
Abstract
Enamel matrix proteins (EMP) have been shown to promote regeneration of periodontal ligament and root cementum, and sometimes to enhance the differentiation of bone-forming cells in vitro and new bone growth in vivo. However, the inconsistent and unpredictable effects of EMP that have been reported for bone regeneration may be due to the highly variable composition of this heterogeneous material, which is comprised mainly of amelogenin and amelogenin-derived peptides. The present study has therefore examined the effects of naturally occurring low-molecular-weight (LMW) and high-molecular-weight (HMW) fractions of Emdogain® (EMD; Institut Straumann, Basel, Switzerland), a commercially available form of EMP, on osteogenic differentiation of bone precursor cells in vitro. In addition, the effects of chemically synthesized specific components of LMW and HMW—namely, the tyrosine-rich amelogenin peptide (TRAP), a specific amelogenin isoform derived by proteolytic clipping, and a leucine-rich amelogenin peptide (LRAP), an isoform derived by alternative splicing—on bone-forming cell activity were also investigated. Our findings demonstrate that while TRAP suppressed the formation of bone-like mineralized nodules, LRAP upregulated osteogenic differentiation. Furthermore, synthetically produced TRAP and its unique C-terminal 12 amino acid sequence (TCT) also suppressed bone-forming cells, whereas LRAP and its unique C-terminal 23 amino acid sequence (LCT) markedly enhanced terminal differentiation of bone-forming cells. These findings suggest that the differential effects of amelogenin-derived peptide sequences present in EMP could be of potential clinical value, with the novel bioactive TCT peptide as a useful tool for limiting pathological bone cell growth and the unique LCT sequence having therapeutic benefits in the treatment of periodontal and orthopedic diseases.
Introduction
Recent analysis of Emdogain® (EMD; Institut Straumann, Basel, Switzerland), a commercially prepared lyophilized form of EMP, has demonstrated the presence of low-molecular-weight (LMW) and high-molecular-weight (HMW) fractions. 16 Both EMD and the two fractions LMW and HMW are naturally occurring, and all of them have been prepared from crude EMP extracts by a precisely controlled patented process that involves heat treatment of EMP. The LMW fraction comprises mainly an amelogenin-derived <6-kDa TRAP and the HMW fraction contains >6-kDa peptides, including the amelogenin peptide LRAP, sheathlin, and full-length amelogenin. 1 While sheathlin has been shown to have a role in cementum development and repair, 17 the effects of these amelogenin peptide-containing subfractions of EMD (LMW and HMW) and chemically synthesized amelogenin peptides (TRAP and LRAP) on osteogenesis have not been investigated. The present study has therefore examined the response of bone-forming cells to both naturally occurring and chemically synthesized EMD components under conditions that facilitate terminal osteogenic differentiation in vitro.
Materials and Methods
Isolation of primary human PDL and alveolar bone cells
Human PDL was obtained from periodontally healthy adult patients undergoing routine extractions at the Eastman Dental Hospital, as previously described. 18 The participants signed informed consent, in accordance with the protocol approved by the Joint Research and Ethics Committee of the Eastman Dental Institute and Hospital. Briefly, tissue was separated from the surface of the middle portion of the root and digested with 3 mg/mL of collagenase type I and 4 mg/mL of dispase (both from Sigma, Gillingham, United Kingdom) for 1 h at 37°C. Single-cell suspensions were obtained by passing the cells through a 70-μm sieve (Falcon, Becton-Dickinson, Cowley, United Kingdom) and cultured in α-modified Eagle's medium (α-MEM) (Gibco Life Technologies Ltd., Paisley, United Kingdom), containing 10% fetal bovine serum (FBS) (PAA Laboratories, Yeovil, United Kingdom) supplemented with 200 U/mL penicillin, 200 μg/mL streptomycin, and 2 mM L-glutamine (all from Gibco Life Technologies Ltd.) at 37°C in a humidified atmosphere of 5% CO2 in air. Three separate PDL cell populations from three different healthy donors (all male, aged between 18 and 25) were used between passages 3 and 6.
Primary human alveolar bone (AB) cells were grown from cortico-lamellar bone of the maxilla of patients undergoing routine extractions at the Eastman Dental Hospital, as previously described. 19 Briefly, the bone chips were cut into ∼1–2 mm3 pieces, placed in 48-well plates, and incubated at 37°C in a humidified atmosphere of 5% CO2 in air for 15 min to enable them to adhere to the culture plates. Low-glucose–Dulbecco's modified Eagle's medium (low-glucose-DMEM; Gibco Life Technologies Ltd.) containing 10% fetal calf serum (FCS; PAA Laboratories) supplemented with 200 U/mL penicillin, 200 μg/mL streptomycin, and 2 mM L-glutamine (all from Gibco Life Technologies Ltd.) was then added, and the explants were cultured until the outgrowth of the adherent cells reached confluence. Cells were used between passages 2 and 5 in the experiments.
