
Abstract
Biologic scaffold materials composed of extracellular matrix (ECM) have been shown to promote the formation of site-specific, functional, host tissue following placement in a number of preclinical and clinical studies. Endotoxin contamination of biomaterials is thought to result in deleterious immune responses that may affect the remodeling outcome when present in significant quantities. However, the exact amount of endotoxin contamination within or upon an ECM-based biologic scaffold that is required to elicit adverse effects in recipients is currently unknown. The present study examined the in vitro and in vivo effects of endotoxin contamination within an ECM scaffold derived from porcine dermis upon the host immune response and the downstream ability of the scaffold material to promote constructive tissue remodeling. Test articles with endotoxin values that exceed the current U.S. Food and Drug Administration (FDA) limit had similar or decreased immune responses both in vitro and in vivo when compared with devices that were below the current FDA limit. Dermal matrices spiked with large doses of endotoxin (100 ng/mL), equivalent to 10–20 times the FDA limit, elicited a robust immune response in vitro. However, by 35 days postimplantation, no difference in tissue remodeling was detected, regardless of the amount of endotoxin present within the material. These results suggest that current endotoxin standards may fall well below levels that induce an adverse acute proinflammatory response and associated long-term deleterious effects upon tissue remodeling outcomes.
Introduction
Lipopolysaccharide (LPS) or endotoxin forms part of the cell wall of gram-negative bacteria. Endotoxin is a ubiquitous contaminant and is abundant in the parent tissues (such as the small intestine or dermis) from which many biologic surgical mesh materials, such as ECM-based scaffolds, are manufactured. Differences in the source materials, method of decellularization, and terminal sterilization may all affect the content of potentially immune modulating factors, like endotoxin, within an ECM scaffold. It has been assumed that endotoxin contamination within an ECM scaffold material may have deleterious effects upon the host immune and remodeling responses, 20 presumably through the activation of the toll like receptor 4 (TLR4) signaling pathway by LPS. 21 This would be in direct contrast to the anti-inflammatory immune responses that have been associated with constructive remodeling of ECM scaffolds.4,16,17,19
A recent study using Puramatrix-loaded hexamethylenediisocyanate-crosslinked dermal sheep collagen discs implanted subcutaneously into rats suggested that endotoxin contamination may delay foreign body reactions and degradation. 20 The amount of endotoxin contamination within or upon biologic scaffolds required to elicit an immune or remodeling effect in recipients is unknown. In addition, extrapolation from prior studies is difficult due to the differences in the model and the use of synthetic, crosslinked, or metallic biomaterials, each of which elicit context-dependant host and tissue remodeling responses. Consequently, the standards 22 for the amounts of endotoxin that can be considered either safe or minimal risk in biologic ECM biomaterials are unclear.
The objective of the present study was to determine the effect of deliberate endotoxin contamination of a noncrosslinked porcine dermal biologic scaffold device upon the host response to the material both in vitro and in vivo and upon the ability of the material to promote constructive tissue remodeling.
Materials and Methods
Study overview
The host immune and tissue remodeling responses to LPS were examined using in vitro and in vivo test systems. In the in vitro system, THP1 cells (a human monocyte cell line) were seeded onto dermal ECM scaffolds that had (1) less than 10 EU LPS (X), (2) greater than 20 EU LPS (X20), (3) or less than 20 EU LPS but were deliberately contaminated by soaking in a solution containing 100 ng/mL LPS (X100) for 15–20 min. The in vitro immune response to each material was determined by assessment of cell proliferation, cell death, differential expression of the LPS receptor TLR4 mRNA, and secretion of chemokines CCL2 and CCL4 and cytokines interleukin 10 (IL-10) and tumor necrosis factor alpha (TNF-α) into in vitro cell culture supernatants.
In the in vivo system, a rodent abdominal wall defect model was used to examine the host response to each implanted dermal ECM device. The same devices as in the in vitro model (X, X20, and X100) were implanted. Body temperature was monitored four times daily for 3 days, daily up to 1 week, and then weekly until sacrifice. Animals were sacrificed on days 1, 3, and 35 following implantation, at which time the systemic response was assessed by complete blood counts and investigation of the local tissue remodeling response by histologic methods.
Dermal ECM scaffolds
Each of the dermal ECM scaffolds used in this study were comprised of XenMatrix and were received sterile from the manufacturer (CR Bard, Davol, Inc., Warwick, RI). However, the devices varied in endotoxin content as follows: (1) ≤10 EU of endotoxin (X), (2) ≥20 EU (X20) of endotoxin, (3) ≤10 EU of endotoxin but soaked in a solution containing 100 ng/mL (equivalent to 1000 EU/mL) LPS (Sigma-Aldrich, St. Louis, MO) for 15–20 min prior to seeding/culture. Endotoxin levels were confirmed by the limulus assay prior to use. Test articles were provided sterile by the manufacturer and were stored at −80°C until use.
