
Abstract
Mouse 3T3 feeder layer has been utilized for epidermal and corneal epithelial cell culture to promote tissue-like cell stratification. However, the molecular mechanism underlying epithelial–feeder layer interactions remains poorly understood. Here, the feeder layer activity of six different mouse cell lines was examined in terms of the colony-forming efficiency (CFE) of primary limbal epithelial cells, including corneal epithelial stem/progenitor cells. When epithelial cells and feeder layers were separated by culture inserts, the CFE was significantly lower than that of epithelial cells, which were cultured with feeder cells on the same dish surfaces, implying that direct contacts between these cells and/or pericellular extracellular matrix (ECM) deposition by feeder layers have an important role in feeder layer activity. With TaqMan polymerase chain reaction assay, the gene expression of 29 ECM molecules and 32 cadherin family genes was profiled in two highest and two lowest cell lines in the CFE for limbal and oral mucosal epithelial cells. A significant difference in the expression correlated with the CFE was observed in six ECM molecules and four kinds of cadherin family genes. In these results, type VI collagen was confirmed to be able to promote the colony formation of epithelial cells in vitro effectively.
Introduction
Clinically, the use of expanded epithelial stem cells using the 3T3 feeder method has been applied in the treatment of patients suffering from a variety of diseases, including burns 4 , giant congenital nevi 5 , or severe ulcers. 6 More recently, the use of both expanded limbal epithelial and oral mucosal epithelial cells has been effectively used for treating patients with total limbal stem cell deficiencies for the long-term restoration of corneal transparency and visual acuity.7–10 These results, therefore, also imply the functional importance of interactions with 3T3 feeder layers in the maintenance of epithelial stem and progenitor cells. Previous reports have revealed that specific soluble factors secreted by fibroblast feeder layers have a distinct effect on epithelial cell dynamics.11–15 However, even with the determination of soluble agents that can enhance cell proliferation, the underlying molecular mechanism regarding the effects of fibroblast feeder layers remains poorly understood.
To understand the mechanism between feeder layers and epithelial cells, the colony-forming efficiency (CFE) of six different fibroblast feeder cell lines were assayed for determining cell lines effectively promoting colony formation of epithelial cells. Moreover, the expression of extracellular matrix (ECM) molecules and cadherin family molecules in the effective fibroblast cell lines, as candidate molecules to promote the colony formation of epithelial cells, was identified by quantitative gene expression analysis.
Materials and Methods
Preparation for feeder layers
Mouse fibroblast-like cell lines, A9 (adult skin), B82 (adult skin), L cl 1D (adult skin), 3T3-Swiss albino (3T3-Swiss, embryo), C3H/10T1/2-clone 8 (10T1/2, embryo), and NIH-3T3 (embryo), were examined. NIH-3T3 was obtained from Dainippon Pharmacy (Osaka, Japan), and other cell lines were obtained from the Japan Health Science Research Resource Bank (Osaka, Japan). Before feeder layer preparation, these cell lines were seeded at a cell density of 1/8 confluence onto 100-mm cell culture dish in the Dulbecco's modified Eagle's medium (DMEM) supplemented 10% fetal bovine serum (FBS; Moregate BioTech, Queensland, Australia). After 2-day culture, subconfluent cells were treated with 10 μg/mL of mitomycin C (MMC; Wako Pure Chemicals, Osaka, Japan) in a serum-free DMEM for 2 h at 37°C and washed with Dulbecco's phosphate-buffered saline (PBS). Then, MMC-treated cells were harvested by 0.25% trypsin and 0.1% EDTA, and plated onto six-well plates at cell numbers of 5×104, 1×105, and 2×105 cells/well or onto six-well cell culture inserts at cell numbers of 5×104, 1×105, and 2×105 cells/insert.
Preparations for rabbit epithelial cells
All experimental protocols were approved by the animal welfare committee of the Tokyo Women's Medical University. Rabbit limbus, including about 1-mm width of cornea and conjunctiva, was isolated by scissors and treated with 200 U of dispase (Godo Shusei, Chiba, Japan) for 1 h at 37°C. Oral mucosa was excised from the buccal cavity of rabbit and treated with 1000 U of dispase at 4°C over night. Then, limbal and mucosal epithelium was separated from stroma, and the epithelium was treated with 0.25% trypsin and 0.1% EDTA for 20 min at 37°C. The disaggregated cell suspension was filtrated to 40-μm-pore-size cell strainer (BD Biosciences, Franklin Lake, NJ).
