
Abstract
Glandular organs feature ramified structures that are important for regulating physiological transport. The aim of this study was to develop a biomaterial-assisted, serum-free culture system to generate branching structures in explants of glandular organs. The fetal mammary gland (MG) was selected as the model organ to study the formation of glandular structure. Among the many biomaterials tested, chitosan demonstrated a superior effect in promoting branch formation in MGs. The morphogenetic effect toward MG branching was chitosan specific and not observed with other analogs with similar chemical compositions or structures. The molecular weight and specific linkages in the chitosan polymer were important parameters in mediating the morphogenetic effect. MG explants from different anatomical locations effectively promoted structure formation. Blocking endogenous fibroblast growth factor 10 (FGF10) inhibited the morphogenetic effect of chitosan, indicating that the chitosan effect was FGF10 dependent. This work demonstrates the feasibility of creating a serum-free system that is competent in facilitating tissue morphogenesis in MG. MG tissue structure can be efficiently generated in a biocompatible system, which was assisted by biomaterials.
Introduction
The mammary gland (MG) is a typical glandular organ featuring a complex branching structure. Without an intact tissue structure in the MG, lactation cannot proceed smoothly, and breastfeeding becomes difficult. Dysfunction or loss of the MG is encountered in many clinical situations such as in congenital anomalies, acquired or traumatic injury, and therapeutic complications.3,4 Until now, no effective treatment has been created to recover MG structure.
The extensive tree-like structure of the MG is generated by branching morphogenesis, a developmental process characterized by repetitive rounds of epithelial branch formation. During the development of the embryonic MG, the mammary placodes are present at embryonic day 10 (E10). 5 Around E13, a spheroid-like mammary epithelial bud is formed adjacent to the epidermis. Along the anteroposterior axis of the body, five pairs of mammary placodes are identified.6–8 In female embryos, the placodes have a developmental quiescent period before E16.9,10 After that, the epithelial bud starts to proliferate and sprouts out before giving rise to the rudimentary branching structure of the MG.6–9 The MG develops in a monopodial pattern and generates a wide-spread ductal system.6,11 When the highly ramified ductal system has been established, alveoli are differentiated all over the branching ducts and become the sites of milk synthesis.12,13
The tissue structure of the MG is developed by branching morphogenesis. Therefore, to engineer the organ, it is preferable to recapitulate the process of structure formation. For decades, several groups have constructed systems for MG structure formation.6–8,10 Many of these systems use a serum-containing medium to isolate and culture MG explants. In the system established in Kratochwil and Schwartz, horse serum and chick embryo extract are fundamental components of the culture medium.6–8 In the study by Hens et al., fetal bovine serum is used for mammary bud culture of harvested embryos. 14 Although the use of serum-free culture had been attempted for the morphogenesis of MGs, serum-containing solutions are still required for harvesting mammary cells.15–17 Additionally, Matrigel, a matrix isolated from mouse sarcoma cells, is used to support the mammary epithelium in serum-free systems.15–17 Although these systems have shown success in facilitating the structure formation of cultured MGs, the added ingredients and matrix render these systems only suitable for laboratory investigation.
There is great interest in creating a biocompatible environment for tissue regenerative purpose. Previously, we and other groups have demonstrated the feasibility of using a biomaterial to assist tissue structure formation.18–21 Many components of extracellular matrices such as collagen and laminin are widely used as biocompatible scaffolds for facilitating tissue structure formation.22–24 This is an appealing approach to regenerate tissue, because it facilitates tissue morphogenesis and structural formation without the addition of an exogenous supplement of serum or bioincompatible extracts. In the current study, this concept was applied to the MG, an important glandular organ. The feasibility of creating a biocompatible environment to promote MG structure formation was investigated. By defining the culture environment, a serum-free biomaterial system capable of facilitating MG structure formation in a biocompatible manner was explored.
Materials and Methods
MG explants
Tissue explants were retrieved from murine embryonic MGs of Institute for Cancer Research strains at the indicated gestational ages. The gestation age was defined in days with the plug detection being defined as day 0 (E0). 6 Animal protocols were approved by the National Taiwan University Animal Care and Use Committee. MG explants were dissected by the methodology described previously.6,8,14,25 In the retrieval of MG explants, small pieces of the adjacent mesenchyme and skin usually accompanied the epithelial MG explants. 6 The assays in this study were performed with female embryos only to prevent the possibility of sex dimorphism. 6 MG explants were placed onto a Nuclepore filter (Whatman, Cat Mo. 110405; VWR) suspended in an air–liquid interface on the cultured medium. 26 The serum-free medium is made of the Dulbecco's modified Eagle's medium: Nutrient Mixture F-12 (Gibco) medium supplemented with 150 μg/mL vitamin C, 100 U/mL penicillin, and 100 μg/mL streptomycin.19,27 Vitamin C is beneficial to branching morphogenesis of ex vivo culture of glandular organs and was supplemented in the system.2,28 The explants were cultured at 37°C in a humidified chamber with 5% CO2 and 95% air atmosphere.
Evaluation of MG structure formation
Cultured MG explants were evaluated at the indicated time points. The assays in which MG explants were not specifically designated utilized the explants harvested from the first pair of MGs for comparison. The branching numbers of cultured explants were measured by counting the epithelial branches independently by different individuals who were blinded to the experimental conditions. The counting results were averaged and presented as the change in the ratio. The ratio is obtained by dividing the number of branches measured after 48 h in culture by that measured at 0 h. The values were determined after at least three-time repeats, and at least five explants were used for each experiment. The values were tested for statistical significance using the t-test, paired t-test, and analysis of variance. It has been reported that during MG development, the branching pattern varies between individuals. 29 Therefore, branching patterns and related parameters were not included for comparison in the current study.
