
Abstract
Despite the fact that the peripheral nervous system is able to regenerate after traumatic injury, the functional outcomes following damage are limited and poor. Bone marrow mesenchymal stem cells (MSCs) are multipotent cells that have been used in studies of peripheral nerve regeneration and have yielded promising results. The aim of this study was to evaluate sciatic nerve regeneration and neuronal survival in mice after nerve transection followed by MSC treatment into a polycaprolactone (PCL) nerve guide. The left sciatic nerve of C57BL/6 mice was transected and the nerve stumps were placed into a biodegradable PCL tube leaving a 3-mm gap between them; the tube was filled with MSCs obtained from GFP+ animals (MSC-treated group) or with a culture medium (Dulbecco's modified Eagle's medium group). Motor function was analyzed according to the sciatic functional index (SFI). After 6 weeks, animals were euthanized, and the regenerated sciatic nerve, the dorsal root ganglion (DRG), the spinal cord, and the gastrocnemius muscle were collected and processed for light and electron microscopy. A quantitative analysis of regenerated nerves showed a significant increase in the number of myelinated fibers in the group that received, within the nerve guide, stem cells. The number of neurons in the DRG was significantly higher in the MSC-treated group, while there was no difference in the number of motor neurons in the spinal cord. We also found higher values of trophic factors expression in MSC-treated groups, especially a nerve growth factor. The SFI revealed a significant improvement in the MSC-treated group. The gastrocnemius muscle showed an increase in weight and in the levels of creatine phosphokinase enzyme, suggesting an improvement of reinnervation and activity in animals that received MSCs. Immunohistochemistry documented that some GFP+ -transplanted cells assumed a Schwann-cell-like phenotype, as evidenced by their expression of the S-100 protein, a Schwann cell marker. Our findings suggest that using a PCL tube filled with MSCs is a good strategy to improve nerve regeneration after a nerve transection in mice.
Introduction
The recent use of artificial nerve conduits filled with a variety of proregenerative substrates shows great promise as an alternative to nerve autografts: in particular, the polycaprolactone (PCL) conduit is biodegradable, has a low risk of being rejected, and has already been shown to be a suitable substrate for the survival and differentiation of Schwann cells,6–9 and the combination of PCL with the use of different cell types, such as fibroblasts, Schwann cells, or bone marrow-derived cells, reportedly improves peripheral nerve regeneration after severe injuries.2,8,10–13
We argue that the use of PCL conduits filled with bone marrow-derived mesenchymal stem cells (MSCs) has potential as an effective alternative therapy, since these cells are multipotent, and have the capacity for differentiating into multiple cellular lineages such as chondrocytes, osteoblasts, and adipocytes; according to some authors MSCs express neurotrophic factors and can also differentiate into nerve cells.14–16 Bone marrow-derived MSCs can be aspirated from patients and are easily isolated and manipulated in vitro, avoiding risks of transplant rejection. The present study is aimed at evaluating regeneration and neuronal survival when transected sciatic nerves in mice are treated with the use of PCL conduits transplanted with MSCs: we also assess the ability of the MSCs to differentiate into Schwann-like cells and to restore target organ function. Our findings suggest that the use of a PCL tube filled with MSCs is a highly promising strategy to improve the reconstruction of a transected peripheral nerve in mice.
Materials and Methods
Animals
Twelve C57BL/6 female mice (20–23 g) were randomly divided into two groups: those with MSC grafts (MSC-treated group, n=6) and those with acellular grafts and treated with a culture medium (Dulbecco's modified Eagle's medium [DMEM] group, n=6). All animal use and care protocols were approved by the Ethics Committee for the Use of Experimental Animals of the Universidade Federal do Rio de Janeiro (Protocol DHE003).
MSC culture
MSCs were prepared from C57BL/6 mice expressing the green fluorescent protein (GFP mice), allowing the subsequent identification of these cells. The bone marrow of GFP mice was harvested from their femurs and tibias using 22G or 30G syringes, respectively, and Hank's buffered saline solution (HBSS). Marrow was centrifuged for 5 min at 2000 rpm, and the pellet was seeded into 25-cm2 culture flasks at a density of 5×107 containing the DMEM supplemented with 20% fetal bovine serum and 100 U/mL penicillin. Nonadherent cells were removed by replacing the medium and adherent cells were continuously cultured, passaged three times, and then used for the experiments, at a density of 1×105 cells in 2 μL. After surgery, a sample of the remaining cells was plated overnight to assess cell viability.
Surgical procedure
All surgeries were performed under deep anesthesia using i.p. injection of ketamine (100 mg/kg) and xylazine (15 mg/kg). The left sciatic nerve was exposed without damaging the surrounding muscles and transected at a mid-thigh level. A 10.0 monofilament nylon suture was used to suture 1 mm of the proximal nerve stump through the epineurium into a policaprolactone conduit (5 mm long). The MSC-treated group received an injection of MSC in the DMEM (1×105/2 μL, n=6) into the conduit and the DMEM group received only the DMEM (2 μL, n=6). Injections were made with a Hamilton microsyringe. One millimeter of the distal stump was sutured to the other end of the conduit, leaving a 3-mm gap between the stumps.