Colony-forming assay
Isolated cells were suspended in a keratinocyte culture medium composed of a basal mixture of three parts of DMEM and one part of nutrient mixture F-12 Ham (Sigma-Aldrich, St. Louis, MO) supplemented with 5% FBS, 0.5% insulin-transferrin-selenium (Gibco-Invitrogen, Carlsbud, CA), 10 ng/mL epidermal growth factor (Invitrogen, Carlsbud, CA), 0.4 μg/mL hydrocortisone (Wako Pure Chemicals), 5 μg/mL transferrin (Gibco-Invitrogen), 2 nM triiodothyronine (Wako Pure Chemicals), and 1 nM cholera toxin (Calbiochem, Darmstadt, Germany). Suspended limbal epithelial cells were seeded at a cell density of 10 or 20 cells/cm2 onto 6-well plates with feeder layer cells (Fig. 1A). For establishing an indirect contact culture condition, feeder layer cells were put in a culture insert of which the bottom was a membrane having pores (0.4 μm in diameter) (Fig. 1B). Oral mucosal epithelial cells were also seeded at a cell density of 100 or 300 cells/cm2 with direct contact with feeder layer cells. After 11- to 14-day culture, cells were fixed with 3.7% formalin and stained with Rhodamine B. CFE was calculated by dividing the observed number of colony with the initial cell number and expressed as means with standard deviations.

Schematic illustration of colony-forming assay (CFA) under two different culture conditions.
Real-time polymerase chain reaction analysis
mRNA expression was profiled with real-time polymerase chain reaction (PCR) analysis. Subconfluent cultured cells on a 100-mm culture dish were treated with ISOGEN (Nippon gene, Tokyo, Japan). Total RNA extraction was performed according to the manufacture's protocol accompanied with this reagent. Reverse transcription (RT) reactions were performed using the following conditions: 1 μg of total RNA, 500 μM of each dNTP, 25 μg/mL of oligo(dT)12–18, 200 U of SuperScriptTM II Reverse Transcriptase (Invitrogen), and a corresponding buffer; milli-Q water was added into the tubes for adjusting the total volume to be 20 μL; thermocycler conditions were 65°C for 5 min; 42°C for 50 min; and 70°C for 15 min. To remove RNA complementary to the cDNA, samples received two units of RNase H (Invitrogen) and were incubated at 37°C for 20 min. To analyze mRNA expression, 6-FAM dye-labeled TaqMan prove (Applied Biosystems, Foste City, CA) were used. Real-time RT-PCR was performed according to the manufacture's protocol accompanied with a 7500 Real-Time PCR System (Applied Biosystems).
Preparation of cell culture surface coated with type VI collagen
Type VI collagen (BD Bioscience) was diluted with PBS. Six- and 12-well cell culture plates were coated with 1 and 0.5 mL of type VI collagen (0.5–10 μg/mL) at 4°C over night, respectively. After the incubations, remaining solutions on six-well plates were aspirated, and the coated plates were rinsed with sterile pure water. MMC-treated NIH-3T3 were seeded on the coated wells at a cell number of 2×105 cells/well for the colony-forming assay (CFA) of rabbit oral mucosal epithelial cells as described above. Type VI collagen-coated 12-well plates were treated with 1% bovine serum albumin (BSA) in sterile pure water for 4 h and washed with sterile pure water. The 12-well plates were used for cell adhesion assay.
Cell adhesion assay
Cell adhesion assays were carried out with rabbit oral mucosal epithelial cell suspensions in serum free basal medium of KCM for 60 min in a humidified incubator at 37°C. The epithelial cells were seeded on type VI collagen-coated 12-well plates at a density of 4×104 cells/cm2. After the incubation, the epithelial cells were washed with 10% FBS-DMEM and with PBS, and harvested with 0.25% trypsin and 0.1% EDTA. The total numbers of adhesive cells were determined with a hemocytometer. Percent of adhesive cell was calculated by dividing the total number of adhesive cell by the initial seeded number of cell. The numbers of cells on BSA-coated wells were assumed as the control, and all data were expressed as means with standard deviations.
