
Abstract
Bone allografts are commonly used by orthopedists to provide a mechanical support and template for cellular colonization and tissue repair. There is an increasing demand for bone graft substitutes that are safe and easy to store but which are equally effective in supporting new bone growth. In this study, we compared three different human bone allografts: (1) the cryopreserved allograft (frozen), (2) the gamma-irradiated and cryopreserved allograft (γ-irradiated), and (3) the solvent dehydrated and γ-irradiated-processed bone allograft (Tutoplast® Process Bone [TPB]). Human mesenchymal stromal cells (hMSCs) have the potential to differentiate into osteogenic, chondrogenic, and adipogenic lineages. Our results showed that hMSC seeding efficiency was equivalent among the three bone allografts. However, differences were observed in terms of cell metabolism (viability), osteoblastic gene expression, and in vivo bone formation. Frozen allografts had the higher frequency of new bone formation in vivo (89%). Compared with frozen allografts, we demonstrated that TPB allografts allowed optimal hMSC viability, osteoblastic differentiation, and bone formation to occur in vivo (72%). Further, the frequency of successful bone formation was higher than that obtained with the γ-irradiated allograft (55%). Moreover, after hMSC osteoinduction, 100% of the TPB and frozen allografts formed bone in vivo whereas only 61% of the γ-irradiated allografts did. As healthcare teams around the world require bone-grafting scaffolds that are safe and easy to store, the TPB allograft appears to be a good compromise between efficient bone formation in vivo and convenient storage at room temperature.
Introduction
Whereas ceramics are safe and are chemically close to the natural mineral structure of bone, these scaffolds showed variations in their properties of resorption, osteoconduction, and osteoinduction. Moreover, they are very different from the structure of authentic bone and the biomechanical properties of bone allografts are superior to all ceramic substitutes. 12 As orthopedists commonly use allograft to provide a template for cellular colonization and tissue repair, researchers and manufacturers have developed different processes to ensure bone allograft biosafety and easy storage at room temperature.11,13–15 These processes include the complete removal of donor cells and the elimination of pathogens and antigenic agents.4,13–15 Finally, the processed bone is γ-irradiated without affecting its biomechanical properties.11,12
The success of the graft may depend on the quality of the bone bed from which most of the revascularization arises. Allografts processing, preservation, and sterilization may reduce its osteoinductive capacity, and when allografts are used alone, bone repair is less effective than an autograft because the osteoinductive proteins and osteogenic cells of normal bone are missing. To enhance the osteoinductivity after processing, various types of cells and tissue factors are available. Mesenchymal stromal cells (MSCs) are multipotent cells that can differentiate into osteogenic, chondrogenic, and adipogenic lineages and produce trophic factors for tissue repair. 16 The combination of human mesenchymal stromal cells (hMSCs) with biomaterials has previously shown its efficiency on ectopic bone formation, as well as on large bone defects in several experimental animal models.17–22 In contrast, few reports have studied hMSC implantation on allografts.
The aim of this study was to assess and compare the in vitro and in vivo properties of human bone allografts by “vitalizing” them with hMSCs. We compared human allografts processed by three different methods: (1) the cryopreserved allograft (frozen), (2) the γ-irradiated and cryopreserved allograft (γ-irradiated), and (3) the solvent-dehydrated and γ-irradiated-processed bone (Tutoplast® Process Bone [TPB]). We tested the cell-seeding efficiency, metabolic activity, and osteoblastic gene expression of hMSCs in the allografts. We assessed the expression of markers for early osteoblastic differentiation (Runx2, alkaline phosphatase [ALP], bone morphogenetic proteins [BMPs]) and late osteoblastic differentiation (bone sialoprotein [BSP], osteocalcin [Oc], and osteopontin [Op]) using quantitative reverse transcription-polymerase chain reaction (RT-PCR). Finally, we evaluated the extent of bone formation in vivo in an ectopic SCID mouse model.
Materials and Methods
Bone substitute
With the consent of donors, femoral heads were taken from patients undergoing hip arthroplasty and stored at −80°C in the tissue bank of “Etablissement Francais du Sang” (EFS) Ile de France. The human frozen femoral head can follow 3 different processes of biosafety:
1. The bone tissue is tested for bacteria and viruses (frozen bone).
2. The bone tissue is tested for bacteria and viruses, and γ-irradiated at 25,000 Gy (γ-irradiated bone).
3. The bone tissue is Tutoplast processed (Tutogen Medical). The Tutoplast process consists in a delipidization, an osmotic cell destruction treatment, hydrogen peroxide treatment, and washing cycles for removal of the noncollagen proteins followed by a solvent dehydrated step and finally a γ-irradiation procedure (TPB). 14
Frozen, γ-irradiated and TPB femoral head was provided by EFS. The femoral head cancellous bone was cut manually into fragments from 2 to 4 mm. Cancellous bone particles of equivalent size, volume, and weight were used in this study to ensure a comparable surface area for in vitro and in vivo analyses (Table 1).
