
Abstract
Human neural stem/progenitor cells (hNSCs/NPCs) are a promising cell source for neural tissue engineering because of their ability to differentiate into various neural lineages. In this study, hNSC/NPC differentiation was evaluated on piezoelectric, fibrous scaffolds. These smart materials have an intrinsic material property where transient electric potential can be generated in the material upon minute mechanical deformation. hNSCs/NPCs cultured on the scaffolds and films differentiated into β-III tubulin-positive cells, a neuronal cell marker, with or without the presence of inductive factors. In contrast, hNSCs/NPCs cultured on laminin-coated plates were predominantly nestin positive, a NSC marker, in the control medium. Gene expression results suggest that the scaffolds may have promoted the formation of mature neural cells exhibiting neuron-like characteristics. hNSCs/NPCs differentiated mostly into β-III tubulin-positive cells and had the greatest average neurite length on micron-sized, annealed (more piezoelectric), aligned scaffolds, demonstrating their potential for neural tissue-engineering applications.
Introduction
Neural stem/progenitor cells (NSCs/NPCs) are a relatively undifferentiated population of cells that have the ability to differentiate into a broad range of specialized neurons and glia in the central nervous system (CNS). 4 NSCs/NPCs from the subventricular zone of the lateral ventricles and the subgranular cell layer of the hippocampal dentate gyrus of rodents and primates have been well characterized. 4 Therapies have used NSCs/NPCs for neural disorders such as Huntington's and Parkinson's disease, 5 amyotrophic lateral sclerosis,6,7 multiple sclerosis, 8 acute brain injury, 9 SCI,10–12 and brain tumors. 13 NSCs transplanted into rats with SCI have been shown to repair damaged tissues and improve functional recovery.11,12 Grafted NSCs/NPCs remyelinated the damaged tissue and traveled along the spinal cord. 2 Glial scar formation, lack of neurotrophin factors, and the presence of various inhibitory molecules that inhibit axonal regeneration may also prevent endogenous or transplanted NSCs/NPCs to function. 14 Hence, stem cell transplants in the CNS have been proposed for replacing and remyelinating the damaged tissues, providing neuroprotection or creating a conducive environment by secreting neurotrophic factors for endogenous cells to regenerate.11,12 One of the main challenges for stem cell-based therapies is cell survival and directing cell differentiation. 15 Co-grafting NSCs/NPCs with other cell types, growth factors, or other therapeutic agents have gained interest and are beginning to show promise in enhancing NSC/NPC survival and neural differentiation in the spinal cord.16,17 Alternative strategies being explored are in the use of engineered three-dimensional scaffolds that use appropriate biomaterials that can influence neural stem cell proliferation and differentiation.18,19
Local electric fields have been measured during neural development or after nerve injury in numerous vertebrate systems. 20 Bioelectrical control mechanisms may be especially relevant for the nervous system, since developing and regenerating nerve fibers occur via interaction with the extracellular environment before reaching their target structures. 21 During tissue development or regeneration, the fields created by electrically charged extracellular matrix materials may provide signals for outgrowth or remodeling. 22 Electric fields have been shown to influence growth and orientation of neurons in vitro 23 where the electric field was generated via electrodes. Piezoelectric polymers can induce a transient change of surface charge without requiring additional energy sources or electrodes and have been shown to yield a higher level of neuronal differentiation and neurite outgrowth in mouse neuroblastoma cells as compared to nonpiezoelectric controls. 22
In this study, the differentiation of human NSCs/NPCs (hNSCs/NPCs) on fibrous piezoelectric scaffolds was studied using immunocytochemistry and quantitative real-time (RT)–polymerase chain reaction (PCR). Fibers were made of polyvinylidene fluoride trifluoroethylene (PVDF-TrFE), which has well-known piezoelectric characteristics and proven biocompatibility.22,24–26 In addition, to our knowledge, this is the first study characterizing hNSCs/NPCs on fibrous scaffolds, wherein the physical features of the fiber, specifically the fiber diameter and orientation, varied. Aligned fibrous scaffolds have been shown to direct neurite outgrowth and enhance neurite length for neuronal cell types when compared to random fibrous scaffolds.27–29 PVDF-TrFE films were also used to evaluate the influence of the polymer chemistry versus the fiber structure in neural differentiation. We hypothesized that piezoelectric and aligned fibrous scaffolds would stimulate hNSC/NPC differentiation and neurite extension.