Statistical analysis
Statistical analysis was computed with the aid of StatView5.0 software. The Kruskal-Wallis test was performed for analyses of CFA of limbal epithelial cells (n=6) and oral mucosal epithelial cells (n=4), real-time RT-PCR of feeder cells (n=3), and cell adhesion assay (n=3). Scheffe's test was performed for CFA and real-time RT-PCR. Statistical analyses of CFA (n=4) and cell adhesion assay (n=3) using type VI collagen coated plates were carried out according to Dunnett's test.
Result
Feeder layer activity
The feeder layer activities of six murine cell lines were examined with primary limbal epithelial cells. Proliferations of the six murine cell lines were completely inhibited by treatment with MMC (data not shown). All examined cells promoted the colony formation of limbal epithelial cells, whereas these epithelial cells formed no colonies without feeder layer cells at a seeding density shown in the Materials and Methods section (data not shown). Here, the CFEs of these six murine cell lines were quantified under two different coculture conditions: direct contact (Fig. 1A) and no contact conditions (Fig. 1B). Limbal epithelial cells and feeder layer cells were cocultured on a culture dish (direct contact condition), and the two types of cells were separated by culture inserts (no contact condition). All feeder layer calls were able to form the colony of limbal epithelial cells even in the indirect condition (the inlet of Fig. 2), indicating that soluble factors secreted these feeder layer cells had a colony-forming activity. However, significantly higher CFEs were obtained under the direct contact condition than no contact condition (Fig. 2).
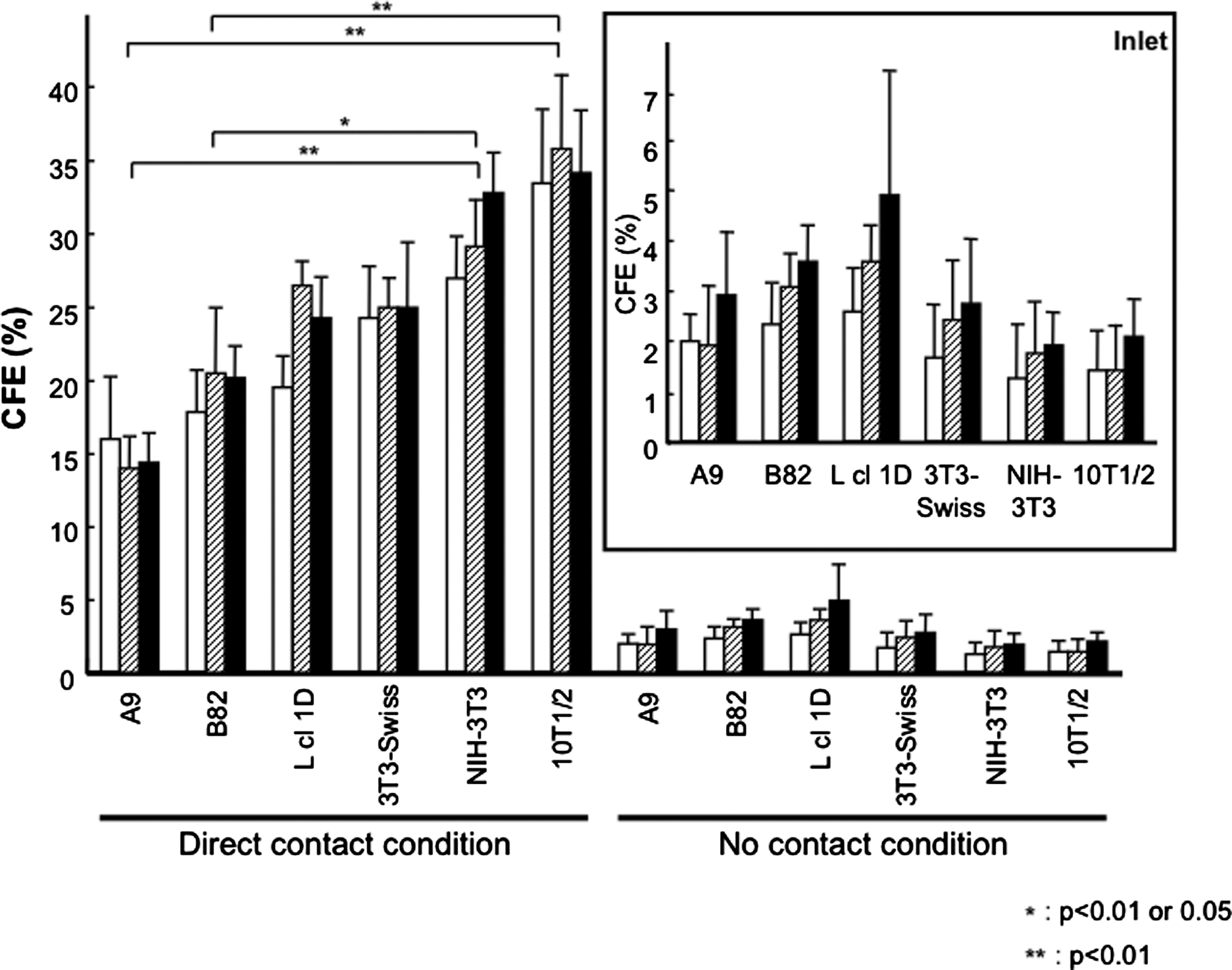
Colony-forming efficiency (CFE) of six murine fibroblast cell lines for limbal epithelial cells. Rabbit limbal epithelial cells were seeded on six-well plates at a cell density of 10 cells/cm2 under direct contact condition, and a cell density of 20 cells/cm2 under indirect contact condition was described in Figure 1. Mitomycin C (MMC)-treated six fibroblast cell lines, including A9, B82, L cl 1D, 3T3-Swiss, NIH-3T3, or C3H/10T1/2 (10T1/2), were used as feeder layer cells. CFE was calculated by dividing the number of colony with the number of initial seeding cell, multiplied by 100, and expressed as means with standard deviations. The open bars show CFE (%) of the feeder cells at a seeding density of 5×104 cells/well or cells/insert; the hatched bars, 1×105 cells/well or cells/insert; the close bars, 2×105 cells/well or cells/insert. Inlet is an enlarged graph showing the results of CFE under the indirect contact conditions.