TPB, Tutoplast® Process Bone.
Platelet lysate preparation
Platelet apheresis collections performed at the EFS (Rungis, France) were biologically qualified in accordance with the French legislation. The platelet count in each product was measured automatically (with an ABX penta 60 Cþ, Horiba ABX). Only samples containing 109 to 2.5×109 platelets/mL were retained. They were frozen at −80°C and subsequently used to obtain platelet lysate (PL) containing platelet-released growth factors. Different batches were obtained (with two to five apheresis collections) to adjust the concentration at 50×106 platelet/mL. This concentration in platelets was previously shown to allow an amplification of hMSCs three times faster compared with fetal bovine serum. 18
Bone marrow cell cultures
Human bone marrow (BM; 3–5 mL volumes) was obtained from iliac crest marrow aspirates of donors and orthopedic patients (Henri Mondor Hospital, AP-HP, Creteil, France) after receiving their informed consent. BM from 9 donors (27 to 52 years old) was used. Nucleated cells from fresh marrow were seeded at 2×105/cm2 in 225 cm2 flasks. hMSCs were expanded in α-modified Eagle's medium (αMEM glutamax) (PAA Pasching) containing 5% PL (αMEM-5%PL) as previously described. 18 The medium was supplemented with 0.5% Ciprofloxacin (Bayer Pharma), and 2 IU/mL Heparin (Sanofi-Aventis) was added to avoid clot formation. The culture medium was changed twice a week and the cultures were maintained in a humidified atmosphere with 5% CO2 at 37°C. When cells reached 80%–90% confluence (passage 0 [P0]), they were detached using trypsin/EDTA (PAA Pasching) and then replated at 1000 cells/cm2 until 80%–90% confluence (passage 1 [P1]). Cell samples were used to confirm the hMSC characteristics as previously described. 18 Briefly, all the MSCs used in this study were positive for CD90, CD105, and CD73, and negative for CD34 and CD45 and were able to differentiate into osteogenic, chondrogenic, and adipogenic lineages (data not shown).
Seeding of MSCs
Bone substitutes were humidified in αMEM at 37°C for 2 h prior to seeding the cells. Then, bone substitutes were loaded individually with 3×105 hMSCs in 20 μL of αMEM-5%PL into untreated 96-well culture plates for 3 h at 37°C in a 5% CO2 atmosphere. Bone substitutes were then cultured individually in 1 mL of αMEM-5%PL into untreated 24-well culture plates for 7 days. Half of the samples were cultured in αMEM-5%PL supplemented with ascorbic acid 5×10−5 M, β-glycerophosphate 10−2 M, and dexamethasone 10−7 M for 7 days to induce osteoblastic differentiation. Cell-free scaffolds were incubated under similar conditions and served as controls.
Seeding efficiency
Cell-seeding efficiency was performed using an indirect method. After 3 h of contact with 300,000 hMSCs, the bone substitute was removed and cells that did not attach to the bone substitute were quantified by DNA Picogreen® labeling. Then, the cells that adhered to the bone substitute were deduced by subtraction. Briefly, the cells that remained floating in the medium and that adhered to the bottom of the plate were lysed with Tris EDTA 0.1% Triton X100 and 0.2 mg/mL of proteinase K. Samples were incubated overnight at 52°C and then frozen-thawed three times, vortexed, and sonicated for 10 min. In parallel, a standard curve with cell range from 0 to 300,000 hMSCs was carried out simultaneously. DNA samples were labeled with picogreen (Invitrogen) that only bound double-strand DNA. After 10 min of incubation at room temperature in the dark, the fluorescence was measured at 520 nm on a spectrofluorimeter (Victor Wallac, PerkinElmer). The standard curve was then used to quantify the cells.
MTS assay
After 7 days of culture on the bone scaffolds, the cell viability was assessed using MTS assay (Promega). Cellularized bone scaffolds were transferred to a 96-well plate containing 120 μL of MTS reagent. After 1 h at 37°C in 5% CO2, 100 μL was collected and absorbance was measured at 490 nm with an ELISA plate reader (Victor Wallac, PerkinElmer). Uncellularized bone scaffolds served as a negative control.
Scanning electron microscopy
Three hours and 7 days after seeding, bone substitutes were fixed with 2.5% glutaraldehyde (Sigma-Aldrich) and then dehydrated with increasing concentrations of ethanol. The dehydration was completed with 60 min of incubation with hexamethyldisilazane (HMDS; Sigma-Aldrich). Samples were then air-dried, sputtered with nano-gold film, and analyzed with scanning electron microscopy (JEOL JSN-6301F) at LISA laboratories (Paris-Est University, Créteil, France).