In addition to the PDL and AB cells, BMSC (Lonza, Slough, United Kingdom) and a recently isolated clonogenic cell population from adult human PDL that was shown to exhibit characteristic features of a bone progenitor/stem cell (clone 7) 20 were also used in the present study to examine the effects of EMD components on a range of bone-forming cells. BMSC and clone 7 were cultured in the same way as the PDL and AB cells, described above, and used between passages 3 and 6.
Treatment of cells with EMD and the EMD fractions
EMD, an EMD-derived fraction <6 kDa (LMW proteins; fraction P5, the last ∼10% of the total protein separated by size-exclusion chromatography 16 ) and an EMD-derived fraction >6 kDa (HMW proteins; a combination of fractions P1–P4, the first ∼30% of total protein separated by size-exclusion chromatography 16 ) were all provided by Institut Straumann. They were diluted in 0.1% acetic acid and added directly to the cells when they reached ∼90% confluence. EMD and the EMD LMW and HMW fractions were used at a final concentration of 100 μg/mL throughout the experiments, unless otherwise noted. These concentrations were determined from their dose effects on terminal PDL cell osteogenesis in vitro, as described below.
Effects of the EMD, LMW, and HMW on terminal differentiation of bone-forming cells
PDL cells were seeded into 24-well plates at a density of 2.5×104 cells/well and cultured in nonselective growth medium (GM) containing low-glucose-DMEM supplemented with 10% FCS, 200 U/mL penicillin, and 200 μg/mL streptomycin for 3 days. Osteogenic medium (OM) was then added, consisting of GM supplemented with 0.1 mM L-ascorbic acid 2-phosphate, 10 mM β-glycerophosphate, and 10 nM dexamethasone (all from Sigma). EMD, LMW, and HMW with concentrations of 1, 3, 10, 30, and 100 μg/mL were added to the cultures, and the medium was changed every 3 days.
Similarly, AB, BMSC, and clone 7 were also seeded into 24-well plates at a density of 2.5×104 cells/well and cultured in GM for 3 days until 90% confluent; the medium was then replaced with OM containing EMD, LMW, and HMW (all 100 μg/mL) and was changed every 3 days. Control cultures were incubated with OM alone.
The formation of mineralized nodules, a measure of terminal osteogenic differentiation, was determined by alizarin red S staining of calcium-containing deposits, as follows. After 3 weeks of culture in OM in the presence of EMD, LMW, and HMW, the cells were fixed with 10% formaldehyde for 15 min and washed with distilled water. The samples were then incubated with 2% alizarin red S (pH 4.2) (Sigma) for 15 min at room temperature, washed with ice-cold methanol, air-dried, and photographed. The level of alizarin red staining was quantified by absorbance at 562 nm (A562), after eluting the stain for 2 h with 10% cetylpyridinium chloride in 10 nM sodium phosphate buffer, pH 7.0.
Quantitative-polymerase chain reaction analysis of osteogenesis-associated genes
PDL cells were seeded into 24-well plates at a density of 2.5×104 cells/well and cultured in GM for 3 days. GM was then replaced with OM and the cells cultured in the presence of EMD, LMW, and HMW and the medium changed every 3 days. On days 4, 7, 10, and 14, total RNA was extracted and quantitative-polymerase chain reaction (Q-PCR) carried out to measure the early marker gene runt-related transcription factor-2 (Runx2) (day 4), a key transcription factor involved in the in vitro intramembraneous ossification/AB development of PDL cells, the intermediate osteogenic marker gene OP (day 7) and the late OC (day 14) and BSP (day 14) genes, as described below. (The Sox-9 gene is considered to be involved primarily in cartilage template development and endochondral ossification/long bone formation 21 and was therefore not measured here.) The different time points selected for the above transiently expressed osteogenic markers and terminal osteogenic differentiation (noted above) were based on previously reported studies22–24 and on our own kinetic experiments (not shown here) in which PDL cells were cultured in OM in the absence and presence of EMD, LMW, and HMW for 1, 4, 7, 10, 14, and 21 days.
Total RNA was isolated from PDL cells using the RNeasy Mini Kit (Qiagen, Crawley, United Kingdom) according to the manufacturer's instructions. For reverse transcription, first-strand cDNA was synthesized using 1 μg of total RNA, as previously described, 20 with primers obtained from Applied Biosystems (Foster City, CA). Q-PCR analysis was carried out using the ABI Prism® 7300 sequence detector (Applied Biosystems), the TaqMan® Gene Expression Assay consisting of the unlabelled specific PCR primers, and TaqMan MGB probes with FAM™ dye labeling in a 96-well-plate format. Thermal cycler conditions were used as recommended by the manufacturer, and the data were collected and analyzed by the SDS 1.2 software (Applied Biosystems). All PCRs were performed in triplicate and each of the gene cycle threshold (ct) value normalized to the glyceraldehyde 3-phosphate dehydrogenase (GAPDH) ct values detected simultaneously on the same plate.