Under the direct contact condition, 10T1/2 and NIH-3T3 showed the two highest CFE, and A9 and B82 showed the two lowest CFE. There were significant differences among the CFE of A9, B82, NIH-3T3, and 10T1/2 at all feeder cell densities under the direct contact condition (p<0.01, Kruskal-Wallis test); nevertheless, there were no significant differences under indirect contact condition. Scheffe's test revealed that the CFEs of 10T1/2 were significantly higher than those of A9 and B82 at all feeder cell densities (p<0.01, Fig. 2), and the CFEs of NIH-3T3 were significantly higher than that of A9 at all feeder cell densities (p<0.01, Fig. 2). The CFEs of NIH-3T3 were significantly higher than those of B82 at cell numbers of 5×104 and 2×105 cells/well (p<0.01, Fig. 2). At a feeder cell number of 1×105 cells/well, there was a significant difference between B82 and NIH-3T3 (p<0.05, Fig. 2).
For confirming the feeder layer activities of murine fibroblasts having the two lowest (A9 and B82) and the two highest (10T1/2 and NIH-3T3) CFEs for other stratified squamous epithelial cells, CFEs of these cell lines were examined by CFA with primary rabbit oral mucosal epithelial cells. Interestingly, no colony formation of oral mucosal epithelial cells was induced by A9 and B82 (Fig. 3A). NIH-3T3 and 10T1/2 were able to induce the colony formation of oral mucosal epithelial cells, but CFEs were significantly lower than those of limbal epithelial cells (Fig. 3). Furthermore, statistical analysis indicated that there was a significant difference among the four feeder layer activities for oral mucosal epithelial cells (p<0.01), and the feeder layer activities of not only A9 and B82 but also NIH-3T3 were significantly lower than that of 10T1/2 under the direct contact condition (p<0.01, Fig. 3). The results indicated that there were possible 10T1/2-specific factors that induced the colony formation of oral mucosal epithelial cells under the direct contact condition.

CFE of A9, B82, NIH-3T3, and C3H/10T1/2 (10T1/2) for oral mucosal epithelial cells. Rabbit oral mucosal epithelial cells were seeded on six-well plates at a cell density of 100 or 300 cells/cm2 under a direct contact condition. As feeder layer cells, MMC-treated A9, B82, NIH-3T3, or 10T1/2 were seeded at a density of 2×105 cells/well. CFE was calculated by dividing the number of colony with the number of initial seeding cell, multiplied by 100.