Quantitative reverse transcription-polymerase chain reaction
Total RNA was isolated from hMSC cultures on day 0 and day 7 using an RNeasy mini-kit as described by the manufacturer (Qiagen). Dnase (Qiagen)–treated RNA was reverse transcribed with RT superscript III (Invitrogen). The cDNA was amplified by TaqMan Polymerase chain reaction (Applied Biosystems) with specific primers (Table 2), and monitored with the ABI Prism 7500 sequence detection system (PerkinElmer/Applied Biosystems). Amounts of cDNA were normalized to GAPDH (ΔCt=CT gene of interest−CT GAPDH). Quantitative PCR was performed in duplicate for each of the nine different hMSC samples analyzed.
ALP, alkaline phosphatase; BMP, bone morphogenetic protein; BSP, bone sialoprotein.
Animal model and implantation procedure
Twenty-one male (7 weeks old) SCID mice purchased from Charles River Laboratories were used in this experiment in accordance with French law on animal experimentation. Isoflurane was used for anesthesia. Subcutaneous dorsal pockets (0.5 cm incisions) were prepared on each mouse. In each pocket, one scaffold was implanted and the skin was closed by suture (Technicon). hMSCs from three different BMs were tested in sixplicate in six independent animals (n=18 scaffolds per group). After 7 weeks, animals were euthanized with an overdose of pentobarbital and the specimens were excised and immediately fixed in 70% ethanol.
Histology
Specimens were decalcified for 3 h in 6.8% nitric acid (VWR) and then rinsed in tap water before being embedded in paraffin. Sections (3 μm) were stained with Masson's trichrome or hematoxylin-eosin and visualized using standard light microscopy. Fifteen sections of each sample were analyzed (5 at the beginning, 5 in the middle, and 5 at the end). New bone was quantified with ImageJ software and new bone density was defined as the ratio of new bone area compared with total implant area ×100.
Statistical analysis
Kruskal-Wallis test was used to compare groups and all data were statistically analyzed using multiple comparisons by the Dunn's test (analysis of variance graphpad software). Comparison of two experimental conditions (two-dimensional [2D] vs. 3D or induced vs. noninduced) was performed using the unpaired nonparametric Mann-Whitney t-test (graphpad software). Differences between the groups with a p-value of ≤0.05 were considered to be significant.
Results
Human MSC seeding efficiency and viability in bone allografts
To compare TPB allograft with γ-irradiated and nonirradiated frozen bone allografts, we first tested hMSC seeding efficiency and metabolic activity. After 3 h of static contact, a mean of 150,000 hMSCs had adhered to the scaffolds (Fig. 1a). Cell-seeding efficiency was equivalent for the three different bone allografts. The effect of the three different bone allografts on cell morphology and distribution was evaluated by scanning electron microscopy (Fig. 2). Three hours after seeding, ball-shaped hMSCs are distributed uniformly around the allografts independently of the allograft preservation and sterilization conditions (Fig. 2d–f). After 7 days of culture on the bone allograft, scanning electron microscopy analysis showed that the cells had fully covered the scaffold and cell multilayer can also be observed (Fig. 2g–i). We next performed MTS assay to evaluate the hMSC viability on the bone allografts. Our results showed that hMSCs on the TPB allografts had the highest metabolic activity compared with those on frozen or γ-irradiated bone allografts (Fig. 1b).

Seeding and metabolic activity of human mesenchymal stromal cells (hMSCs) on bone substitutes.

Scanning electron microscopy. Frozen bone
Gene expression in bone-allograft-seeded hMSCs
We investigated the effects of the different bone allografts on osteogenic gene expression in the hMSCs using quantitative RT-PCR. hMSCs cultured for 7 days on bone allografts (3D) were compared with hMSCs cultured in 2D flasks (n=9). The frozen, γ-irradiated, and TPB bone allografts were able to induce with high significance (p≤0.001) the expression levels of late osteoblastic genes, including Op (4922-, 1690-, and 386-fold, respectively), BSP (22-, 36-, and 31-fold), and BMP2 (546-, 45-, and 38-fold) (Fig. 3). Oc which is a very late osteoblastic gene coding for matrix bone protein was found upregulated by all three scaffolds (10-, 4-, and 3-fold, respectively), with a high significant induction only for frozen bone allografts (p≤0.001). Oc and BMP2 were induced to significantly higher levels in the frozen allografts compared with the TPB allografts (3- and 15-fold, with a p≤0.05 and p≤0.01, respectively). All the genes were more highly expressed in γ-irradiated allografts compared with the TPB allografts, but the differences were not statistically significant. Runx2, osteonectin (On), BMP4, and BMP6 were not significantly upregulated (Fig. 3). Further, ALP was more significantly downregulated in frozen allografts compared with the γ-irradiated or TPB bone allografts (p≤0.01).
