
Abstract
With the aim of discovering new molecules for induction of bone formation and biomineralization, combination of bioinformatics and simulation methods were used to design the structure of artificial peptides based on proline-rich domains of enamel matrix proteins. In this study, the effect of such peptides on the differentiation toward the osteogenic lineage of human umbilical cord mesenchymal stem cells (hUCMSCs) was evaluated with or without osteogenic supplements (hydrocortisone, β-glycerol phosphate, and ascorbic acid) and compared to the effect of the commercially available enamel matrix derivative (EMD). It was hypothesized that the differentiation toward the osteogenic lineage of hUCMSCs would be promoted by the treatment with the synthetic peptides when combined with differentiation media, or it could even be directed exclusively by the synthetic peptides. Osteoinductivity was assessed by cell proliferation, bone morphogenetic protein-2 secretion, and gene expression of osteogenic markers after 1, 3, and 14 days of treatment. All peptides were safe with the dosages used, showing lower cell toxicity. P2, P4, and P6 reduced cell proliferation with growing media by 10%–15%. Higher expression of early osteoblast markers was found after 3 days of treatment with EMD in combination with osteogenic supplements, while after 14 days of treatment, cells treated by the different synthetic peptides in combination with osteogenic supplements showed higher osteocalcin mRNA levels. We can conclude that osteogenic differentiation of hUCMSCs is promoted by short-term EMD treatment in combination with osteogenic supplements and by long-term treatment by the synthetic peptides in combination with osteogenic supplements, showing similar results for all the peptide variants analyzed in this study.
Introduction
In this study, two other variants of our designed consensus peptide (peptide 4 and peptide 5) and the analogous peptide to human amelogenin (peptide 6) were investigated for the osteoinductive potential of hUCMSCs, in a continuing effort to advance bone regeneration therapies and follow up our previous research findings.
The new variants of the consensus peptide have been designed with respect to different properties of composing amino acids other than prolines. Several serine residues have been integrated in the sequences of peptides 4 and 5. Breakdown of bone and connective tissue matrix is mainly due to matrix metalloproteinases and serine proteases. 3 Therefore, a serine rich peptide with prolines would compete through substrate inhibition and slow down bone resorption. Moreover, apart from serines, replacement by cysteine residues has been studied in the peptide 5. Cysteines comprise sulfur atoms that may bind to metal surfaces by forming sulfur bridges; 4 therefore, this peptide would be optimized to functionalize implant surfaces to favor osteointegration and to promote osteoblastic differentiation.
We have used hUCMSCs to evaluate the effect of the synthetic peptides. hUCMSCs are undifferentiated cells that have the potential to differentiate into osteoblasts among a variety of cell lineages. To promote the osteogenic differentiation of hMSCs, dexamethasone, β-glycerolphosphate, and ascorbic acid (osteogenic supplements) are added to the culture media. 5 We hypothesized that the differentiation toward the osteogenic lineage of hUCMSCs could be synergistically enhanced by the treatment with the synthetic peptides when combined with differentiation media or it could even be directed exclusively by the synthetic peptides. To address this hypothesis, hUCMSCs were treated with the different synthetic peptides for 1, 3, and 14 days in both the presence and absence of osteogenic supplements. The osteoinductivity and potential synergistic effects were evaluated by cell proliferation, gene expression of osteogenic markers, and bone morphogenetic protein-2 (BMP-2) secretion to the culture media.
Materials and Methods
Peptide design
The precise selection of the proline-rich regions from amelogenin and ameloblastin was described in details in a previous study. 2 We designed our peptides with constraints on length (25 AA), conservation of proline positions, and with systematic variation of other composing amino acids (Table 1).
P2, Peptide 2; P4, Peptide 4; P5, Peptide 5; P6, Peptide 6; S, Ser; P, Pro; L, Leu; V, Val; Q, Gln; M, Met; H, His; C, Cys.
3D models of the designed peptides
The designed peptides (P2, P4, P5, and P6) should differ in their physical, chemical, and structural properties due to the different amino acid compositions, except the conserved proline positions in the sequence. We utilized a recently developed Web-based method of peptide design 6 to explore possible correlation between the structure and biological response to the particular peptide. The sequences of three designed peptides were modeled by MOBYLE server 7 and only the best energy conformer of each peptide was used for further analysis and optimization. The three resulted peptides and their structural and dynamical properties were explored by molecular dynamic simulations (MD) by means of AMBER 9.0 package 8 using ff99SB force field 9 in TIP3 10 explicit water environment. MD simulations were performed according to the following conditions: dielectric constant eps ∼ r, 508 solvent molecules, equilibration–100ps (0–300K), production phase–500ps, and NPA Hamiltonian equations of motion.
Preparation of peptides and enamel matrix derivative
Enamel matrix derivative (EMD) was kindly supplied by Straumann GMbH (Basel, Switzerland). EMD was dissolved to 10 mg/mL in 0.1% acetic acid in phosphate-buffered saline (PBS). Aliquots were prepared and stored at −20°C until use. Four synthetic peptides (Table 1) were designed and purchased from Eurogentec (Seraing, Belgium). One vial containing 10–21 mg of the selected synthetic peptides was delivered in a freeze-dried pellet form and dissolved to 5 mg/mL in 0.1% acetic acid in PBS. Final concentrations of 50 μg/mL of EMD and peptides were used to assay their effect on cytotoxicity, proliferation, and differentiation of hUCMSCs cells in an in vitro system.
hUCMSCs isolation
hUCMSCs were isolated from umbilical cords obtained in the process of human umbilical cord blood donation under the Concordia Cord Blood Donation Program. The samples were obtained after informed consent and with the approval of the Ethical Committee of Balearic Islands (CEIC-IB). To isolate hUCMSCs, the cord was rinsed several times with sterile saline, the cord blood was drained, and clots flushed from the vessels. Next, the cord was cut into small pieces and incubated with 0.075% collagenase type IA (Sigma, St Louis, MO) for 2 h. Then, 0.125% trypsin was added for 30 min with gentle agitation at 37°C. The digested mixture was then passed through a 100 μm filter to obtain cell suspensions. Cell pellets were resuspended in NH4Cl-based erythrocyte lysis buffer, incubated for 10 min at room temperature, and washed in PBS. Finally, the cells were resuspended in Dubelcco's modified Eagle's medium with low glucose and Glutamax (DMEM-LG; Gibco, Grand Island, NY) and 20% FBS (HyClone, Thermo Scientific, Logan, UT) and plated in noncoated 25 cm2 cell culture flask. Cultures were maintained in a humidified atmosphere with 5% CO 2 at 37°C. After 3 days of culture, the medium was replaced and nonadherent cells were removed. The medium was changed twice weekly thereafter. Once 80% confluence had been reached, adherent cells were replated at a density of 1×104 cells/cm2.