Profiling of mRNA expression of 29 ECM molecules
Gene expression of 29 ECM molecules in A9, B82, NIH-3T3, and 10T1/2 was quantified by real-time RT-PCR analysis with specific TaqMan probes. Twenty-nine of ECM molecules consisted of 19 procollagen genes, 6 proteoglycan core protein genes, and 4 tenascin genes (Table 1). The mRNA expression of these molecules was normalized with Gapdh mRNA expression, and statistical analysis was carried out according to Kruskal-Wallis test for selecting candidate molecules to promote the growth and colony formation of epithelial cells. As the results, gene expression of 15 ECM molecules was significantly different among the feeder cells (p<0.05). Therefore, Scheffe's test was performed to find significant differences among the expression in each feeder cells. As the results, there was significantly different expression among six molecule genes; type II collagen alpha1 chain (Col2a1), type VI collagen alpha1 chain (Col6a1), type XII collagen alpha1 chain (Col12a1), type XVII collagen alpha1 chain (Col17a1), osteoglycan (Ogn), and tenascin C (Tnc) (Fig. 4). There are no significant differences in the comparison of these six gene expressions between A9 and B82 (Fig. 4). The mRNA expression of Col2a1, Col6a1, Col17a1, and Ogn in 10T1/2 was significantly higher than those in A9, B82, and NIH-3T3 (p<0.05 or 0.01, Fig. 4). Additionally, Col12a1 and Tnc expression in 10T1/2 was significantly higher than those in A9 and B82 (p<0.05 or 0.01, Fig. 4), although there were no significant differences between 10T1/2 and NIH-3T3. Interestingly, Tnc expression was detected only in 10T1/2 and NIH-3T3, and the expression levels were almost the same.

The results of real-time polymerase chain reaction (PCR) analysis of the expression of extracellular matrix molecules. Expression of mRNA of type II collagen alpha1 chain (Col2a1), type VI collagen alpha1 chain (Col6a1), type XII collagen alpha1 chain (Col12a1), type XVII collagen alpha1 chain (Col17a1), osteoglycin (Ogn), and tenascin C (Tnc) in A9, B82, NIH-3T3, and C3H/10T1/2 (10T1/2) was presented as means of Gapdh(%) with standard deviation. ND represents nondetected expression.
Profiling of mRNA expression of 32 cadherin family genes
mRNA expression of the 32 cadherin family genes in the four murine fibroblasts, consisted of A9, B82, NIH-3T3, and 10T1/2, was quantified by real-time RT-PCR analysis with specific TaqMan probes and normalized with Gapdh mRNA expression. Thirty-two cadherin family genes consisted of 19 cadherin genes, 8 desmosome protein genes, and 5 other cadherin family genes (Table 2). In the results of the Kruskal-Wallis test, four cadherins were graded as candidates for the factors to promote the colony formation of epithelial cells. Expression of cadherin 2 (Cdh2), cadherin 11 (Cdh11), cadherin 13 (Cdh13), and cadherin-like 26 (Cdh26) in four feeder cells showed significant difference (p<0.05, Fig. 5). Additionally, Scheffe's test found that mRNA expression of Cdh2 and Cdh26 in 10T1/2 and NIH-3T3 was significantly higher than the expression in A9 and B82 (p<0.01, Fig. 5). Furthermore, the expression of Cdh11 and Cdh13 in 10T1/2 was higher than that in A9, B82, and NIH-3T3 (p<0.01, Fig. 5). However, there were no significant differences among the expression of these cadherins in NIH-3T3, A9, and B82.

The results of real-time PCR analysis of the expression of cadherin family genes. Expression of mRNA of cadherin2 (Cdh2), cadherin11 (Cdh11), cadherin13 (Cdh13), and cadherin-like26 (Cdh26) in A9, B82, NIH-3T3, and C3H/10T1/2 (10T1/2) was presented as means of Gapdh (%) with standard deviation. ND represents nondetected expression.
