx2 and OCN (Fig. 3I), or the adipogenic markers, PPARγ2 and c/EBPα (Fig. 3J), increased, respectively.
Selection and characterization of HIM-DFCs subclones
A total of 48 clones were obtained from limited dilution experiments performed using HIM-DFCs. Based on the cell morphologies associated with each of these clones, and ALP staining assays that were performed, the 48 clones were classified as HDF1, 2, or 3. These designations represented the three ALP staining patterns that were observed: long, spindle-like fibroblasts (HDF1), polygonal-like undifferentiated cells (HDF2), or short, spindle-like osteoblasts (Fig. 4A, B). HDF3 clones were also associated with the strongest ALP staining, whereas HDF1 clones exhibited the weakest ALP staining (Fig. 5A, B). In contrast, HDF2 clones had the highest percentage of Ki67-stained cells, whereas HDF1 and HDF3 clones had similar, lower levels of Ki67 staining (Fig. 4C, D).
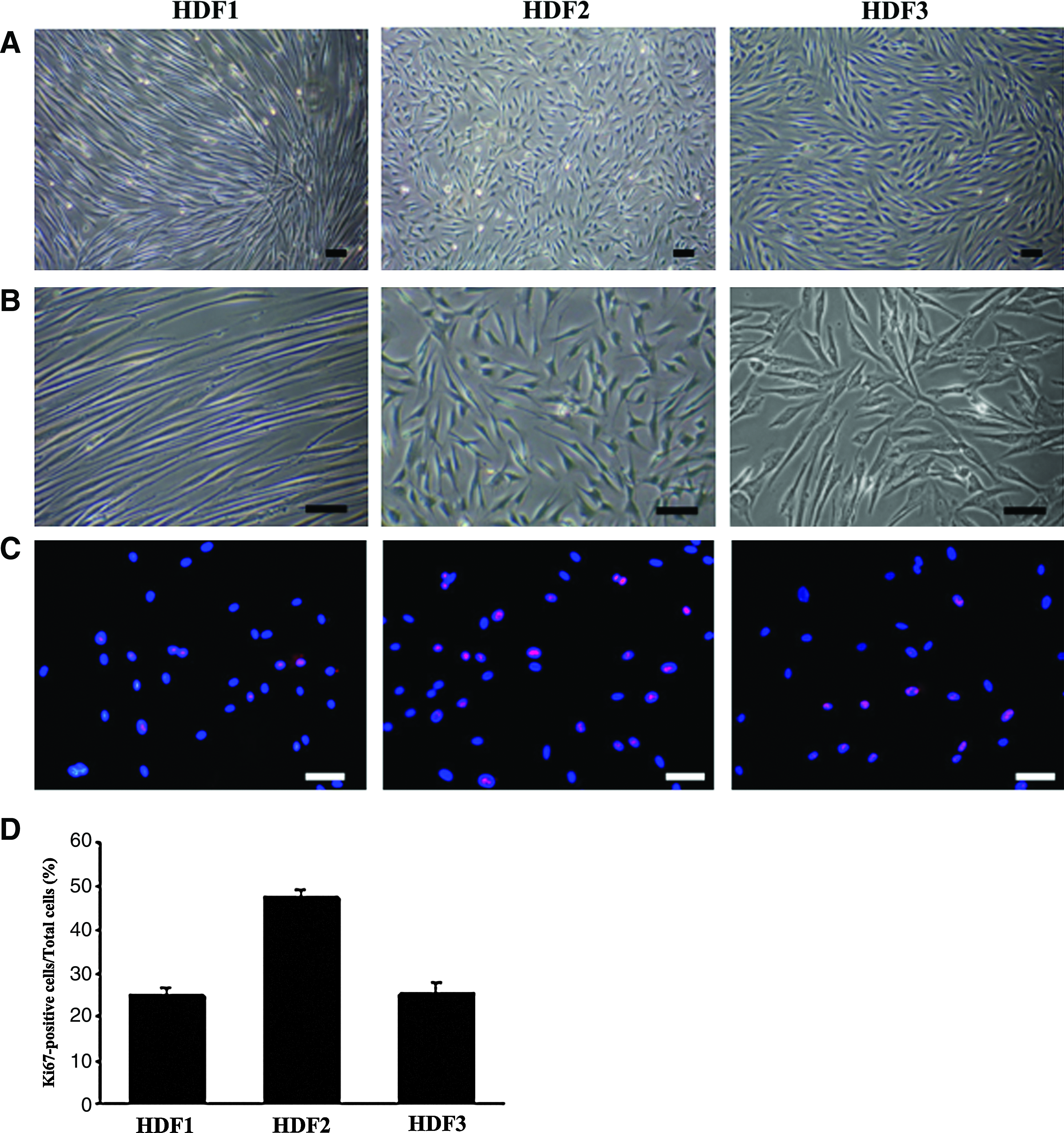
Characterization of three HIM-DFC subclones that are representative of HDF1, HDF2, and HDF3. The morphologies
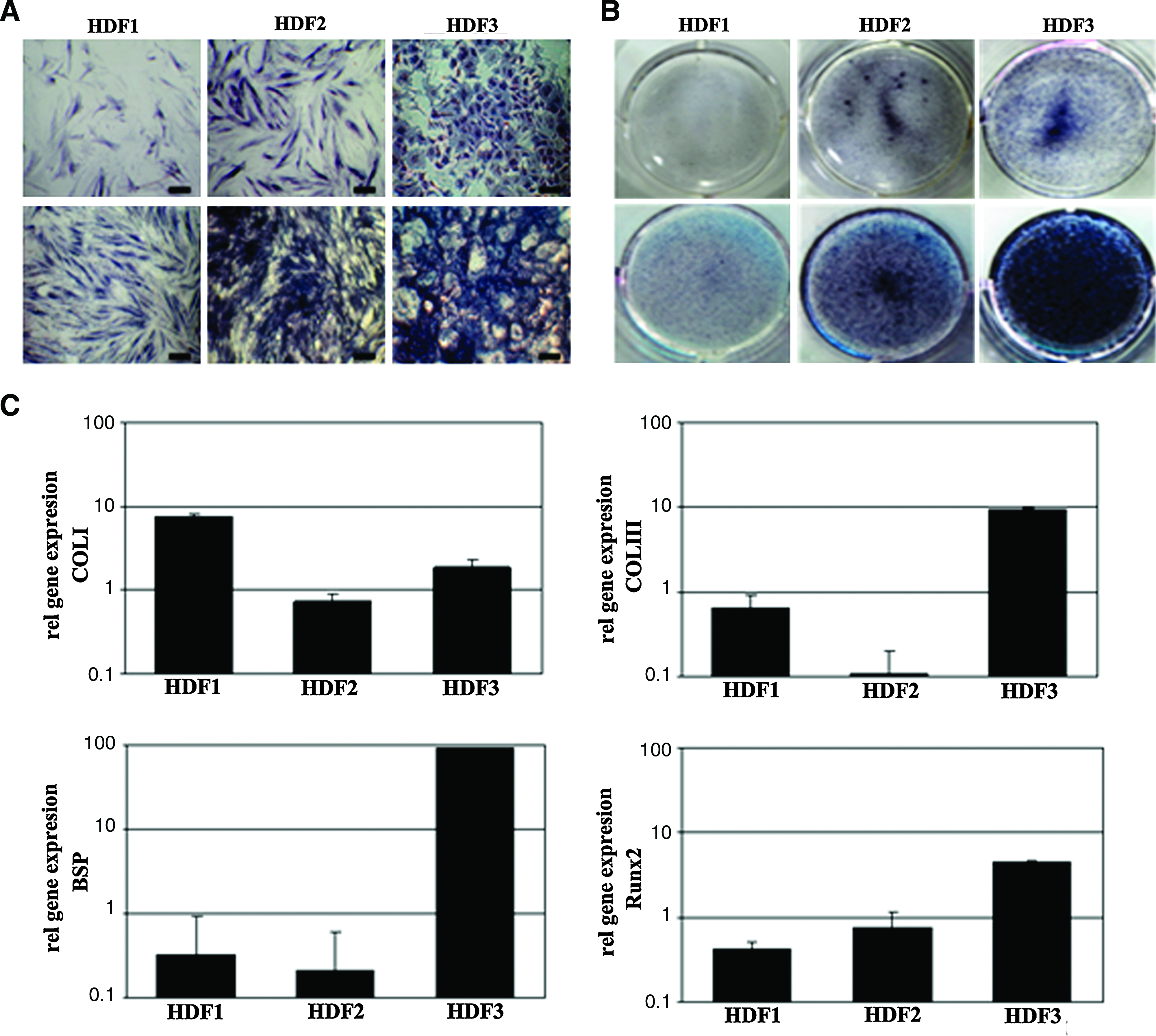
Mineralization associated with each of the three subclone types analyzed.