Six weeks later, the animals were anesthetized with ketamine and xylazine, and then perfused transcardially with 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4). The regenerated left sciatic nerve within the tube, the L4 spinal cord, and the L4 dorsal root ganglion (DRG) were removed and prepared for immunohistochemistry, light microscopy, and transmission electron microscopy.
Morphological analysis
Transmission electron microscopy
The middle segment of the regenerated nerves was immersed for 2 h in 2.5% glutaraldehyde in 0.1 M phosphate buffer (pH 7.4), washed in 0.1 M cacodylate buffer (pH 7.4), and postfixed for 90 min in 1% osmium tetroxide containing 0.8% potassium ferrocyanide and 5 nM calcium chloride in 0.1 M cacodylate buffer (pH 7.4). The segments were washed in 0.1 M cacodylate buffer (pH 7.4) and stained in 1% uranyl acetate overnight in the dark, dehydrated in increasing concentration of acetone, infiltrated with Poly/Bed 812 resin (Polysciences, Inc.), and polymerized at 60°C for 48 h. Semithin (500 ηm) and ultrathin (70 ηm) cross sections were obtained with an ultramicrotome (MT-6000-XL; RMC, Inc.). The semithin sections were stained with toluidine blue and examined on a light microscope (Zeiss Axioskop 2 plus), while the ultrathin sections were collected on copper grids and contrasted in 5% uranyl acetate and 1% lead citrate and analyzed on a transmission electron microscope operated at 80 kV (Zeiss 900).
Myelinated fibers quantification
Photographs of the semithin cross sections taken under the light microscopy at a 63× magnification were used for quantification. The entire nerve diameter was photographed and the total number of myelinated fibers in each regenerated nerve was counted using ImageJ Software (1.42q).
Tissue preparation for light microscopy and immunohistochemistry
The distal segment of the regenerated sciatic nerve, the L4 DRGs, and L4 spinal cord were removed from perfused mice, washed in 0.1 M phosphate buffer (pH 7.4), cryoprotected with 10%, 20%, and 30% sucrose for 24 h each, and mounted in Tissue-Tek® O.C.T. Compound (Sakura Finetechnical). Frozen longitudinal serial sections of DRGs (12 μm) and transverse serial sections of the sciatic nerve (10 μm) and the spinal cord (20 μm) were cut on a cryostat (Leica CM, 1850) and mounted onto gelatin-coated slides.
Neuronal quantification
Slides with DRGs and spinal cord (8–10 sections per slide) were stained with cresyl violet for quantification of primary sensory neurons and motor neurons, respectively. Sections were photographed on the Zeiss Axioskop 2 plus microscope at a 20× magnification. All nucleoli of neurons in the DRGs sections and of neurons in the ventral horn of the spinal cord were counted using ImageJ Software (1.42q). Counting from 8–10 sections allowed the quantification of almost all their structures.
Immunohistochemistry analysis
Sections through the distal segment of the regenerated sciatic nerve were prepared to assess the presence of the GFP+ -grafted cells and the expression of the Schwann cell marker S-100. Slides were washed for 5 min in 0.1 M phosphate-buffered saline (PBS; pH 7.4), incubated in blocking solution containing 10% normal goat serum (Invitrogen), and 3% bovine serum albumin in 0.3% Triton X-100 and 0.1 M PBS for 1 h and then washed twice in 0.3% Triton X-100 with 0.1 M PBS for 5 min each. Sections were incubated in the primary antibody (anti S-100, 1:100; Sigma) overnight at 4°C in a humid chamber followed by two washes in 0.3% Triton X-100 with 0.1 M PBS for 15 min each. Slides were incubated in the secondary antibody (Alexa Fluor 546 goat anti-rabbit IgG, 1:600; Invitrogen) for 2 h at room temperature and washed three times in PBS (pH 7.4) for 10 min each. Nuclei were stained with DAPI for 5 min (1:10,000; Sigma); the slides were washed three times in PBS (pH 7.4) followed by 5-min wash with distilled water and finally mounted with Fluoromount (Sigma). The primary antibody was omitted for negative controls. All sections were analyzed under confocal microscopy (Disk scanning unit; Olympus).
Trophic factor quantification
We evaluated the trophic factor levels in the tissue, by performing an immunofluorescence analysis for the brain-derived neurotrophic factor (BDNF), the nerve growth factor (NGF), neurotrophin 3 (NT-3) and NT-4 on the spinal cord, DRG and sciatic nerve sections. We used the same protocol described above and the primary antibodies used were rabbit anti-human BDNF (Preprotech; 1:100), rabbit anti-human NGF (Preprotech; 1:100), goat anti-mouse NT-3 (Preprotech; 1:100), and goat anti-mouse NT-4 (Preprotech; 1:100). After primary incubation, the slides were washed and incubated with the appropriate antibodies Alexa 488 goat anti-rabbit (Sigma; 1:600) or Alexa 488 rabbit anti-goat (Sigma; 1: 600) for 2 h at room temperature, followed by three washes, and coverslipped with Fluoromount (Sigma). The immunofluorescence images were analyzed with the Image-Pro Plus (version 6.0), by evaluating the ratio between the stained area and the total field area. We took pictures from all the sections using 20× objective. After that, the trophic factors BDNF, NGF, NT-3, and NT-4 were quantified using these images. Data are presented as mean±standard error of the mean.