Preparation of chitosan-containing culture system
Preparation of a chitosan-containing culture system generally followed the described methodology. 30 Chitosan (Sigma) was dissolved in 1 M acetic acid. It was mixed with a serum-free medium and neutralized with sodium hydroxide for a final concentration of 0.3 mg/mL. A mock medium was prepared in the same manner as the chitosan-containing medium without adding chitosan. In our preliminary data and in a previous study, 21 it was shown that the aforementioned serum-free medium and mock medium had similar morphogenetic effects. They were used as the control in the following assays.
Preparation of biomaterials and chitosan related substrates
Type I collagen (Cat. 354242, rat tail) and laminin were obtained from BD Biosciences. Glycosaminoglycans, including heparan sulfate (H7640) and chondroitin sulfate (C9819), were purchased from Sigma. Poly-
Immunofluorescence
For immunofluorescent staining, MG explants were fixed with ice-cold 4% paraformaldehyde in phosphate-buffered saline (PBS), and thoroughly washed. Then, the fixed MG explants were pretreated with 0.3% Triton-X-100 and 10% bovine serum albumin in PBS at 37°C for 2 h, and then incubated with selected primary antibodies. Because the tissue explants were used for staining, a detergent with a higher concentration was used for permeabilization.31,32 Fibroblast growth factor receptor 2b (FGFR2b, 1:100; R&D Systems) monoclonal antibody was used for detecting the receptor expression in cultured explants. Primary antibodies were visualized by fluorescence-conjugated secondary antibodies (1:100; Chemicon). The fluorescence was photographed by using a confocal microscope (Leica SP-5).
Antisense oligodeoxynucleotide assay
Antisense experiments were performed with oligodeoxynucleotides (ODNs) with a phosphorothioate modification. The nucleotide sequences used in this study were the following: fibroblast growth factor 10 (FGF10) antisense (351–366), 5′-AGTATCCATTTCCACAT-3′, and scrambled sequence, 5′-GGCAAATTTGGAAATTG-3′. 33 Scrambled and antisense ODNs (2 μM) were added to the system in the beginning of culture. MG explants were cultured with indicated ODNs for 48 h and photographed. To determine if gene expression of FGF10 was suppressed, reverse transcription polymerase chain reaction was performed with the following primers. FGF10: forward, 5′-GGATACTGACACATTGTGCCTCAG-3′ and reverse, 5′-TGTTTTTTGTCCTCTCCTGGGAG-3′. Glyceraldehyde 3-phosphate dehydrogenase was used as a control and detected using the following primers: forward, 5′-GGAAGGTGAAGGTCGGAGTC-3′ and reverse, 5′-CAGTAGAGGCAGGGATGATG-3′. Total RNA was extracted from cultured explants with TRI reagent (Sigma) and was reverse transcribed for cDNA synthesis (Fermentas). The results were detected and analyzed by an imaging system (Alpha Innotech).
Results
Branching structure formation of MGs in vivo
To demonstrate in vivo structure formation during MG development, MG primordiums were isolated at the indicated stages from E13 to E17. Consistent with previous studies, a resting period of branching development was noted in MGs before E16.9,10 In E13, E14, and E15 stages, MG rudiments appeared without showing any branches. In the following developmental stages, branching morphogenesis was initiated in MGs. Two branches were noted in E16 MG rudiments while more branches were formed at E17 (Fig. 1). This indicates that the structure formation of MG was active in the stages after the resting period.
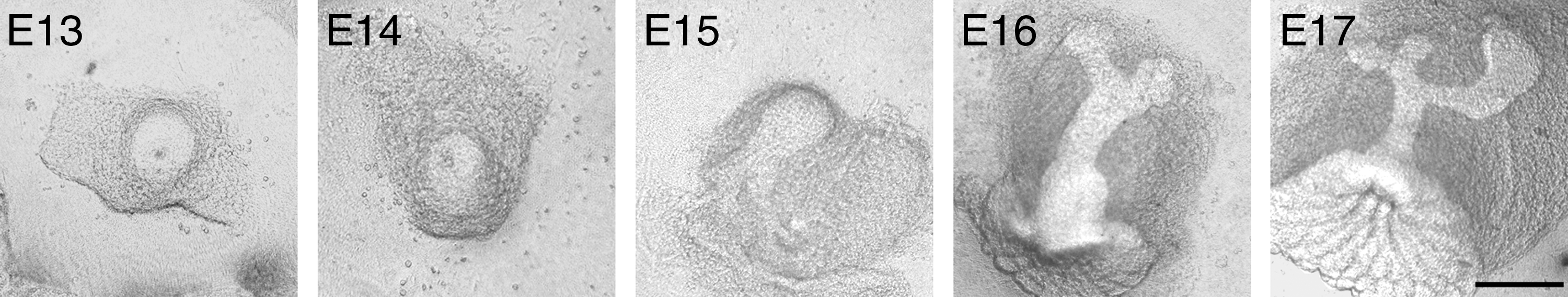
Morphogenesis of MG in vivo. The MG primordium has different morphologies, depending on the stage of development in vivo (scale bars=100 μm). MG, mammary gland.