Cell surface profile of the hUCMSCs used for the studies was confirmed by standard flow cytometry as previously described 2 ; cells did not express CD34, CD45, CD31, and HLA-DR, whereas were positive for CD105, CD90, and CD73, showing a typical profile of MSCs. 11
Cell culture
hUCMSCs were seeded in 24-well plates and grown to confluence in a “growing media” consisting of DMEM/Glutamax/LG, supplemented with penicillin (50 IU/mL), streptomycin (50 μg streptomycin/mL), and 20% fetal bovine serum (HyClone). Cells were cultured at 37°C in a 95% air–5% CO2 atmosphere.
At confluence (designated as day 0), cells were treated with 0.1% acetic acid-PBS (control), with synthetic peptides (50 μg/mL) or with EMD (50 μg/mL), either in “growing media” or in “differentiation media” consisting of growing media supplemented with hydrocortisone (200 nM), ascorbic acid (50 μg/mL), and β-glycerophosphate (10 mM). Media were replaced every Monday and Thursday. Treatments were added after changing the media to keep the concentration of peptides and EMD in solution throughout the whole experiment. Culture media were collected after 1 day of treatment to test cytotoxicity, and after 14 days to analyse BMP-2 release. Cells were harvested at days 1, 3, and 14 to analyze gene expression of several osteoblast differentiation markers using real-time reverse transcriptase–polymerase chain reaction (RT-PCR).
Determination of cell viability: lactate dehydrogenase activity
Lactate dehydrogenase (LDH) activity in the culture media was used as an index of cell death. LDH activity was determined spectrophotometrically after 30 min incubation at 25°C of 100 μL of culture and 100 μL of the reaction mixture by measuring the oxidation of nicotinamide adenine dinucleotide (NADH) at 490 nm in the presence of pyruvate, according to the manufacturer's kit instructions (Roche Diagnostics, Mannheim, Germany). Results were presented relative to the LDH activity in the medium of cells cultured in growing media and treated with 0.1% acetic acid-PBS (low control, 0% of cell death) and of cells treated with triton X-100 1% (high control, 100% of death), using the equation:
Cell proliferation
Proliferation rate was analyzed after the incorporation of bromodeoxyuridine (BrdU) as described by the manufacturer (Roche Diagnostics). To test the effect of the different synthetic peptides and EMD on cell proliferation, 2×103 cells were seeded in each well (96-well plates) for 24 h. After cell attachment, culture medium was changed and different treatments peptide/EMD (50 μg/mL) and BrdU were added to the wells. Cells were cultured for additional 4 days to allow the incorporation of BrdU to the proliferating cells. Media were replaced every Monday and Thursday. Treatments and BrdU were added after changing the media to keep the concentration of peptides and EMD in solution throughout the whole experiment. Values were expressed as a percentage of control cells, which were set to 100%.
RNA isolation and RT-PCR analysis
Total RNA was isolated using the NucleoSpin® TriPrep kit (Macherey-Nagel), following the instructions of the manufacturer. Total RNA was quantified at 260 nm using a Nanodrop spectrophotometer (NanoDrop Technologies, Wilmington, DE).
The same amount of total RNA (0.27 μg) from each sample was reverse transcribed to cDNA using High Capacity RNA-to-cDNA kit (Applied Biosystems, Foster City, CA) according to the protocol of the supplier. Each cDNA was diluted and aliquots were stored at −20°C until the PCR reactions were carried out.
Real-time PCR was performed using the LightCycler FastStart DNA Master PLUS SYBR Green I (Roche Diagnostics) following the manufacturer's instructions. Real-time PCR was done for two reference genes: glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and β-actin, and five target genes: collagen type I (COLL-I), alkaline phosphatase (ALP), runt-related transcription factor 2, (RUNX2), decorin (DCN), and osteocalcin (OC). A negative control without cDNA template was run in each assay. Each reaction contained 500 nM of the corresponding oligonucleotide primers (h-β-actin-F: 5′-AAGGGACTTCCTGTAACAATGCA-3′; h-β-actin-R: 5′-CTGGAACGGTGAAGGTGACA-3′; h-GAPDH-F: 5′-TGCACCACC-AACTGCTTAGC-3′; h-GAPDH-R: 5′-GGCATGGACTGTGGTCATGAG-3′; h-COLL-I-F: 5-CCTGACGCACGGCCAAGAGG-3′; h-COLL-I-R: 5′- GGCAGGGCTCGGGTTTCCAC-3′; h-ALP-F: 5′-CCGCTATCCTGGCTCCGTGC-3′; h-ALP-R: 5′-GGTGGGCTGGCAGTGG-TCAG-3′;h-RUNX2-F: 5′- CTGTGCTCGGTGCTGCCCTC-3′; h-RUNX2- R: 5′- CGTTACCCGC-CATGACAGTA-3′; h-DCN-F: 5′- ATCTCAGCTTTGAGGG-CTCC-3′; h-DCN- R: 5′- GCCTCTCTGTTGAAACGGTC-3′; h-OC-F: 5′-GAAGCCCAGCGGTGCA-3′;h-OC-R: 5′- CACTACCTCGCTGCCCTCC-3′; hWNT7B-F: 5′- TCTGCTTTGGCGTCCTGTACGT-3′; hWNT7B-R: 5′- CCGACTCTGGCAGATGGCACG-3′), 5 μL of LightCycler FastStart DNA Master PLUS SYBR Green I (Roche Diagnostics), and 3 μL of cDNA in a final volume of 10 μL.
The amplification program consisted of a preincubation step for denaturation of the template cDNA (5 min, 95°C), followed by 45 cycles consisting of a denaturation step (10 s, 95°C), an annealing step (10 s, 60°C), and an extension step (10 s, 72°C) for β-actin, GAPDH, COLL-I, DCN, RUNX2, and WNT7B; or followed by 45 cycles consisting of a denaturation step (10 s, 95°C), an annealing step (5 s, 68°C), and an extension step (12 s, 72°C) for ALP and OC. After each cycle, fluorescence was measured at 72°C. A negative control without a cDNA template was run in each assay.