Materials and Methods
Scaffold and film fabrication
Electrospinning was conducted using a typical setup as previously described. 30 About 15% and 25% (w/v) of poly(vinylidene fluoride trifluoroethylene) (65/35) (PVDF-TrFE; Solvay Solexis, Inc.) in methyl ethyl ketone (MEK; Mallinckrodt Baker) were used for electrospinning to fabricate nano- and micron-sized fibers, respectively. Random and aligned fibers were collected on a grounded metal plate or a fast-rotating drum, respectively. The scaffolds were annealed to enhance piezoelectric characteristics 25 where the electrospun scaffolds were kept at 135°C for 96 h and quenched with ice water. Untreated electrospun scaffolds were denoted as as-spun scaffolds. All samples were kept in vacuum for at least 48 h before use for cell culture. For preparing the films, 25% (w/v) PVDF-TrFE solution was deposited on a metal sheet and drawn with a draw bar to create the films. The films were air-dried for 2 days prior removing from the metal plate and were further vacuum-dried for 2 days before usage. Annealed films were processed in the same manner as described for the fibrous scaffolds.
Scaffold characterization
Film and scaffold morphology was evaluated using a scanning electron microscope (SEM; LEO 1530 Gemini). The samples were mounted onto the SEM stubs and were coated with gold using sputter coating (Bal-Tec 020 HR Sputtering Coater) at 40 μA for 20 s.
The Instron 3342 Single Column System was used to determine the Young's modulus (or stiffness) and maximum tensile stress of all scaffold groups. The scaffolds were cut into 70-×10-mm strips with a testing area of 40×10 mm. Ten samples were tested for each scaffold group using a displacement rate of 10 mm/min. 31 The thickness of each sample was calculated as the average of the thickness of the two ends and the middle section of each strip. 32 All scaffolds were sterilized with ethanol and rinsed with phosphate-buffered saline (PBS) before the mechanical testing.
Surface chemistry was examined by Fourier transform infrared (FTIR) spectroscopy using the Perkin Elmer FTIR-ATR 100 series for unprocessed powder, films, and electrospun. All samples were scanned in the range of 1600–400 cm−1 with a resolution of 4 cm−1.
The presence of a Curie transform, which is unique to the piezoelectric β-phase crystals in PVDF-TrFE, was evaluated with thermally stimulating depolarization current (TSDC). The dipole in the β-phase crystal rotates into a nonpiezoelectric crystal conformation when the Curie temperature is reached. TSDC measures the solid phase–phase transition as a depolarization current peak. The PVDF-TrFE electrospun mats or powder were sandwiched between two pieces of Teflon film and heated from −60°C to 140°C in the absence of an externally applied field with a heating ramp of 7°C/min using TherMold TSC 9000 TSC/RMA (TherMold). The crystallinity of the scaffolds was determined using differential scanning calorimetry (DSC, TA Q100). The samples were heated from −60°C to 200°C and cooled to −60°C and then heated again to 200°C with a 7°C per minute temperature change. The crystallinity of samples was calculated using the following relation:
where Hfs is the average of the calculated heat of fusion at melt (n=3), and Hft is the heat of fusion for 100% crystallinity of the polymer (45 J/g 33 ).
The porosity of the random scaffolds was estimated by the density difference between the scaffold and the unprocessed material. The thickness of a 6-mm-diameter scaffold disk was measured and weighed to determine the density of the scaffold (ρscaffold, n=5). The porosity was calculated as follows 34 :
where ρraw is 1.78 g/cm3. The mean pore diameter was determined by the capillary flow analysis (1100 AEX Capillary Flow Porometer; Porous Materials, Inc., Analytical Services Division).
hNSC/NPC culture on scaffolds
The experimental groups for the cell studies were cells cultured on (1) 25% PVDF-TrFE film, (2) nanosized random, (3) nanosized aligned, (4) micron-sized random, and (5) micron-sized aligned scaffolds. The scaffolds and films were punched into 6-mm disks using a biopsy punch and were sterilized with ethanol followed by several PBS washes. The scaffolds and films were preconditioned in a DMEM (Invitrogen, Inc.) with 10% fetal bovine serum (Thermal Scientific Hyclone) for 1 day before seeding to improve cell attachment. They were rinsed with PBS several times immediately before seeding. Laminin-coated plates served as a control in this study. About 100 μL of laminin (6.5 μg/mL, Cultrex® Mouse Laminin I; R&D Systems, Inc.) was added to each well of a 96-well plate and incubated at 37°C overnight. Laminin-coated wells were then rinsed with hNSC/NPC-culturing media (described in the next section) once immediately before seeding.