The effect of serum in the branching structure formation of MGs in vitro
To verify whether MG structures formed in culture could mimic in vivo development, an established system widely used for culturing MGs and other glandular organs was used.8,34,35 The components of the system are shown in Figure 2A. Isolated MG explants were placed in a filter and cultured at the air–medium interface. The air–liquid interface is beneficial for explants to proceed branching morphogenesis and generate glandular structures by optimizing gas supply and surface tension.36,37 Because of the MG morphogenetic resting period during in vivo development, MG explants harvested at E15 were used for the following assays. When the culture was prepared with serum, cultured MG explants successfully initiated the branching process, and after 48-h culture, had sprouted many branches. When a serum-free medium was used, MG explants sprouted two branches in the first 24 h of culture. In the following culture period, no additional branches were found (Fig. 2B). Quantitative analysis demonstrated that the number of MG branches was different between serum-containing and serum-free conditions (Fig. 2C). In agreement with previous studies,6–8 the results show the importance of serum in the in vitro culture system for MG structure formation.
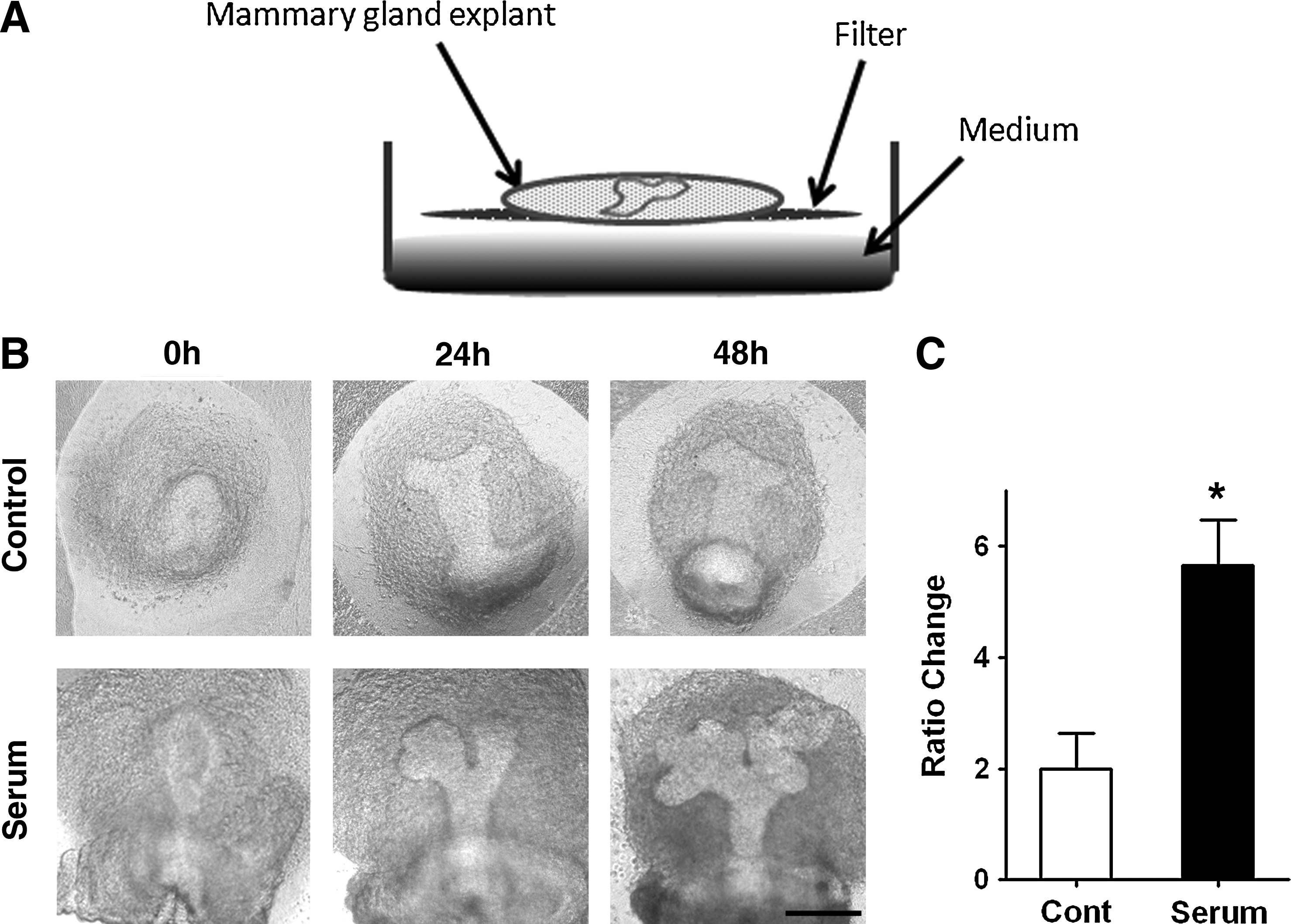
Serum effect in the structure formation of MG in vitro.
Screening of biomaterials to facilitate structure formation of MG explants
Although serum was essential in the culture system of MGs, it might not be suitable for clinical tissue regeneration. For regenerative purpose, a serum-free system might be preferable. Therefore, a serum-free system composed of biomaterials was explored. In this study, to increase the interaction between biomaterials and culture MG explants, biomaterials were prepared in soluble forms. Figure 3A illustrates the serum-free system that was prepared by mixing soluble biomaterials with the medium used for MG culture. To determine which biomaterials were capable of facilitating structure formation of cultured MG explants, biomaterials that have been previously demonstrated to be able to regulate tissue morphogenesis were tested. These were type I collagen, laminin, and glycosaminoglycans, which are natural substrates well known for their roles in branch formation during organogenesis.2,38 We also tested agarose, which has been used to engineer substrates for mammalian cell patterning, 39 and polylysine, a polypeptide commonly used to enhance cell adhesion. 40 After 48 h of culture in a serum-free culture medium, some morphogenetic-promoting effects on MG explants could be observed in the presence of type I collagen and laminin, but not in other substrates (Fig. 3B). However, in the chitosan-containing system, MG explants developed significantly more branches than in the control (Fig. 3B). Quantitatively, chitosan had the strongest effect on promoting MG structure formation (Fig. 3C).