Culture of THP1 cells on dermal ECM scaffolds
THP1 cells, a human myelomonocytic cell line, were used to evaluate the immune response in vitro. THP1 cells were cultured in RPM1640 (Invitrogen, Carlsbad, CA) with 10% fetal bovine serum (HyClone Laboratories, Inc., South Logan, UT) and 5% Penicillin/Streptomycin (10,000 U/10,000 μg/mL) (Invitrogen, Carlsbad, CA). All cell culture was performed at 37°C, 20% O2, and 5% CO2. Dermal scaffolds were cut into 1.5 cm2 discs (average thickness 0.02 cm and volume 0.045 cm3) in an aseptic manner, and these devices were cultured with 1×106 THP1 cells in a total volume of 1 mL of culture media. In the in vitro X100 group, LPS was added to the medium of THP1 cells and test article to make a concentration of 100 ng/mL. THP1 cells were cultured on test devices for 1, 2, or 3 days, with supernatants and cell pellets being collected at each time point. Supernatants were stored at −20°C until use. Cell pellets were washed twice in sterile saline, and then stored in trizol (Invitrogen, Carlsbad, CA) at −80°C until RNA extraction with chloroform-phenol.
Analysis of cell proliferation and death
Prior to separation of supernatants and cell pellets, the number of live cells was determined using trypan blue staining (Invitrogen, Carlsbad, CA) and a hemocytometer. Cell death was also calculated by determining the quantity of double-stranded DNA released into the supernatant using a Quant-iT PicoGreen kit (Invitrogen, Carlsbad, CA).
Reverse transcriptase–quantitative polymerase chain reaction
Extracted RNA was converted to cDNA using a cDNA synthesis kit (Invitrogen, Carlsbad, CA) according to manufacturer's instructions. Quantitative polymerase chain reaction (qPCR) was also performed on samples using a iQ5 real-time PCR machine (BioRad, Hercules, CA) and a reaction mix of 2 μL cDNA, 2 μL each primer, and 14 μL SYBR green PCR reaction mix (Applied Biosystems, Foster City, CA). Primers of genes of interest and housekeeping genes were TLR4 (NM_138554.3), forward GCCCTGCGTGGAGGTGGTTC and reverse AGCTGCCTAAATGCCTCAGGGGA (134 base pairs); and 23S rRNA (NM_001025.4), forward GCACAGCCCTAAAGGCCAACCC and reverse TCACCAACAGCATGACCTTTGCG (243 base pairs). Primers were designed using the NCBI primer BLAST tool (www.ncbi.nlm.nih.gov/tools/primer-blast/index.cgi) and synthesized by Eurofins MWG Operon (Huntsville, AL). A relative method of analysis (2−ΔΔCt) was used to evaluate mRNA expression. 23
Concentration of supernatants and western blotting for TNF-α and IL-10
Supernatants from THP1 cells seeded onto all devices and on tissue culture plastic (TCP) (∼800 μL) were concentrated by lyophilization (Dura-Dry MP, Cincinnati, OH) and resuspended in 150 μL of deionized water. These concentrated supernatants were then used for western blotting. Samples were normalized by loading of 50 μg of total protein as determined by BCA assay (Pierce, Rockford, IL) into each well of a 10% acrylamide gel, with concentrated supernatant from THP1 cells on TCP used as a control. Gels were then transferred to blotting paper using a semidry transfer system (BioRad, Hercules, CA), proteins were blotted using SnapID (Millipore, Billerica, MA), and blots were visualized using Supersignal Pico chemiluminescence kit (Pierce, Rockford, IL) with a Kodak image station 2000R and software (Kodak, Rochester, NY). TNF-α (Abcam, Cambridge, MA) and IL-10 (Abcam) were used at concentrations of 1/200 in 0.01% milk/TBST, with secondary antibodies also used at 1/200 in 0.01% milk/TBST.
ELISAs for Chemokines CCL2 and CCL4
Chemokines CCL2 and CCL4 are markers of proinflammatory responses 24 and are associated with increased fibrosis following tissue injury.25,26 Therefore, these markers were investigated as potential markers of an adverse host inflammatory response in the present in vitro studies. Cell supernatants collected from THP1 cultured on test devices were analyzed for CCL2 and CCL4 (SABiosciences, Frederick, MD) by ELISA according to the manufacturer's directions.
Implantation of dermal ECM in a bilateral partial abdominal wall defect model
Sprague Dawley rats (female, 200–300 g) were anesthetized and prepared for sterile surgery (including clipping of hair, chlorhexidine/iodine disinfection of surgical site, and sterile drapes). A bilateral partial thickness abdominal wall defect model was used, with the same test article being implanted on both sides. In brief, for each implant a ventral midline incision was made and the skin and subcutaneous tissue was bluntly dissected to allow better access to the ventrolateral abdominal wall. A bilateral 1.5 cm by 1.5 cm defect was then created in the external and internal oblique muscle layers, leaving the underlying tranversalis fascia and peritoneum intact. The size-matched test device was then sutured into the defect at each of the four corners with 4-0 polypropylene. The dermis and epidermis were closed over the defect with single interrupted sutures of 4-0 vicryl. Rats were given enrofloxacin (34 mg/rat/day) and buprenorphine (0.06 mg/rat/day) daily for 2 days after surgery. Wounds were monitored for dehiscence and seroma formation twice daily. Each rat was implanted with two identical implants (X, X20, or X100, described previously), one on each side of the midline (Table 1).
X, dermal ECM scaffolds with ≤10 EU endotoxin; X20, dermal ECM scaffolds with ≥20 EU endotoxin; X100, dermal ECM scaffolds with ≤10 EU endotoxin and spiked with 100 ng/mL LPS; LPS, lipopolysaccharide.