Analysis of gastrocnemius muscle: weight and creatine phosphokinase activity
Before perfusion, the gastrocnemius muscles from both limbs were dissected and immediately weighed to analyze its trophism as an indirect measurement of nerve regeneration and muscle reinnervation. The muscles were then placed in physiological saline solution (135 mM NaCl, 5 mM KCl, 2 mM CaCl2, 1 mM MgCl2, 15 mM NaHCO3, 1 mM NaH2PO4, and 11 mM glucose) containing 0.1% albumin, cut into small pieces, and homogenized (in a 1:10 dilution). The creatine phosphokinase activity was determined with a use of a diagnostic kit (Procedure No. 47 UV; Sigma Chemical Co.) with a light spectrophotometer (UV 1240; Shimazu). Values were expressed in units of a released enzyme per gram of muscle tissue per hour (U/g/h), where one unit corresponds to the amount of enzyme that catalyzes the transformation of 1 μmol of substrate at 25°C. 17
Functional assessment
Motor function was evaluated with use of the sciatic functional index (SFI), based on a protocol described by Inserra et al.
18
This is an indirect method to access reinnervation of hindlimb muscles after sciatic nerve transection. Two, 4, and 6 weeks after surgery, the animals' pawprints were registered, and two measurements were taken: (i) the print length (PL), corresponding to the distance from the heel to the third toe, and (ii) the toe spread (TS), corresponding to the distance from the first to fifth toe. Both measurements were taken from injured (E, from experimental) as well as noninjured (N, from normal) sides and the SFI was calculated according to the following equation:
The SFI oscillates around zero for normal nerve function, when it is equal to −100 represents total loss of function of that nerve.
