
Abstract
Numerous studies on engineering cartilage utilizing chondrocytes from juvenile animal sources have been reported. However, there are many unknown aspects of engineering cartilage using human chondrocytes—especially from middle-aged or elderly adults—which are critical for clinical application of tissue engineering in the field of orthopedic surgery. The primary aim of this study was to engineer neocartilage tissue from 50–60-year-old human chondrocytes in comparison to engineered cartilage made from juvenile swine chondrocytes (JSCs). Articular chondrocytes from middle-aged, nonarthritic humans and juvenile swine were isolated and placed in culture for expansion. The chondrocytes (passage 1) were mixed in fibrin gel at 40–60×106 cells/mL until polymerization. Cells/nodule constructs and devitalized cartilage–cells/hydrogel–devitalized cartilage constructs (three-layered model) were implanted into subcutaneous pockets of nude mice for 12, 18, and 24 weeks. The specimens were evaluated histologically, biochemically, and biomechanically. This allowed for direct comparison of the cartilage engineered from human versus swine cells. Histological analysis demonstrated that samples engineered utilizing chondrocytes from middle-aged adults accumulated basophilic, sulfated glycosaminoglycans (sGAG), and abundant type II collagen around the cells in a manner similar to that seen in samples engineered using JSCs at all time points. Biochemical analysis revealed that samples made with human cells had about 40%–60% of the amount hydroxyproline of native human cartilage, a trend parallel to that observed in the specimens made with swine chondrocytes. The amount of sGAG in the human chondrocyte specimens was about one-and-a-half times the amount in native human cartilage, whereas the amount in the samples made with swine chondrocytes was always less than native cartilage. The biomechanical analysis revealed that the stiffness and tensile of samples made with human cells were in a pattern similar to that seen with swine chondrocytes. This study demonstrates that chondrogenesis using articular chondrocytes from middle-aged adults can be achieved in a predictable and reliable manner similar to that shown in studies using cells from juvenile animals and can form the basis of engineering cartilage with degradable scaffolds in this patient population.
Introduction
The field of cartilage tissue engineering has sought new approaches in which chondrocytes are transferred on biocompatible scaffolds to produce new extracellular matrix (ECM) for the repair of cartilage defects.2–4 Numerous groups have developed methodologies to generate cartilage tissue through cell encapsulation into hydrogels or seeding them onto scaffolds. However, the vast majority of these studies have used cells from juvenile animal sources—cells that are believed to have a strong propensity to form new ECM. Our work with a variety of hydrogels demonstrates successful tissue engineering of cartilage using swine, rabbit, or sheep cells from healthy young animals for the evaluation of chondrogenesis.5,6 Although cartilage can be easily generated with chondrocytes from juvenile animals, these favorable results may not accurately model the clinical conditions of patients requiring treatment for articular cartilage lesions; many of the patients requiring chondral defect repairs are middle aged or older. Several studies have reported that articular chondrocytes from aged human and animal sources have an impaired ability to generate new ECM in vitro. Less is known, however, whether aged human chondrocytes can be reinvigorated to produce neocartilage much like that observed with juvenile animal chondrocytes. More importantly, this should be studied in comparative studies in vivo.
Although studies utilizing juvenile animal cell sources provide fundamental information for the field of cartilage tissue engineering, further studies using human chondrocytes are required for the transition of bench top work into clinical trials. Building on a solid foundation of in vitro and in vivo animal studies, we hypothesized that human cartilage could be engineered with similar morphological, biochemical, and biomechanical properties as engineered cartilage generated by juvenile swine chondrocytes (JSCs). In this study, articular chondrocytes from middle-aged healthy, nonarthritic, human donors were encapsulated in fibrin gel (FG) and compared with articular JSCs. The aims of this study were (i) to determine whether middle-aged human articular chondrocytes encapsulated in hydrogel could form neocartilage, (ii) to evaluate whether the neocartilage could integrate with existing human cartilage, and (iii) to assess whether the tissue-engineered cartilage generated by middle-aged human chondrocytes was comparable to that generated by JSCs, morphologically, biochemically, and biomechanically.
Materials and Methods
Cartilage harvest and cell isolation
All actions were approved by the Institutional Animal Care and Use Committee (IACUC) and IRB of the Massachusetts General Hospital. Human articular cartilage (age range: 50–60 years old, average age: 55 years old) was obtained from National Disease Research Interchange. Native cartilage measuring 1×1×0.2 cm was collected from each sample for control assays. The remainder tissue was finely minced and digested using 0.1% collagenase type II (Worthington Biochemical, Lakewood, NJ) for 16–18 h at 37°C. After digestion, the chondrocytes were rinsed and washed twice with PBS. Cell viability was assessed using the trypan blue dye exclusion. Cell isolations with >90% viability were plated in 150-cm2 culture flasks at an initial density of 6×103 cells/cm2 and cultured in standard chondrocyte media consisting of Ham's F-12 medium with L-glutamine supplemented with 10% fetal bovine serum (Sigma-Aldrich, St. Louis, MO), 50 mg/mL ascorbic acid, 100 U/mL penicillin, and 0.1 mM nonessential amino acids at 37°C in 5% CO2. Media were changed three times per week. The cells were removed from the flasks with 0.5% trypsin-EDTA (wt/vol) (Gibco, Carlsbad, CA) when they reached 90% confluence and washed twice with PBS. Articular cartilage was also harvested from the knees of 3-month-old swine and processed in the same fashion as human chondrocytes. Passage 1 (P1) cells were used for the construct assembly.