Temperature was monitored four times daily with an IR-B152 Small Rodent Infrared Thermometer (Braintree Scientific, Braintree, MA) for the first 3 days. From days 4 to 7, temperatures were monitored daily, and then weekly until sacrifice.
Euthanasia, blood collection, and explant harvest
Animals were euthanized by inhalation of carbon dioxide followed by cervical dislocation. Blood was collected immediately by cardiac puncture into ethylenediaminetetraacetic acid tubes and processed by Idexx Laboratories (Westbrook, ME) for a complete blood count. Explants were photographed in situ, explanted, and stored in 10% nonbuffered formalin for histologic processing.
Histologic analysis
Fixed tissue was paraffin embedded, and then 5-μm transverse sections were then cut through the entire explant and stained with hematoxylin and eosin (H&E) and Masson's trichrome. Representative images were generated with a Nikon eclipse E600 microscope (Nikon, Melville, NY) and a Nuance multispectral imaging system (CRi, Woburn, MA). Six high-powered representative fields (400×on H&E-stained sections) in each section were scored for inflammatory cell infiltration into the device at days 1 and 3 by five blinded investigators based upon the criteria detailed in Table 2. Explants from day 35 were scored for constructive remodeling, with a higher score being indicative of a more proinflammatory reaction. That is, a lower score indicates a higher degree of constructive remodeling. Constructive remodeling was assessed in six high-powered representative fields (100×on H&E-stained sections) by five blinded investigators using criteria listed in Table 3.
HPF, high powered field.
Statistics
Analysis of variance (ANOVA) with a Tukey post hoc test (Minitab 15 statistical software) was performed to determine significance. Percentage and ratio data were log transformed prior to ANOVA. Semiquantitative scoring was assessed by a Kruskal-Wallis and then a Mann-Whitney test (Minitab 15 statistical software). A p-value of less than 0.05 was considered a significant result.
Results
In vitro evaluation
Total cell counts and cell death