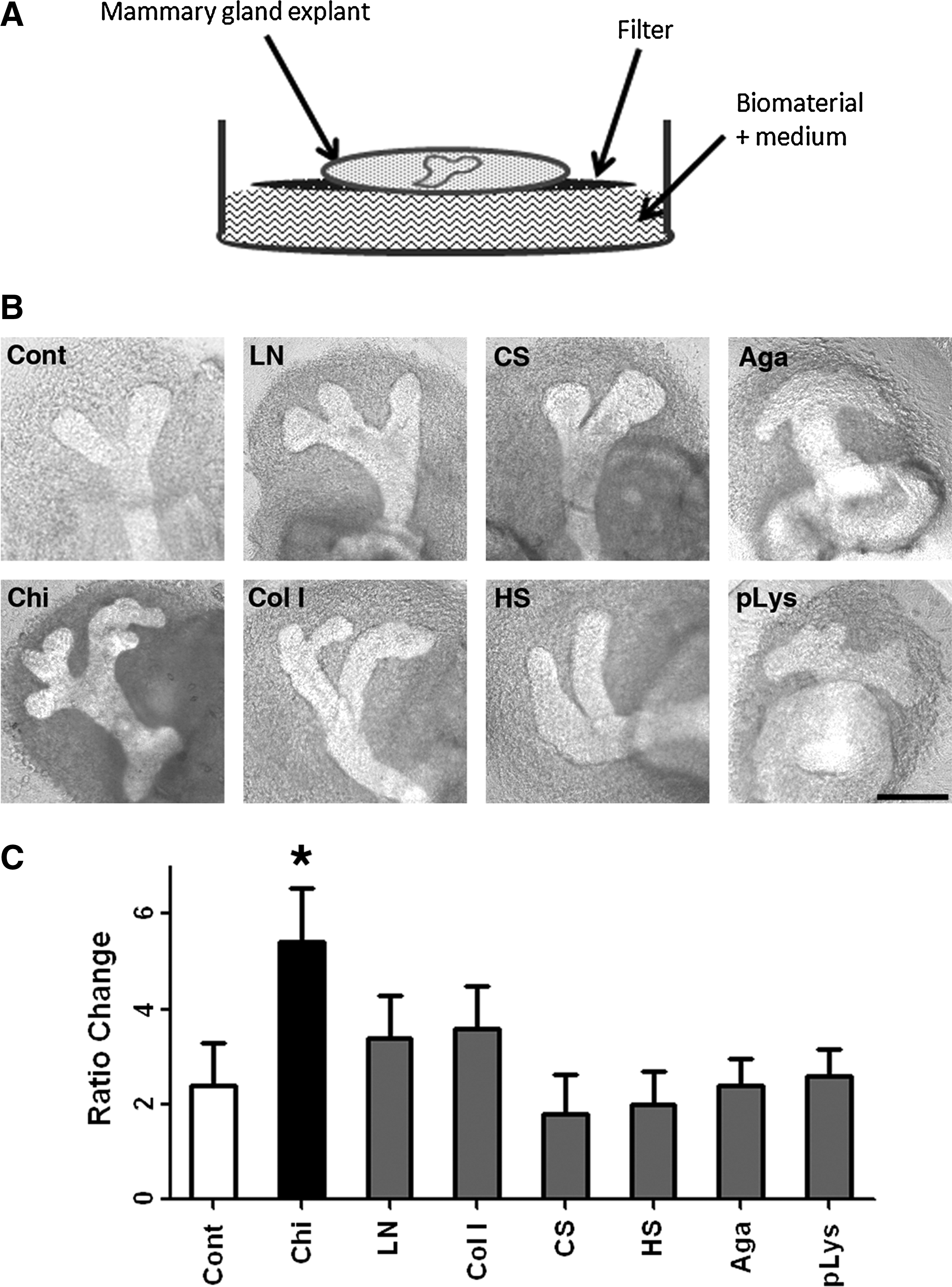
The biomaterial-assisted serum-free system utilized to regulate structure formation of MG explants.
Chemical properties of chitosan in regulating MG structure formation
To elucidate the chemical properties of chitosan that was responsible for promoting MG structure formation, chitosan-related analogs and chemicals were tested. Because chitosan is composed of a combination of GlcNAc and GlcN, these two chemical units were prepared separately. No branching-promoting effects were observed in explants grown with these constituents in the culture medium (Fig. 4A). To determine whether the effect of chitosan was caused by chemicals with similar structures, analogs of chitosan such as GalNAc, GalN, ManNAc, and ManN were tested in the culture system. Our results show that these chemicals, prepared under the same conditions, did not exhibit any branch-promoting effect on cultured MG explants (Fig. 4B). These results suggest that the branch-promoting effect on MG was specific to chitosan rather a common phenomenon mediated by similar chemicals.