Poietics Normal Human Neural Progenitors (hNSCs/NPCs), isolated from 16–20-week fetal whole brain, and all components in the media used in this study were purchased from Lonza, Inc. One day before seeding, the cells were thawed and incubated in a neural progenitor maintenance medium (NPMM) for 24 h. The NPMM contained an NPC basal medium, Neural Survival Factor-1 (NSF-1), gentamicin and amphorericin B, 3 μg/mL human epidermal growth factor, and 1 μg/mL human fibroblast growth factor. The neural progenitor differentiation medium (NPDM) contained an NPC basal medium, NSF-1, gentamicin and amphorericin B, and 25 ng/mL brain-derived neurotrophic factor.
The scaffolds and films were seeded with a neurosphere suspension (15,000 cells/cm2) and were then kept in a humidified 37°C incubator with 5% CO2 where the medium was changed every 3 to 4 days. The cells were kept in either a control (NPMM) or induction (NPDM) medium for 9 days.
RNA isolation and gene expression
Homogenization and DNA digestion of the lysate for P0 neurospheres and cells on scaffolds and films at days 1 and 9 in culture were performed by QIAshredder (Qiagen, Inc.) and RNase-Free DNase Set (Qiagen), respectively. Total RNA was isolated using the RNeasy Micro Kit (Qiagen) following the manufacturer's instructions. The One Stem QuantiTech SYBR Green RT-PCR Kit (Qiagen) and the MX4000 detection system (Stratagene, Inc.) were used to conduct quantitative RT-PCR analysis. The reverse transcription cycle was performed at 50°C for 30 min followed by PCR activation for 15 min at 95°C. A three-step amplification cycle consisted of 15 s of denaturation at 94°C, 30 s of annealing at 55°C, and 30 s of extension at 72°C, repeated for 40 cycles. A melting curve analysis was included for each reaction. The analysis was performed in triplicate, and the values were normalized to the relative amount of the housekeeping gene RPLPO (ribosomal protein, large, PO). 35 Gene expression for β-III tubulin, glial fibrillary acid protein (GFAP), and RPLPO (housekeeping gene) was evaluated. The logarithmic value of the ratio of mRNA/RPLPO expression at days 9 to 1 (log D9/D1) was calculated.
Immunofluorescent staining and analysis
At day 9, the cells were fixed with 4% paraformaldehyde for 20 min and were blocked with PBS containing 0.1% Triton X-100, 10% normal donkey serum, and 1% bovine serum albumin (BSA) for 45 min at room temperature. They were incubated with a primary antibody cocktail containing 1:25 mouse anti-neuron-specific β-III tubulin (100 μg/mL; R&D Systems), 1:350 rabbit anti-human nestin (1 mg/mL; Abcam, Inc.), and 1:25 sheep anti-human GFAP (100 μg/mL; R&D Systems) overnight at 4°C. After rinsing, the samples were incubated with a secondary antibody cocktail containing 1:200 of NorthernLights™ anti-mouse IgG-NL557 (Lonza), donkey anti-rabbit IgG-alexa flour® 488 (Invitrogen), and donkey anti-sheep IgG-alexa flour 405 (Invitrogen) for an hour at room temperature. They were rinsed with 0.1% BSA twice for 10 min each and stored in PBS until needed. Between each step, the samples were rinsed with 0.1% BSA for 10 min. The samples were counterstained with the ProLong® Gold antifade reagent with DAPI (Invitrogen) to stain the nuclei and prevent photo bleaching just before imaging. Fluorescent images of the samples were taken with a confocal fluorescent microscope (C1si). Series of images were taken with depth ∼60–80 μm using the z-stack function. The images were then volume rendered to create a two-dimensional image for analysis.
Confocal images taken at 40×were used to calculate the percentage of positively stained cells. Four images were taken randomly per scaffold, and eight scaffolds were used for each scaffold type in each medium. A total of 32 images were used to calculate the percentage of positively stained cells and to measure the neurite length for each group. The total number of cells (DAPI) were counted manually using ImageJ (1.42q). The cells were considered positive for nestin, β-III tubulin, or GFAP by stain and corresponding morphology. The fraction of positively stained cells per image was calculated as the number of positively stained cells divided by the total number of cells. The average of the fraction of positively stained cells was then calculated for each group. Only processes that were immnunopositive for β-III tubulin were considered as neurites for measurement. Neurite extension was measured as the distance between the tip of the neurite and the point entering the cell body.