Construct assembly and implantation
Articular chondrocytes from both healthy human donors and young swine were suspended in a FG solution. To make the FG, bovine fibrinogen (Sigma, St. Louis, MO) was prepared in sterile saline at a concentration of 80 mg/mL. 3 Thrombin (GenTrac, Middleton, WI) was diluted to 50 U/mL in saline. One hundred twenty-five microliters of cells in the fibrinogen solution and an equal volume of thrombin solution were placed into a bone wax well measuring 0.8×0.8 cm and mixed with the micropipette. Final nodule construct volume was 0.8×0.8×0.35 cm. Based on our previous experience, the cell concentration of swine chondrocytes was 40×106/mL3,7,8 and the human chondrocytes was 60×106/mL.
To evaluate the integration of neocartilage with native articular cartilage, three-layered constructs were assembled as previously described. 7 The FG solution containing either human or swine chondrocytes was prepared in the same way as explained previously. Discs of human and swine cartilage were made with a 6-mm biopsy punch and devitalized by a previously documented procedure using five freeze-thaw cycles. 9 Subsequently, 100 μL of the cell/FG was placed on the top of one disc and a second disc was placed on the top of the cells in the gel, creating a cartilage–cell/hydrogel–cartilage construct to determine cartilage integration. A 7–0 monofilament suture was used to secure the layers in place. After the in vivo period, the suture was removed and constructs were tested for integration.
All constructs (nodule constructs, n=60; three-layered constructs, n=72) were implanted into subcutaneous pockets on the backs of nude mice (Massachusetts General Hospital). Samples were harvested at 12, 18, and 24 weeks.
Histological analysis
Nodules (n=3) and three-layered constructs (n=3) were randomly selected from each of the time points and were placed in 10% phosphate-buffered formalin for 24 h. They were subsequently embedded in paraffin and sectioned. Sections were prepared from a minimum of two areas from each specimen and were stained with hematoxylin and eosin (H&E) for evaluation of gross morphology of the neotissue. Other sections were stained with safranin-O and toluidine blue to verify the production of sulfated glycosaminoglycans (sGAG). The sections were evaluated in a blinded manner by three independent reviewers, to determine the presence or absence of neocartilage formation.
Immunohistochemical analysis
Mouse anti-human collagen type I (COL I) antibody (Accurate Chemical & Scientific Corp., Westbury, NY) and mouse anti-human collagen type II (COL II) antibody (Developmental Studies Hybridoma Bank, Iowa City, IA) were used. Prior to use, the antibody was diluted 1:1000 in 1% bovine serum albumin (Gibco) in PBS. For negative control, N-universal negative control (DAKO, Glostrup, Denmark) was used. The incubation time was 60 min. Sections were washed extensively in PBS and an appropriate secondary antibody (Dako) was used for 20 min, followed by wash in PBS (3×5 min). Detection of the immunoreaction was achieved by adding 3, 3-diaminobenzidine (Dako), which was used as substrate chromogen for 5 min. Slides were washed in distilled water and counterstained with hematoxylin to visualize the cell nuclei.
Biochemical analysis
Quantification of sGAG was determined with the dimethylene blue method and the hydroxyproline was determined using the chloramine-T method in the nodule constructs as previously described by our laboratory. 8 Biochemical analysis was performed on four to six samples per group at each time point. One 40–50 mg piece of engineered cartilage tissue was collected from each nodule specimen and those pieces were weighed and lyophilized for 24 h. Dehydrated specimens were digested with papain solution (125 μg/mL papain type III, 100 mM phosphate, 10 mM L-cysteine, and 10 mM EDTA [pH 6.3]) at 60°C for 16 h. Both sGAG and hydroxyproline content of the specimens were determined by using shark cartilage (Sigma) as a standard. Samples and standards were analyzed in duplicate and reported as a percentage of wet tissue weight as well as dry tissue weight.
Biomechanical analysis
Samples were frozen at −80°C immediately after harvest for biomechanical testing and thawed at room temperature in PBS with proteinase inhibitors immediately before testing, when the fibrotic capsule surrounding the constructs was removed.
Compression testing (nodule constructs)
Samples were cut into disks with a 6-mm diameter using a disposable biopsy punch and placed in a confining chamber that was mounted in an Enduratec EL2100 mechanical testing frame using a 10-N load cell (Bose, Eden Prarie, MN). Samples were equilibrated at room temperature in 0.15 M PBS (pH 7.4) containing 100 U/mL penicillin G and 100 mg/mL streptomycin.
After mounting each disk in the confined compression chamber, the distance between the porous platen and the chamber was decreased until a signal of ∼5 g (50 mN) was detected by the load cell. This distance was taken to be the sample thickness. Each disk was compressed by 10 sequential increments of 2.5%–3.0% static strain, up to a maximum of 25%–30% total strain. After each increment, the load was recorded every 0.5 s for 100 s. Stress relaxation data were fit to a poroelastic model of material behavior to calculate the equilibrium modulus. 10
Tensile testing (three-layered construct model)
The integration of engineered cartilage with native cartilage was assessed by pulling three-layered constructs to failure in tension, a technique described by our group in several studies over the last decade.7,11,12 The flat ends of the constructs were attached to Plexiglas rods, using quick-setting cyanoacrylate glue. The rods were then mounted in the jaws of the Enduratec EL2100 mechanical testing frame. One milliliter of PBS was placed around the constructs to prevent sample dehydration. Tensile displacements were applied to the constructs at a rate of 4–10 μm/s, corresponding to a bulk strain rate of 0.15%/s for each sample, and the resultant load was recorded at a rate of 5 points/sec. Samples were pulled to failure in tension as indicated by either visible separation of cartilage disks or a measured load of <0.05 N. Applied displacements and measured loads were normalized to sample thickness and area, respectively. Using these data, a stress–strain curve was constructed for each specimen. From the stress–strain curve, the ultimate tensile strength (σUTS) was determined by inspection as the highest stress supported by the sample, and the failure strain (ɛf) was determined by inspection as the strain at which further increases in strain produced lower stresses. The failure energy (Ef), defined as the area under the stress–strain curve until failure, was calculated by a Reimann sum method with the partition element given by the strain interval between data points. The dynamic tensile modulus (M) for the given strain rate of each sample was calculated as the slope of the linear portion of the stress–strain curve, using a standard least squares algorithm, with r2 values >0.9 as criteria for acceptable linearity.
