
Abstract
The bone marrow niche for mesenchymal stem cells (MSCs) contains different amounts of bone and fat that vary with age and certain pathologies. How this dynamic niche environment may affect their differentiation potential and/or healing properties for clinical applications remains unknown, largely due to the lack of physiologically relevant in vitro models. We developed an enabling platform to isolate and study effects of signaling interactions between tissue-scale, laminated hydrogel modules of multiple cell types in tandem. We applied this platform to co- and tri-culture of primary human MSCs, osteoblasts, and adipocytes over 18 days in vitro. Each cell type was analyzed separately with quantitative polymerase chain reaction (qPCR) and histochemistry for several mesenchymal lineage markers. Distinct expression dynamics for osteogenic, adipogenic, chondrogenic, and myogenic transcriptional regulators resulted within each cell type depending on its culture setting. Incorporating this data into multivariate models produced latent identifiers of each emergent cell type dependent on its co- or tri-culture setting. Histological staining showed sustained triglyceride storage in adipocytes regardless of culture condition, but transient alkaline phosphatase activity in both osteoblasts and MSCs. Taken together, our results suggest novel emergent phenotypes for MSCs, osteoblasts, and adipocytes in bone marrow that are dependent on and result in part from paracrine interactions with their neighboring cell types.
Introduction
Toward this end, use of biomaterials may provide a way to recreate these 3D environments, while allowing the study of complex cellular interactions. This includes the application of methods for high-throughput, multivariate analyses of high-content data (e.g., from gene microarrays, suspension arrays, time-of-flight-mass spectrometry, and microscopy images)7–11 that yield system-level information of complex cellular processes at or close to a single-cell level. However, innovative strategies that more closely mimic in vivo microenvironments need to be further coupled with the sophisticated methods outlined above. 12 Moreover, use of primary human cells (rather than immortalized mammalian cell lines), co-culture systems containing two or more cell types to permit better simulation of interactions within realistic microenvironments, and tissue-scale 3D culture systems have the potential to yield further progress toward making regenerative medicine a reality. Therefore, in these studies we employed an extension of novel photopatterning techniques (previously developed in our laboratory 13 ) to generate and assemble 3D laminated hydrogel modules of three different primary human cell types (mesenchymal stem cells [MSCs], osteoblasts, and adipocytes) into millimeter-scale co- and tri-culture constructs.
These cell types reside in close proximity within the same bone marrow niche, motivating their use in this platform as a model of interactions between them. MSCs, which are endowed with the ability to differentiate into many elements of the skeletal system, 14 have been used clinically as cytotherapies both for musculoskeletal and for other disorders. 15 The MSC differentiation programs of adipogenesis and osteogenesis are reciprocally regulated in cultures of MSCs; both master transcriptional regulators RUNX2 and PPARγ are present in low levels in undifferentiated cells, and differentiation toward one lineage completely suppresses genes associated with the other lineage.16,17 This phenomenon has been exploited in past work to evaluate how cell shape, 18 substrate stiffness, 19 and the 3D biomaterial network structure 20 differentially regulate MSC fate under defined media conditions. Differentiation toward either pathway is also regulated by a complex set of paracrine signals16,17 derived from or regulated by cells in the neighboring bone marrow niche environment, including osteoblasts, adipocytes, haematopoietic stem cells, and endothelial cells.16,21 Conditioned media and 2D co-culture studies have provided some insight into how one cell type (osteoblasts or adipocytes) affects the function and differentiation of MSCs.22–24 However, these experiments only model static, one-way interactions and there is currently no systematic means to examine the effects of multidirectional and dynamic crosstalk over time between multiple cell types simultaneously in 3D culture in a way that better mimics interactions that occur in vivo.