High rates of THP1 cell proliferation were seen in all groups, a phenomenon that has been previously reported using this model. 27 THP1 cells grown on low endotoxin dermal ECM (X) showed no difference in live cell number or percentage of live cells compared with cells grown on control TCP at any time point (Fig. 1a, b). In comparison, THP1 cells grown on X20 and X100 endotoxin-contaminated matrices had decreased cell number when compared with TCP at day 2 and days 1 and 2, respectively (Fig. 1a). This decrease in cell number compared with THP1 cells alone may be due to the effect of endotoxin contamination of these devices. There was a significant decrease in the percentage of live cells on the X100 samples as compared with all other groups at day 2 (Fig. 1b). The rate of cell death in cultures was further quantified by assessment of the amount of double-stranded DNA in the culture supernatant. At all time points, there were significant differences between the groups, with the greatest amount of double-stranded DNA being found in cultures of THP1 cells on X100 dermal matrix (all time points) and X20 matrix (days 2 and 3), indicating these groups exhibited the highest degree of cell death (Fig. 1c). THP1 cells grown in dermal matrix alone (X) had significantly higher double-stranded DNA compared with TCP at day 1 (p<0.01), but less than groups X20 and X100 (Fig. 1c).
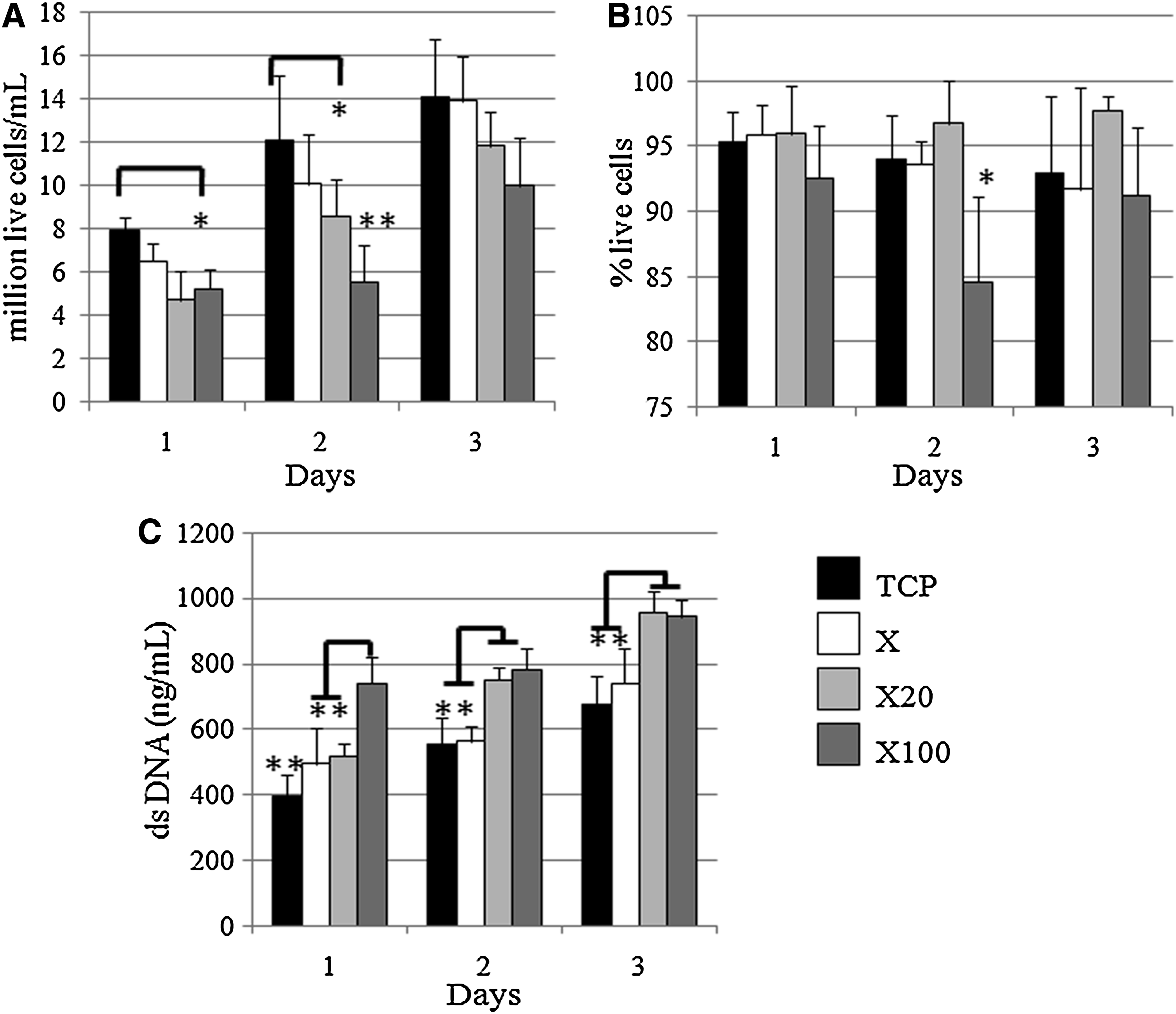
X100 devices decreased the proliferation rate of THP1 cells and increased the cell death.
Secretion of CCL4 and CCL2
No detectable CCL4 was found in supernatant from THP1 cells grown on TCP (data not shown). Greater levels of CCL4 were found in supernatants from the THP1 cells cultured in the presence of 100 ng/mL of LPS only as a control (i.e., no scaffold material; LPS100) than in all other groups at all time points (p<0.01) (Fig. 2a). CCL4 was found in greater amounts in supernatants from the X100 dermal ECM than in the other dermal ECM groups at days 1 and 3 (Fig. 2a).
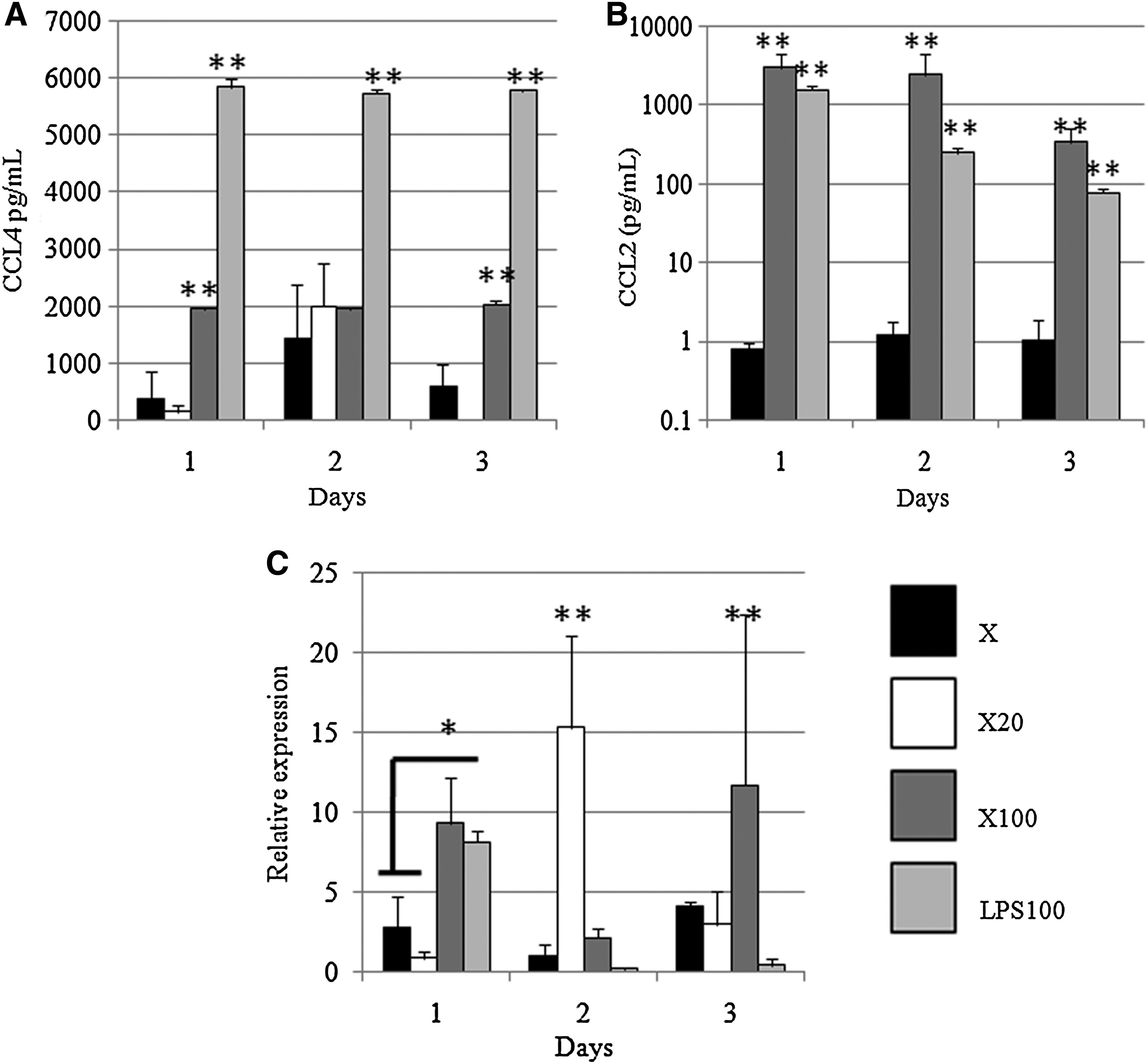
Expression of TLR4 mRNA, chemokines, and cytokines in response to dermal matrices.