Statistical analysis
All analyses were performed using GraphPad Prism 5.01 (GraphPad Software, Inc.). Unpaired statistical analyses were performed by using the Student's t-test for comparisons between two groups, or by the One-Way analysis of variance for comparisons between three groups, followed by the Tukey post-hoc test when necessary. Data are represented as mean values±standard error. A p-value of 0.05 was considered to indicate statistical significance.
Results
MSC therapy enhances the total number of myelinated fibers and results in greater organization of the regenerating nerve
Six weeks after sciatic nerve transection, we analyzed the morphology of the regenerating nerve in the middle segment. The semithin cross sections revealed that the two groups displayed different patterns of organization. The DMEM group had more connective tissue between the bundles of regenerating clusters and dispersed groups of fibers (Fig. 1A), while the group that received MSCs treatment (Fig. 1B) showed larger number of myelinated fibers. Quantitatively, the MSCs treatment resulted in a significant increase in the total number of myelinated fibers (885.5±75.0, p<0.05) compared to the DMEM group that had no cell grafts (592.6±50.5) (Fig. 1C). At the ultrastructural level, the DMEM group displayed fewer myelinated fibers with evidence of damaged axoplasm and irregular myelin sheaths. The perineurium could not be clearly delimitated (Fig. 1D). The regenerated nerve showed better tissue organization in the MSC group: well-organized bundles, regenerating clusters filled with preserved nerve fibers surrounded by layers of compacted perineurium (Fig. 1E).
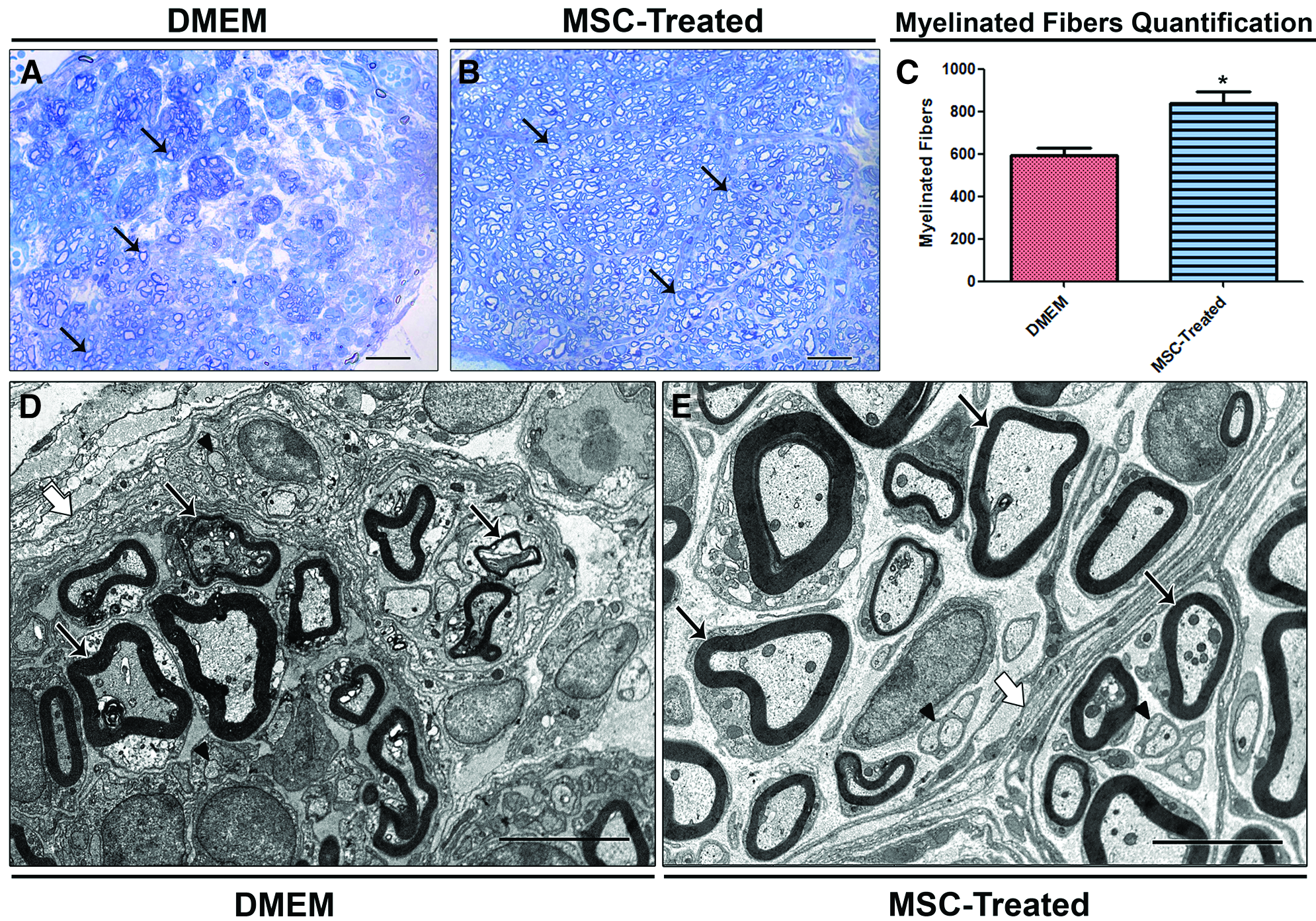