ALP activity assay
PDL cells were cultured in OM in the absence and presence of EMD, LMW, and HMW, with the medium changed every 3 days. Since ALP is involved in a late phase of mineralization, 19 its activity in PDL cells was measured on day 14, as described previously. 19 Briefly, triplicate wells were washed with phosphate-buffered saline (PBS) and the cells incubated with 200 μL of 5 mM p-nitrophenyl phosphate in 50 mM glycine, 1 mM MgCl2, and 150 mM 2-amino-2-methyl-1-propanol buffer, pH 10.5, at 37°C for 1 h. A 50 μL aliquot of 3 M NaOH was added to stop the enzymatic reaction and the absorbance measured at 405 nm (A405).
Effects of the BMP-antagonist noggin and the TGFβ-antagonist UB-505124 on EMD-, LMW-, and HMW-mediated PDL osteogenesis
To determine whether EMD, LMW, and HMW act via a BMP- and/or TGFβ-dependent pathway of osteogenic differentiation, the effect of noggin, a BMP pathway inhibitor,25,26 and UB-505124, a TGFβ pathway inhibitor,26,27 was measured using PDL cells, as described below. The cells were plated and cultured in OM for 2 weeks in the presence of EMD, LMW, and HMW. Replicate test cultures also contained 100 ng/mL of noggin (Peprotech, London, United Kingdom) or 1 μM UB505124. After 2 weeks, the cultures were stained with alizarin red S and the formation of mineralized nodules measured as described above.
Effects of EMD, LMW, and HMW on the Smad pathway of osteogenic regulation
Smad1/5/8, an inactive cytoplasmic protein complex, is activated by phosphorylation to p-Smad1/5/8 and then translocated to the nucleus, where it functions as a transcription factor for the BMP gene, thereby stimulating osteogenic activity.26,28 To determine the effects of EMD, LMW, and HMW on Smad1/5/8 activation to p-Smad1/5/8, flow cytometry (FCM) was used to examine changes in the relative levels of p-Smad1/5/8, and immunocytochemistry was carried out to examine intracellular localization, as described previously.18,19
Briefly, PDL cells were incubated in 24-well plates in GM for 3 days, and GM was then replaced with low-glucose-DMEM with 1% FBS for a further period of 48 h to reduce the levels of endogenous growth factors. To eliminate the presence of any exogenous growth factors, the culture medium was replaced with (1) completely serum-free OM containing low-glucose-DMEM, 0.1 mM L-ascorbic acid 2-phosphate, 10 mM β- glycerophosphate, and 10 nM dexamethasone (all from Sigma); (2) serum-free OM+EMD (100 μg/mL); (3) serum-free OM+LMW (100 μg/mL); and (4) serum-free OM+HMW (100 μg/mL), and further cultured for 1 h. For the FCM, after treatment of the PDL cells with EMD, LMW, and HMW as above, the cells were detached with 0.05% (w/v) trypsin and 0.05 mM (w/v) EDTA (trypsin-EDTA) for 5 min at room temperature, fixed with 1% paraformaldehyde, permeabilized with 0.1% Triton X, and blocked for 20 min using 10% normal goat serum (NGS). Cells were then incubated with 1:100 rabbit polyclonal anti-p-Smad1/5/8 (Insight Biotechnology, London, United Kingdom) for 1 h. After washing, the cells were further incubated with fluorescein isothiocyanate (FITC)-conjugated goat anti-rabbit (1:200) for 1 h at room temperature. FITC-conjugated normal rabbit IgG was used as a negative control. The cells were centrifuged and re-suspended in 200 μL of PBS and the fluorescence intensity of 10,000 individual cells was measured using a flow cytometer (FACScan; Becton-Dickinson). Data were analyzed using the WinMDI 2.8 software program.
For immunocytochemistry, after treatment of the PDL cell monolayers with EMD, LMW, and HMW as above, the cells were fixed with 4% paraformaldehyde for 15 min at room temperature and permeabilized using 0.1% Triton X (Sigma) for 15 min at room temperature. They were then treated with 10% NGS in PBS for 1 h, followed by incubation for 1 h at room temperature with 1:100 rabbit polyclonal anti-p-Smad1/5/8 (Insight Biotechnology). After washing, FITC-conjugated goat anti-rabbit was added (1:200) for 1 h at room temperature. Nuclei were stained with Hoechst stain, and p-Smad1/5/8-positive cells were visualized by their green fluorescent staining.
Smad6, a negative regulator of BMP-mediated osteogenic differentiation, exerts its activity by binding to and inhibiting the phosphorylation of the Smad1/5/8 complex.25,28 This prevents translocation to the nucleus and thereby prevents its function as a BMP transcription factor, as noted above. 25 To examine the effects of EMD, LMW, and HMW on the Smad6 gene, the PDL cells were cultured in OM in the presence of these proteins (all used at 100 μg/mL) and gene expression assessed by conventional PCR on day 4, as previously described. 18 GAPDH, a housekeeping gene, was used as an endogenous control. The following are the sequences of the forward and reverse primers (from Sigma-Genosys, Pampisford, United Kingdom)—Smad6 forward: AAAGACGCACTTTGGCTTA, reverse: CGAATACTTTATTATCGAGTGACTG; GAPDH forward: CCACCCATGGCAAATTCCCATGGCA, reverse: CTGGACGGCAGGTCAGGTCCACC. Smad6 primers were used at an annealing temperature of 57°C for 30 cycles and GAPDH primers were used at an annealing temperature of 55°C for 25 cycles.