In an analysis of gene expression for the three subclone types, real time-PCR analysis detected high levels of COL-I, COL-III, BSP, and Runx2 mRNA in HDF3 clones. In contrast, HDF1 clones were associated with high levels of COL-I and COL-III mRNA, and lower levels of BSP and Runx2 mRNA. For HDF2 subclones, low levels of COL-I, COL-III, and BSP mRNA were detected, although high levels of Runx2 mRNA were also detected (Fig. 5C).
In FCM analyses, HIM-DFCs and one of each HDF1, 2, and 3 subclone was found to highly express CD105, CD90, CD29, CD44, and the mesenchymal stem cell markers, CD146, and Stro1. However, levels of CD34, CD14, and CD45 were undetectable in these cell types, except for the HDF2 clone, which expressed low levels of CD45, a marker associated with stem cells. The HDF2 clone also expressed higher levels of Stro-1 compared with the other two clones, whereas levels of CD146 and Stro-1 were the lowest for the HIM-DFCs. In all three subclone types, similar levels of CD146 were detected (Fig. 6 and Supplementary Fig. S2).
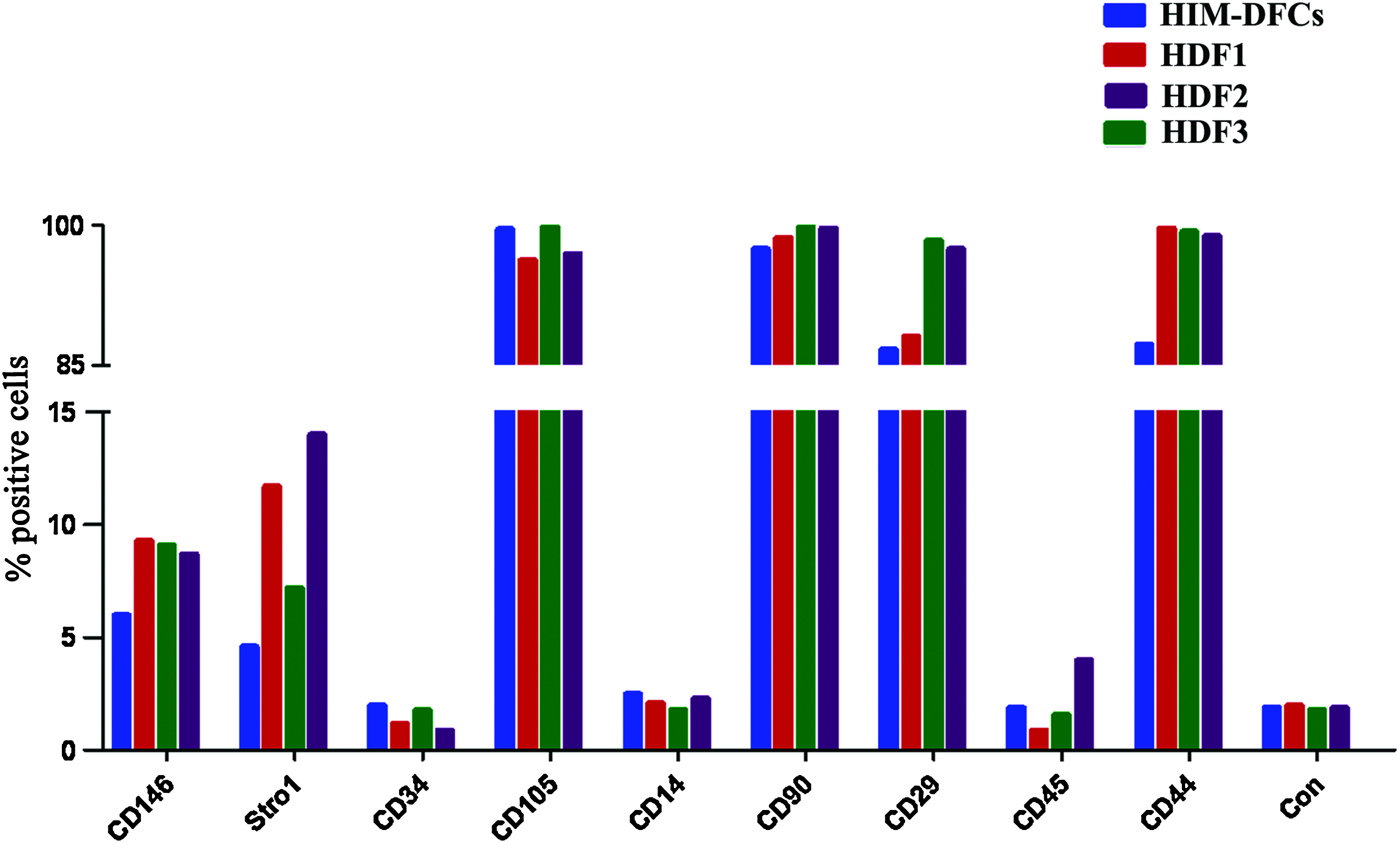
Surface marker expression by HIM-DFCs versus HDF1-3 based on FCM data. Detection of CD146, stro1, CD34, CD105, CD14, CD90, CD29, CD45, and CD44 are compared with a negative control (Con) for HIM-DFCs, a HDF1, a HDF3, and a HDF2. Color images available online at www.liebertonline.com/tea
In vivo generation of a cementum-PDL complex
The differentiation potential of HIM-DFCs, as well as a subclone representative of each HDF1, 2, and 3 category, were evaluated when these cell types were combined with CBB and implanted into the subcutaneous pockets of immunodeficient mice. After 6 weeks, these implants were resected, and histological examinations were performed. Based on these analyses, the HDF1 clone was found to be associated with the formation of rich fiber tissues positive for the markers associated with collagen COL-I and COL-III (Fig. 7A–D and Supplementary Fig. S3), whereas the HDF2 clone was associated with a few fiber tissues and a mineralized matrix positive for COL-III and OCN associated with osteogenesis (Fig. 7E–H and Supplementary Fig. S3). For the HDF3 clone, a rich, mineralized matrix was detected (Fig. 7I–L), and these matrixes were positive for the osteogenesis markers OCN and BSP (Supplementary Fig. S3). In contrast, the population of heterogenous HIM-DFCs that were implanted was associated with the formation of a cementum-PDL-like complex positive for the PDL markers periostin, COL-III, and cementum marker CAP (Fig. 7M–P and Supplementary Fig. S3).