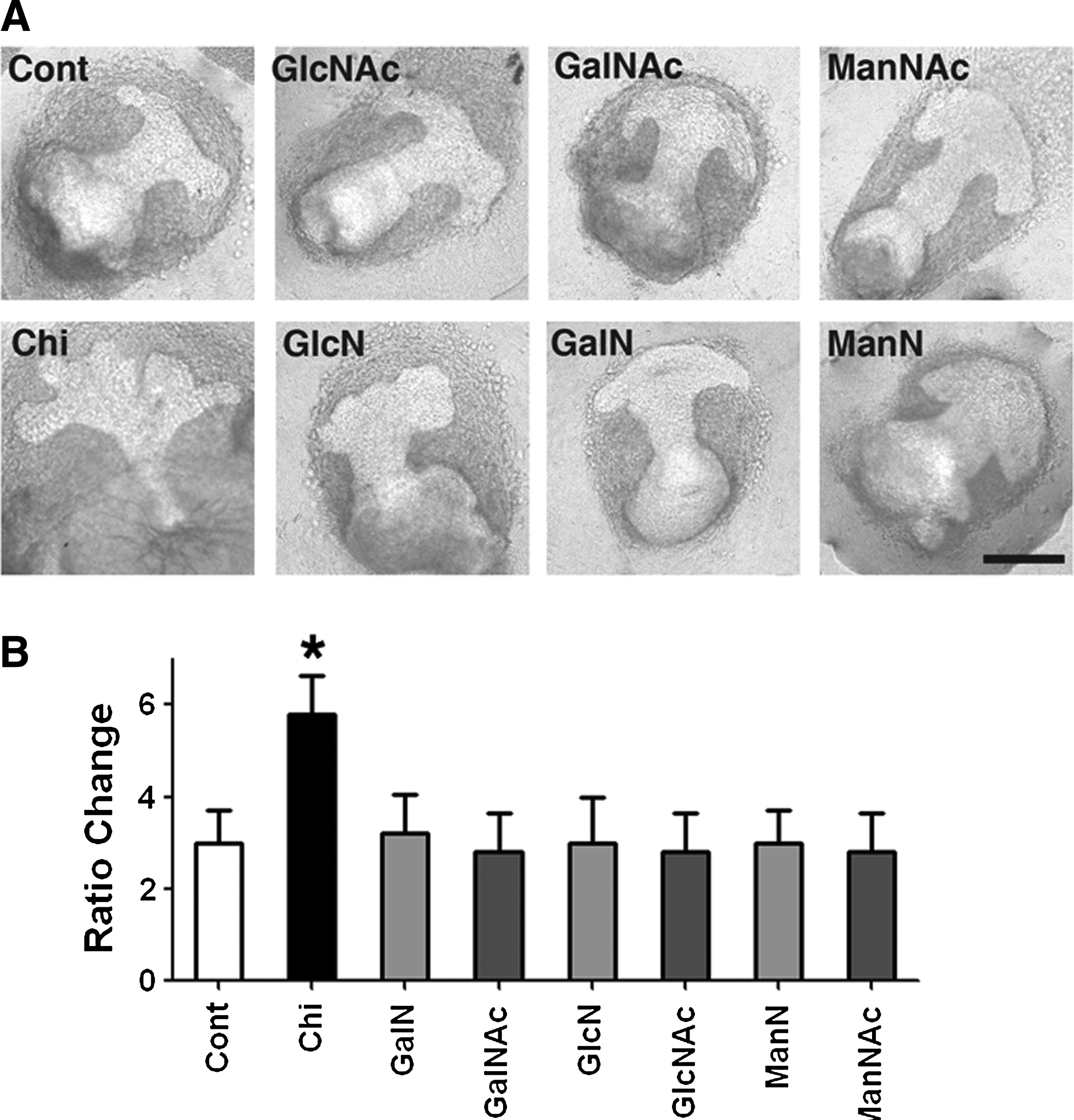
Chitosan-related chemicals in regulating MG structure formation.
The molecular weight of chitosan was demonstrated to be an important parameter in regulating its biological effects.41,42 When chitosan with molecular weights of 810 or 612 kDa was used, cultured MG explants developed a greater number of branches than when cultured with chitosan with lower molecular weights (Fig. 5A). When MG explants were cultured in the medium prepared from chitosan oligomers with molecular weights<5 kDa, the branch number was similar to that of the control (Fig. 5A). These results indicate that the molecular weight of chitosan was critical in mediating the chitosan effect.