Identification of the bioactive components of EMD, LMW and HMW
As noted above, TRAP is a naturally occurring 45-amino-acid proteolytic product of the full-length amelogenin protein, and is the major component of LMW.1,16 LRAP is a naturally occurring 56-amino-acid spliced form of amelogenin, and is a component of HMW. The presence of TRAP and LRAP in LMW and HMW, respectively, was confirmed by MALDI-TOF analysis (data not shown). By sequence comparison, we have identified that although TRAP and LRAP have an identical 33-amino-acid N-terminal sequence, the TRAP has a unique 12-amino-acid C-terminal sequence (designated TCT), while LRAP has a unique 23-amino-acid C-terminal sequence (designated LCT). To examine the biological activities of these peptides, TCT and the parent peptide TRAP, and LCT and the parent peptide LRAP were chemically synthesized (Insight Biotechnology) (Table 3), diluted in cell-culture-grade water (Thermo Scientific, Basingstoke, United Kingdom), and 1 μg/mL of each added directly to the PDL cells, clone 7, AB cells, and BMSC when they reached ∼90% confluence. These concentrations were determined from their dose effects (from 0.1 to 100 μg/mL) on terminal osteogenesis on PDL cells in vitro, as described above.
Statistical analysis
The PCR data are shown as the mean fold-change±standard error (±SE) of three separate experiments compared with that of control cells cultured in OM alone (defined as 1.0). Alizarin red staining intensity is shown as the relative staining intensity±SE of three separate experiments compared with OM alone (defined as 1.0). FCM data are presented as the mean average fluorescence intensity (AFI±SE) of three separate experiments. One-way ANOVA was used to assess statistical significant differences followed by the post hoc Bonferroni test for multiple comparisons (p<0.05) between the means (SPSS 11.0 software, Chicago, IL).
Results
Effects of EMD and the EMD fractions on terminal differentiation of bone-forming cells
Increasing concentrations of the EMD preparation used in the present study were found to progressively stimulate the formation of bone-like mineralized nodules by PDL cells, a key indicator of their ability to undergo terminal osteogenic differentiation in vitro (Fig. 1), with 100 μg/mL of EMD the alizarin red staining intensity increasing by 330% compared with control PDL cells cultured in OM alone (p<0.05). In marked contrast, increasing concentrations of LMW decreased alizarin red staining, with 100 μg/mL resulting in a 30% reduction in staining intensity (p<0.05; Fig. 1). Notably, as with EMD, increasing concentrations of HMW progressively increased terminal osteogenic differentiation of the PDL cells, with 100 μg/mL resulting in a level that was 580% higher than found when the cells were cultured in OM alone, as shown in Figure 1 (p<0.05). These results thus demonstrate that 100 μg/mL of EMD and HMW are stimulatory, whereas 100 μg/mL of LMW is inhibitory to terminal osteogenic differentiation of PDL cells. Since EMD and the EMD fractions were found to have the most pronounced effect on PDL osteogenesis at 100 μg/mL, this concentration was used throughout the experiments reported below.

Effects of Emdogain® (EMD), low molecular weight (LMW) and high molecular weight (HMW), on terminal osteogenic differentiation of periodontal ligament (PDL) and other bone-forming cells. Alizarin red staining of the cells cultured for 3 weeks in osteogenic medium (OM) in the presence of EMD, LMW, and HMW at concentrations 1, 3, 10, 30, and 100 μg/mL for the PDL cells and 100 μg/mL for the PDL clone 7, alveolar bone, and bone marrow-derived mesenchymal stem cells (BMSC). Alizarin red staining intensity is shown as the relative staining intensity±standard error (SE) of three separate experiments using three different healthy donors compared with OM alone (defined as 1.0). *Statistically significant difference compared with OM alone p<0.05. Color images available online at www.liebertonline.com/tea
EMD and the EMD fractions were also found to have similar effects on other bone-forming cells, as shown in Figure 1. Thus, when PDL clone 7 (an osteogenic precursor/stem cell-like clone isolated from adult human PDL) 20 was cultured in OM in the presence of 100 μg/mL of EMD, there was a significant increase (120%; p<0.05) in alizarin red staining intensity observed compared with OM alone. In contrast, in the presence of LMW the staining intensity decreased by 40% compared with OM alone (p<0.05), while, as with EMD, the presence of HMW resulted in a 350% increase in staining intensity (Fig. 1). Similar effects on mineralization were also observed with both AB and BMSC cells, which were stimulated by EMD (staining intensity 2.6 and 2.0, respectively; p<0.05) and also by HMW (270% and 180% increase, respectively; p<0.05) compared with OM alone. In contrast, LMW reduced the staining intensities of these two types of bone-forming cells by 30% and 40%, respectively, compared with OM alone (p<0.05). Thus, the differential effects of the EMD fractions were exerted not only on the heterogeneous PDL cell population but also on the isolated osteogenic clone, AB cells, and the BMSC.