To allow relative quantification after PCR, standard curves were constructed from the standard reactions for each target and housekeeping genes by crossing points (Cp) values, that is, the cycle number at which the fluorescence signal exceeds background, versus log cDNA dilution. The Cp readings for each of the unknown samples were used to calculate the amount of either the target or the reference relative to a standard curve, using the Second Derivative Maximum Method with the LightCycler analysis software 4.0 (Roche Diagnostics). Relative mRNA levels were calculated as the ratio of relative concentration for the target genes relative to that for the mean between the two reference genes (GAPDH and β-actin). Values were expressed as a percentage of control cells cultured in growing media, which were set to 100%.
Release of BMP-2 into the cell culture media
BMP-2 secreted to the culture media after 14 days of treatment was analyzed by enzyme-linked immunosorbent assay (ELISA). Aliquots from the culture media were centrifuged at 1800 rpm for 5 min at 4°C and supernatants were used for BMP-2 determination following instructions described by the manufacturer (Quantikine Immunoassay; R&D Systems, Minneapolis, MN). Total protein in the culture media for each sample was determined using a BCA protein assay kit (Pierce, Rockford, IL) and used for correction of BMP-2 secretion, expressed in pg BMP-2/mg protein.
Statistical analyses
The data were presented as mean values±standard error of the mean. Differences between groups were assessed by student's t-test or by paired t-test and using a statistical software package (SPSS, Chicago, IL). Results were considered statistically significant at the p-values ≤0.05.
Results
Peptide structures
Models of all three new studied peptides showed signature of compact, well packed structures lacking secondary structure elements, as expected due to the rich content of prolines. We hypothesized in our previous paper that the effect of the peptides could be correlated with exposition of their C termini containing conserved proline-rich region (PPLPP). As one can see on the equilibrated structures of the three modeled peptides (Fig. 1), all of them show similar general properties (compactness and C-terminal exposition) and they also share general structural characteristic of their PPLPP region. Peptides 4 and 6 have similar overall topology—two distinct loops making the structures similarly compact. Regarding the compactness (could be measured as radius of gyration) the peptide 5 is not an exception; but it has different topology of loops and makes possible a contact between C an N terminus. All of them (and this is in agreement with previously reported P2 peptide property) expose their terminal PPLPP sequence stretch in a way suitable for further interactions. They are not exactly in the same conformational state regarding their phi/psi torsions of PPLPP backbone, but it is quite visible that compactness of the molecules helps their C terminus to be conformational, similar, and interacting most probably by the same way. Having neither direct structural evidences about the isolated peptides conformations nor about their modus of interaction we cannot unambiguously state that the described structural features are directly responsible for the effect of the peptides in question, but they seem to be good candidates for explanation of their action.
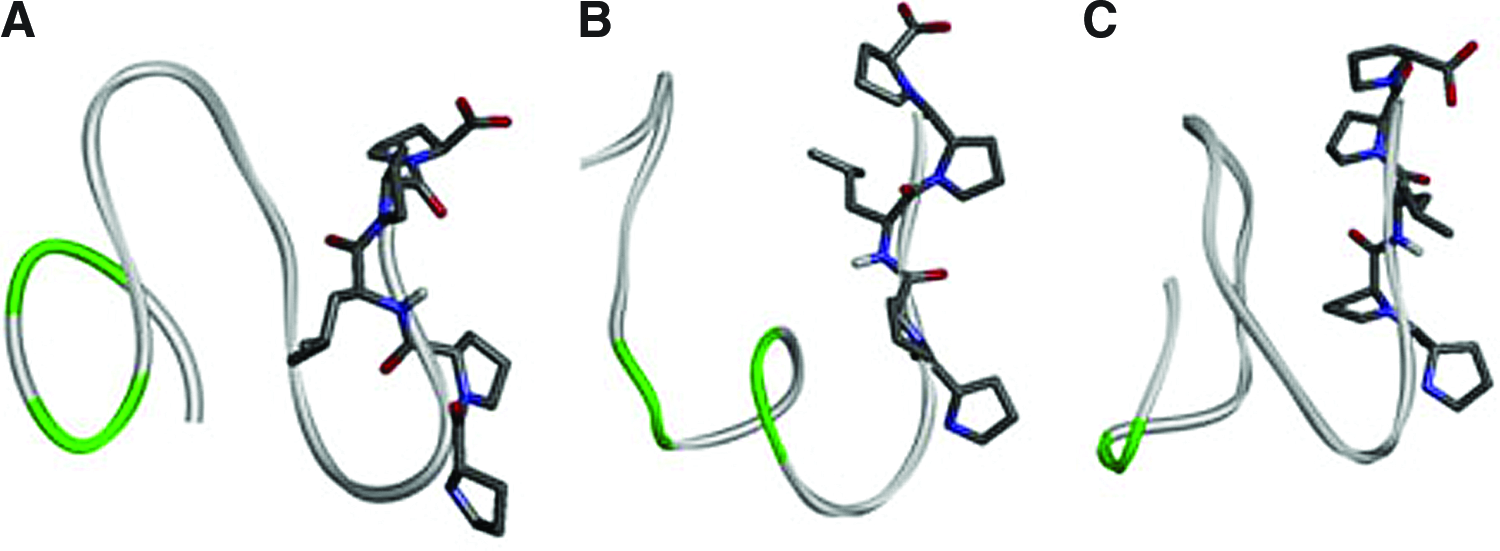
PepFold predicted 3D structures of
Cell viability: LDH activity
The LDH activity in the culture media was used as an index of toxicity (Fig. 2). After 24 h of treatment, peptide 2 improved cell viability of hUCMSCs compared with control in both, growing and differentiation media. Although values did not reach statistical significance, all treatments (EMD and peptides) improved cell viability compared with control in both media.