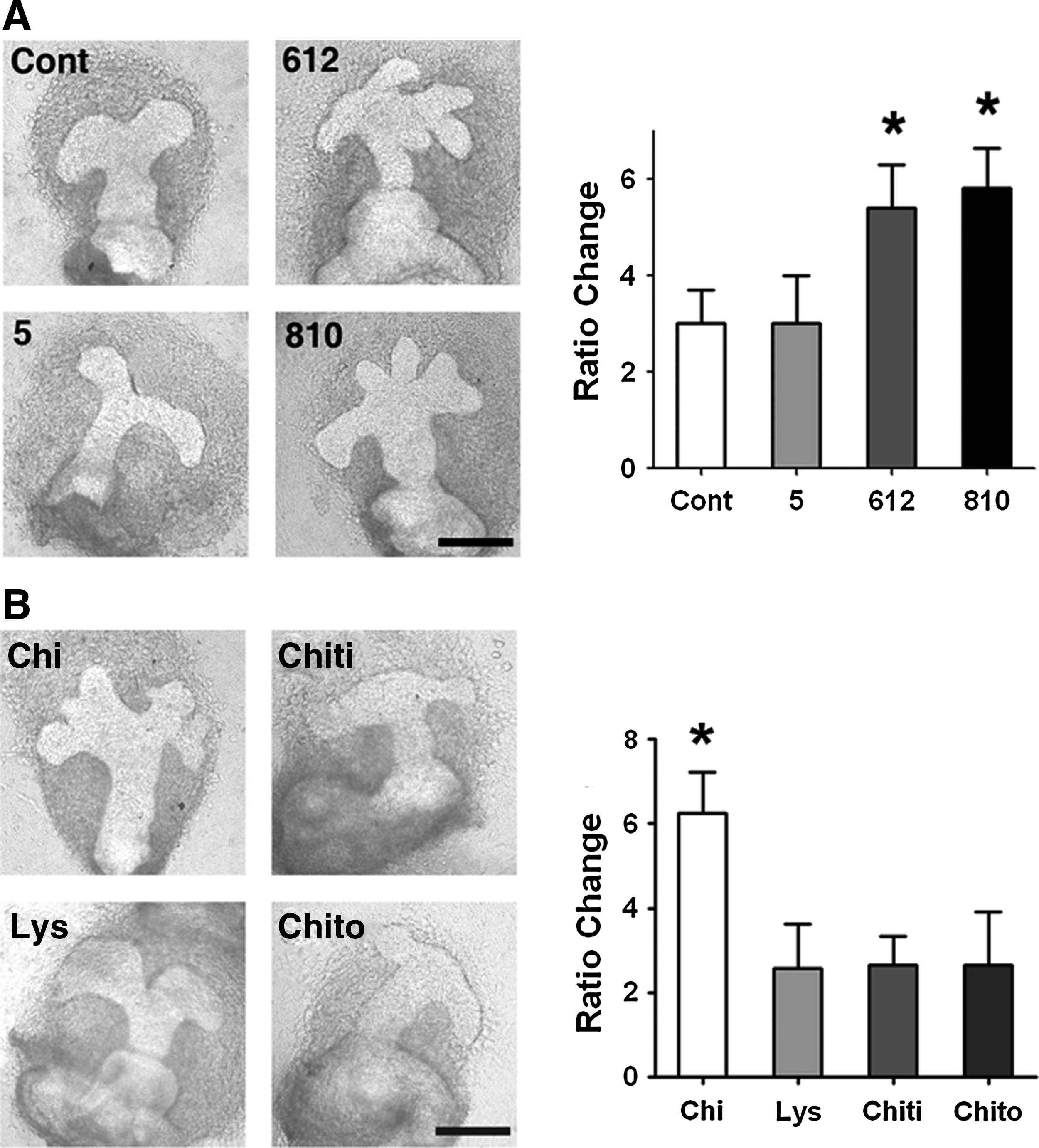
The chemical properties of chitosan required for regulating MG structure formation.
Chitosan is a polymeric biomaterial, and the linkages between individual monomers are important to maintain the chemical content and structure. Similarly, the molecular weight of chitosan is also dependent on polymeric linkages. To test the importance of linkages, we used different enzymes capable of cleaving the specific linkages within chitosan polymers. Lysozyme, which hydrolyzes GluNAc linkages, diminished the MG branch-promoting effect of chitosan (Fig. 5B). Similar effects were observed with chitinase, another enzyme capable of cleaving GluNAc linkages (Fig. 5B). Chitosan also contains another linkage between GluNAc and GluN, which can be digested by chitosanase. No branch-promoting effect on MGs was observed when chitosanase-digested chitosan was used (Fig. 3C). These data show the importance of GluNAc linkage and GluNAc-GluN linkages in chitosan for promoting MG structure formation.
Anatomical factor in chitosan mediated structure formation of MG
During MG development, five different pairs of MGs are generated in distinct anatomical locations. It has been reported that considerable variation existed in the degree of development among MGs from different anatomical positions.5,8 To further verify whether the activities of MG structure formation promoted by chitosan could be affected by an anatomical factor, the cultured explants harvested from different pairs of MG were evaluated in a time-lapse manner. The representative figures showing the cervical pair of MG explants are shown in Figure 6A. After 24 h of culture, the differences in the branching number in cultured MG explants were observed (Fig. 6A). In the following 24 h, branching activity seemed to be suspended, although MG explants continued to grow. In contrast, MG explants in a chitosan-containing system showed successive branching, and the branched number exceeded the control (Fig. 6B). In addition to the cervical MG pair, there are four other MG pairs. MG explants harvested from the different anatomical locations were examined in the chitosan-containing system. Among the different MG pairs, variation existed in the branch number of explants. However, those cultured in the chitosan-containing system developed more branches than the controls grown in the absence of chitosan (Fig. 6C, D). This result indicates that chitosan had a general effect on the structure formation of MGs.

Anatomical factors in chitosan-mediated structure formation of MG.
FGF10 signaling in chitosan-mediated structural formation of MGs
During MG development, the FGF10-FGFR2b signaling pathway plays a key role in inducing branching morphogenesis of placodes. 5 FGF10 had been implicated in instructing epithelial–mesenchymal interactions required for the formation of epithelial branches. When mice are deficient in FGF10, normal MG development is not observed. 5 Therefore, it might be worthy to verify whether FGF10 also plays a role in regulating MG structure formation in the current system. As shown in the MG explants cultured for 1 day, FGFR2b was expressed in the newly developed epithelial branches (Fig. 7A). It implied a probability that FGF10 might be essential in mediating MG structure formation. When FGF10-blocking ODNs were applied, no structure formation could be initiated in the control group. Similar effects were found in the chitosan-containing group. However, scrambled ODNs did not affect the control or chitosan-containing system (Fig. 7B). When the branching number was compared between the control treated with FGF10 antisense ODNs and the chitosan-containing system treated with FGF10 antisense ODNs, no differences were found (Fig. 7D). The results indicate that the promoting effect of chitosan diminished when FGF10 was blocked in cultured MG explants.