Effects of EMD, LMW, and HMW on the expression of bone-associated markers
The results in Table 1 show that EMD significantly upregulated the expression of the early osteogenic transcription factor Runx2 after 4 days (approximately twofold), whereas the presence of LMW had no effect compared with OM alone. As with EMD, HMW also upregulated Runx2 nearly twofold (p<0.05) (Table 1). Similarly, the intermediate OP and late osteogenic marker genes OC and BSP, and ALP activity were all found to be significantly upregulated by both EMD and HMW, whereas LMW significantly downregulated all of these genes except for OP (Table 1). These results thus demonstrate that although LMW had no effect on the early Runx2 and intermediate OP genes, it, nevertheless, suppressed the key late markers OC, BSP, and ALP, and it also suppressed the terminal osteogenic differentiation of PDL and other bone-forming cells as described above. Moreover, it also suggests that LMW and HMW, two different components of EMD, have completely opposite effects on bone-forming cells.
PDL cells were cultured in OM in the presence of EMD, LMW, and HMW for up to 2 weeks. Q-PCR analysis was carried out for the early osteogenic transcription factor Runx2, intermediate matrix marker gene OP, and the late matrix marker genes OC and BSP. ALP activity was also measured at day 14. The PCR data are shown as the mean fold-change±SE of three separate experiments using three different healthy donor cells compared with that of control cells cultured in OM alone (defined as 1.0).
The values shown are relative to the values obtained with OM alone.
Statistically significant difference compared with OM alone (p<0.05).
EMD, Emdogain®; LMW, low molecular weight; HMW, high molecular weight; Runx2, runt-related transcription factor-2; OP, osteopontin; OC osteocalcin; ALP, alkaline phosphatase; BSP, bone sialoprotein; PDL, periodontal ligament; OM, osteogenic medium; Q-PCR, quantitative-polymerase chain reaction; SE, standard error.
Effects of noggin and UB505124 on EMD-, LMW-, and HMW-mediated osteogenesis
To determine whether the action of EMD and the EMD fractions might occur via the BMP- and TGFβ-mediated pathways of osteogenic differentiation, the PDL cells were incubated in OM in the absence and presence of noggin, a BMP pathway inhibitor, and UB505124, a TGFβ pathway inhibitor. The results in Table 2 show that the presence of noggin and of UB505124 alone had no effect on the alizarin red staining intensity of control PDL cells cultured in OM. As before, when the cells were cultured in the presence of EMD alone the staining intensity significantly increased to 2.1. However, the concurrent presence of noggin or UB505124 in the EMD cultures markedly reduced the alizarin red levels compared with cultures incubated with EMD alone (from 2.1 to 1.0 and 1.4, respectively; p<0.05) (Table 2). Moreover, the presence of noggin and UB505124 similarly further reduced the low mineralization activity obtained in the presence of LMW alone (0.1 and 0.3, respectively, compared with 0.6; p<0.05). Notably, the high level of mineralization observed in the presence of HMW (3.5) was unaffected by noggin and UB505124 (3.1 and 3.1, respectively). These results thus suggest that the osteogenic activity of EMD and LMW, but not of HMW, is mediated via BMP- and TGFβ-dependent pathways.
Alizarin red staining of PDL cells cultured for 14 days in OM in the presence of EMD (100 μg/mL), LMW (100 μg/mL), and HMW (100 μg/mL) with and without noggin and UB505124. The numbers are alizarin red staining intensity±SE of three separate experiments using three different healthy donor cells.
Statistically significant difference between noggin-treated verses untreated (p<0.05).
Statistically significant difference between UB505124-treated verses untreated (p<0.05).
Effects of EMD, LMW, and HMW on the Smad pathway of osteogenic regulation
The Smad pathway has previously been shown to modulate BMP-mediated osteogenic differentiation.19,26 In view of the apparent effect of EMD and the EMD fractions on the BMP pathway, shown above, the effect of these proteins on the relative level of p-Smad1/5/8, the active form of the BMP transcription factor, was examined by FCM. As shown in Figure 2A, EMD and HMW were found to have no effect on p-Smad1/5/8 compared with serum-free OM alone, whereas the presence of LMW significantly reduced this activated complex by 42% (p<0.05). Moreover, immunocytochemical studies demonstrated that ∼50% of the cells in the presence of serum-free OM alone and with EMD and HMW expressed p-Smad1/5/8, whereas in the presence of LMW no p-Smad1/5/8-positive cells were detected (Fig. 2B). These results thus indicate that LMW suppresses activation of p-Smad1/5/8, resulting in inhibition of osteogenic differentiation.