Morphological analysis and fiber quantification in the middle segment of regenerated nerve, 6 weeks after transection.
MSC treatment influences on DRG neurons survival
Six weeks after nerve transection and cell grafting, we evaluated neuron survival by counting the cell nucleoli in the L4 DRG and the L4 spinal cord segment (Fig. 2). The DMEM group of sensory DRG neurons (Fig. 2A, B) were fewer significantly (21.5±3.2, p<0.05) than the MSC-treated group (33±3.4) (Fig. 2C), indicating a pronounced preservation of sensory neurons after MSC treatment. Counts of motor neurons in the anterior horn of the L4 spinal cord segments (Fig. 2D, E) revealed a slight, but statistically insignificant increase in the total number of motor neurons in the MSC-treated group (11.4±1.15; 10±1.6) (Fig. 2F).
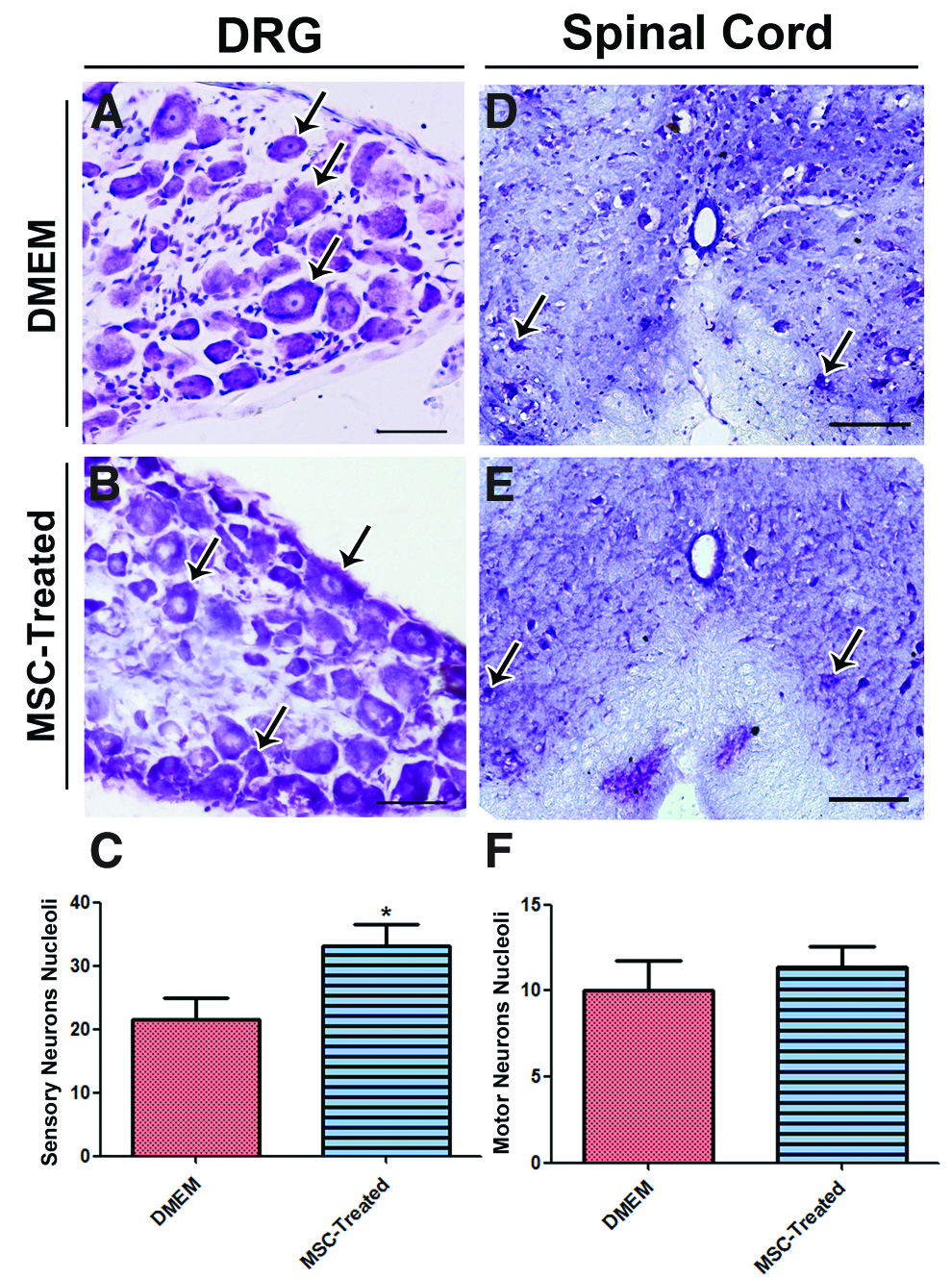
Quantification of neurons nucleoli in dorsal root ganglion (DRG) and spinal cord. Longitudinal sections through DRG
MSC express S-100 protein
To evaluate whether MSCs survived and were still present inside the transplanted tube, and whether they were able to differentiate into Schwann cells 6 weeks after grafting, the distal segment of the regenerated sciatic nerve was stained with an antibody against S-100, a Schwann cell marker (Fig. 3). GFP+ cells were readily visible in the analyzed area, showing that grafted cells had survived and remained inside the PCL tube (Fig. 3A, A′). Some of these GFP+ MSCs also expressed S-100 (Fig. 3B, B′, C, C′), suggesting that they had assumed a Schwann-cell-like phenotype.
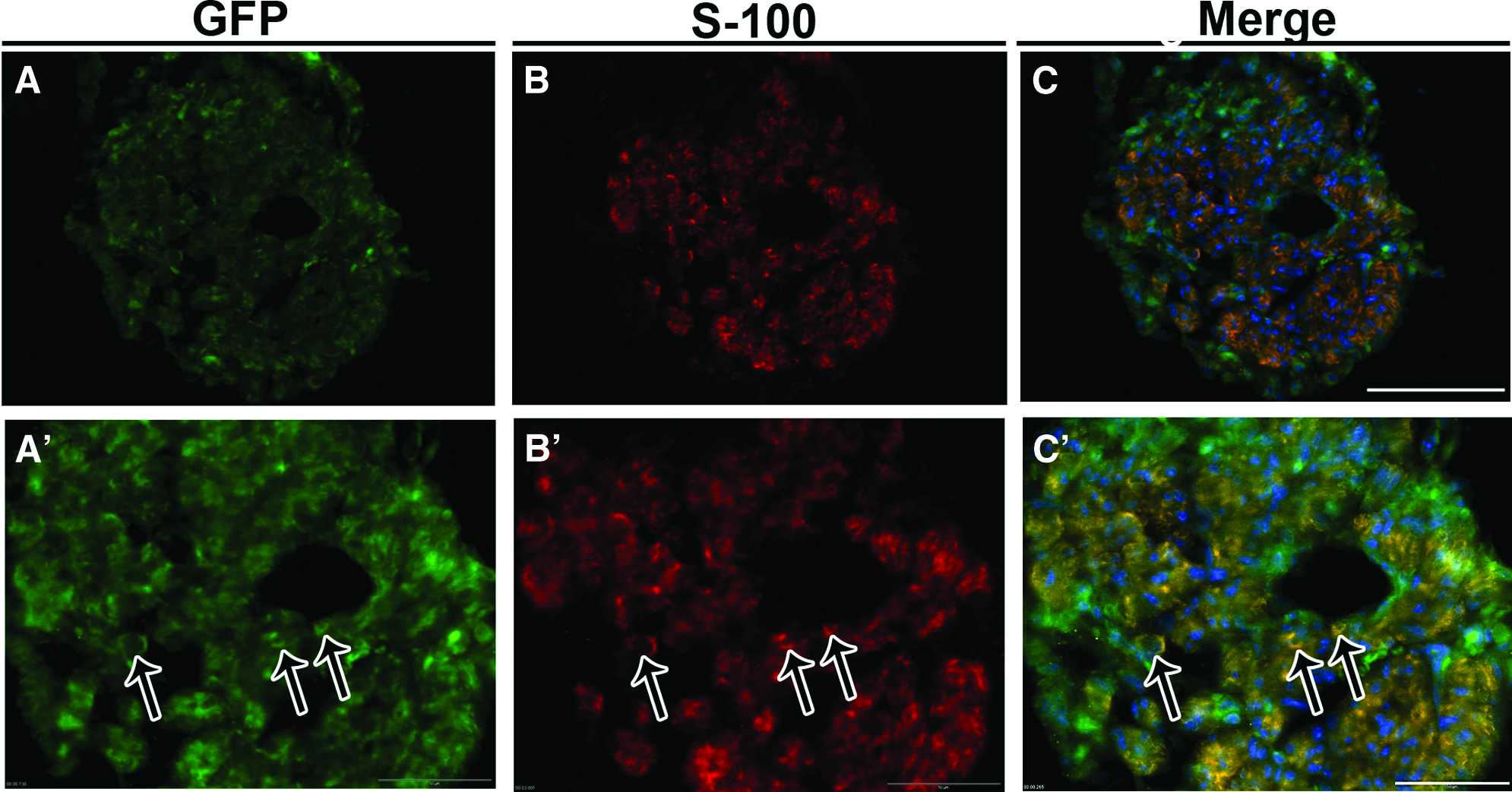
MSC grafting and S-100 protein expression. Confocal images of cross section through the distal segment of regenerating MSC-treated nerve, 6 weeks after surgery.