No CCL2 was detected in supernatants of THP1 cells on TCP or X20 dermal ECM. Very small concentrations of CCL2 were seen in supernatant from the X dermal matrix group (Fig. 2b). The highest secretion of CCL2 was found in supernatant from THP1 cells on X100 dermal ECM (Fig. 2b). The amount of CCL2 detected in supernatants from the THP1 cells with 100 ng/mL of LPS alone control group (LPS100) was also markedly increased when compared with that found in the X dermal matrix group (p<0.01) (Fig. 2b).
TLR4 mRNA expression
Expression of TLR4 mRNA in THP1 cells in response to test devices was compared with that in THP1 cells on TCP (Fig. 2c). At day 1, there was greater expression of TLR4 mRNA in response to X100 dermal matrix and LPS alone control (LPS100) than was observed for other groups (p<0.05). While LPS may induce tolerance for up to 5 days after an initial spike in proinflammatory responses, as seen in response to LPS100, TLR4 mRNA expression was also increased in response to X20 dermal matrices at day 2 and to X100 devices at day 3. Recently, factors intrinsic to biomaterials have been shown to activate TLR signaling pathways on responding immune cells,27–30 which may be the case here. In addition, this explanation is consistent with the increased (i.e., >1) expression seen in THP1 cells in response to X dermal matrix.
IL-10 and TNF-α expression
Both TNF-α and IL-10 were found to be present in concentrated supernatants from THP1 cells cultured on all devices. No difference was found in expression between any of the groups for either TNF-α or IL-10, although there was a decrease in expression over the 3 days (data not shown).
In vivo model
Body temperature after device implantation
Implantation of X100 dermal ECM resulted increased in body temperature compared with X dermal matrix for the most of the first 54 h after implantation (Fig. 3a, b). X20 dermal ECM induced lower changes in temperature than X100 matrices, with significant differences observed at 18, 30, 40, and 54 h postimplantation. X20 dermal matrices induced an increase in body temperature compared with X dermal matrix, but only at 18 h (Fig. 3b). From 60 h onward (Fig. 3a), there was no difference between the test articles in terms of the change in body temperature induced by device implantation.

Change in body temperature for rodents implanted with dermal matrices.
Complete blood counts
Hematocrit, hemoglobin, erythrocyte cell counts, and other related parameters were unchanged over the time period of the study and in comparison to reference ranges. 31 Total leukocyte counts in the lower part of the reference range were found in all groups at day 1 (Fig. 4a) but had increased by days 3 and 35. The percentage of monocytes was within the reference range for all groups at day 1 (Fig. 4b). There was no difference in lymphocytes between all groups at either time point (Fig. 4c) but neutropenia was found in response to X dermal matrix at day 1 and X20 dermal matrix at day 3 (Fig. 4d). When the ratio of lymphocytes to neutrophils was calculated, there was an increase in this ratio in response to X dermal matrix at day 1 (Fig. 4e) (p<0.05). There was an increase in the lymphocyte/neutrophil ratio in all groups at day 35 as compared with the reference range (Fig. 4e).