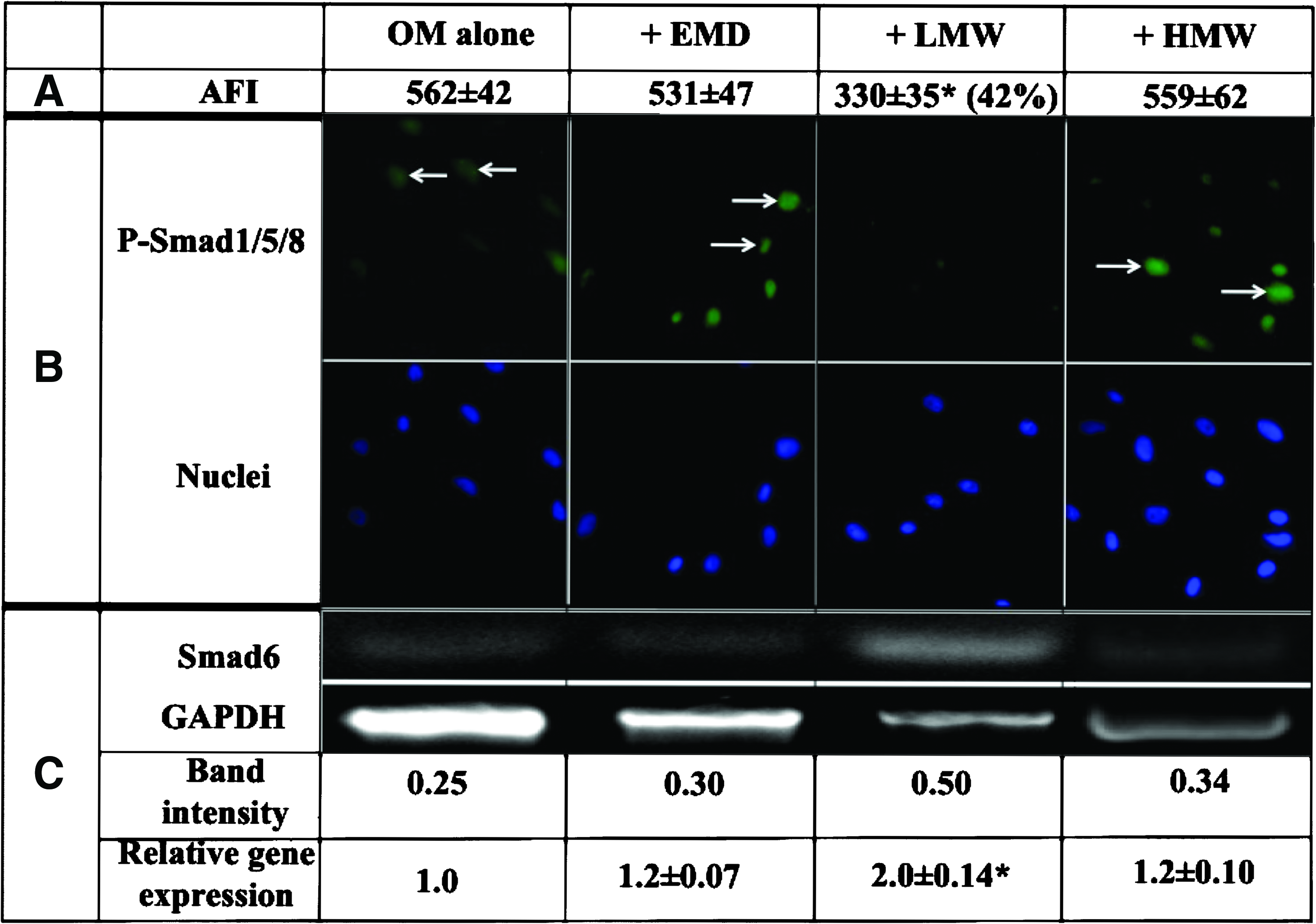
Effects of EMD, LMW, and HMW on the Smad pathway.
Smad6 has previously been shown to be a negative regulator of BMP-mediated osteogenesis that exerts its activity by binding to and preventing the phosphorylation and subsequent nuclear translocation of Smad1/5/8. To determine whether the apparent inhibitory effect of LMW on the level of p-Smad1/5/8, as shown above, was mediated via Smad6, the expression of the Smad6 gene was examined in the absence and presence of EMD and the EMD fractions. The results in Figure 2C show that while EMD and HMW did not cause any marked change in Smad6 gene expression, in the presence of LMW the Smad6 gene was significantly upregulated (twofold; p<0.05) compared with OM alone. These results thus suggest that LMW may inhibit osteogenic activity via stimulating Smad6, a negative regulator of the BMP pathway.
Effects of synthetic amelogenin peptides on bone-forming cells
The amelogenin-derived TRAP and LRAP have previously been identified as components of LMW and HMW, respectively, as noted above. In view of the effects of these fractions on osteogenic differentiation, chemically synthesized TRAP and LRAP were obtained and examined for their effect on the terminal osteogenic differentiation of bone-forming cells. As with LMW, synthetic TRAP (1 μg/mL) significantly reduced bone-like nodule formation and the alizarin red staining intensity of the PDL cells, clone 7, AB cells, and BMSC (40%–60% reduction; p<0.05) compared with OM alone (Table 3). In contrast, as with HMW, 1 μg/mL of LRAP markedly stimulated bone-like nodule formation and increased the staining intensity of the PDL cells, clone 7, AB cells, and BMSC (between 150% and 340% increases; p<0.05) compared with OM alone. These results thus demonstrate that the chemically synthesized amelogenin isoforms TRAP and LRAP exhibit activities that are similar to the EMD fractions LMW and HMW; further, these data suggest that TRAP and LRAP are the active components of EMD that differentially regulate osteogenic differentiation.