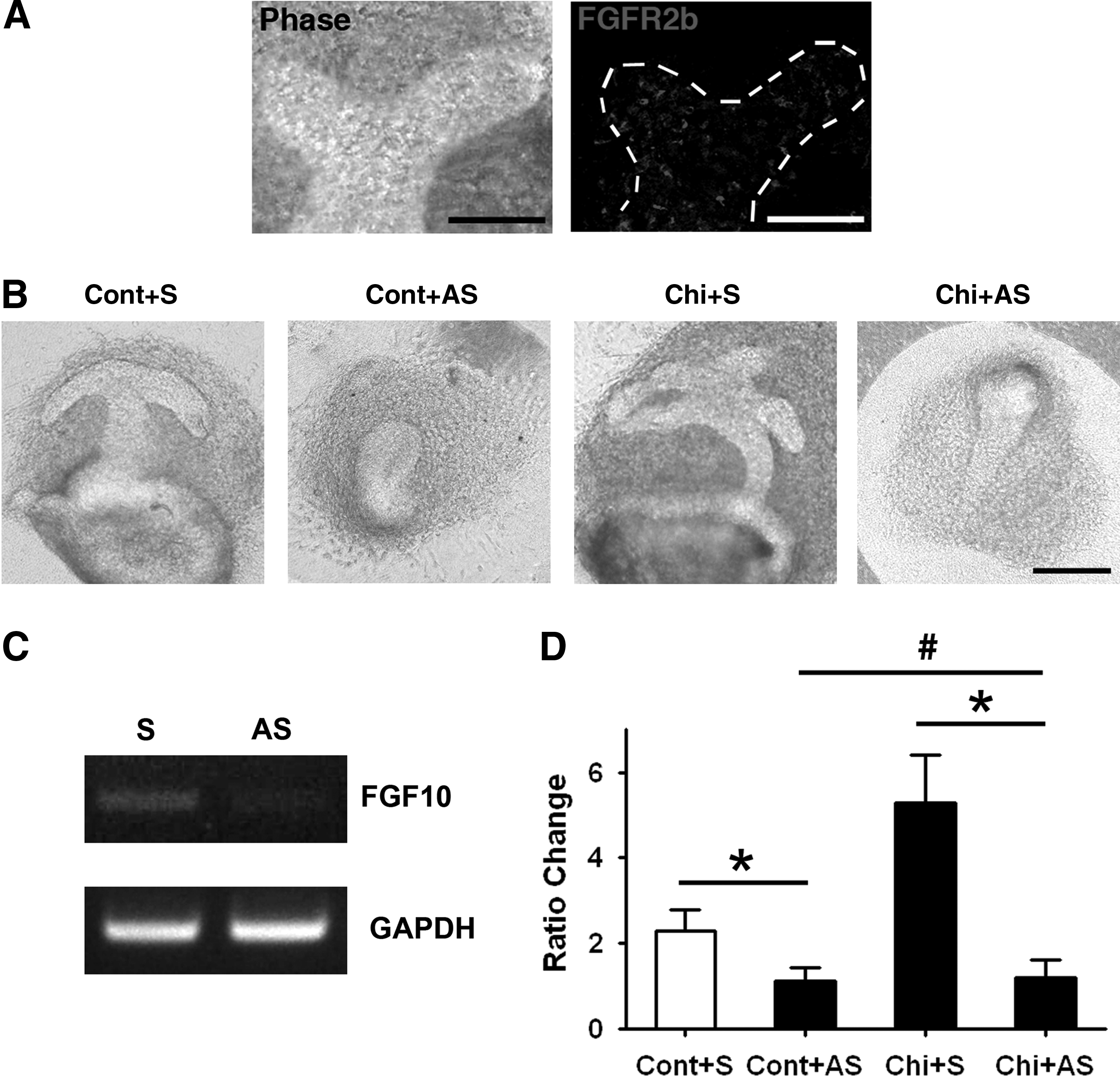
MG structure formation in chitosan-containing system by FGF10 blocking.
Discussion
In the current study, we demonstrated the possibility of creating a biomaterial-assisted system to facilitate the structure formation of MGs. With chitosan, the ramified structure could be generated efficiently. In this system, the structure formation of MG derived from different anatomical locations could be promoted. By coordinating with endogenous tissue-specific morphogens, MG structure formation can be generated in vitro without serum supplement.
Structure formation is essential for many different glandular organs. By forming ramified structures, different cells can be well organized, and the efficacy of physiological transport as well as the cell surface created for physiological demands can increase significantly. Therefore, many glandular organs are generated by branching morphogenesis during organogenesis. Because of these physiological advantages, creating a ramified structure is beneficial to regenerate a functional glandular organ. Although it has been shown that progenitor mammary cells are capable of tissue regeneration after being transplanted into healthy recipients,43,44 diseases or therapeutic interventions may make it difficult to find a competent recipient environment for reconstructing the mammary structure. Although constructing an artificial structure with biomaterial scaffolds may be a straightforward approach, until now, it is still difficult to fabricate an exquisite network structure within a limited space.45,46 In contrast, by facilitating the cell interactions with materials that contribute to structure formation, the architecture can be established by the cultured tissue or cells. Furthermore, cells can be organized and incorporated into the tissue structure simultaneously, which is beneficial for the tissue to be functional.