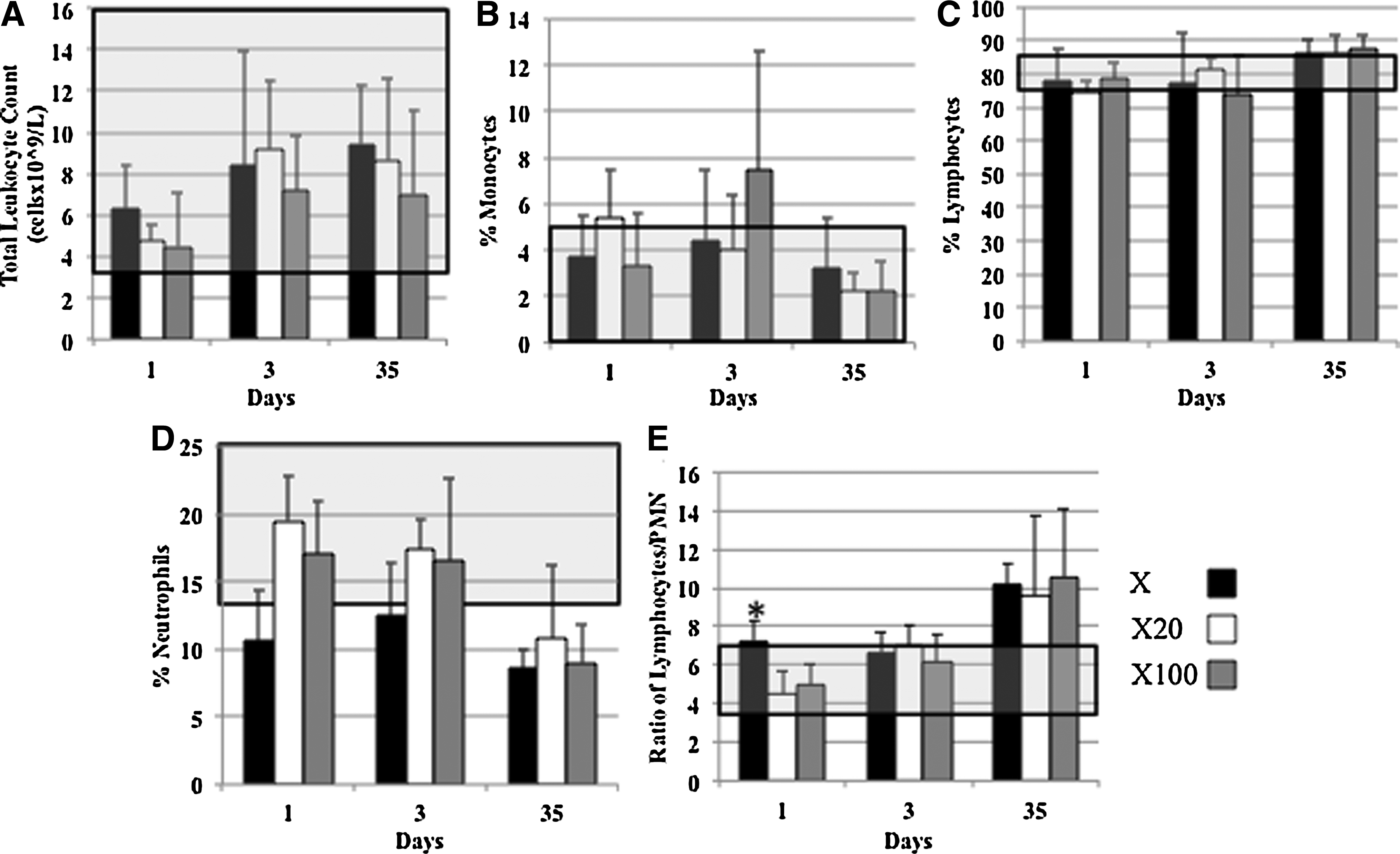
Systemic cell response to dermal matrices.
Inflammatory cell infiltration
Macroscopically, there was no difference noted between any of the test devices. At early time points (days 1 and 3), there was limited integration of the device into the host tissues. Occasional seroma and hematoma presence was noted in some animals from all groups at days 1 and 3, but these were not found at day 35. Devices from the X100 group appeared to have diminished in size compared with the other groups macroscopically at day 35 (data not shown).
At early time points (days 1 and 3), cellular infiltration of all devices was found predominantly at the edges of the implants (Fig. 5a, day 1 only). No fibrous connective tissue deposition response was observed in these early time points. There was edema in the surrounding muscle parenchyma at days 1 and 3 (Fig. 5a). By day 35, cells were found to infiltrate the entire device in all groups (Fig. 5b). Degradation of the scaffold was most pronounced at the periphery with fibrous tissue, angiogenesis, and foreign body giant cells noted (Fig. 5b). Devices from all groups were found to be surrounded by a dense and circumferentially organized connective tissue. Mononuclear cells were still found infiltrating and surrounding the X100 dermal matrices at day 35, but were decreased in number compared with earlier time points.

H&E sections of explants show inflammatory cell infiltration of the devices, with edema of the surrounding muscle parenchyma seen in response to the X100 devices. By 35 days, deposition of host tissue is seen in the periphery of each device, with fibrous tissue surrounding the implant.