Alizarin red staining intensity of the PDL cells, clone 7, AB cells, and BMSC cultured for 3 weeks in OM in the presence of synthetic TRAP (1 μg/mL), LRAP (1 μg/mL), TCT (1 μg/mL), and LCT (1 μg/mL). Alizarin red staining intensity is shown as the relative staining intensity±SE of three separate experiments using three different healthy donor cells compared with OM alone (defined as 1.0).
Statistically significant difference compared with OM alone (p<0.05).
AB, alveolar bone; BMSCs, bone marrow-derived mesenchymal stem cells; TRAP, tyrosine-rich amelogenin peptide; LRAP, leucine-rich amelogenin peptide; TCT, C-terminal 12 amino acid sequence; LCT, C-terminal 23 amino acid sequence.
Alignment of the TRAP and LRAP isoforms shows that the 33-amino-acid sequence at the N-terminal of TRAP and of LRAP is common to each other (Table 4). In view of the differential activities of TRAP and LRAP, it was considered that this overlapping sequence is likely to be devoid of the specific bone-modulating activity exhibited by TRAP and LRAP. In experiments not reported here, we found that concentrations between 0.1 and 100 μg/mL of this null peptide had no effect on cell mineralization in vitro. In marked contrast, however, the 12-amino-acid C-terminal peptide (TCT) unique to the osteogenesis-inhibitory TRAP, and the 23-amino-acid C-terminal peptide (LCT) unique to the osteogenesis-stimulatory LRAP, were found to have potent but opposite osteogenic effects, as shown in Table 4. Thus, 1 μg/mL of TCT was found to strongly inhibit (between 30 and 50% reduction; p<0.05), whereas 1 μg/mL of LCT strongly stimulated terminal osteogenic differentiation of PDL cells, clone 7, AB cells, and BMSC (between 140% and 490% increases; p<0.05), compared with OM alone (Table 4). These results indicate that the unique C-terminal amino acid sequences of TRAP and LRAP, TCT and LCT, respectively, are likely to be the bioactive sequences responsible for the differential activities of the parent amelogenin-derived isoforms.
Amino acid sequences: −NH2→−COOH.
Discussion
A number of studies, including the present, have demonstrated that EMD stimulates bone-associated markers and bone-like nodule formation of PDL and other bone-forming cells.29–31 In contrast, in some reports EMD has also been shown to inhibit the expression of osteogenic genes and mineralized bone-like nodule formation in vitro10,32 and also in vivo.8,11,13 To delineate the putative osteomodulatory components, several attempts have been made to fractionate freshly isolated non-heat-treated porcine EMP.14,15 However, such partially purified preparations may sometimes contain growth factors such as BMP-2 and TGFβ1 that are likely to have pronounced effects on bone cell activity14,15,31 and have also been reported to contain components of differing molecular sizes, including LMW (<6 kDa) proteins.14,15,31
The present study has used two recently isolated fractions of EMD, LMW and HMW, obtained by carefully controlled industrial-scale methodologies. 16 The results obtained here demonstrated that 100 μg/mL of LMW, which comprises <6-kDa EMD proteins, mainly TRAP, strongly suppressed the mineralization of PDL as well as other bone-forming cells, including osteogenic precursor/stem cell clone 7, AB cells, and BMSC. Although LMW did not have any apparent effect on the early and intermediate osteogenic markers Runx2 and OP, respectively, the late markers OC, BSP, and ALP were strongly inhibited by this fraction. In contrast, the presence of HMW, which comprises >6-kDa EMD proteins and is devoid of LMW components, stimulated the expression of early, intermediate, and late osteogenic marker genes and also the terminal differentiation of the bone-forming cells. Thus, the sometimes variable outcome of osteogenic differentiation in vitro and bone regeneration in vivo observed using nonfractionated EMP and EMD8,11,13,29–32 may reflect at least partly differences between these different preparations.