CFA and cell adhesion assay of type VI collagen-coated culture surfaces
The results of real-time RT-PCR analysis indicated possible candidate molecules promoting the colony formation of epithelial cells, since this gene expression was significantly higher in feeder cells having a high CFE. In these candidate molecules, the expression of Col6a1 was focused as a possible candidate molecule, and type VI collagen-coated culture plates were prepared for CFA. Rabbit oral mucosal epithelial cells were seeded on the coated 6-well plates and cocultured with MMC-treated NIH-3T3, because the CFE of oral mucosal epithelial cells and Col6a1 expression of 10T1/2 were significantly higher than those of other feeder cells. Moreover, the gene expression of Col6a1 in NIH-3T3 and 10T1/2 treated with MMC did not affect the culture condition with or without rabbit oral mucosal epithelial cells (data not shown). In the results, type VI collagen showed the dose-dependent induction of CFE of rabbit oral mucosal epithelial cells (Fig. 6A). Statistical analysis showed that CFEs at 0.5 and 1 μg/cm2 type VI collagen were significantly higher than the control surface (0 μg/cm2) (p<0.05 and 0.01, respectively, Dunnett's test, Fig. 6A). Additionally, the cell adhesion assay of rabbit oral mucosal epithelial cells was performed for confirming the type VI collagen-dependent property to primary epithelial cell adhesion. In a way similar to CFA, oral mucosal epithelial cells showed an adhesive dependency on type VI collagen, and there were significant differences (p<0.01, Fig. 6B). Moreover, when the coating densities of 12-well plates were more than 0.1 μg/cm2, the coated surfaces showed significantly higher epithelial cell adhesions than surfaces coated with only BSA (p<0.01, Fig. 6B).
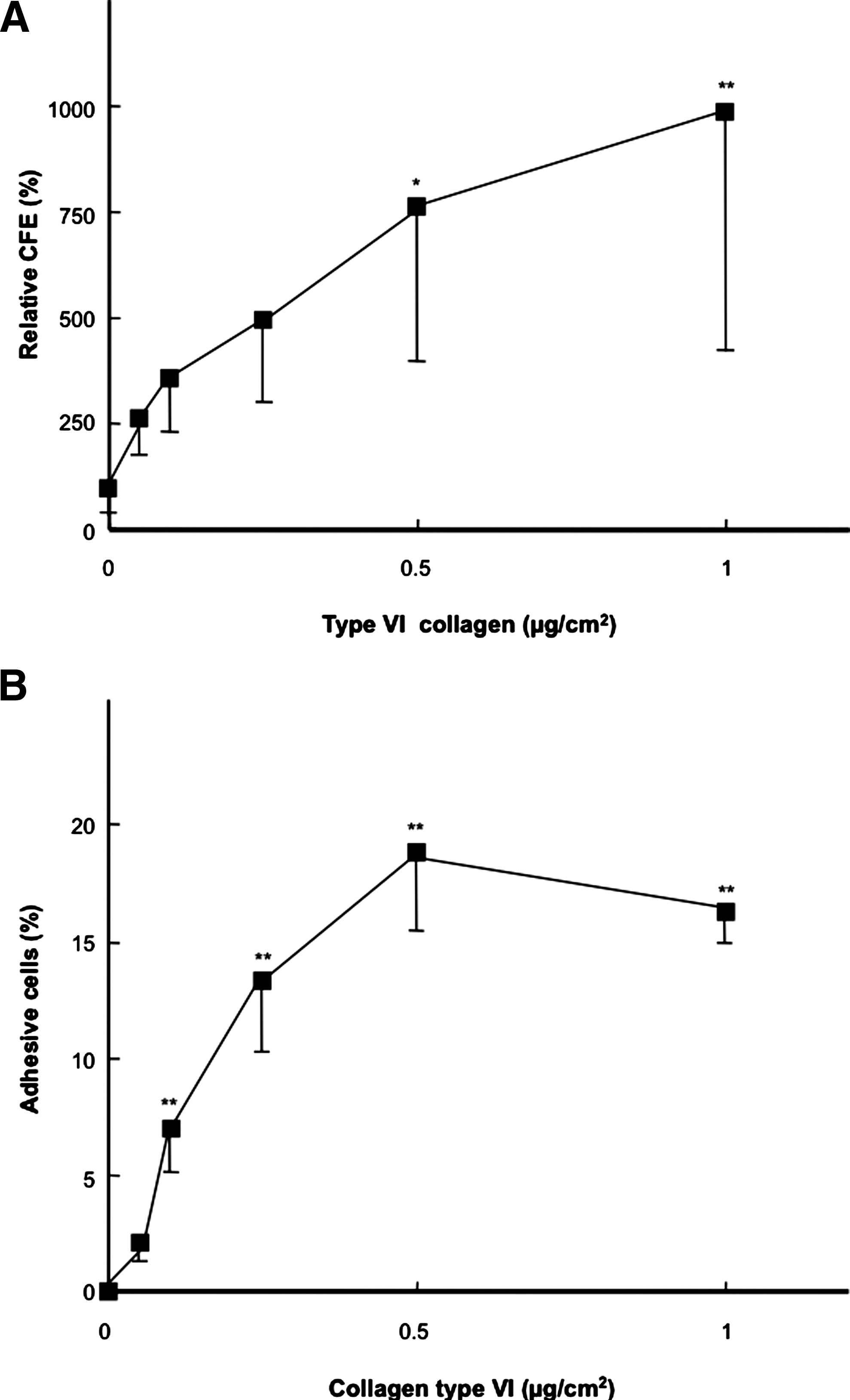
Efficiencies of type VI collagen for the colony formation and cell adhesion of oral mucosal epithelial cells.