The cells infiltrating the devices were a mixture of inflammatory cells—that is monocytes, neutrophils, and lymphocytes (Fig. 6). Semiquantitative scoring of the inflammatory cells indicated that there were less inflammatory cells infiltrating the X20 dermal matrix than the other devices at day 1 (Fig. 6b). By day 3, the X100 device had more inflammatory cells infiltrating than both other groups (Fig. 6b), although the score was not different from that at day 1. Infiltrating cells were found to have decreased in X devices at day 3 compared with day 1 (p<0.01). Semiquantitative scoring of all groups at day 35 assessed the host constructive remodeling response, with a higher score being indicative of a more proinflammatory reaction. X20 dermal matrices had lower scores than X100 devices, but no difference was observed when compared with X test articles (Fig. 6c) (p<0.05). The relatively high scores seen in all groups are consistent with the early host response to ECM, when there is still significant scaffold to be degraded.4,17
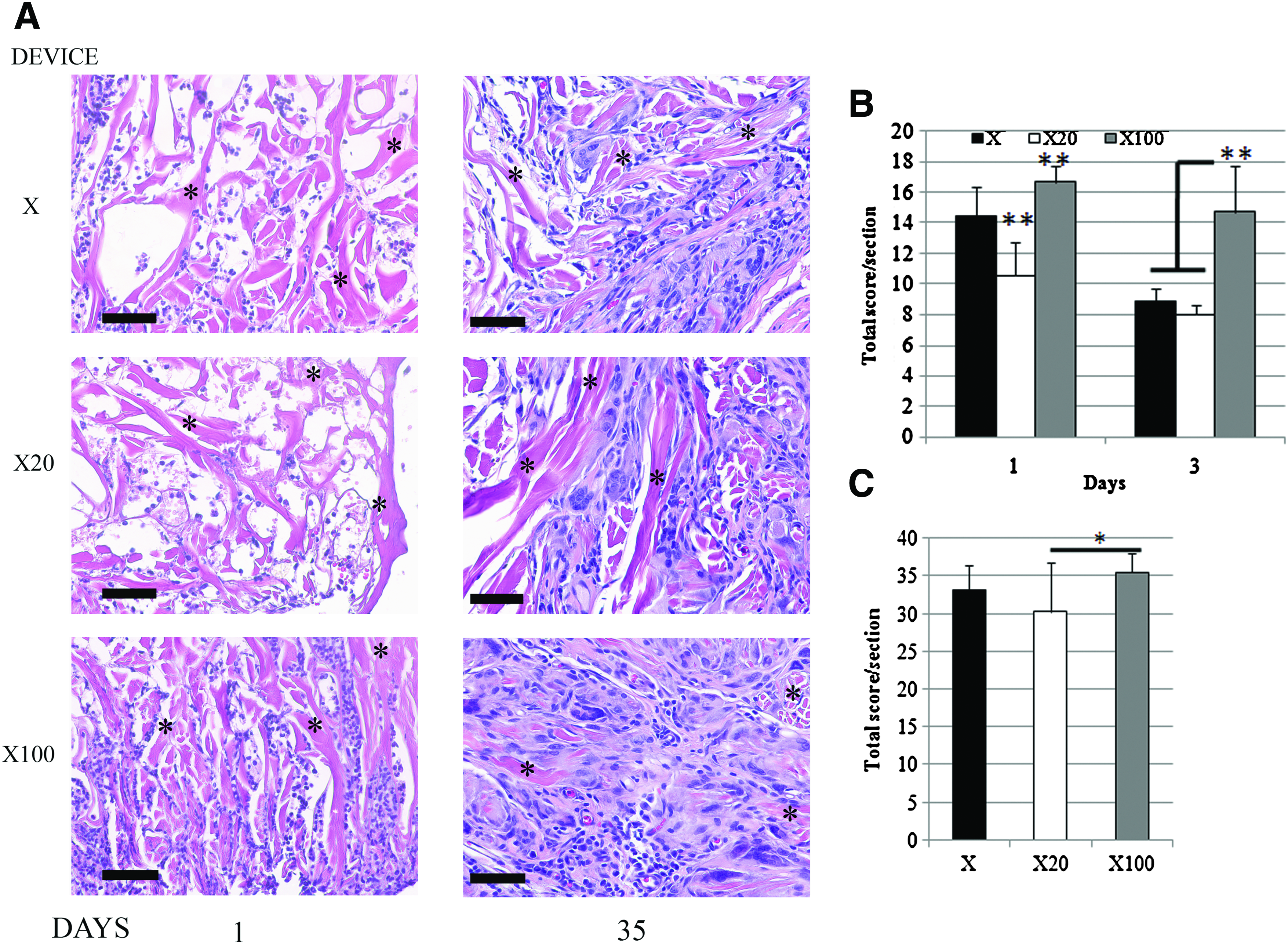
Discussion
Endotoxin is a ubiquitous contaminant that can initiate adverse immune reactions when present in significant quantities on implantable medical devices. The amount of endotoxin within or upon an implanted biomaterial required to elicit an adverse reaction is unknown. The manufacture of biologic scaffolds from xenogeneic tissue sources that are in contact with commensal bacteria (such as intestines and skin) requires rigorous decellularization, aseptic techniques, terminal sterilization, and thorough quality assurance to minimize the residual LPS. Despite these steps, LPS present within these tissues cannot be entirely removed. Therefore, acceptable amounts of endotoxin within or upon a xenogeneic biologic scaffold, which do not elicit a deleterious in vivo proinflammatory reaction nor lead to a foreign body reaction, need to be systematically determined.
In the present study, the effect of endotoxin contamination on the host immune to a dermal scaffold was examined. Increased THP1 cell death was noted in X20 and X100 groups, which may be due to endotoxin induction of pyronecrosis. Despite this increase in the rate of in vitro cell death and in TLR4 mRNA expression, X20 dermal matrices (i.e., >20 EU endotoxin) induced similar expression of chemokines and cytokines as X dermal devices (i.e., <10 EU endotoxin). This expression of proinflammatory chemokines was less than the robust response induced by X100 matrices and cells cultured in media containing 100 ng/mL LPS as a positive control. An in vivo host immune response that was largely indistinguishable from that to X devices was also noted when X20 devices were implanted in a rodent partial abdominal defect model. No differences were found in the long-term host response to X and X20 dermal matrices. These results suggest that the current U.S. Food and Drug Administration endotoxin standards 22 for biomaterials may fall well below the dose required for deleterious effects upon long-term remodeling. The results also demonstrate the caution with which in vitro data should be considered with regard to predicting in vivo responses.