It has previously been shown that the BMP-2 and TGFβ1 pathways are involved in the regulation of bone-forming cell activity.20,22,23,33–35 The present study therefore used noggin, a BMP-2 pathway inhibitor, and UB-505124, a TGFβ1 pathway inhibitor, to determine whether EMD and the EMD fractions might have acted via one or both of these signaling pathways. The results showed that in the presence of noggin the mineralization of the cells was reduced further than in the presence of LMW alone. This suggests that LMW may inhibit osteogenic differentiation at least partly via BMP-2-induced intracellular messengers, consistent with a previous report that in the mouse C2C12 cell line the LMW components in EMD were able to suppress BMP-2-dependent osteogenesis. 15 In addition, the results reported here demonstrate that LMW reduced the intracellular level of the activated BMP-2 transcription factor complex p-Smad1/5/8 in PDL cells and it also stimulated the expression of the Smad6 gene, which is known to act as a BMP-2 pathway negative regulator.25,28 These results thus suggest that LMW in EMD suppresses osteogenesis at least partly by upregulating the inhibitory Smad6 that binds to and blocks the phosphorylation and subsequent nuclear translocation of the Smad1/5/8 complex. In addition, it was observed here that in the presence of the TGFβ1 pathway inhibitor UB-505124 bone-like nodule formation by the cells was further reduced. Although the effects of EMD and the EMD fractions on Smad2/3 and Smad7, other factors involved in TGFβ-mediated osteogenesis,26,27 were not examined here, the present observations, nevertheless, suggest that the TGFβ signaling pathway may be at least partially involved in LMW-mediated inhibition of osteogenic differentiation.
In contrast to previous reports that BMP-2 and TGFβ1 growth factors may be present in a HMW EMD fraction (e.g., 10–20 kDa),14,15,31 in the present investigation it was observed that the stimulatory effects of HMW were unaffected by the BMP-2 and TGFβ1 inhibitors, suggesting that this fraction (6–20-kDa components) is unlikely to contain BMP-2- and TGFβ1-like activities. Moreover, it further indicates that HMW may stimulate bone via signaling pathway(s) independent of BMP-2 and TGFβ1, possibly such as those mediated by parathyroid hormone and the hedgehog and Wnt families.36,37
Since the TRAP and LRAP have been identified as components of LMW and HMW, respectively, the present study also investigated the effects of chemically synthesized TRAP and LRAP on bone-forming cells. As with LMW, synthetic TRAP suppressed, whereas, as with HMW, synthetic LRAP stimulated, the osteogenic differentiation of the PDL cells. Although a previous report has suggested that the C-terminal amino acid sequence of LRAP might be responsible for the bioactivity of the parent protein, 38 hitherto there has been no definitive evidence for the molecular identities of the active peptides. To delineate the specific sequences that are most likely to modulate osteogenic differentiation we compared the TRAP and LRAP amino acid sequences, which revealed that while the 33-amino acid sequence at the N-terminal of both the peptides are identical, the 12-amino acid (TCT) and the 23-amino acid (LCT) sequences at the C-terminals of TRAP and LRAP, respectively, are unique to each. The TCT and LCT sequences were therefore chemically synthesized and examined for osteogenic activity, and found to inhibit and stimulate bone-forming cells, respectively. Moreover, protein homology analysis of the TRAP and LRAP amino acid sequences, using the National Centre for Biotechnology Information (NCBI) protein database and BLAST software (NCBI, Bethesda, MD), showed that these TCT and LCT amino acid sequences were the domains that were the most conserved sequences of the TRAP and LRAP of >100 different animal species, further indicating their possible physiological importance in modulating the process of bone formation. These findings thus demonstrate, for the first time, that the C-terminal TCT and LCT amino acid sequences exhibit, at least in vitro, the osteogenic activities of the respective TRAP and LRAP amelogenin-derived parent peptides examined here.
A number of procedures and materials are available for achieving bone regeneration, including the use of different types of bone graft as well as the FDA-approved BMP-2/INFUSE, but their predictability and efficacy in terms of clinical outcome remain uncertain. Although the present study has not assessed the potential clinical significance of these two new peptides, the novel bioactive 23-amino acid LCT amelogenin sequence identified here can be produced synthetically in large quantities and with high purity, and might be capable of significantly improving therapeutic options for periodontal and orthopedic diseases, while the novel 12-amino acid TCT peptide might prove to be a valuable tool for limiting pathological bone cell growth.
Conclusions
Two main fractions of EMD, LMW and HMW, which contain the amelogenin isoforms TRAP and LRAP, respectively, elicited differential bone-forming responses in vitro. Furthermore, while the naturally occurring LMW and the synthetically produced TRAP and its unique C-terminal TCT sequence suppressed bone-forming cells, the naturally occurring HMW and the synthetically produced LRAP and its unique C-terminal LCT sequence were found to induce terminal differentiation of bone-forming cells. Although the in vivo functionality of there new peptides has thus far not been examined, the in vitro results reported in the present study suggest the possibility that they might have potential clinical benefit in promoting the repair and regeneration of bone defects arising from damage and disease.
Footnotes
Acknowledgments
H.D. Amin is grateful for the financial support provided by a Dorothy Hodgkin Postgraduate Award for research studies (jointly funded by the Engineering and Physical Sciences Research Council, United Kingdom, and by Institut Straumann) and the Research Discretionary Funds of the Periodontology Unit, UCL Eastman Dental Institute. Financial support was also provided by the NIHR Comprehensive Biomedical Research Centre and by the WCU Program of the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (No. R31-10069).
Disclosure Statement
The authors alone are responsible for the content and writing of this article. No competing financial interests exist.