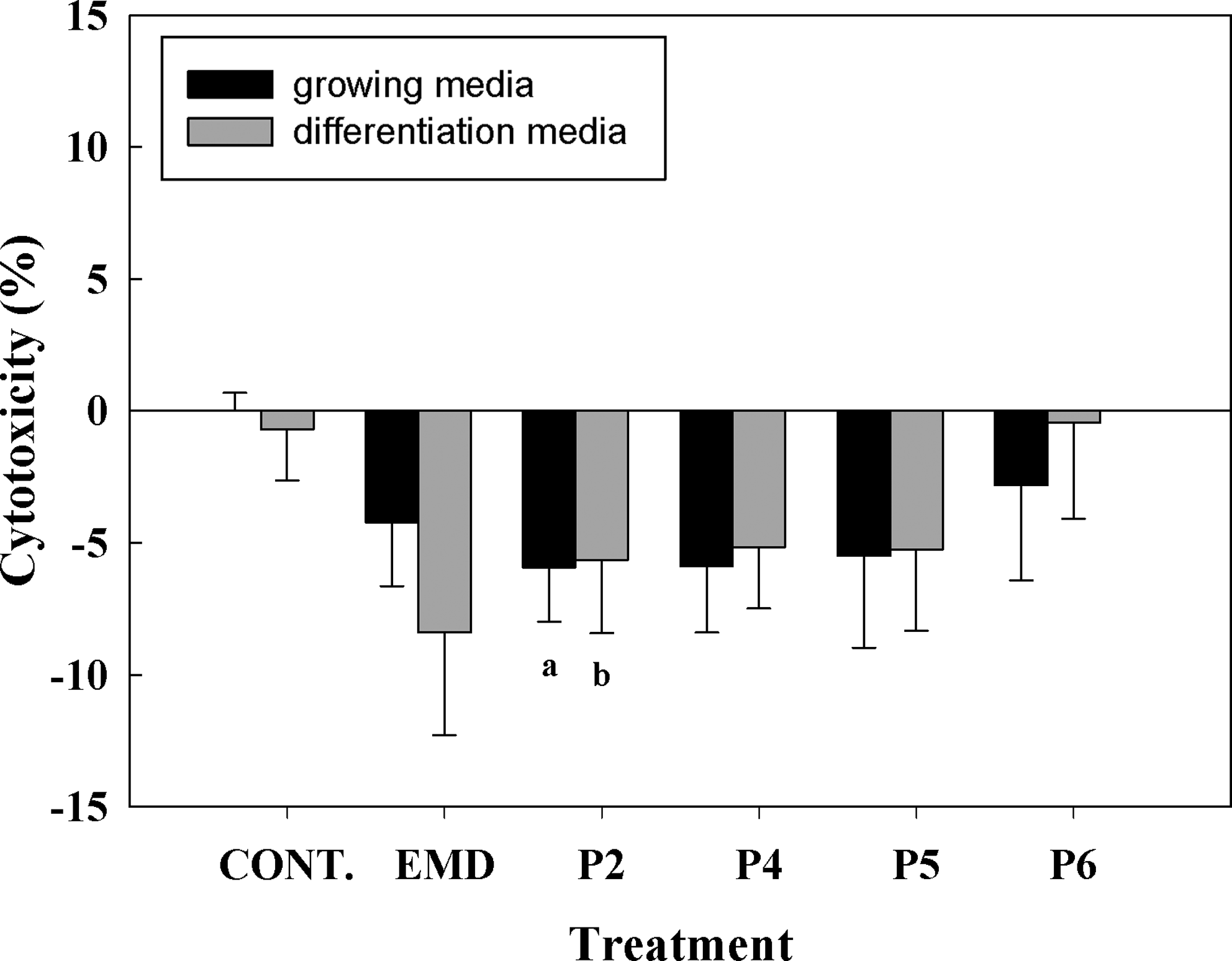
Lactate dehydrogenase activity measured from culture media collected after treatments with the different peptides (50 μg/mL) or enamel matrix derivative (EMD; 50 μg/mL) for 24 h. Values represent the mean±standard error of the mean (SEM) (n=6 from three different donors). Paired t-test: (a) p≤0.05 versus control growing media, (b) p≤0.05 versus control differentiation media.
Cell proliferation
The effect on cell proliferation of 4 days of treatment either with EMD or with synthetic peptides was assessed by analyzing the incorporation of BrdU into the cells cultured in growing media (Fig. 3). No differences were found when comparing EMD-treated cells to control cells. Treatment with P2, P4, and P6 reduced cell proliferation compared with control cells. Moreover, P6 showed significant lower proliferation compared with EMD.
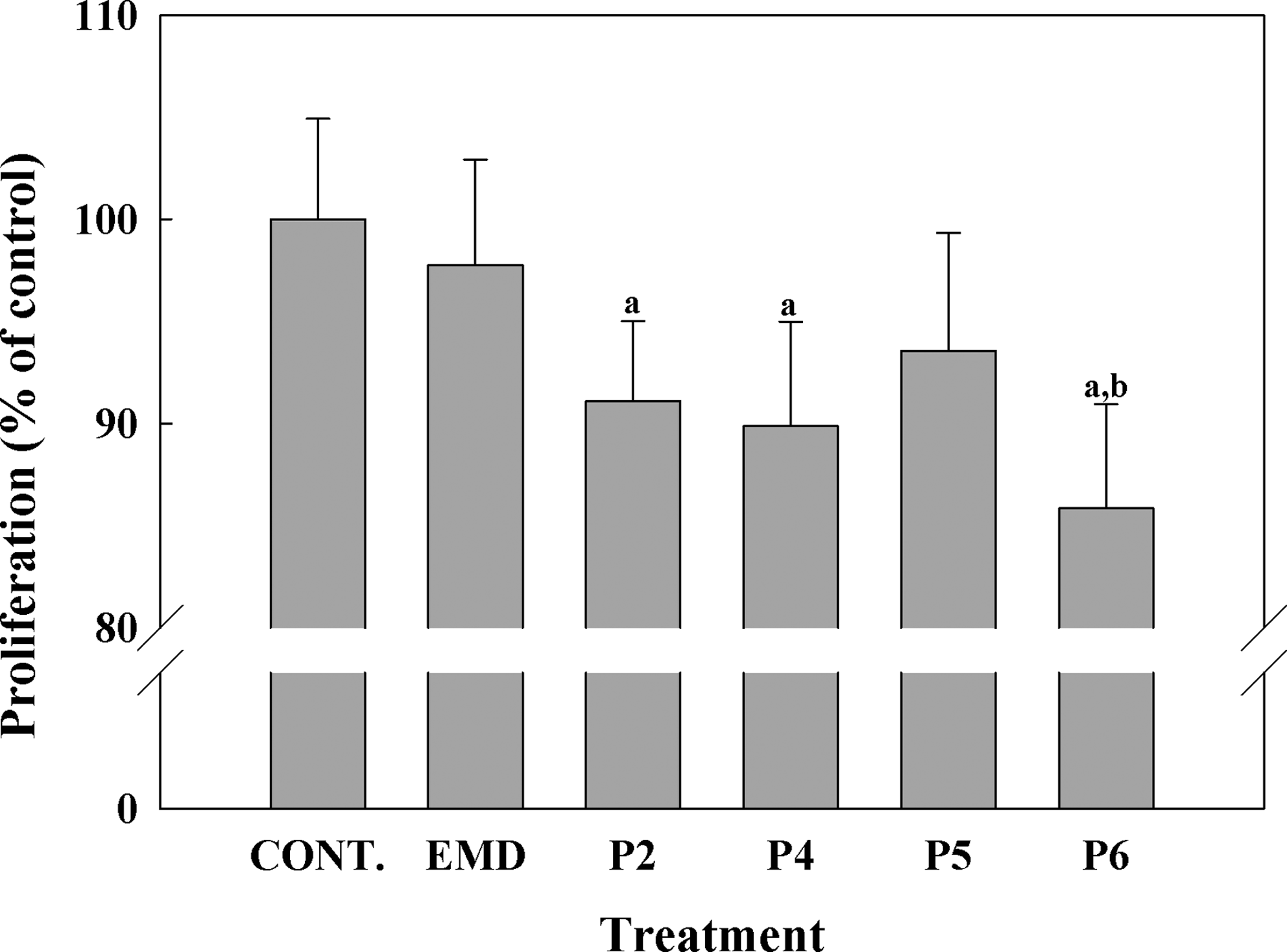
Proliferation of human umbilical cord mesenchymal stem cells (hUCMSCs) cells incubated with the different peptides (50 μg/mL) or EMD (50 μg/mL) for 4 days. Cell proliferation was measured by bromodeoxyuridine incorporation into the cell for 4 days. Values were expressed as a percentage of control cells, which were set to 100%. Values represent the mean±SEM (n=6 from three different donors). Paired t-test: (a) p≤0.05 versus control growing media, (b) p≤0.05 versus EMD growing media.