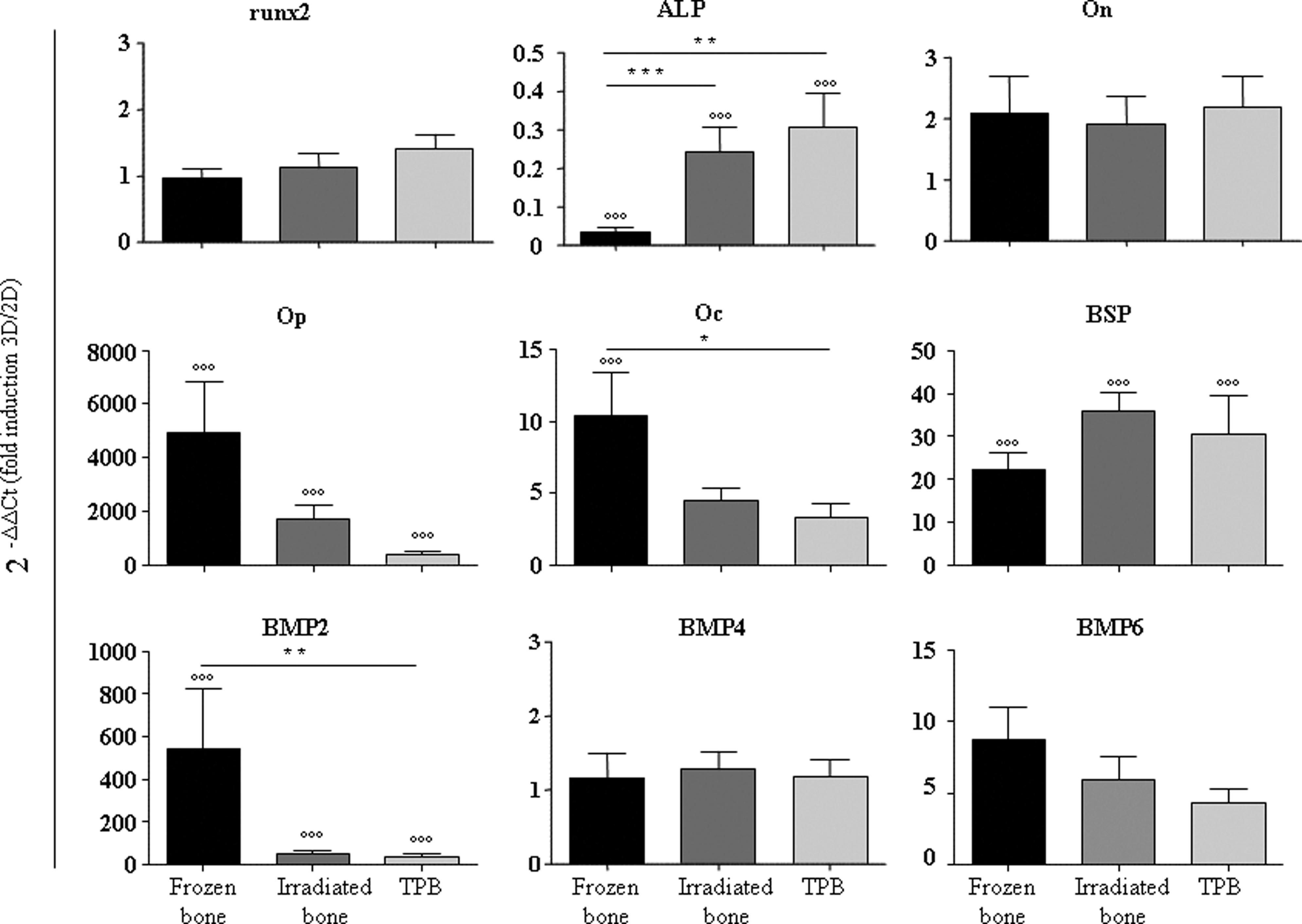
Osteoblastic gene expression of hMSCs cultured on bone substitutes. Osteoblastic (Runx2, alkaline phosphatase [ALP], bone sialoprotein [BSP], osteopontin [Op], osteocalcin [Oc], osteonectin [On], and bone morphogenetic proteins [BMPs] 2, 4, and 6) gene expression is determined by quantitative (TaqMan) RT-PCR on hMSCs on frozen bone, γ-irradiated bone, or TPB (three-dimensional [3D]). hMSCs cultured for 7 days on bone allografts (3D) were compared with hMSCs cultured in 2D flasks. Values for all genes were normalized to GAPDH (ΔCt) and to the gene expression for hMSCs cultured in flask (2D) (2−ΔΔCt (fold induction 3D/2D)). hMSCs from nine different bone marrow (BM) samples were used for qRT-PCR and each experiment was done in duplicate. Mean data±SEM are presented.*/p<0.05; **/p<0.01; ***/°°°p<0.001 (ANOVA test [*] was performed between the three allografts. Mann-Whitney test [°] was performed when 3D was compared with 2D. RT-PCR, reverse transcription-polymerase chain reaction.