Statistical analysis
All the statistical analysis was performed using two-way ANOVA to test the differences in mean value of GAG and hydroxyproline content as well as the biomechanical parameters of the neocartilage and three-layered construct model constructs over the three time points in this study. Mean values were reported with standard deviations. Significance was assigned at p<0.05.
Results
All samples were recovered from nude mice at 12, 18, and 24 weeks. For the nodule constructs, the initial shape was conserved over time in all the experimental groups. The specimens resembled cartilage in color and texture and were resistant to external compression. For the three-layered constructs, the native articular cartilage disks maintained their glistening white surface throughout the duration of implantation. Neotissue formation in the intradisk space ranged from pinkish white to pearlescent in all the experimental groups. Although the appearance of the neotissue was notable in the intradisk space, there were no obvious gaps between the new cartilage and the native cartilage disks (Fig. 1).

Gross appearance. Nodule construct made with elder human articular chondrocytes before
Histological evaluation
For the nodule constructs H&E staining of samples made with chondrocytes from both middle-aged human sources and JSCs exhibited a similar histomorphology characteristic of cartilage (cells within basophilic ground substance) at all time points. The presence of sGAG was also evident with intense safranin-O and toluidine blue staining at all time points from all the experimental groups (Fig. 2).
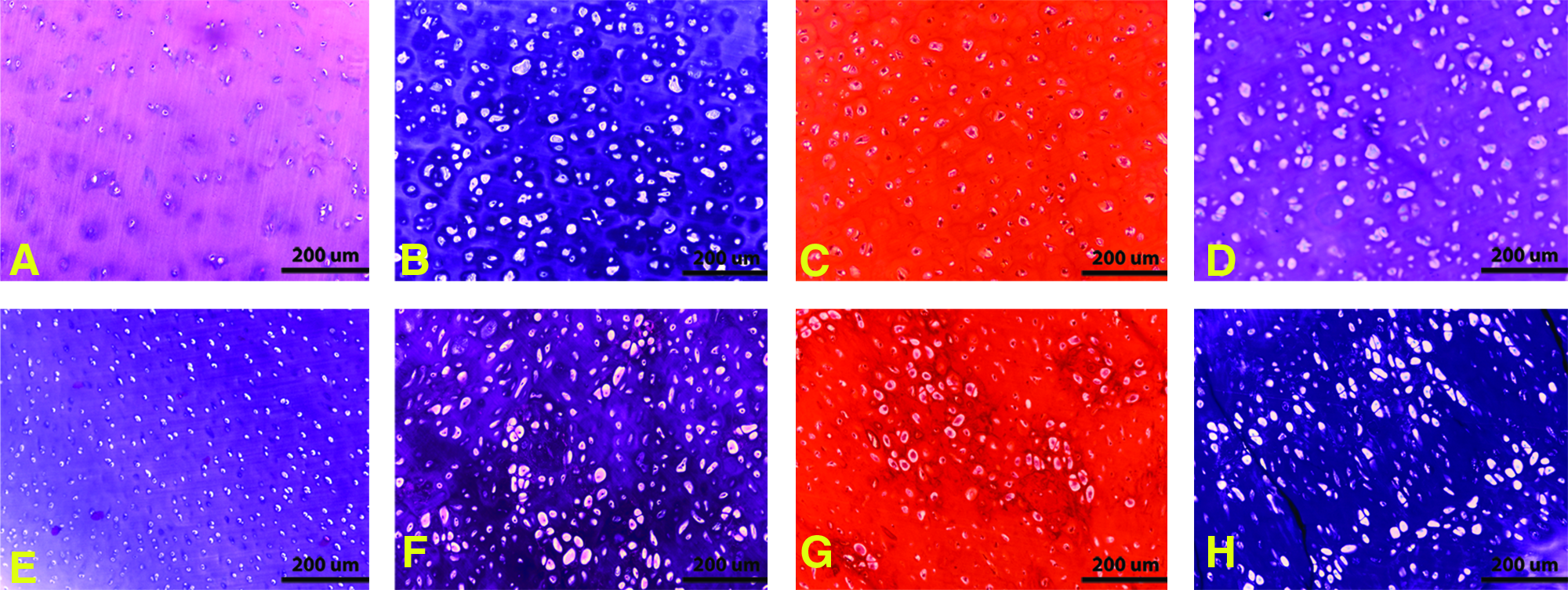
Histological evaluation of nodule samples (after 18 weeks in vivo). Samples made with elder human articular chondrocytes were stained with hematoxylin and eosin (H&E)
For the three-layered constructs, H&E staining of samples made with native cartilage disks and chondrocytes from both middle-aged human sources and JSCs revealed similar new ECM formation of neocartilage tissue within all intradisk areas. Increased red staining with safranin-O as well as increased blue staining with toluidine blue revealed the presence of abundant sGAG in the newly formed tissue. Staining was less intense in the native cartilage due to the freeze-thaw devitilization process used to make the nonviable native cartilage disks, which was consistent with the findings in our previous experiments.7,13 In all of the experimental groups, the new matrix formed between the native cartilage disks appeared to fill all surface irregularities and in multiple samples there was evidence of neocartilage penetrating into the adjacent cartilage disks. There were no gaps observed at the interfaces of the neotissue and the native cartilage disks (Fig. 3).