mRNA levels of different osteogenic markers
The effect of synthetic peptides or EMD treatment (50 μg/mL) on mRNA levels of different osteogenic markers was analyzed by real-time RT-PCR after 1, 3, and 14 days in hUCMSC cell cultures. Five different osteogenic markers were analyzed (Figs. 4–6), including collagen-I, alkaline phosphatase, RUNX2, decorin, and osteocalcin. In addition to the osteogenic markers, WNT7B was also analyzed. After short cell treatment, EMD induced higher collagen-I mRNA expression levels than the synthetic peptides in cells cultured in growing media; when cells were cultured in differentiation media, this difference was only seen after 3 days of treatment. No significant differences in collagen-I mRNA levels were found at day 14, nonetheless, although values did not reach statistical significance, EMD treatment in growing media induced higher levels of COLL-I mRNA expression compared with control and to peptide treatment.
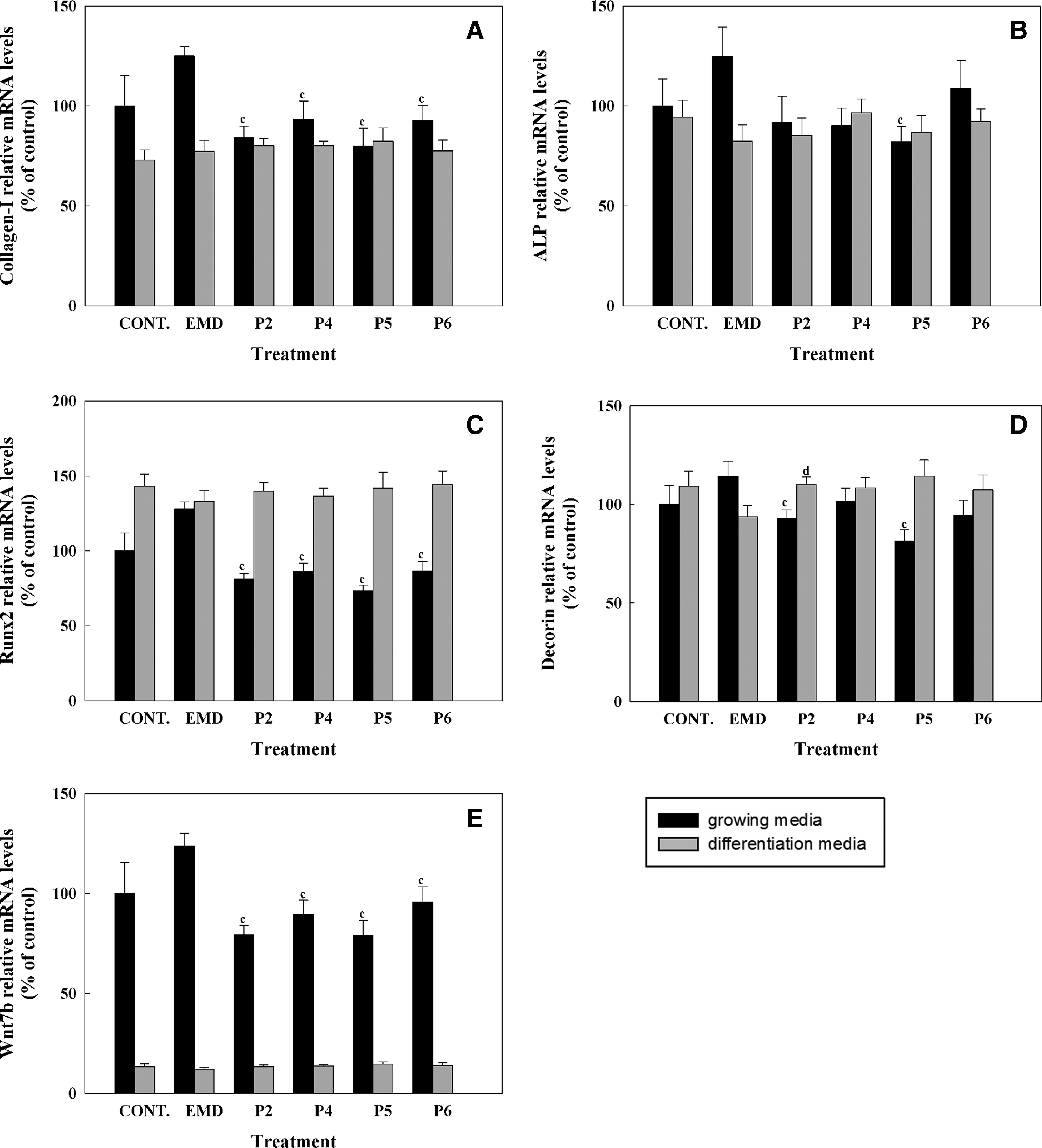
Gene expression levels of collagen-I
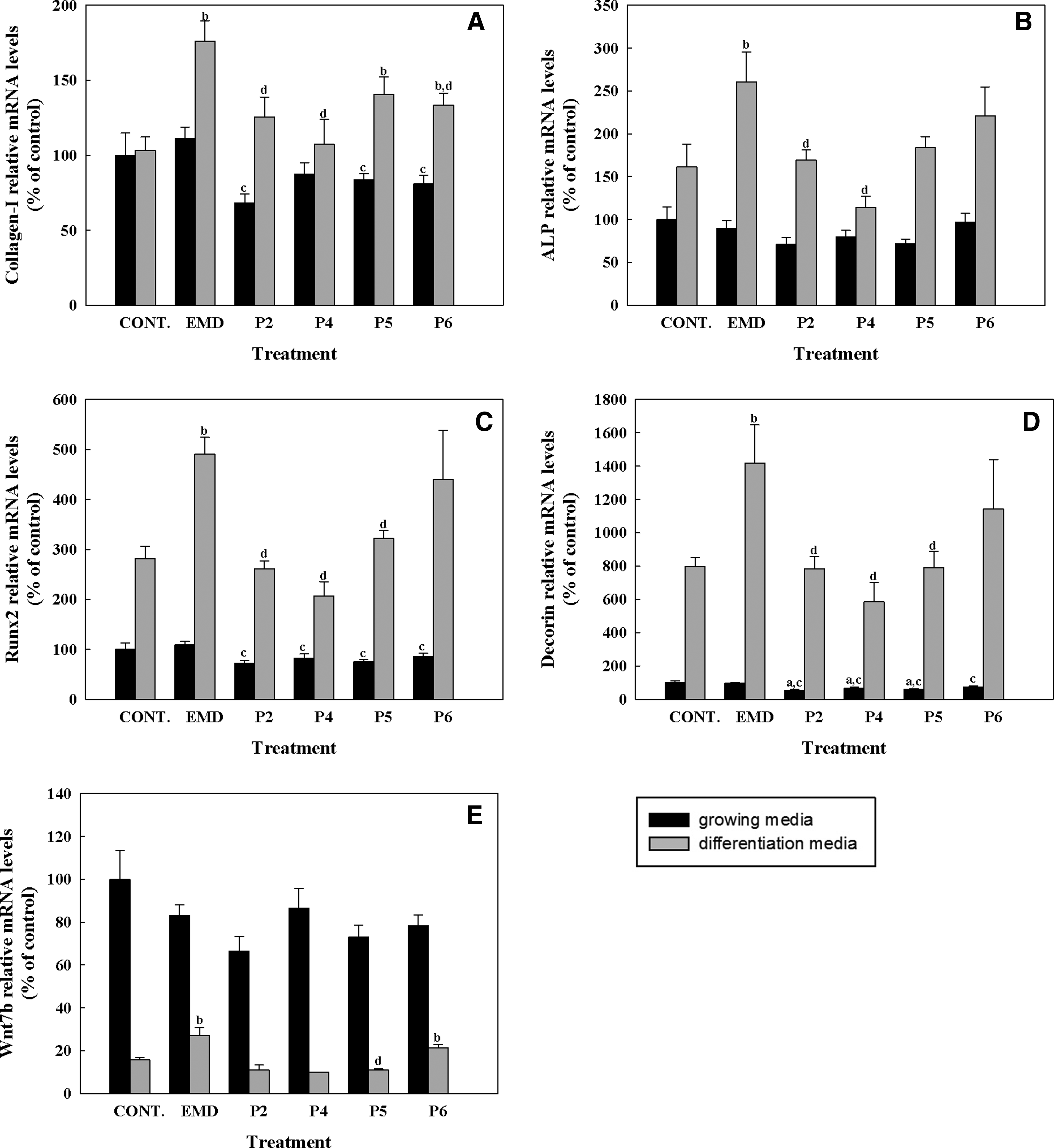
Gene expression levels of collagen-I
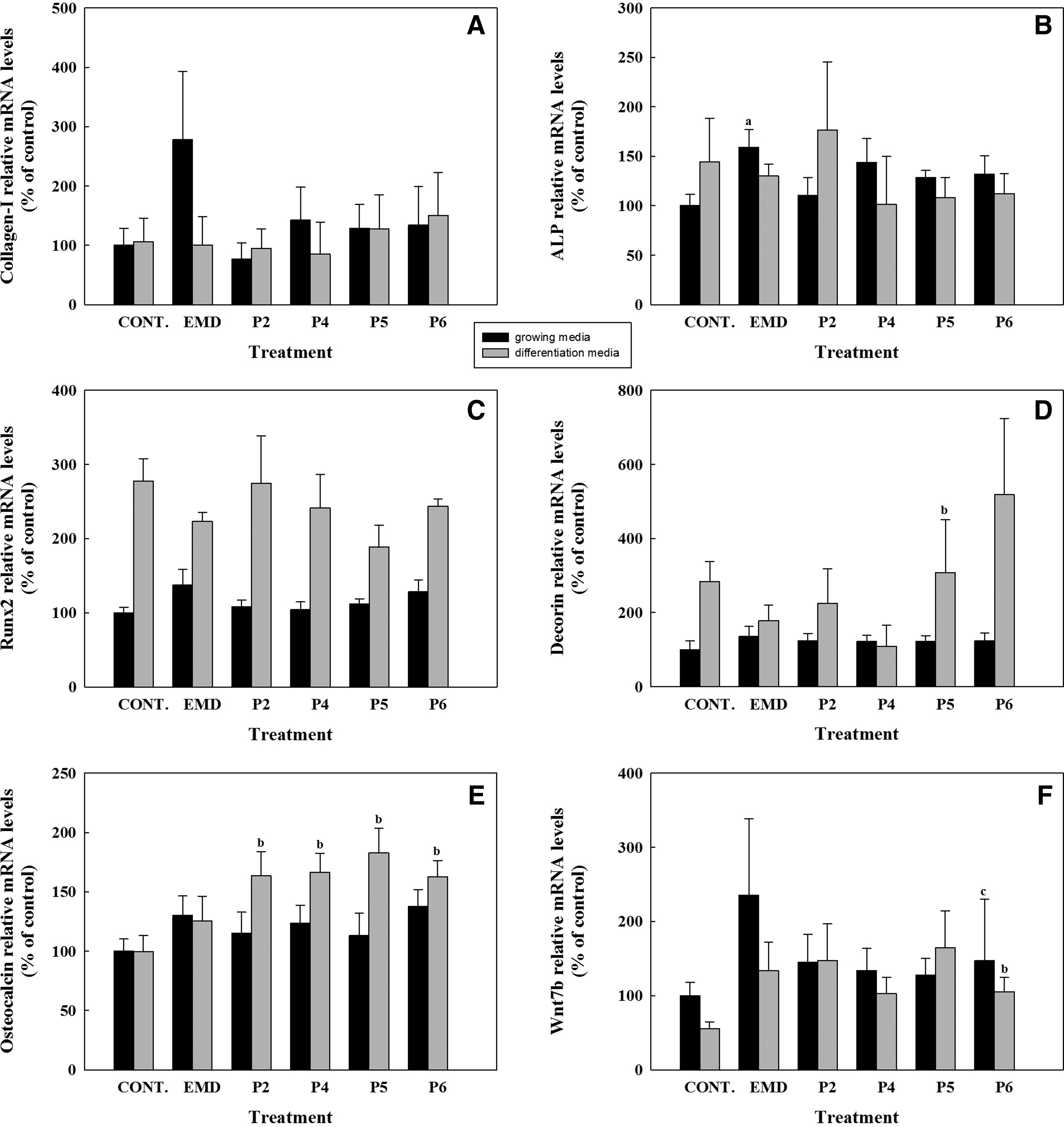
Gene expression levels of collagen-I
ALP mRNA levels were increased in cells cultured in differentiation media for 3 days for all the treatments compared with growing media, and significantly higher levels were found in cells treated with EMD compared with the control and to P2 and P4. At day 14, EMD treatment induced a higher expression of ALP mRNA levels compared with the control in growing media. No differences were found on ALP mRNA levels in cells treated with the different peptides after 14 days of treatment.