Impact of the scaffolds on ectopic bone formation in vivo
We investigated the impact of the bone allografts on bone formation in vivo by using an ectopic bone implantation model in immunodeficient SCID mice. Frozen, γ-irradiated, and TPB bone allografts were tested alone or combined with hMSCs isolated from three different BM donors. Seven weeks after implantation, the bone allografts were collected and then decalcified and stained using Masson's Tri-chrome and hematoxylin-eosin (Fig. 4 and Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/tea). Neovascularization occurred on unseeded allografts as well as on bone allografts seeded with hMSCs. There was no evidence of inflammatory reaction and only minimal indications of bone resorption were detected. In the unseeded controls, no new bone formation was observed but loosely organized connective tissues were present (Fig. 4a–c). In contrast, for bone allografts seeded with hMSCs, we detected new bone formation in all three different bone allografts adjacent to the scaffold (Fig. 4d–i). This new tissue contained osteocyte-like cells and osteoblast-like cells lining the surface. BM-like elements could also be observed (Fig. 4g–i).
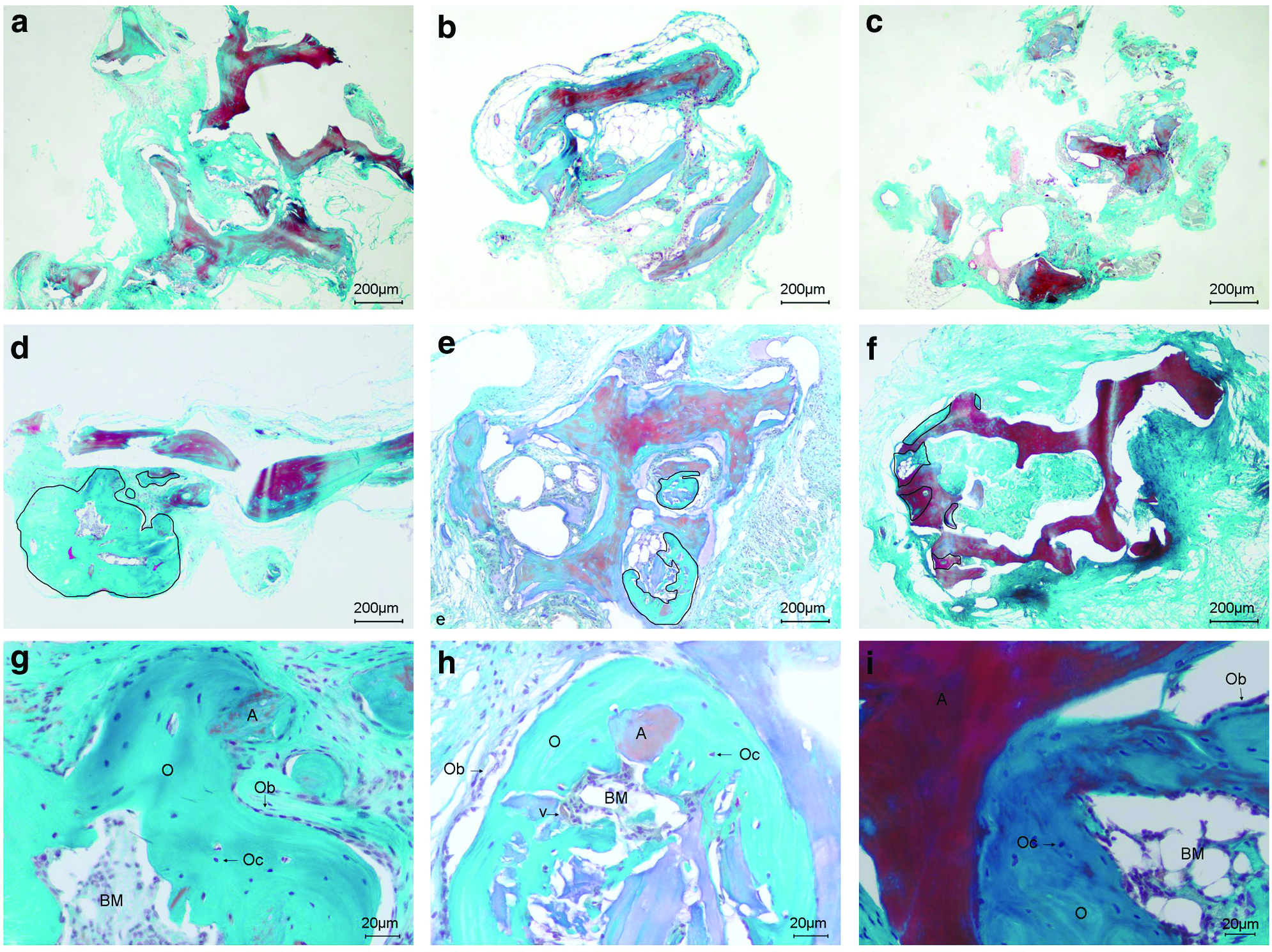
Histological analysis of new bone formation after 7 weeks of ectopic implantation. Scaffolds alone without hMSCs
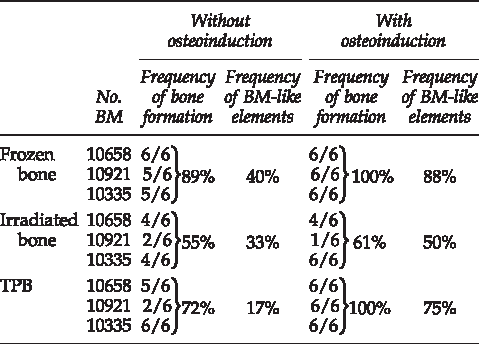
Bone formation occurred significantly more frequently in the frozen bone allografts than in the γ-irradiated bone allografts (89% vs. 55%, p<0.001; Table 3). However, no statistically significant differences in the frequency of bone formation were observed between the TPB allografts and the frozen bone allografts (72% vs. 89%). For scaffold with bone formation, a quantitative analysis was performed. Compared to the total surface bone area, an average of 33.2%±5.6% (mean±SD), 20.8%±8% (mean±SD), and 18.4%±6.8% (mean±SD) of new bone formation was obtained, respectively, with frozen, irradiated, and TBP bone allografts (Fig. 5a). BM-like elements were evident in association with bone formation with a frequency of 40%, 33%, and 17%, respectively (Table 3). All together, our results indicated that frozen bone allograft exhibited the highest frequency and density of new bone formation. However, no significant differences were observed between the TPB and the frozen bone allografts (Fig. 5a; Table 3).