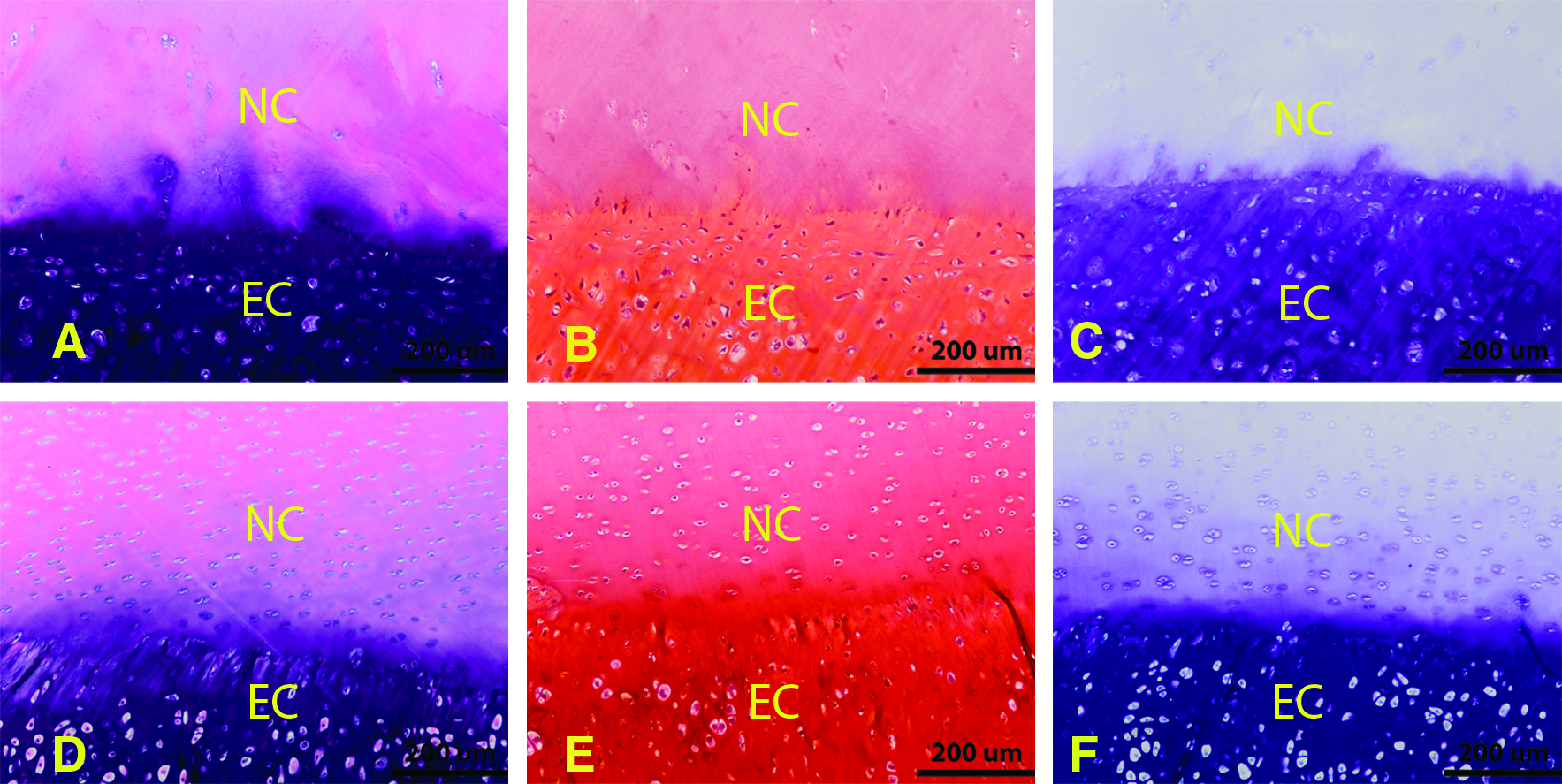
Histological evaluation of three-layer constructs (after 18 weeks in vivo). Human cell source samples were stained with H&E
Immunohistochemical staining
Immunostaining for COL I was negligible in both the nodule constructs and the three-layered constructs. By contrast, immunostaining for type II collagen demonstrated uniform staining throughout the new ECM on the nodule constructs. The three-layered constructs also showed the presence of type II collagen in both the new cartilage matrix and the native cartilage disks (Fig. 4).