Discussion
In the present study, six different mouse fibroblasts were employed as feeder layers. Three of them (A9, B82, and L cl 1D) are subclones of L-cells derived from adult mouse skin, and the rest of them (3T3-Swiss, NIH-3T3, and 10T1/2) were derived from mouse whole embryo. As a whole, embryo-derived fibroblasts showed a relatively higher CFE than skin-derived cells. However, more studies that are extensive are required to conclude that the CFE is dependent on cell origin. Under the indirect condition, all the examined fibroblasts showed a similar, but weak CFE, whereas limbal epithelial cells made no colony without feeder cells. This observation suggested that soluble molecules secreted by feeder layer cells were able to migrate through the micropores (0.4 μm) of culture inserts and affect the CFE of limbal epithelial cells. Similar observation was previously obtained with a conditioned 3T3-cell culture medium. 16 Although the conditioned medium stimulated the colony formation of epithelial cells, the CFE was significantly lower than that under normal coculture condition. The observation in this study suggested that ECM molecules and/or cell–cell interaction between epithelial cells and feeder cells could be candidate factors to promote the colony formation of epithelial cells, and implied the importance of (1) the cell–cell interactions between 10T1/2 and limbal epithelial cells, and NIH-3T3 and the cells, and (2) the extracellular and/or pericellular matrix produced by 10T1/2 and NIH-3T3 in their colony forming activities, since these cells showed significantly a higher CFE under the direct condition.
In previous study, the importance of the cell–cell interaction was indirectly demonstrated, 17 18 and in the present study, the results of CFA also indicated the importance of cell–cell interaction. In the results of real-time RT-PCR, the gene expression of Cdh2 in NIH-3T3 and 10T1/2 was significantly higher than the expression in A9 and B82. Cadherin-2, as known a neural-cadherin (N-cadherin), was originally found in neural tissues 19 , and its cDNA sequence has been determined. 20 In basal epithelial cells of the limbus, there are N-cadherin-positive limbal epithelial cells, and the limbal epithelial cells show a high CFE. 21 Moreover, the CFE of limbal epithelial cells cocultured with NIH-3T3 feeder layer expressing low N-cadherin was significantly lower than that with normal NIH-3T3 feeder cells. 22 Therefore, the expression of Cdh2 in feeder cells may relate to the efficiency of colony formation of epithelial cell cocultured with the feeder cells. In addition to the result of expression of Cdh2, the expression of Cdh11, 13, and 26 was significantly different among feeder cells correlated with CFE. These results may indicate that these cadherin molecules have an important role for the colony formation of epithelial cells. Interestingly, cadherin-11, as known an osteoblast-cadherin (OB-cadherin), is expressed in carcinoma and reported to correlate to epithelial cell motility. 23 Therefore, OB-cadherin may have an important role for epithelial cell functions in a specific appearance such as wound healing, development, and expansion in vitro.
Reduced CFE under the indirect condition also suggested that ECM depositions were one of key factors to promote epithelial cell growth. Then, mRNA expression of 29 ECM molecules in A9, B82, NIH-3T3, and 10T1/2 was quantified by real-time RT-PCR analysis. Based on the results of CFA and RT-PCR, Tnc was selected as a candidate molecule, since Tnc mRNA expression in 10T1/2 was significantly higher than the expression in A9 and B82. Tnc was reported to induce epithelial–mesenchymal interactions in the development of not only kidney, gut, and tooth24–26 but also cornea and oral mucosa during wound healing in humans.27–29 Additionally, the downregulation of Tnc is important to mammary epithelial cell differentiation in vitro. 30 Since these previous studies may indicate an important role in the culture of limbal epithelial cells, A9 and B82 were seeded on Tnc-coated culture dishes as feeder cells for confirming a possible correlation between Tnc and CFE for epithelial cells. In consequence, there was no Tnc effect on promoting the colony formation of limbal epithelial cells cocultured with A9 of B82 (data not shown). In a previous study, cultured corneal epithelial cells also express Tnc in vitro, and Tnc was deposited on the culture surface. 31 Therefore, the results of CFA using a Tnc-coated culture surface in this study may correlate to the previous report.