In many previous attempts to generate MG tissue structure in vitro, serum is an essential component.6–8,16,47 It is well known that many morphogens are involved in the developmental program of establishing the tissue structure of glandular organs.48,49 Thus, to recapitulate the process of structure formation, supplementation of essential morphogens is imperative. In many studies that establish MG structure in vitro, the addition of exogenous serum or other morphogen-abundant substrates had been widely used.6–8,10,14–17 Serum is competent in providing many essential growth factors and morphogens for in vitro cultured tissue and cells. However, it is a complex mixture with a variety of undefined components with undetermined concentrations. These properties render the effect of serum in biological responses complicated and difficult to clarify. Additionally, the potential for serum-transmitted diseases and inconsistent quality from different batches are major concerns in engineering tissue for clinical applications. To this end, creation of a serum-free system is preferable. In addition to biocompatibility issues, a serum-free system provides the possibility to clarify biological reactions and tissue interactions specifically, making it possible to fine-tune and optimize the system for tissue regeneration.
In a serum-free system, maintaining and supporting the essential factors for tissue structure formation is imperative. It is appealing to create an environment that is bioactive in both mediating cell interaction and tissue response. 21 Substrates capable of regulating or enhancing endogenous morphogens are of interest when designing biomaterial approaches. In the current study, we screened various biomaterials and found that chitosan promoted tissue structure formation of MGs. Some morphogenetic-promoting effects on MG explants could be observed in the presence of type I collagen and laminin. Collagen and laminin are essential extracellular matrices for branching morphogenesis, and are reasonable to contribute to structural formation of MG explants.38,50 Chitosan is a biocompatible polymer derived from deacetylated chitin. 51 It has been extensively used in biomedical applications because of its ability to form a polyelectrolyte complex with many bioactive factors.41,52–54 The polycationic nature of chitosan enables it to interact with many anionic molecules that are important in mediating cell and tissue responses.55,56 Because of these features, chitosan is competent in immobilizing and increasing the local concentration of morphogenetic factors, which allow morphogens to act on cells precisely and contribute to tissuegenesis. Similar morphogenetic effects have been shown in other ramified tissues,19,30 and might also account for chitosan's promoting effect on MG structure formation in this study. Together with the findings of other ramified tissue, these results demonstrate the general potential of chitosan in mediating structure formation of branching organs.
Chitosan-related biological responses are largely dependent on its molecular weight.41,42 High-molecular-weight chitosan absorbs peptide and loads protein more easily.57,58 In our previous studies, high-molecular-weight chitosan had a greater branch-promoting effect on salivary glands. 30 Similarly, a successful branching-promoting effect was induced in MGs by chitosan with a higher, but not a lower, molecular weight. In addition, the branch-promoting effects of chitosan diminish when digested with enzymes. It had been reported that the ability of chitosan to form a polyelectrolyte complex is affected by molecular weight. 59 Enzymatic digestion produces lower-molecular-weight forms of chitosan, and therefore may not provide a sufficient molecular composition to generate the polyelectrolyte complex that in turn affects the chitosan-mediated morphogenetic effects. Therefore, to create a system supported by chitosan, the molecular weight is a critical parameter that needs to be precisely determined.
FGF10 is essential in guiding tissue structure formation during MG development. 5 FGF10 plays a principal role in defining the ectodermal cells destined to form MG and acts as a dominant chemoattractant that regulates MG epithelial developmental behaviors. In the current study, the morphogenetic effect of chitosan diminished when FGF10 was blocked. At E15 stages of which the explants are harvested, FGF10 expression is observed in the mammary fat pad precursor surrounding the mammary epithelial bud. 5 Because the current culture system is free from serum or other exogenous growth factors, the source of FGF10 is presumably from MG explants. Chitosan can interact with different members of the FGF family, such as FGF10, to promote biological responses.21,60,61 It is likely that MG explants produce FGF10, and FGF10 morphogenetic effect is concentrated and enhanced by the chemical interaction with chitosan in an appropriate spatiotemporal manner for branching morphogenesis. This effect suggests the possibility that the underlying mechanism behind chitosan's effect is due to its interaction with FGF10.
During MG development, there is a diverse potential for branching activities in MG explants harvested from different anatomical locations.8,62 At the molecular level, FGF10 is only involved in the initial structure formation of MGs originating from all pairs, except the fourth one, which is asynchronous in MG structure formation. 5 Intriguingly, in the current system, all MG pairs could be induced to form tissue structure. This suggested the possibility that chitosan may be effective in coordinating and enhancing the effect of other morphogens implicated in MG morphogenesis rather than FGF10. Accordingly, we previously demonstrated that chitosan is able to cooperate with different morphogens to enhance their biological efficacy. 21 This feature is beneficial for tissue regeneration, because a diversity of morphogen requirements may exist. Therefore, there is potential for chitosan-containing systems to be utilized for structure formation of different glandular organs.
Conclusion
This study demonstrates the feasibility of using a biomaterial approach to facilitate structure formation of glandular organs without serum or exogenous morphogens. The current system is competent in efficiently promoting structure formation of MG. By coordinating with endogenous morphogenetic factors, tissue structure formation can be recapitulated in a biocompatible system using chitosan.
Footnotes
Acknowledgments
The authors thank the National Science Council of the Republic of China, National Taiwan University Hospital, and National Health Research Institutes for their financial support of this research. The authors also thank the staff of the Eighth Core Lab, Department of Medical Research, National Taiwan University Hospital, for technical support during the study.
Disclosure Statement
No competing financial interests exist.