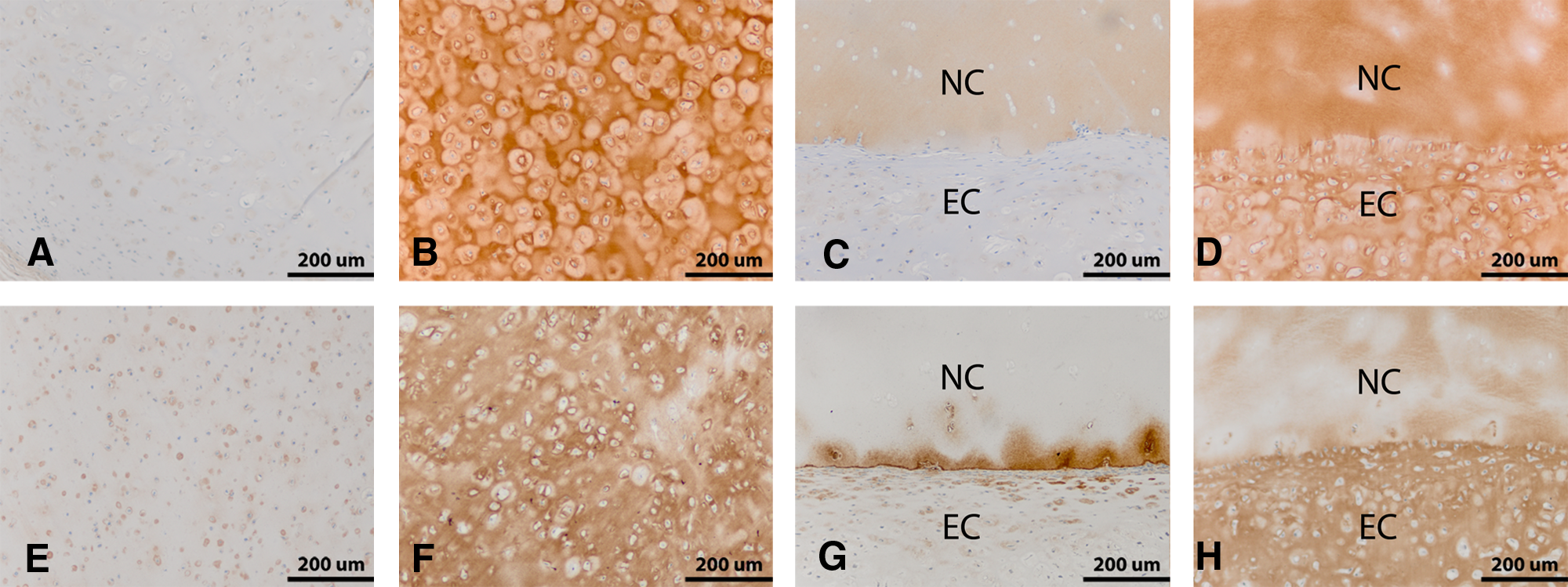
Immunohistochemical evaluation of nodule and three-layer samples (after 18 weeks in vivo). Nodule constructs made with elder human articular chondrocytes were stained for type I
Biochemical analysis
GAG content
The amount of GAG in the native JSCs was 453±32 μg/mg dry tissue (Fig. 5). However, the amount of GAG measured in the native human cartilage (211±31 μg/mg dry tissue) was less than half of that measured in the native swine tissue. At the 12-week time point, the amount of GAG in the swine chondrocyte nodules was 264±12 μg/mg dry tissue, or slightly more than half of the native swine cartilage, whereas the amount of GAG in the nodules made with human chondrocytes was 326±26 μg/mg dry tissue—about 154% of the amount measured in the native human tissue. A similar result was observed for the human chondrocyte nodules at 18 weeks, but the amount in the swine chondrocyte constructs at 18 and 24 weeks was only about 11%–15% of the amount measured in the native swine.
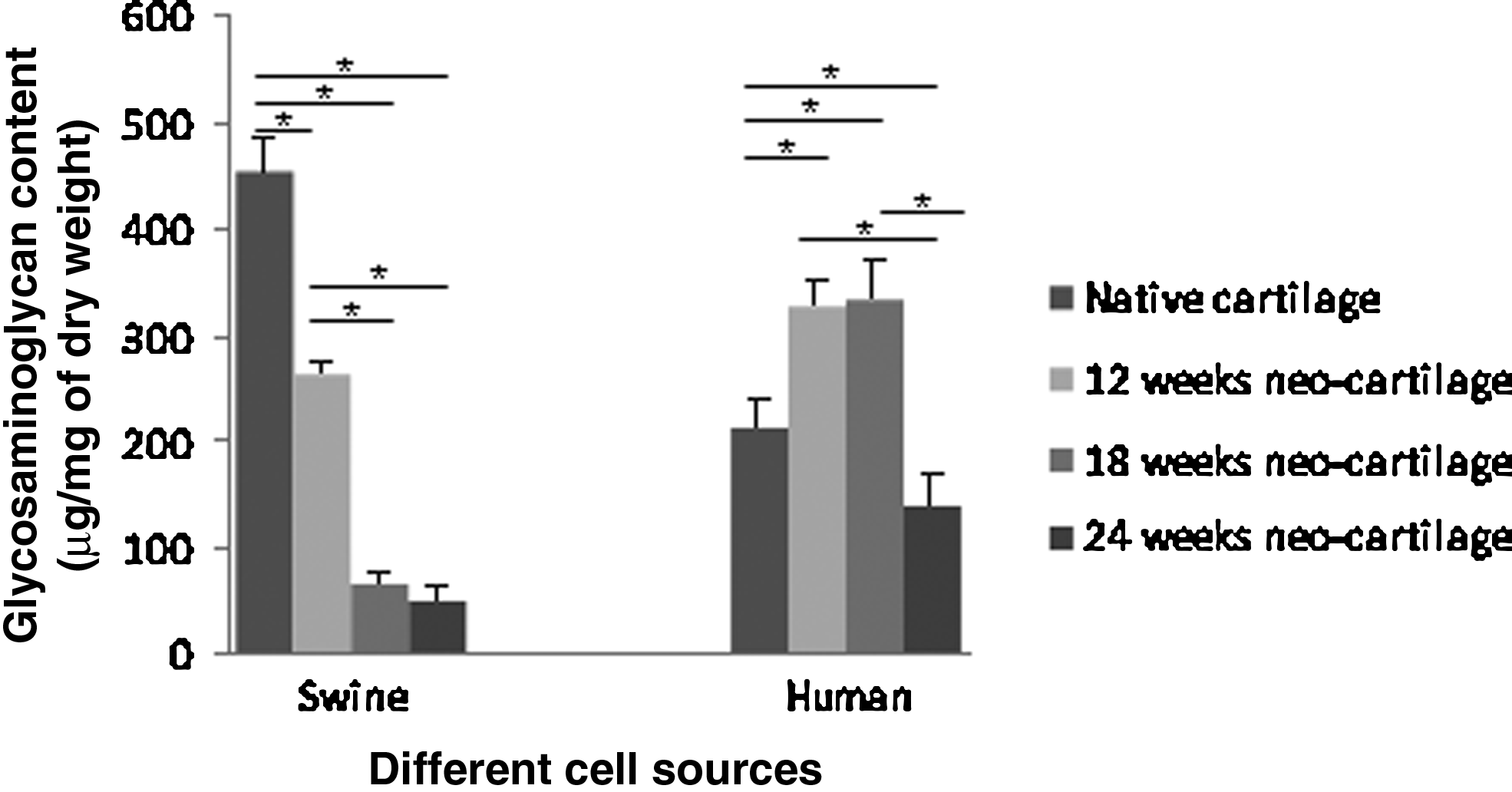
Glycosaminoglcyan (GAG) contents are shown in both wet weight and dry weight (n=4–5). There was statistically significant difference in the amounts of GAG between tissue-engineered cartilage and corresponding native cartilage at all the time points (p<0.05). For swine source samples, there was no statistically significant difference in the amounts of GAG between specimens harvested at 18 and 24 weeks (p>0.05); however, both of which had a statistically significant difference comparing with the specimens harvested at 12 weeks (p<0.05). For human source samples, there was no statistically significant difference in the amounts of GAG between specimens harvested at 12 and 18 weeks (p>0.05); however, both of which had a statistically significant difference comparing with the specimens harvested at 24 weeks (p<0.05). (*Indicates statistically significant difference; p<0.05).