Cell therapy increased the levels of trophic factors in the tissue
We investigated the amount of trophic factors (BDNF, NGF, NT-3, and NT-4) in the spinal cord, DRG, and sciatic nerve (Fig. 4). The trophic-factor staining showed low immunoreactivity, because they spread very fast and they are soluble in the tissue. However, we could observe a difference in the staining pattern among the groups. In relation to the spinal cord ventral horn (Fig. 4A, D, G, J), there was a significant difference in the fluorescence intensity between the groups only for NGF (Fig. 4D). The immunoreactivity for the other trophic factors was similar. The DRG (Fig. 4B, E, H, K) and the sciatic nerve (Fig. 4C, F, I, L) of the MSC-treated group presented significantly higher levels of all analyzed trophic factors in relation to the DMEM group.
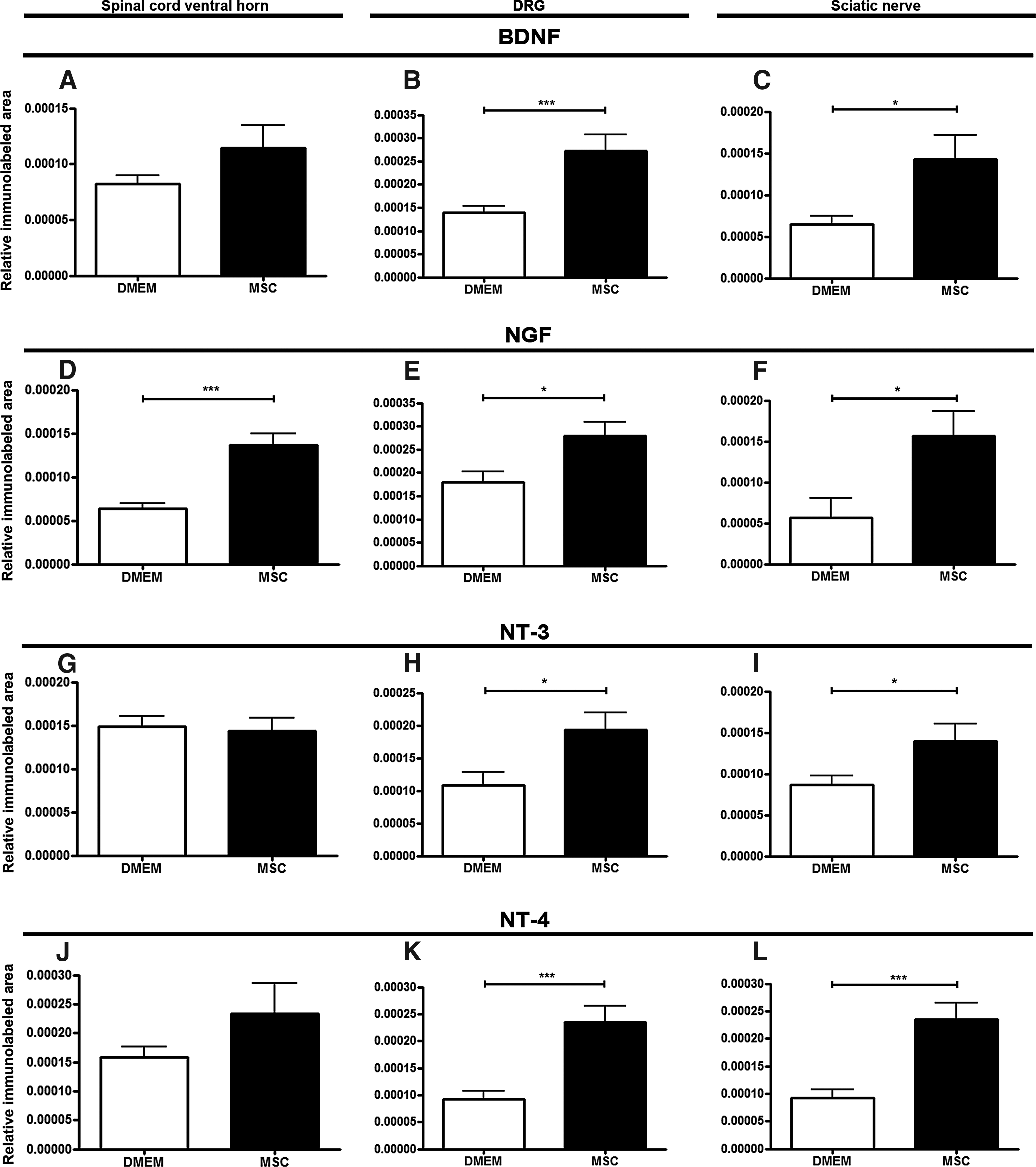
Quantification of trophic factors immunolabeled area.