Various differentiation states detected for HIM-DFCs in vivo. Each image is representative of resected tissues that were paraffin embedded, sectioned, and then stained with hematoxylin and eosin
Discussion
Dental follicle tissues play a key role in both tooth eruption and the formation of PDTs. In the latter case, PDTs represent heterogeneous tissues that are composed of a PDL, AB, and cementum that cover the dentin. Correspondingly, DFCs are the precursor cells of PDL cells, cementoblasts, and osteoblasts, which lead to the formation of PDLs, cementum, and ABs, respectively. Therefore, heterogeneous DFCs have the potential to be seeding cells for the regeneration of PDTs.
In the present study, expression of Notch1, Stro-1, ALP, COL-I, COL-III, and OCN was detected in HIM-DFCs. Earlier, both Notch1 signaling and Stro-1 have been shown to be necessary for the maintenance of a reservoir of undifferentiated cells.16–18 Thus, it is significant that HIM-DFCs were also found to express these markers. ALP, COL-I, and OCN are also markers of osteogenic or cementoblastic differentiation, thus suggesting that HIM-DFCs have the potential to differentiate into osteoblasts and cementoblasts.19–21 Moreover, COL-I and COL-III represent functional markers of PDL fibroblasts. 22 Therefore, these results suggest that HIM-DFCs contain heterogeneous cell types that include PDL fibroblasts, osteoblasts, or cementoblasts, and undifferentiated cells. 23 Further, based on the different degrees of ALP staining observed for the HIM-DFC colonies analyzed, cells in various differentiation states were detected. 24 The ultrastructural characteristics of organelles associated with the HIM-DFC populations analyzed were also consistent with these observations. In combination, these heterogeneous biological characteristics support the use of HIM-DFCs as seeding cells for the generation of engineered PDTs that consist of cementum, PDL, and AB.