Statistical analysis
SPSS (SPSS v.16.0; IBM) was used for statistical evaluation. All data were analyzed using a one-way analysis of variance with the Tukey's test for multicomparisons. General linear regression was performed to determine the effect of fiber diameter, alignment, stiffness, crystallinity, and piezoelectricity on neurite extension and the percentage of nestin+, β-III tubulin+, or GFAP+ cells. Probability (p) values<0.05 were considered statistically significant differences.
Results
Electrospun scaffold and film characterization
The PVDF-TrFE-annealed film had a rough morphology as compared to the as-cast film (Fig. 1). Annealing did not alter the fiber morphology or diameter significantly (Fig 1). Average fiber diameters of nano- and micron-sized scaffolds were ∼750 nm and 3.3 μm, respectively. The average fiber alignment was ∼90% for all aligned scaffolds. The fibers in the aligned scaffolds were oriented and densely packed; hence, the porosity and pore size were only evaluated on random fibrous scaffolds. All as-spun random mats had an average porosity of 51% and a pore size of 3 μm. Average porosity of annealed scaffolds was 74% and was significantly greater than as-spun random mats. The average pore size of annealed random mats was 4 μm, but not statistically different from as-spun random mats.

SEM images (magnification 1500×, scale bar=20 μm) of electrospun polyvinylidene fluoride trifluoroethylene (PVDF-TrFE) nanosized, as-spun and annealed random
For a similar fiber size, aligned and annealed scaffolds have higher Young's moduli than the random and as-spun scaffolds, respectively (Table 1). Annealed micron-aligned scaffolds had the highest Young's modulus (p<0.05). The annealed scaffolds also had a higher crystallinity than the as-pun scaffolds. All electrospun scaffolds had a depolarization peak in the TSDC spectra at ∼100°C, whereas a current movement in the unprocessed powder TSDC spectra occurred around the melting point. All materials had peaks at 1285 and 1430 cm−1 in the FTIR spectra.
Annealed scaffolds and films had higher crystallinity than the as-spun scaffolds, as-cast film, and unprocessed powder (p<0.05) 1Highest modulus among all scaffolds except for aligned, annealed nanosized PVDF-TrFE (p<0.05). Annealed random2 and aligned1,3 had higher moduli than as-spun random and aligned, respectively (p<0.05). *Aligned as-spun and annealed scaffolds had higher maximum tensile stress than the random scaffolds in their respective groups (p<0.05).**Aligned, annealed micron-sized PVDF-TrFE scaffolds had the highest maximum tensile stress among all groups except for aligned, annealed nanosized PVDF-TrFE scaffolds (p<0.05).
PVDF-TrFE, polyvinylidene fluoride trifluoroethylene.
hNSCs/NPCs on scaffolds and films
hNSCs/NPCs received from Lonza were free-floating neurospheres (Fig. 2A). hNSCs/NPCs seeded on the laminin-coated surface predominantly had a large flat-cell morphology and were immunopositive for the neural stem cell marker nestin (nestin+) in the control media at day 9 (Fig. 2B). In the induction media, cells had small cell bodies with long processes that were mostly nestin+ with a few cell bodies immunopositive for the neuronal marker β-III tubulin (β-III tubulin+) at day 9 (Fig. 2C).