Quantitative analysis of bone formation after 7 weeks of ectopic implantation. New bone formation was assessed on three different samples from each allograft.
The bone formation was assessed by the presence or the absence of new bone area on 15 sections for each scaffold (5 sections at the beginning, 5 in the middle, and 5 at the end). hMSCs from 3 different BMs were tested in sixplicate in six independent animals (n = 18 bone substitute samples per group). For scaffold with bone formation the frequency of BM-like elements was counted on 10 different sections.
BM, bone marrow; hMSCs, human mesenchymal stromal cells.
Impact of the hMSC commitment on bone formation in vivo
To evaluate the impact of the hMSC commitment on bone formation in vivo, we induced strong osteoblastic differentiation in the cells by adding osteoinductor (ascorbic acid, beta-glycerophosphate, and dexamethasone) to the culture medium for 7 days. After osteoinduction, the osteogenic genes Runx2, ALP, BSP, Op, BMP2, and BMP6 were induced between 2- and 12-fold in seeded TPB allografts compared with the levels of expression in uninduced cells. In contrast, osteoinduction had almost no effect on the expression of these genes in hMSCs seeded on frozen or γ-irradiated bone allografts, and only the ALP and BMP6 genes were upregulated. In fact, the Op, BMP2, and BMP4 genes in these cells were downregulated (Fig. 6).

Osteoblastic gene expression of osteoinduced hMSCs cultured on bone substitutes. Osteoblastic (Runx2, ALP, BSP, Op, Oc, On, and BMPs 2, 4, and 6) gene expression is determined by quantitative (TaqMan) RT-PCR on hMSCs cultured in osteogenic (ascorbic acid, β-glycerophosphate, and dexamethasone) platelet lysate (PL) medium on frozen bone, γ-irradiated bone, and PB. Values for all genes were normalized to GAPDH (ΔCt) and to the gene expression for hMSCs 3D cultured in basal PL medium (2−ΔΔCt (fold induction osteoinduced/nonosteoinduced)). hMSCs from nine different BM samples were used for qRT-PCR and each experiment was done in duplicate. Mean data±SEM are presented. */°p<0.05; **/°°p<0.01 (ANOVA test [*] was performed between the three allografts. Mann-Whitney test [°] was performed when osteoinduced cells were compared with nonosteoinduced cells.).