Cell therapy enhanced reinervation of the gastrocnemius muscle (weight and creatine phosphokinase levels analysis)
To assess muscle innervation and the muscle activity, the gastrocnemius muscle was weighted and its creatine phosphokinase levels were measured 6 weeks after sciatic nerve transection (Fig. 5). Despite the fact that MSC-treated as well as DMEM groups had lost weight when compared to the uninjured side (0.108±0.002), muscle atrophy was more evident in the gastrocnemius of the DMEM mice (0.052±0.004, p<0.05), with a statistically significant increase in the weight of the muscle from MSC-treated mice (0.072±0.005) (Fig. 5A).
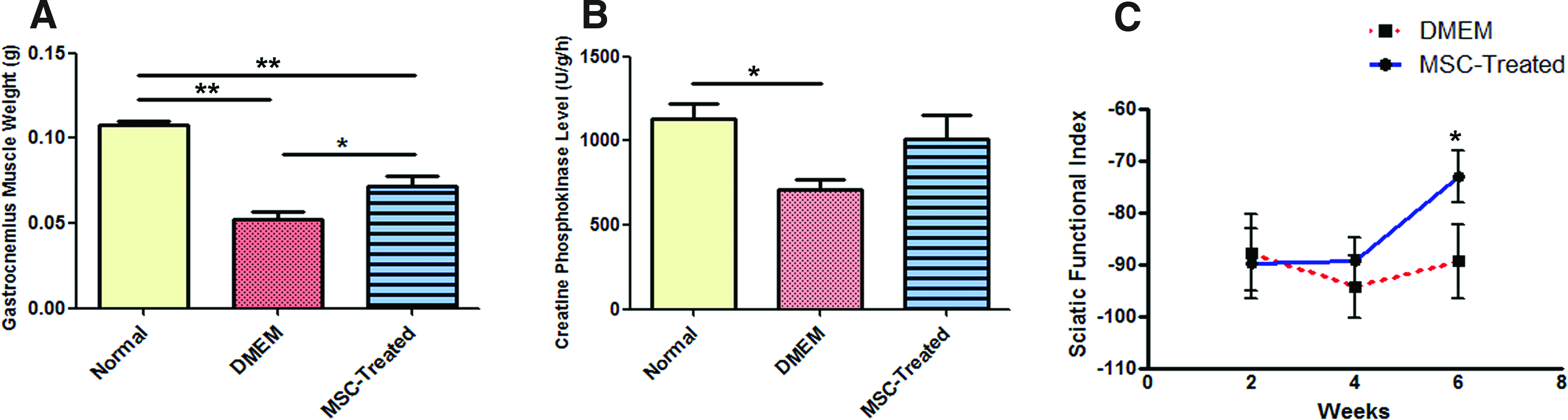
Muscle activity and functional analysis. Gastrocnemius muscle weight
The levels of creatine phosphokinase of both groups were lower than on the normal uninjured side (1130±86.68); however, only the DMEM group showed a significant loss of creatine phosphokinase (708.3±58.39, p<0.03), while the value for the MSC-treated group was closer to the normal, with no significant difference (1013±139.4) (Fig. 5B).
MSC treatment improved functional recovery
To assess the efficacy of the MSC therapy on motor function after nerve transection, animals were evaluated with use of the walking-track analysis 18 2, 4, and 6 weeks after surgery. The SFI was compared between MSC-treated and DMEM groups until the fourth week. Animals that received MSCs maintained the same pattern of function, while the DMEM had a loss of function. After the fourth week, both groups started to show an improvement; the recovery was statistically significant for the MSC-treated group at the week 6 postnerve transection (Fig. 5C).
Discussion
The PNS has the capacity for regeneration after lesions. However, following a severe injury such as full nerve transection, functional recovery is not complete. Such partial recovery leads to limited functional results, with disabilities and decreased quality of life for the affected patients, being the regeneration results disappointing and far from satisfactory. 2 A number of studies have been published, with attempts to improve the regeneration process. Such trials have involved use of Schwann cells transplantation, macrophages recruitment, and delivery of neurotrophic factors and cytokines. 19 Our findings document that bone marrow-derived MSCs inserted into a policaprolactone tube, along with the proximal and distal ends of transected nerve, are able to increase local levels of trophic factors, as well as improve sciatic nerve regeneration, as evidenced by an increase in myelinated fibers number, enhanced sensory neurons survival, and better gastrocnemius and motor function recovery.