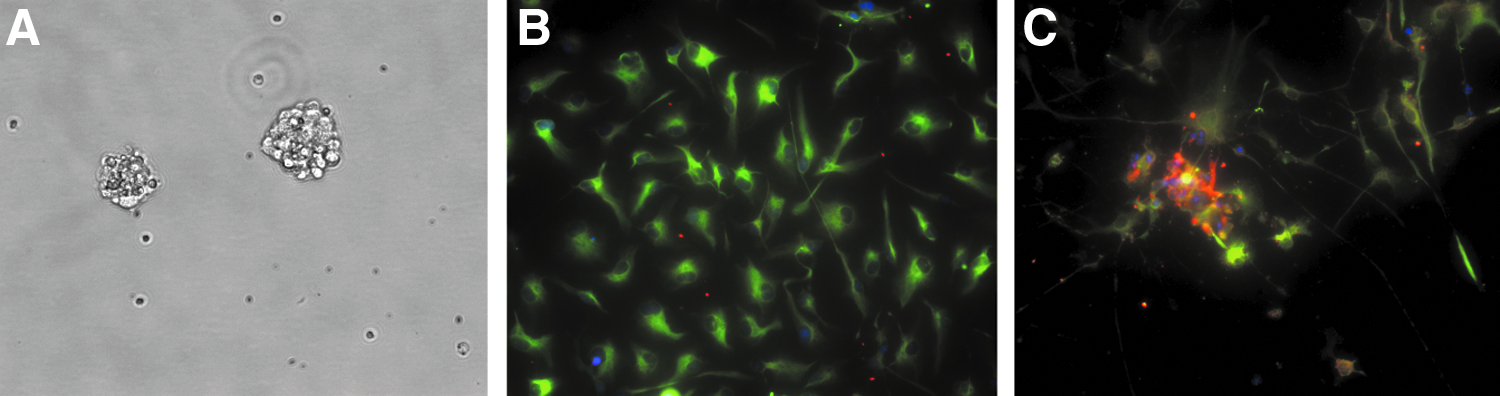
Bright-field image (20×) of a neurosphere suspension in flasks before cell seeding onto scaffolds/laminin surfaces at day 0
Cell bodies appeared smaller on the as-cast and annealed films (Fig. 3) in the control media as compared to the laminin-coated surface (Fig. 2B). Cells were mostly nestin+ on as-cast films (Fig. 3A), but were both nestin+ and β-III tubulin+ on annealed films (Fig. 3B). Fluorescent images of the cells on films in the induction media were absent because no cells were observed after the staining process. A negligible fraction of the immunopositive astrocytic marker GFAP (GFAP+) was observed on both laminin and films (Figs. 2 and 3).
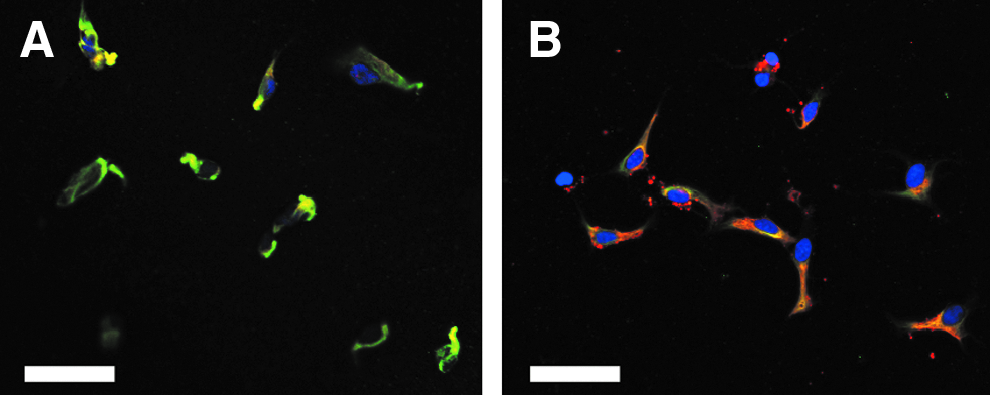
Confocal fluorescent image (40×, scale bar=50 μm) of cells on as-cast
On all electrospun scaffolds, cells were predominantly β-III tubulin+, and a small fraction was nestin+ in both medium conditions and have a negligible fraction of GFAP+ at day 9 (Fig. 4). A linear relationship was determined between piezoelectricity and crystallinity of the fiber with the percentage of β-III tubulin+ cells (p<0.05, Pearson's goodness-of-fit test). Fiber alignment was linearly correlated with the percentage of nestin+ cells (p<0.05, Pearson's goodness-of-fit test), and no linear relationship was observed between the various properties of the fibers and the percentage of GFAP+ cells. Small spheres were observed on all scaffolds in both media, and nestin+ cells were lower in the induction media than the control medium, but were not statistically significant (Fig. 5A). The fraction of β-III tubulin+ cells was lowest on laminin-coated plates in the control medium (Fig. 5B). In the induction medium, annealed nanorandom and micron-aligned scaffolds had more β-III tubulin+ cells than on the laminin-coated surface (p<0.05). Neurites extended along the fibers on the aligned scaffolds in the induction group (Fig. 4). The average neurite length on annealed micron-aligned scaffolds in the induction medium was longer than on as-spun nanoaligned in the control medium and as-spun nano- and micron-random scaffolds in the induction medium (Fig. 6). A linear relationship was determined between fiber alignment, piezoelectricity, stiffness, and crystallinity of the scaffolds with neurite extension (p<0.05, Pearson's goodness-of-fit test).