RUNX2 mRNA levels were higher in cells cultured with differentiation media for the different days analyzed. At day 3, EMD treatment induced significantly higher levels of RUNX2 compared with the control and to peptide treatment. Moreover, in cells cultured in growing media EMD treatment for 1 and 3 days induced higher RUNX2 levels compared with peptide treatment.
After 3 days of treatment, decorin expression showed the same profile as RUNX2, showing higher levels in cells cultured in differentiation media and a higher induction in cells treated with EMD compared with control and with peptide treatment. After 14 days of treatment, P5 treatment induced a higher expression of decorin mRNA levels compared with the control in differentiation media. No differences in mRNA levels were found with treatment with any other peptides or with EMD treatment at day 14.
WNT7B mRNA levels were found to be higher in cells cultured in growing media than in cells cultured in differentiation media, mainly after 1 and 3 days of culture. Moreover, after 1 day of treatment a significant reduction of WNT7B mRNA levels in cells treated with the peptides compared to cells treated with EMD was found. At day 3, cells cultured in differentiation media and treated with EMD showed higher levels of WNT7B, reaching significance when compared with the control and to P5. P6 showed significantly higher mRNA levels compared with the control in differentiation media at days 3 and 14 and lower WNT7B mRNA levels compared with EMD in growing media at day 14.
Osteocalcin mRNA was not detected after 1 and 3 days of culture. After 14 days of treatment, all peptides induced a higher expression of osteocalcin mRNA levels compared with the control in differentiation media. No differences were found on osteocalcin mRNA levels in cells treated with EMD compared to the control in any of the media.
Release of BMP-2 into the cell culture media
BMP-2 protein levels in culture media after 14 days of treatment were analyzed by ELISA (Fig. 7). The levels of BMP-2 in cells treated with the synthetic peptides were lower than in cells treated with EMD or control cells. However, statistical significance was only found in cells cultured in growing media for P2 and P5. No differences were found among culture media used with and without osteogenic supplements.

Bone morphogenetic protein 2 (BMP-2) protein levels in culture media of hUCMSCs treated with the different peptides (50 μg/mL) or EMD (50 μg/mL) for 14 days. Data expressed as pg BMP-2/mg protein. Values represent the mean±SEM (n=6 from three different donors). Paired t-test: (a) p≤0.05 versus control growing media, (c) p≤0.05 versus EMD growing media.
Discussion
In this study, synthetic peptides sharing a common polyproline sequence and the commercially available EMD were investigated for osteoinductive potential of hUCMSCs. hUCMSCs provides a multipotent cell source capable to differentiate into osteoblasts when cultured with osteogenic supplements. 5 In a previous study, we have shown that the peptide 2 variant enhanced the differentiation toward the osteogenic lineage of hUCMSCs. However, the individual effect of the synthetic peptide was not investigated. Thus, to provide a more comprehensive evaluation, the present study aimed to further investigate the ability of synthetic proline-rich peptides and of EMD to promote the osteogenic differentiation of hUCMSCs individually or in combination with osteogenic supplements.
Bone tissue formation, occurs throughout life, and is involved in bone remodeling in adults. 12 Mesenchymal stem cells are the main source of osteoprogenitor cells, being involved in normal skeletal homeostasis 13 and in reparative mechanisms of bone. 14 Since MSCs are directly involved in the bone regenerative process and are capable of propagating along the osteogenic lineage, primary cultures of hUCMSCs were chosen for our studies. However, variables among different donors must be taken into account when using primary cells; to overcome this problem and to improve validity of our data, we have used three different donors.
We have previously demonstrated the safety of peptide 2 after short- and long-time in vitro cell treatment. 2 Here, we confirmed that the new variants of our consensus peptide evaluated in the present study did not impair cell viability after a short period of treatment. Thus, we can conclude that the synthetic consensus peptides at a dose of 50 μg/mL are safe regardless of the composition of the non-proline aminoacids.
In consistence with our previous study, 2 treatment with the synthetic peptides reduced cell proliferation compared with untreated cells, while no differences on cell proliferation on cells treated with EMD compared to control were found. The lack of effect of EMD on cell proliferation might be due to an enhanced cell differentiation, in fact, previous studies have shown that EMD might exert a different effect depending on the maturation state of the cells.15,16The effect of EMD on MSCs proliferation has been evaluated in few studies with controversial results; different studies have proved that EMD enhanced proliferation,17–19 while others have failed to find EMD-induced differences in terms of cell proliferation. 20 These inconsistencies in results might be due to differences in cell source and differences in experimental designs between different studies.