Hydroxyproline content
The amount of hydroxyproline in the native human tissue was 39±10 μg/mg dry tissue and that in the native swine tissue was 35±5 μg/mg dry tissue. These initial amounts were not statistically different from each other (p>0.05). The amount of hydroxyproline in the nodule specimens made with middle-aged human articular chondrocytes at 12, 18, and 24 weeks was 19±4, 15±6, and 25±2 μg/mg dry tissue, respectively. By comparison, the amount of hydroxyproline in the nodule specimens made with articular JSCs at 12, 18, and 24 weeks was 18±2, 16±4, and 29±1 μg/mg dry tissue, respectively. There were no statistical differences between the amounts of hydroxyproline for human chondrocytes versus swine chondrocytes at each time point. However, the amount of collagen in the test specimens at each time point for both human and swine chondrocytes was statistically different from the level in the native cartilage (p<0.05) (Fig. 6).

Hydroxyproline contents are shown in both wet weight and dry weight (n=4–5). There was statistically significant difference in the amounts of hydroxyproline between tissue-engineered cartilage and corresponding native cartilage at all the time points (p<0.05). There was also statistically significant difference in the amounts of hydroxyproline among the three experimental groups harvested at 12, 18, and 24 weeks (p<0.05) for both swine and human source samples. (*Indicates statistically significant difference; p<0.05).
Biomechanical analysis
Equilibrium modulus (applied to nodule constructs)
The equilibrium modulus of cartilage nodule samples grown from human and swine chondrocytes showed similar values at 12 and 18 weeks (Fig. 7). However, both the human and swine samples experienced a small drop in modulus at 24 weeks. It should be noted that the modulus of the cartilage with either JSCs or human cells never exceeded 10% of the modulus of native cartilage (juvenile swine cartilage: 457±11 kPa; human native cartilage: 332±12 kPa). The equilibrium modulus of samples grown from human and swine chondrocytes differed by 10%–40% across the time of the study, with no statistical difference between the two cell types at any time point.
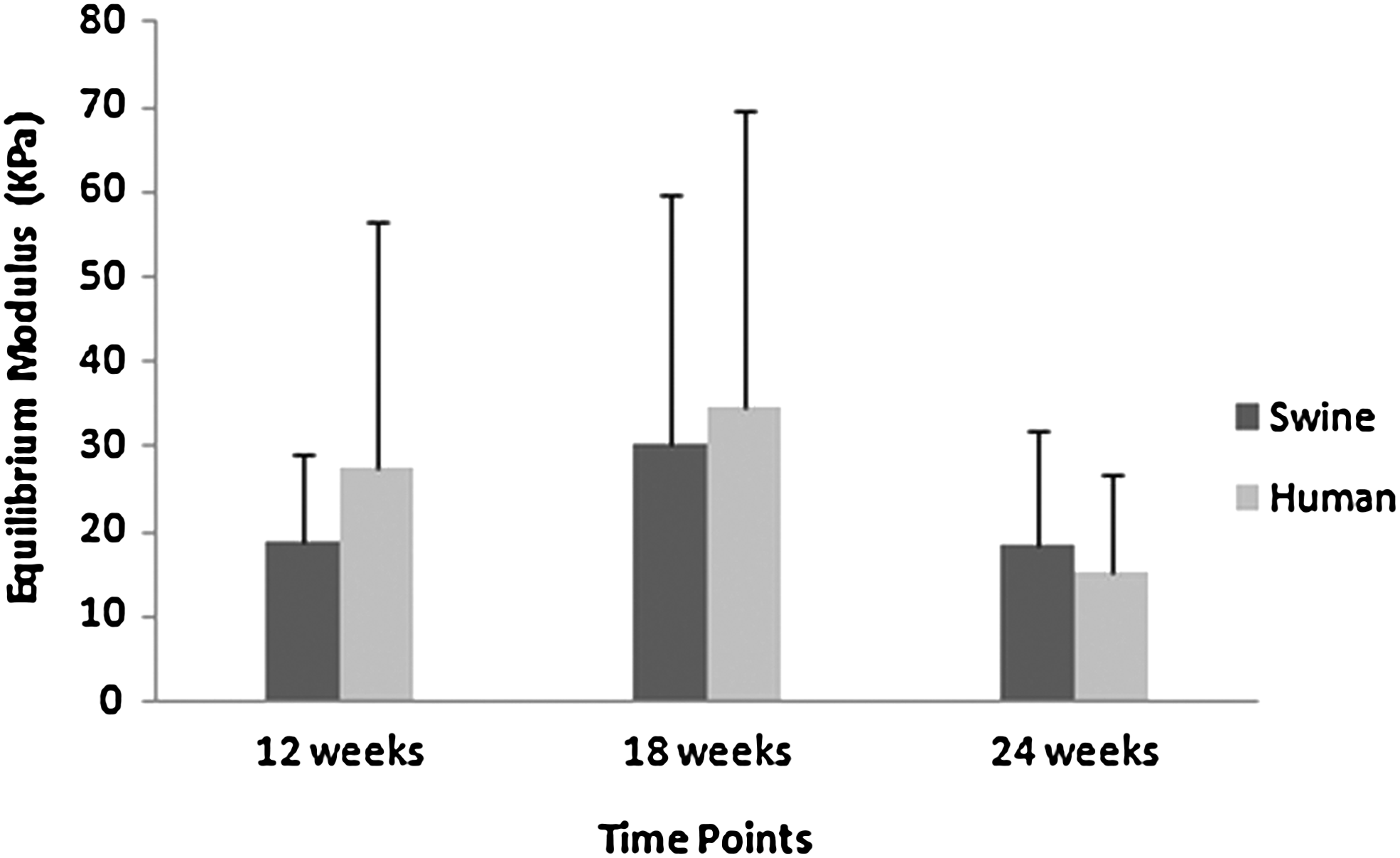
Equilibrium modulus (n=8–12). There was no statistically significant difference in the equilibrium modulus between the human chondrocyte and swine chondrocyte groups at each time point (p>0.05).