The results of the present study also demonstrate that subcloning of isolated HIM-DFCs could be used to obtain follicle cells in different states of differentiation. 23 For example, in this study, subclones were categorized as HDF1, 2, or 3 depending on the cell morphologies observed and the degree of ALP staining associated with each subclone type. In addition, high levels of COL-I, COL-III, BSP, and Runx2 mRNA were detected in type III clones, and a combination of high levels of COL-I and COL-III mRNA, and lower levels of BSP and Runx2 mRNA, were detected in HDF1 clones. These results suggest that HDF3 clones are related to the differentiation of osteoblasts and cementoblasts in the periodontium,25–28 whereas HDF1 clones may be related to the differentiation of fibroblasts in the periodontium.29,30 Classic developmental theory holds that dental follicle tissues can be divided into three layers, with loose connective tissues present between each layer. HDF1 subclones were found to exhibit fibroblast-like characteristics and weak osteogenic differentiation, thus implying that HDF1 subclones may be the precursor cells of fibroblasts present in PDLs. 23 Outer and inner layers of dental follicles will also develop into AB tissues and cementum, respectively. Correspondingly, HDF3 subclones were associated with strong osteogenic characteristics, which indicate that these cells may be precursor cells of AB cells and cementoblasts. 23 In contrast, HDF2 subclones exhibited characteristics of an undifferentiated cell type, 23 and this would be consistent with the need for a stem cell population to be present in PDTs. 17 Further, in this study, high levels of mesenchymal cell markers were found to be expressed by the three passage #30 subclone types analyzed, as well as the heterogeneous population of HIM-DFCs. However, these four cell types were also found to have high levels of the mesenchymal stem cell markers, CD146 and Stro1, which revealed that they still maintained strong stem cell characteristics.31,32
Although previous studies have reported the biological characteristics of DFCs in vitro,33–37 inconsistent results have been reported their differentiation properties in vivo. For example, DFCs have been shown to contribute to the formation of heterotopic fiber bundles, and not osteoid tissues.18,38,39 However, other studies, including our previous work, have suggested that DFCs contribute to the formation of cementoid-like tissues or a mineralized matrix after heterotopic transplantation.2,40–44 In combination with the analysis of biological characteristics of HIM-DFCs in vitro performed in the present study, we hypothesize that these inconsistencies may be due to differences in the heterogeneity of DFCs, as well as the microenvironment in which they are maintained. In other words, osteoid tissues would form if HDF3 subclones were a predominant cell population in the heterogeneous DFC population transplanted, and fiber bundles would form if HDF1 subclones were predominant.
To further evaluate the differentiation properties of HIM-DFCs and associated subclones in vivo, heterogeneous HIM-DFCs, as well as HDF1, 2, and 3 HIM-DFC subclones, were combined with CBB and implanted into subcutaneous pockets of athymic nude mice. After 6 weeks, the heterogeneous population of HIM-DFCs was associated with the generation of a cementum-PDL-like complex. In contrast, HDF1, 2, and 3 subclones were associated with the formation of single tissues. Based on these results, it is apparent that a heterogeneous HIM-DFC population is needed to regenerate PDTs, rather than a single cell type. The balance between the different subcolony subtypes of heterogeneous HIM-DFCs may also determine the formation of different target tissues in combination with the influences of the microenvironment. However, the ability to isolate different cell types from the heterogeneous population of HIM-DFC cells obtained from extracted molars suggests that specific cell types could be obtained for regeneration of a specific tissue when needed.
In this study, heterogeneous HIM-DFCs were observed to generate PDT complexes in combination with CBB, whereas single subclone types contributed to the generation of single PDTs, such as a PDL, AB, or cementum. However, for various PDT defects, or tooth loss, that are caused by different periodontal diseases, it is important to determine how the interactions between different subclone cell types of HIM-DFCs determine the type of tissue that can be regenerated. Further, it remains unclear how the microenvironment of HIM-DFCs contribute to the balance between the cell types present that result in tissue generation. Meanwhile, whether chromosome of seeding cells are stable and normal is also key for tissue engineering periodontal tissues. 45 However, it is still unclear whether there are any abnormalities in karyotype of DFCs used as seeding cells in this study. Therefore, additional studies are needed to address these important considerations.
Conclusion
In summary, the present study investigated the heterogeneity and characteristics of HIM-DFCs and associated subclones, both in vitro and in vivo. Based on these experiments, HIM-DFCs appear to be a valuable resource for the generation of PDTs. Additional studies will need to be performed to confirm the results of the present study, particularly the cytological basis for using the 30th passage of heterogeneous HIM-DFCs and associated subclones. However, the potential for HIM-DFCs to generate tissues to treat different disease with PDTs looks promising.
Footnotes
Acknowledgments
This study was supported by grants from the Nature Science Foundation of China (30725042, 81020108019, and 31030033) and the National Basic Research Program (973 Program) (2010CB944800).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.