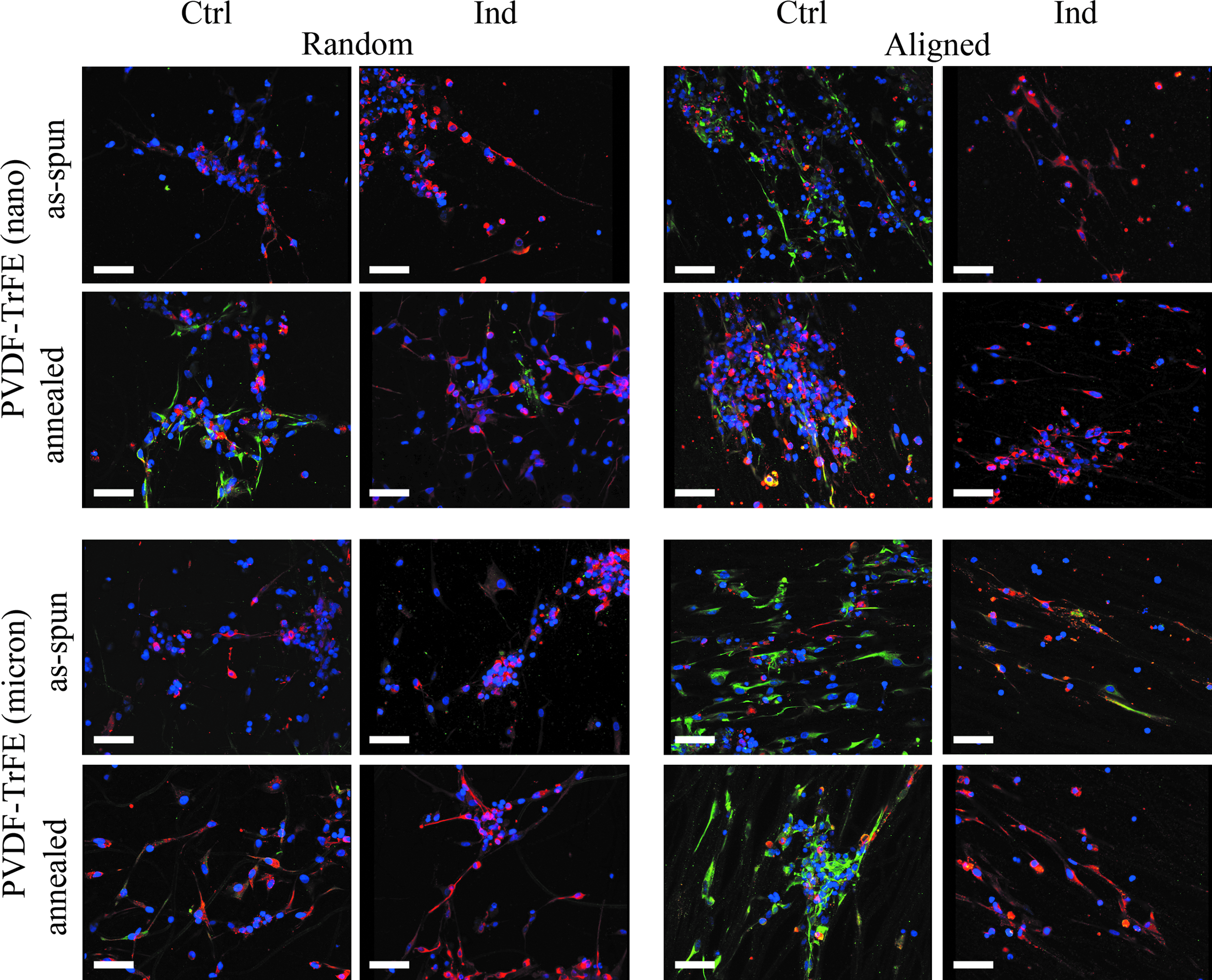
Confocal fluorescent images (40×, scale bar=50 μm) of cells seeded onto electrospun nano- and micron-sized PVDF-TrFE as-spun and annealed, random and aligned scaffolds stained with anti-nestin (green), anti-β-III tubulin (red), anti-GFAP (blue), and counterstained with DAPI (blue) in control and induction media at 9 days in culture. Color images available online at www.liebertpub.com/tea

Fraction of positively stained anti-nestin

Average neurite extension of cells on electrospun PVDF-TrFE nano- and micron-sized, as-spun and annealed, random and aligned in control and induction media at day 9. *Average neurite length on PVDF-TrFE micron-sized annealed, aligned scaffolds in induction media was longer than nanosized as-spun aligned in control media and nano- and micron-sized as-spun, random scaffolds in induction media.