In vivo, when hMSC allografts were osteoinduced for 7 days prior to implantation, the frequency of bone formation increased to 100% for the frozen and TPB bone allografts. (Table 3, Supplementary Fig. S2). In contrast, osteoinduction had no significant effect on the frequency of bone formation in the γ-irradiated allografts (Table 3). A quantitative analysis showed a significant increase of new bone density ratio on frozen and TPB bone allografts compared with the noninduced scaffold (1.6- and 2.49-fold increase, respectively, p≤0.05; Fig. 5b vs. 5a). A higher new bone density ratio was observed in frozen (55%±4.5% [mean±SD]) compared with TPB (45.8%±7.2% [mean±SD]) or irradiated (30.2%±5.5% [mean±SD]) bone allografts (Fig. 5b). Significant differences were observed only between frozen and γ-irradiated allografts. Frequency of BM-like elements was upregulated in all three allografts with a significant increase in TPB allografts (3.7-fold) (Table 3).
Discussion
The combination of hMSCs with biomaterials has previously shown its efficiency for ectopic bone formation, as well as on large bone defects in several experimental animal models.17–22 Whereas coral and ceramic are safe and chemically close to the natural mineral structure of bone, these scaffolds showed variations in their properties of resorption, osteoconduction, and osteoinduction. Moreover, they are very different from the structure of authentic bone and the biomechanical properties of bone allografts are better than all ceramic substitutes. 12 As orthopedists commonly use allograft to provide a template for cellular colonization and tissue repair, demands for bone graft substitutes that are safe, easy to store, and work efficiently have been increasing steadily. Because allograft processing, preservation, and sterilization may reduce its osteoinductive capacity, we compared allografts processed by three different methods: (1) the cryopreserved allograft, (2) the γ-irradiated and cryopreserved allograft, and (3) the solvent-dehydrated and γ-irradiated human TPB allograft. Our results showed that hMSCs can survive on an allograft tissue and that hMSC seeding efficiency was equivalent for the three bone allografts. However, differences were observed in terms of cell metabolism (viability), osteoblastic gene expression, and in vivo bone formation. Our results also showed that the 3D mineral bone substrate had an impact on hMSC osteoblastic commitment. Further, the osteoblastic commitment had an impact on in vivo bone formation.
We found that the late osteoblastic genes Op, BSP, and Oc were upregulated in the 3D-cultured cells at day 7. This gene expression pattern was associated with de novo bone formation as shown in Table 3 and Figure 4. Op is known to be upregulated before BSP and its increased level of expression corresponds to the onset of mineralization.23–26 The higher expression level of Op, compared with that of BSP or Oc, may be interpreted as evidence that these cells were undergoing terminal differentiation. The early genes Runx2, ALP, or On were not upregulated; in fact, ALP was significantly downregulated. However, the basal levels of expression for these genes were already high in the 2D-cultured hMSCs (data not shown). The use of PL in our culture medium may have contributed to these osteoblastic gene expression patterns as we had previously shown that PL can prime hMSC differentiation toward the osteoblastic lineage. 18 The decrease in ALP expression and the high expression of Op may reflect a transitional step from an early immature phenotype to a more differentiated one. In addition, the 3D culture conditions stimulated the expression of the strong osteoinductive agents BMP2 and BMP6.27–31 Secretion of BMP2 and BMP6 by hMSCs could be an advantage for autocrine and paracrine induction of osteoblastic differentiation in vitro and in vivo. 32 The increase in the expression levels of late osteoblastic genes observed in the 3D-cultured cells indicated that the hMSCs had matured into osteoblasts even in the absence of exogenous osteoblastic induction. Our results confirmed previous work showing that ceramic 3D environment induced hMSC osteoblastic differentiation. 26 Moreover, our results indicated that the gene expression patterns induced by 3D culturing alone appeared to be sufficient to support bone formation in vivo.
The chemical composition of the matrix itself may play a role in determining how cells respond as a consequence to their interactions with the extracellular matrix. 32 Frozen and γ-irradiated bone allografts both have protein and lipid molecules in their matrices. In contrast, TPB contains only the mineral part of bone. Unexpectedly, neither freezing nor γ-irradiation nor TPB treatment differentially impacted cell-seeding efficiency. However, after 7 days of in vitro culture, we found that the metabolic activities of the cells seeded on cryopreserved allograft (frozen and γ-irradiated bone) were lower than that of cells cultured on TPB. The storage conditions at −80°C may have modified the proteins and lipids in the scaffold and influenced the cellular responses. 33 Further, the number of viable cells was lower on γ-irradiated allografts than on the frozen bone. This may have been due in part to the lipid peroxidation caused by γ-irradiation. As described by Moreau et al., this treatment causes severe damage to nucleic acids and proteins, impairing their functions and leading to the loss of cell viability. 34 On the other hand, the high metabolism of the cells seeded on TPB after 7 days indicated that lipids and proteins in the matrix were not required for cell survival and/or proliferation.