Tensile properties (applied to three-layered constructs)
The tensile properties of three-layered samples made with swine chondrocytes differed from those made with human chondrocytes over time (Fig. 8). The UTS and the tensile modulus made from JSCs increased over time and were consistently higher than those with human cells, with greater than a three-fold difference in UTS at 24 weeks (p<0.05) and more than a seven-fold difference in tensile modulus at 24 weeks (p<0.05). The UTS and tensile modulus of the human chondrocyte samples decreased from 18 to 24 weeks.

Ultimate tensile strength and tensile modulus (n=8–12). There was no statistically significant difference between human chondrocyte and swine chondrocyte groups at both 12 and 18 weeks (p>0.05). However, there was statistically significant difference at 24 weeks (p<0.05). (*Indicates statistically significant difference; p<0.05).
In contrast to the trends seen in tensile strength and modulus data, the failure strain was higher in samples made from adult human cells, with a 90% difference measured at 24 weeks. The failure energy of samples made from human and swine cells was similar at 12 weeks, but was ∼40% higher in swine samples at 18 weeks and more than two-fold higher at 24 weeks (p<0.05) (Fig. 9).

Failure modulus and failure energy (n=8–12). There was no statistically significant difference between human chondrocyte and swine chondrocyte groups at both 12 and 18 weeks (p>0.05). However, there was statistically significant difference at 24 weeks (p<0.05). (*Indicates statistically significant difference; p<0.05).
Discussion
Most of the published work on engineering cartilage on biodegradable scaffolds, both in vitro and in vivo, has used juvenile animal chondrocytes. This may be due to the ease of acquiring large numbers of chondrocytes from young animals for experimentation and the difficulty in obtaining sufficient amounts of healthy human tissue. Our own studies have predominantly employed JSCs to generate cartilage in vivo in nude mice or in situ in young adult animals.5,7,8,14 Nonetheless, these studies have built a solid foundation of experimental animal data demonstrating that the concepts of tissue engineering can be employed to produce new cartilage tissue that can potentially heal defects in the articular surface of joints.
ACI has established that chondrocytes can be harvested, expanded in culture, and implanted into cartilage defects to stimulate a reparative process.15,16 Previous studies have suggested, however, that articular chondrocytes isolated from aged human and animal cartilage sources could have an impaired capacity to produce ECM.17–20 This is crucial since patients sustaining injuries may more likely be middle aged or older. Far less is known about engineering cartilage for joint surface regeneration in these age groups, which makes the study of chondrocytes from middle-aged humans necessary and clinically relevant.
Our team has extensive experience using different hydrogels for encapsulating animal chondrocytes to generate ECM for possible knee cartilage repair.5,9,21–24 While it is well established that human chondrocytes will make new ECM, few studies have made a direct comparison between human and animal chondrocytes to engineer neocartilage tissue. 25 In this study, we made a direct comparison between the ECM generated by JSCs and that made by chondrocytes from middle-aged human sources when both were encapsulated in the same fashion in a FG polymer. The results indicate that articular chondrocytes from middle-aged human donors do, in fact, have the capacity to generate tissue-engineered cartilage in vivo when encapsulated in this hydrogel scaffold. Generally, the engineered neocartilage produced with articular chondrocytes from these middle-aged human donors has histological, biochemical, and biomechanical characteristics similar to that of neocartilage made with articular chondrocytes from JSC donors. Thus, the data generated from this study suggest that it would be possible to use autologous articular chondrocytes to engineer neocartilage for joint resurfacing and repair in the middle-aged population.
The histological and immunohistochemical results in constructs made with human chondrocytes were indistinguishable from those using JSCs. The histological staining of the ECM was uniform across the specimens in both groups and the safranin-O images demonstrated the production of sGAG typically observed in native cartilage. COL II, a significant component of the ECM of hyaline articular cartilage, defines the biomechanical properties of this tissue. The immunostaining of the native cartilage from both groups showed intense staining for COL II and faint-to-negligible staining for COL I as would be expected. The engineered cartilage specimens from both groups showed similar staining patterns indicating the production of COL II and little staining for COL I. There were no observable differences between the engineered specimens made with JSCs and those made with middle-aged human chondrocytes.