Increased TLR4 expression has been associated with fibrosis,32,33 chronic graft dysfunction, and increased proinflammatory polarization. 34 However, TLR signaling pathways have also been correlated with the early stages of hepatic regeneration. 33 Stated differently, long-term activation of TLR4 signaling pathways may be correlated with a deleterious host response and a foreign body reaction. TLR4 mRNA expression in THP1 cells was found to increase at day 1, and then decrease at day 2, with a smaller increase in expression at day 3 in response to X100 test devices. This variable expression pattern may be explained by the induction of tolerance to LPS and the possible role of endogenous TLR ligands within the dermal matrix27–30 or damage-associated molecular pattern (DAMP) molecules released by increased cell death seen in this group at day 3. However, the dose of LPS required to achieve this response was 100 ng/mL (∼500–1000 EU/mL depending on the endotoxin detection method), which well exceeds the FDA limit for endotoxin in medical biomaterials. These results appear to contrast with a recent study where endotoxin (at 100 ng/mL and higher) was found to delay the progression of a foreign body reaction to a crosslinked ovine dermal collagen biomaterial in vivo. 20 This disagreement may be explained by the differences in the type of endotoxin and the nondegradable biomaterial used in the other study.
Prolonged TLR4 signaling can induce tolerance so as to limit the extent of the proinflammatory reaction by preventing reactivation of immune cells. Proinflammatory genes, such as TNF-α and CCL2, are downregulated, while IL-10 is inducible during tolerance, 35 which is consistent with the immune response to X20 and X dermal scaffolds. Both IL-10 and TNF-α were found to be secreted by THP1 cells in vitro with no difference detected between groups. Previous studies have also detected increased IL-10 secretion in response to LPS spiked biomaterials, which was associated with a decreased foreign body reaction. 20 Induction of a tolerance to low doses of endotoxin or other TLR ligands within the biomaterial may explain the decreased immune response seen to the X20 test articles seen in this study.
Tissue damage itself may induce a significant immune reaction from the host; consequently, the effect of the injury alone on the induction of host immune responses cannot be ignored in our in vivo rodent model. Tissue damage results in the release of DAMPs, which act as danger signals to activate the immune system by proinflammatory pathways such as TLR. Inflammatory cells were seen to infiltrate the edges of the wound in defect only day 2 controls, and these cells were observed surrounding necrotic myotubes. Similar patterns were noted in all groups implanted with a dermal ECM, regardless of endotoxin level. Intramuscular implantation of a biomaterial, as in this study, has been correlated with increased immune reactions when compared with a subcutaneous model, 36 suggesting that the greater injury induced in the intramuscular model may contribute to the activation of the immune response. To separate the effect of the injury and the response elicited by the test article, the present study focused on the cells infiltrating the dermal matrices, to gauge the effect of this microenvironment and not that of the injury on the host response.
In the present study, a myelomonocytic cell line (THP1 cells) was chosen to model the host response in vitro. Mononuclear cells (including monocytes) have been found to infiltrate the interface of biologic scaffolds and the host within days of implantation in vivo.2,37 Of these cells, macrophages are vital for the constructive remodeling of the biologic scaffold. 17 THP1 cells have been used previously to evaluate biomaterials (in both stimulated and unstimulated forms).27,38 As monocytes are among the first responder cells to an implanted biomaterial, THP1 cells were not differentiated before exposure to test devices so as to assess the acute reaction, and prior stimulation with PMA, cytokines, or growth factors may change the immune response. However, there are limitations to the use of cell lines. To overcome the limitations of using a cell line, an in vivo model was also used to assess the host response to endotoxin contamination of dermal matrices.
The role of neutrophils in the acute host response to biomaterials has been suggested by previous studies2,37 but is still unclear. In the present study, alterations in the systemic leukocyte count, and in particular the neutrophil count and lymphocyte/PMN ratio suggested that neutrophils may also play a role in the response, particularly to noncontaminated dermal matrices. Decreases in systemic neutrophil counts may reflect increased migration of neutrophils to the implantation site. However, as no neutrophil-specific staining and quantification was undertaken, it is difficult to draw any conclusions. The role of neutrophils in the host immune and constructive immune responses clearly requires further research.
In conclusion, the present study has examined the role of endotoxin contamination within a dermal ECM scaffold on the host response. Test articles with amounts of endotoxin that exceed the current FDA limit (X20) were found to have similar or decreased immune responses both in vitro and in vivo when compared with devices that fell under the endotoxin limit (X). In contrast, dermal matrices spiked with 100 ng/mL endotoxin (a dose equivalent to 500–1000 EU/mL) elicited a robust immune response from THP1 cells in vitro and an increased acute proinflammatory reaction in vivo. However, at 35 days after implantation, no difference was detected between the implantation groups regardless of the endotoxin level within the test article. These results suggest that the current endotoxin standards may fall well below those levels required to induce an acute proinflammatory response with associated long-term deleterious remodeling outcomes. In addition, this study suggests that receptors such as TLR4 may have a role in the immune cell interaction with ECM biomaterials and that these scaffolds may contain endogenous ligands for this receptor (apart from any endotoxin). Understanding those signaling pathways responsible for the host immune/biomaterial interaction may have important implications for future biomaterial development.
Footnotes
Acknowledgments
The authors wish to acknowledge the assistance of Joe Hanke, Teri Gasser, Amy Wiester, Jennifer Kerr, Amie Hanz, Michael Valles, and Bradley Kurtik from the McGowan Animal Facility for the help with the animal subjects, and Deanna Rhoads for her help with histology.
Disclosure Statement
No competing financial interests exist.