The log D9/D1 mRNA/RPLPO ratio >1 indicated an increase in expression and a ratio <1 indicated a decrease in expression of mRNA from days 1 to 9. Cells expressed less β-III tubulin on most of the scaffolds in both media (Fig. 7A). GFAP mRNA was expressed the highest for cells on annealed micron-random scaffolds in the control medium (Fig. 7B). Lower expression of GFAP mRNA occurred on the micron- and nanoaligned scaffolds in both control and induction media, which corresponded with decreased β-III tubulin mRNA expression. GFAP mRNA expression was higher in the induction than in the control medium on annealed films.

Logarithmic value of the D9/D1 RNA/RPLPO ratio of β-III tubulin
Discussion
This is the first study that evaluated hNSC/NPC differentiation on fibrous scaffolds, specifically electrospun micron- and nanosized fibrous scaffolds that also have piezoelectric characteristics. The piezoelectric characteristics of the films and scaffolds increased from as-cast or as-spun to annealed films and scaffolds. Enhanced piezoelectric characteristics occurred during the annealing processing as shown by the increase in the presence of the piezoelectric β-phase crystal and crystallinity as previously described.25,26 Annealing also resulted in an increase in the scaffold's Young's modulus or stiffness, which is often observed with an increase in crystallinity. The depolarization peak in TSDC confirmed the presence of the β-phase crystals by the dipole movement at the Curie transform. Peaks in the FTIR spectra of all scaffolds also demonstrated the characteristic bands of β-phase crystals. 36 In piezoelectric materials, the accumulation of transient electric charge occurs in response to applied mechanical deformation. In these fibers, piezoelectricity may be induced via minute deformations of the fibers due to cell attachment and migration, which have been shown in other non-neural cell types in vitro. 37 Cells have been shown to contract their matrices up to 1–3 μm. 38 Enhanced piezoelectricity due to annealing may generate more transient electric charge in response to the same mechanical deformation.
The hNSCs/NPCs attached to the films and scaffolds, differentiated, and migrated outward from the spheres, which is typical behavior observed for these cells.39,40 Most cells migrating away from the sphere were β-III tubulin+ in induction media, similar to a previous study. 41 β-III tubulin+ cells with neuron-like morphology in the control media suggested that annealed films induced neuronal differentiation. The annealed films had a rougher surface as compared to as-cast films, which could be contributed to the nonuniform crystal formation during the annealing process, as demonstrated by Rodriguez et al. 42 Cells on as-cast films were β-III tubulin+, but had a different morphology than those cells on laminin in the control media. Although cells on the annealed films did have a neuron-like morphology and were positive for β-III tubulin, the results may also be contributed to the increase in roughness as compared to the as-cast films. Nevertheless, the difference in the cell morphology on the as-cast films as compared to the laminin surfaces suggests an influence of the piezoelectric property on hNSC/NPC morphology and/or differentiation.
The fraction of nestin+ cells was highest on laminin-coated plates in both the control and induction media, suggesting that most cells remained as proliferative hNSCs.43,44 Higher nestin+ cells on the laminin-coated surface suggested that the fibrous structure of the scaffolds may also enhance differentiation. Similar results were observed for rat NSCs stained for nestin on electrospun polyethersulfone (PES) fibers and tissue culture plastic. 45 PES fibers similar in size to the nanosized scaffolds reported here promoted neural differentiation, and no GFAP+ cells were observed. Rat NSCs attached poorly on the 1452-nm PES fiber, and large aggregates were developed. However, in this study, smaller aggregates were found on the nanosized scaffolds where most cells appeared to have migrated outward on the micron-sized scaffolds. Annealed and micron-sized scaffolds had a greater pore size and porosity as compared to as-spun and nanosized scaffolds, respectively, which may allow better cell migration. hNSCs/NPCs that migrated outward from the spheres demonstrated a preference for differentiation into β-III tubulin+ cells with a neuron-like morphology. Stiffness has been previously described to direct nonhuman NSC/NPC differentiation, with neuronal differentiation preferred on softer hydrogel matrices.19,46,47 In this study, the Young's modulus or stiffness of the scaffolds was greatest for all annealed scaffolds, yet its effect on differentiation was unclear. Although stiffness was linearly correlated to neurite extension, the cell response could have been due to an increase in piezoelectricity. Other scaffold features such as fiber orientation or alignment influenced cell and neurite orientation similar to studies using neuronal cell types. 27 The decrease in nestin+ cells with an increase in β-III tubulin+ cells with neuron-like morphology on scaffolds suggests that the scaffolds may have induced hNSC differentiation into neuron cells, and the number of β-III tubulin+ cells on scaffolds was greater than on laminin-coated plates. No differences were detected in the total neurite length between all random and aligned fibrous scaffolds, but neurites were longer on annealed micron-aligned and nanorandom scaffolds. Hippocampal neurons similarly favored micron-scale electrospun poly(lactic acid-co-glycolic acid) fibers for initial attachment and neurite guidance. 27 Fiber morphology may not be the only factor influencing neurite outgrowth but also the piezoelectric property/chemistry of the material, as demonstrated by the cellular response on the films and the linear correlation observed between the properties of the scaffolds and neurite extension.