Recent studies reported the beneficial effects of bone marrow stromal cells in nerve regeneration.3,12,20–22 Advantages of using these cells include ease of derivation (by aspiration from donor), reduced risk of rejection, ability to proliferate in vitro, and their multipotency, which allows them to differentiate into cells of bone, cartilage, and muscle tissues.23–26 Some studies have shown that MSCs are also able to differentiate into neuron-like and Schwann-like cells in vivo and in vitro, as evidenced by their expression of some neural specific markers.20,21,27–29 Thus, these cells hold great promise for therapeutic application. We have similarly demonstrated that implanted MSCs are also able to transdifferentiate into Schwann-like cells, resulting in increased myelination of regrowing fibers.
Currently, the most common method for nerve repair in neurosurgery is use of a nerve autograft, considered to be the gold standard procedure for clinicians. However, some limitations, such as the restricted nerves availability, the necessity of a second surgery for its removal and loss of function, have stimulated additional studies for development of alternative biological tubular prothesis. The PCL conduit has already been shown to be favorable for enhancing nerve regeneration, guiding nerve fiber growth, and allowing the diffusion of nutrients into its lumen, in addition to being biodegradable.6,8,30 Some investigators propose that following substantial loss of nervous tissue, nerve regeneration can be enhanced by filling the guide tube with different substances like collagen fibers, collagen sponge, 31 fibroblasts, neurotrophic factors, and stem cells.
Several authors discuss the use of MSCs inside different types of tubular prothesis to enhance the regeneration of transected sciatic nerves.20,29,32,33 Pereira Lopes et al. 12 showed that bone marrow stromal cells grafted into a collagen tube successfully increase the number of Schwann cells and myelinated fibers following sciatic nerve transection, leading to functional recovery. In accordance with these authors, other researchers attribute improved regeneration to the release of neurotrophic factors by MSCs, including NGF, BDNF, ciliary neurotrophic factor (CNTF) and glial cell line-derived neurotrophic factor (GDNF), CNTF, and NT-3. They also claim that these cells cause an increase in levels of TrkA and the growth-associated protein 43 (GAP 43).19,21,22,33 TrkA, TrkB, TrkC (tyrosine kinase receptors), and p75 NTR are the cell surface receptors, which bind to one or more of the neurotrophins BDNF, NGF, NT-3, and NT-4. This neurotrophin-receptor complex activates multiple signal transduction pathways that are important to mediate protective effects such as neuron survival and differentiation.34,35
Neurotrophins are known as proteins that regulate neuronal survival, promote axonal and neurite growth, and Schwann cell proliferation.36–38 These molecules induce axonal growth and regeneration, via independent mechanisms, such as by taking part in the polimerization and accumulation of F-actin in the growth cones. As seen here, the MSCs were able to promote an increase in these neurotrophins expression in the site of injury, with an increase in the number of Schwann cells and enhanced axonal growth, thus facilitating the regeneration process.34–38
We also investigated the survival of sensory neurons because these cells are more susceptible to death after a peripheral lesion than motor neurons, which usually survive after peripheral lesions.39–41 According to Jivan et al., 41 16 weeks after transection of the C7 spinal nerve, along with the autograft, 8% of the motor neurons die, while 42% of DRG neurons disappear. The authors report that distal lesions, like sciatic nerve transection, do not lead to significant death in spinal cord neurons, whereas proximal lesions (like avulsions) cause neuronal death in the DRG. They argue that this difference in neuronal responses might be related to the presence of receptors for neurotrophic factors on axotomized neurons. Other reports claim that neurotrophins and their receptors confer a variable vulnerability on neuronal subpopulations, and suggest that these factors could support neuron survival.42–44 These findings may explain the results of our study, where we saw no significant difference between the MSC-treated and DMEM groups in the total number of motor neurons in the ventral spinal cord, while the MSC-treated animals had a pronounced preservation of sensory neurons, with higher levels of the neurotrophins when compared to the untreated group.
Following peripheral nerve lesion, successful regeneration is not related only to axonal growth, but also with the possibility that these axons must find their targets and reinnervate their peripheral targets to re-establish function. Denervated muscle progressively loses weight and atrophies, with a clear decrease in levels of the creatine phosphokinase enzyme.45,46 The creatine phosphokinase enzyme is responsible for the reaction that converts phosphocreatine and adenosine diphosphate to creatine and adenosine triphosphate, 45 and plays a key role in storage and supply of energy in muscle cells. Decreases in this enzyme levels are related to loss of function and activity of the stricken muscle, making the creatine phosphokinase enzyme a relevant indicator of tissue activity. 47 Following sciatic nerve transection, the gastrocnemius muscle is denervated, accompanied by a loss weight and decrease in its levels of creatine phosphokinase. Our data show that treatment with MSCs leads to an increase in muscle weight and its creatine phosphokinase levels, suggesting a recover in the activity and function of the gastrocnemius. In addition to these findings, the motor function analysis, assessed by the SFI, was significantly better in MSC-treated animals compared to animals that did not receive the cells, 6 weeks after transection. Together, these results suggest that the use of MSCs can improve muscle's innervation with a significant restoration of its function.
In conclusion, we have demonstrated that MSCs grafted inside a PCL tube guide have the potential to improve sciatic nerve regeneration. This treatment also prevents the death of sensory neurons, restores gastrocnemius muscle function, and improves motor function in mice. Our study provides important data about the use of stem cell therapy in nervous system lesions, bringing new perspectives for their treatment.
Footnotes
Disclosure Statement
No competing financial interests exist.