In addition to the histology, the biochemical analysis revealed that the encapsulated cells were generating GAG, particularly in the early phase of in vivo incubation. The amount of GAG measured in the native JSCs was nearly twice that measured in the native human cartilage. The samples made with JSCs demonstrated moderate amounts of GAG in the 12-week samples—about half that of the native cartilage. For unexplained reasons, the swine specimens harvested at 18 and 24 weeks had much less GAG. By contrast, in the groups of specimens made with middle-aged human chondrocytes, the amount of sGAG was about 30%–40% higher than native cartilage at 12 and 18 weeks. This suggests that these aged cells liberated from their native matrix can be reinvigorated to produce large amounts of GAG as has been reported previously in chondrocytes from old-aged sheep. 26 The amount of GAG measured at 24 weeks remained about 75% of native human cartilage. These results indicated that the middle-aged human chondrocytes encapsulated in FG polymer retained a strong ability to produce high amounts of GAG. A drop in the amount of GAG was noted in the later time points in constructs from both cell sources. This decline may be due to the lack of biomechanical forces in this subcutaneous implant model. This decline was noted by week 18 in the samples made with juvenile swine cells, whereas the constructs made with human cells did not decline until the 24 week specimens. These results may be due to inherent differences between the species or the different ages of human versus the swine cells used in this experiment.
Whereas the GAG content of native swine cartilage was nearly double that of native human cartilage, the amount of collagen in the human cartilage and JSCs was not significantly different. The total amount of collagen from all the engineered groups was lower than that found in the native cartilage—generally about 50%. These results are in accordance with most of the published data obtained regarding tissue engineering cartilage with chondrocytes from young donors and chondrocytes from different anatomical sources.8,27 Therefore, it seems that the low production of collagen by chondrocytes on scaffolds is a fundamental problem of tissue engineering cartilage and not specifically dependent on donor age and species.
In addition to morphological and biochemical evidence that the engineered cartilages from both sources of cells were somewhat comparable, we sought to compare the biomechanical parameters of the new ECM generated by chondrocytes from juvenile animals and those from middle-aged humans. The reported equilibrium moduli of engineered cartilage generally are an order of magnitude lower than the native cartilage (juvenile swine cartilage: 457±11 kPa; human native cartilage: 332±12 kPa) from which the chondrocytes are collected. The results from confined compression testing of the nodules in this study show that the ECM from both groups is less stiff than native cartilage, but that the moduli in both groups are comparable. The weakness of the ECM could also be an artifact of the long-term subcutaneous implant model where the cartilage is not subjected to the normal biomechanical forces found in the joint.
The capacity of the cartilage to integrate with the existing cartilage is another characteristic important for the clinical application of engineered cartilage. Using a three-layered construct, we sought to determine whether the new cartilage would integrate with native cartilage by employing a tensile test. The histological results demonstrated that the neotissue filled the intradisk space without any obvious gaps. Mechanical testing revealed that the UTS increased over time in the group using swine cells. Similar results were obtained in specimens using human chondrocytes at 12 and 18 weeks, but the UTS of human specimens at 24 weeks was much lower than the swine specimens.
It is also noteworthy that the temporal trends in compressive and tensile mechanical properties were quite different over the course of 24 weeks. Compressive properties declined from 18 to 24 weeks, when the equilibrium modulus was 2%–4% of native tissue and may be related to the drop in GAG production. In contrast, the tensile modulus of swine samples at 24 weeks (1.7 MPa) was 10%–20% that of native cartilage. 28 This is consistent with observations of continued collagen deposition at later time points and may suggest that long-term implantation in this system becomes more favorable for collagen network formation and less favorable for proteoglycan maintenance. Further, this is also consistent with other studies demonstrating that proteoglycan loss can enhance collagen network integrity, particularly in immature tissues. 29 Although the collagen content of specimens in the nodules from human and swine cells was not significantly different, the tensile properties of the three-layered human cell constructs show a weaker integration at 24 weeks. The collagen produced in the nodules may not be parallel to the collagen production in the three-layered constructs. Another explanation may be that the collagen produced between the cartilage disks in the three-layered constructs might not contribute to the integration strength in a linear fashion. Collectively, these data suggest that there are long-term (+18 weeks) remodeling processes that occur in tissue-engineered cartilage samples that affect the mechanical behavior of the tissue.
The results from this study demonstrated that chondrogenesis using articular chondrocytes from middle-aged people can be achieved in a predictable and reliable manner. Although the data are not identical to cartilage engineered with JSCs, there were many similarities noted in histological appearance, compressive moduli, and collagen production. Since most patients with articular cartilage lesions are not juveniles, but are quite often middle aged or older, it makes this head-to-head study comparing JSCs to human cells from middle-aged people to engineer cartilage more meaningful because it moves these cell–scaffold technologies closer to possible clinical application. Although the study design could have included groups from juvenile humans or older-aged animals, it was not logistically possible because obtaining cartilage from that human age group (5–15 years) is nearly impossible and old age, large animals are not commercially available. Further, the aim of this study was to determine whether cells from middle-aged, or even older, people have the capacity to generate cartilage ECM that is comparable to that observed by cells from juvenile animals, which has been the focus of most reported experimental cartilage engineering studies. Evaluating these parameters will help in developing strategies to apply tissue engineering approaches clinically.
Footnotes
Acknowledgments
This work was supported by a grant from National Football League Charities. Special thanks to Dr. Xing Zhao's wife Ying Zhou, Ph.D., candidate at the Boston University School of Medicine, for assistance on the statistical analysis.
Disclosure Statement
No competing financial interests exist.