Cultured oral mucosal epithelial cells are also considered to be a suitable and useful cell source for regenerative medicine for human corneal ocular surface.8,9 Therefore, for confirming the feeder layer activities of A9, B82, NIH-3T3, and 10T1/2 for colony formation of oral mucosal epithelial cells, CFEs of these cell lines were examined by CFA with primary rabbit oral mucosal epithelial cells. The feeder layer activity of 10T1/2 for rabbit oral mucosal epithelial cells was significantly higher than the others, and in the results of real-time RT-PCR, the expression of Col6a1 in 10T1/2 was also significantly higher than that in other feeder cells. Although, the CFE of NIH-3T3 for rabbit oral mucosal epithelial cells was ∼10 times lower than that of 10T1/2, MMC-treated NIH-3T3 seeded on type VI collagen-coated plates showed a significantly higher CFE than that seeded on normal plates. Especially, at a type VI collagen-coating density of 1 μg/cm2, the CFE of NIH-3T3 for rabbit oral mucosal epithelial cells was significantly higher (∼10 times) than that of NIH-3T3 seeded on the normal surface. Interestingly, the percentage of adhesive cells was higher than the percentage of colony formation of NIH-3T3 seeded on the type VI collagen-coated surface. These results indicated that the oral mucosal epithelial cells adhering type VI collagen could partially induce colony formation on the coating surface under the direct contact condition with MMC-treated NIH-3T3. Type VI collagen is expressed in many connective tissues of, particularly, blood vessel, the kidney, skin, the liver, and muscle. 32 Additionally, mRNA expression of type VI collagen is investigated during wound healing, and the results indicate that the expression is regulated in a time-dependent fashion. 33 Moreover, there are many reports that show the cell adhesion of various types of mesenchymal cells to type VI collagen-coated surfaces and a specific binding mode between cells and this molecule. 34 In the present study, type VI collagen indicated the induction of colony formation and adhesion for primary oral mucosal epithelial cells in vitro. Actually, type VI collagen is one of the major components of connective tissue in human oral mucosa 35 and cornea 36 , and considered to have important roles in development and wound healing in cornea.37,38
In conclusion, CFA was demonstrated to compare the CFE between six different mouse cell lines under two conditions: direct contact between limbal epithelial cells and feeder layer cells and indirect one. Consequently, the differences in CFE between different feeder layer cells were demonstrated to be caused by cell–cell contacts between epithelial cells and feeder layer cells and/or deposited pericellular matrix. There upon, mRNA expression of 29 ECM molecules and 32 cadherin family genes in A9, B82, NIH-3T3, and 10T1/2 was quantified by real-time RT-PCR analysis. As the result, the mRNA expression of six ECM molecules (Col2a1, Col6a1, Col12a1, Col17a1, Ogn, and Tnc) and four cadherin (Cdh2, Cdh11, Cdh13, and Cdh26) genes was expressed in feeder cells having a high efficiency to promote the colony formation of epithelial cells. These results may suggest that the expression of these molecules is a candidate to promote colony formation and to maintain epithelial stem/progenitor cells in vitro.
Footnotes
Acknowledgments
The authors thank Dr. N. Ueno, Dr. J. Yang, and Ms. M. Tezima (Institute of Advanced Biomedical Engineering and Science, Tokyo Women's Medical University, Tokyo) for their useful comments. This study was supported by the Formation of Innovation Center for Fusion of Advanced Technologies in the Special Coordination Funds for Promoting Science and Technology Cell Sheet Tissue Engineering Center (CSTEC) and the Global COE program, the Multidisciplinary Education and Research Center for Regenerative Medicine (MERCREM), from the Ministry of Education, Culture, Sports, Science and Technology (MEXT), Japan.
Disclosure Statement
No competing financial interests exist.