β-III tubulin is a type of microtubule that is used as a neuronal cell marker in the developing and mature nervous system.48,49 In a study using P19 embryonal carcinoma cells, gene expression of β-III tubulin was detected at day 2, and by day 12, a reduction in gene expression was observed while protein levels remained high. 50 A similar pattern was observed in the rat cerebral cortex, where gene expression decreased with increasing neural maturity. The change in protein levels was insignificant, suggesting that the β-III tubulin proteins were stabilized by a post-translational mechanism. 51 In this study, β-III tubulin gene expression was low, but protein expression was observed on annealed nanorandom and micron-aligned scaffolds, suggesting hNSCs/NPCs may have differentiated into mature neuronal cells. Further investigations are required to identify the neuron population and the function of these cells. The neurotransmitter released or presence of receptors and electrophysiology are needed to be characterized to confirm neural differentiation. On all scaffolds, gene expression for GFAP was detected, but only negligible amounts of GFAP+ cells were observed. A similar phenomenon was observed during rat brain development, where GFAP+ appeared late in gestation, but GFAP gene expression was first detected at low levels at day 15 of the fetus. 52 Hence, a longer culture time may be needed to detect GFAP with immunostaining to correlate with the increase in GFAP gene expression.
A mixed population of cells may have been present in the P0 neurospheres, and all cells may not have differentiated into mature cell types. 53 The presence of both NSC markers and mature cell type markers suggests an asymmetric proliferation producing both mature cell types and stem cells or precursors. Future studies may need to examine proliferation in parallel with differentiation of the hNSCs on the various scaffolds. The inconsistency in gene expression could also be contributed to the mixed population of P0 neurospheres.
Studies of transplanted NSCs/NPCs were mostly derived from the brain and were able to differentiate effectively into astrocytes and oligodendrocytes, but rarely into neurons.54,55 The limitations of NSCs/NPCs transplantation are cell survival and inefficient differentiation into neurons after several passages. 56 It is suggested that an adult SCI environment is not conducive for neural differentiation 56 ; therefore, an NSC/NPC-only transplant may have a limited clinical advantage. An hNSC/NPC- PVDF-TrFE transplant may be of benefit, because this study demonstrated that hNSCs/NPCs predominantly differentiated into cells exhibiting neuron-like characteristics on the piezoelectric scaffolds while maintaining a population of nestin+ cells. The presence of the scaffold may promote differentiation into neurons and also maintain a stem cell population at the SCI site. Although PVDF-TrFE is a nondegradable polymer, tubes and channels made of poly(2-hydroxyethyl methacrylate) (PHEMA) or PHEMA-co-methyl methacrylate,57–62 poly[N-(2-hydroxypropyl)methacrylamide],63–67 and poly(acrylonitrile-co-vinylchloride)68–72 have been evaluated for repairing SCI. The nondegradable property provides structural stability, 73 which may be advantageous for functional repair. In addition, the piezoelectric activity of the material may be induced in vivo by bulk mechanical deformation from the cerebrospinal fluid circulation and in response to body motion.
Conclusions
This study demonstrated that the differentiation of hNSCs/NPCs was enhanced on the annealed piezoelectric PVDF-TrFE scaffolds. The cells differentiated into neurite-bearing β-III tubulin+ cells, and the average neurite extension was highest on annealed micron-aligned scaffolds. This finding suggested that contact guidance combined with piezoelectric properties may promote neurite extension and neuronal differentiation. The results of this study demonstrate the potential use of electrospun PVDF-TrFE for neural tissue-engineering applications.
Footnotes
Acknowledgment
The authors would like to thank the New Jersey Commission on Spinal Cord Research for financial support.
Disclosure Statement
No competing financial interests exist.