γ-Iirradiation has been reported to change osteoinductive proteins in the matrix and to impair osteoblastic differentiation.35,36 However, the influence of γ-irradiation on the biological properties of hMSCs is controversial.35–40 In our study, the osteoblastic gene expression of cells cultured on γ-irradiated bone was equivalent to that of cells grown on TPB. As well, the area size of new bone formation is not different either. Considering that TPB does not contain protein, this result supports the hypothesis that γ-irradiation impaired or abolished the activity of osteoinductive proteins in the γ-irradiated and cryopreserved bone allograft.35,36 As osteoblastic gene expression was equivalent for cells in the TPB and γ-irradiated allografts, it was surprising that the frequency of bone formation was significantly lower in γ-irradiated allografts than in the TPB (55% vs. 72%). These differences may be due to the relative numbers of viable cells on the allografts before the implantation, as they were significantly lower on the γ-irradiated scaffolds than on the TPB. This result supports the hypothesis that new bone formation depends on the number of viable MSCs on an allograft.
Osteoblastic gene expression was significantly higher in hMSCs seeded on frozen bone compared with those seeded on TPB. Further, the frequency and the area size of new bone formation and the frequency of bone-marrow-like elements were higher on frozen bone compared with the TPB allografts. However, these results were not significantly different. We concluded that frozen bone partially retains osteoinductive properties, suggesting that proteins in the matrix play a beneficial role by triggering hMSC differentiation toward the osteoblastic lineage. Several residual growth factors can be found within the extracellular bone matrix.41–43 Among them, BMPs could play a role in the formation of new bone in frozen allografts.41,42,44 TPB provides only an osteoconductive scaffold and lacks any osteoinductive proteins. When hMSCs cultured on TPB were fully induced using an exogenous agent, osteoblastic gene expression (Runx2, ALP, BSP, Op, BMP2, and BMP6) was upregulated. This was associated with a higher frequency of bone formation (100%), similar to that of frozen bone. This was also associated with higher area size of new bone formation and higher frequency of bone-marrow-like element. These results support previous work showing that committing the hMSCs toward osteoblastic differentiation before scaffold implantation in mice is important in order to obtain optimal bone formation. 45
Synthetic bone substitutes include ceramics, mixtures of collagen and ceramic, coral derivatives, and bioactive glass. These are potentially attractive alternatives to natural bone, but because of their varied properties of resorption and variations in their osteoconductive, osteoinductive, and mechanical properties, these materials are considered to be less useful than allografts. 12 Moreover, previous in vitro data from Seebach et al. show that TPB produced better overall results than a synthetic scaffold. 46 Also, we showed that TPB allowed osteoblastic bone formation with a frequency, area size, and bone commitment almost equivalent to frozen bone. TPB bone substitute appears to be a good compromise between easy storage, safety, and in vivo bone formation efficiency. It has been shown that adding proteins like VEGF and/or BMP to a scaffold potentiates bone formation. 47 It could be interesting to test the association of TBP with growth factors and cells and observe whether the results are equivalent to the gold standard autologous bone graft.
Conclusions
Our study highlighted differences in the osteoconductive and osteoinductive properties of various bone allograft substitutes currently used in orthopedic applications. Compared with the gold standard allograft (frozen bone), we showed that TPB allowed optimal hMSC seeding and the frequency and the area size of bone formation in vivo were not statistically different. In addition, the frequency of bone formation in vivo was better than that obtained with γ-irradiated allografts. As healthcare teams around the world require bone-grafting scaffolds that are safe and easy to store, the TPB bone substitute appears to be a good compromise between easy storage, safety, and in vivo bone formation efficiency.
Footnotes
Acknowledgments
This work was partially supported by EFS Ile de France, the French Research Ministry and by the 7th Framework Program of the European Commission through the project REBORNE #241879. Laura Coquelin was supported by Tutogen during 6 months. The authors are grateful to the EFS Ile de France which kindly provided the cryopreserved, the γ-irradiated and the Tutoplast®-processed bone allograft. The authors are grateful to Pr. E. Allaire and Dr. M. Gervais for the animal platform facilities. We thank the SEM platform of University Paris-Est, with special thanks to Dr. Patrick Ausset for his precious help. We would like to thank Dr. N. Ortonne for his help on histological analysis and Pr. Luc Douay and Dr. Charles-Henri Flouzat-Lachaniette for their critical review of the manuscript.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.