To investigate the osteoinductivity of the synthetic peptides and of EMD, we analyzed the expression of different osteoblast markers of hUCMSCs after 1, 3, and 14 days of treatment in the presence and absence of osteogenic supplements in the culture media. Five different marker genes were analyzed, including collagen-I, RUNX2, ALP, decorin, and osteocalcin. Higher osteopromotive effect was found for EMD after a short-term treatment of 3 days in cells cultured with osteogenic supplements, since we found higher expression levels of all the early osteoblast markers analyzed compared to both, control and peptide-treated cells.
However, after a long-term treatment of 14 days, this osteopromotive effect of EMD was not found any longer in the presence of osteogenic supplements, while some osteopromotive effect was observed in cells cultured in growing media. Type I collagen and alkaline phosphatase are considered early markers of differentiation,21,22 and at day 14 we found that EMD-treated cells cultured without osteogenic supplements had higher expression levels of COLL-1 and ALP than control cells, while no differences were found in cells treated with the synthetic peptides. These results are in agreement with reports from other authors.18,23–25 Similar response to treatments was found for RUNX2, an osteoblast specific transcription factor, 26 and for decorin, a small leucine rich proteoglycan highly expressed in extracellular bone matrix. 27 In all experimental groups, RUNX2 and decorin mRNA levels were higher when cells were cultured with the osteogenic supplements and no differences were found among treatments, except for P5, that induced decorin expression in cells cultured in differentiation media.
Among the osteoblast markers, osteocalcin—a major noncollagenous protein of bone highly expressed in mature osteoblasts—is currently considered the most specific and the latest of expressed markers, in fact, is the only gene that is only expressed by fully differentiated osteoblasts and regulates their function.28–30 In fact, at days 1 and 3 osteocalcin mRNA levels were not detected by real-time RT-PCR in hUCMSCs, in the presence or absence of osteogenic supplements in the culture media. We found that treatment with any of the synthetic peptides increased osteocalcin mRNA levels but only when cells were stimulated with the osteogenic supplements after 14 days of treatment. No significant differences were found on osteocalcin expression levels in cells treated with EMD, in accordance to other reports.17,18 The finding that the synthetic peptides induce an increase in osteocalcin mRNA levels indicates that cells are at a more mature stage of the differentiation process. Moreover, the relevance of this gene has been demonstrated in a recent in vivo study, in which osteocalcin was found to be the best predictive marker for osseointegration of titanium implants (Monjo et al. submitted).
Osteocalcin results indicate that although the synthetic peptides are not osteoinductive in hUCMSCs, they are osteopromotive in combination with osteogenic supplements after long-term treatment. These results, together with our previous observations 2 suggest a different mechanism of action for the synthetic peptides and EMD. Here, we have observed that EMD is osteopromotive after short-term treatment in cells cultured with osteogenic supplements, while after long-term treatment some osteopromotive effect is seen in cells cultured in growing media. It would be possible that longer EMD treatment than 14 days could induce the osteogenic differentiation of hUCMSCs in the absence of osteogenic supplements. On the other hand, synthetic peptides have shown an osteopromotive effect in combination with osteogenic supplements after long-term treatment.
Since it has recently been shown that platelet-released supernatant stimulates mesenchymal stem cell differentiation toward an osteoblastic phenotype, and that this effect could be mediated by BMP-2 through the Wnt-signaling pathway, 31 we analyzed BMP-2 secretion of hUCMSCs and WNT7B mRNA expression levels to test whether the synthetic peptides or EMD would exert their action through a similar mechanism. The obtained results are not conclusive, though we observe higher WNT7B mRNA expression levels in cells treated with EMD and cultured in growing media after 1 or 14 days of treatment and in cells cultured in osteogenic supplemented media after 3 days of treatment. Moreover, these data correlate with higher levels of BMP-2 secretion after 14 days of treatment. The finding that WNT7B mRNA expression levels were higher in cells cultured in the absence of the osteogenic supplements is in agreement with the reported observation that the canonical Wnt signaling maintains an undifferentiated proliferative state of MSCs. 32
Consistent with the similar structural properties of the different synthetic peptides evaluated in this study, no differences have been found in the activity among them. In agreement with previously reported P2 peptide property, 2 all of the synthetic peptides exposed their PPLPP stretch in a way suitable for interactions. The short sequence of PPXPP in the C-terminal region of peptides has been shown to participate in the transactivation activity of transcription factors and/or co-activators.33,34 We have previously hypothesized that the mode of action of the synthetic peptides might involve interaction with a receptor capable of influencing intracellular signaling cascades, and that the accessibility and structural rigidity of this short consensus sequence (PPXPP) may be of importance in the signaling activity of our synthetic peptides. Actually, we are currently evaluating the significance of the PPLPP motif in the osteopromotive effect of the designed synthetic peptides and we expect to clarify its biological importance in future works.
Our findings that synthetic polyproline-rich peptides promote osteogenic differentiation of hUCMSCs after long-term treatment are relevant in the development of new treatments for tissue regeneration that could be more cost effective. The use of natural proteins like EMD in tissue engineering and regenerative medicine has some associated disadvantages including immunogenicity, relatively high cost, large molecular weight (100–200 amino acids), instability in vivo, and difficulty in sterilization. In contrast, synthetic peptides could be advantageous due to their ease of synthesis and handling, and their low immunogenic activity.
Conclusion
Osteogenic differentiation of hUCMSCs is promoted by EMD after short-term treatment. In contrast, synthetic polyproline-rich peptides promote osteogenic differentiation of hUCMSCs after long-term treatment regardless of the composition of the non-proline aminoacids spacing the proline motifs.
Footnotes
Acknowledgment
This work was supported by the Norwegian Research Council (Grant Number 171058), the Conselleria de Comerç Industria i Energia de les Illes Balears (BA-2009-CALT-0001-PY), the Conselleria d'Innovació, Interior i Justicia de les Illes Balears (AAEE0044/09), the Spanish Ministry of Health's Fondo de Investigación Sanitaria (FIS-PI07/1021), the Ministerio de Ciencia e Innovación del Gobierno de España (Torres Quevedo contract to M.R. and J.M.R., and Ramón y Cajal contract to M.M.) and Eureka-Eurostars Project Application E!5069 NewBone, and Interempresas Internacional Program (CIIP20101024) from the Centre for the Development of Industrial Technology (CDTI). It was also supported by the Grant Agency of the Czech Republic (Grant number P302/10/0427), and it was also a part of the research projects No. Z40550506 and Instutional Research Concept No. AV0Z505200701.
Disclosure Statement
No competing financial interests exist.