To address these limitations, we encapsulated primary human MSCs (hMSCs), osteoblasts, and adipocytes into tissue-scale co- and tri-culture constructs, as described previously. In particular for this study, we employed poly(ethylene glycol)-based hydrogels that maximized diffusion of soluble factors between cell types, and that were cultured in media without exogenously added differentiation cues. We hypothesized that this would enable us to specifically evaluate effects of soluble paracrine signals derived solely from the encapsulated cells. As a proof-of-principle experiment, we evaluated two co-culture configurations (MSCs flanked by adipocytes or osteoblasts on both sides) and a tri-culture configuration (one module each of osteoblasts, MSCs, and adipocytes; see Fig. 1). We hypothesized that each culture environment would uniquely affect the differentiation of encapsulated MSCs and functional responses of osteoblasts and adipocytes as a result of continuous paracrine crosstalk and feedback. After 1, 7, and 18 days in culture, whole constructs were either analyzed using histochemical staining to assess osteogenic or adipogenic differentiation, or sectioned with a scalpel to separate cell populations for quantitative polymerase chain reaction (qPCR) analysis of mRNA expression for genes from several mesenchymal lineages. We further incorporated the relative expression levels of each gene assessed at each time point into a series of multivariate analyses. This provided the means to ascertain the covariance between genes and to determine how the dynamics of these co-variant genes correlate with possible emergent cell phenotypes.
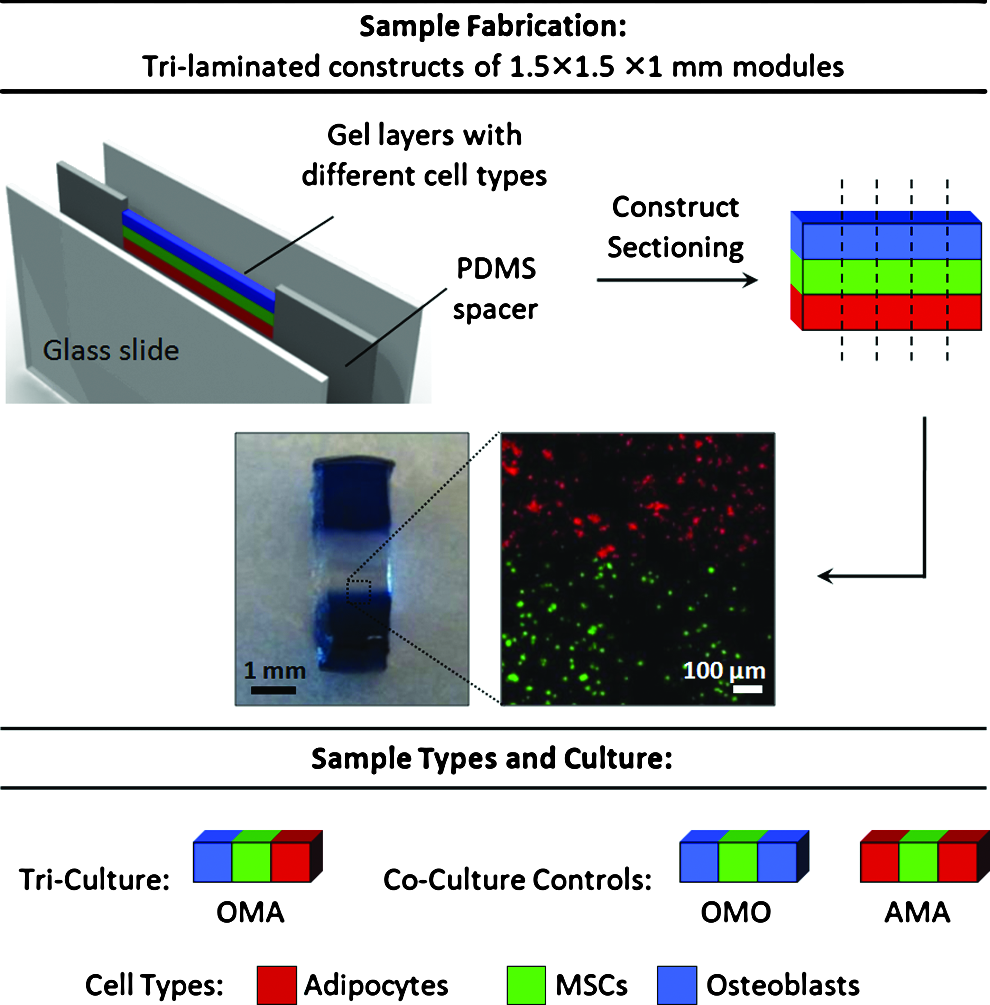
Sample fabrication and study design. Fabrication of co- and tri-culture constructs using the techniques outlined in the Materials and Methods section yields sample sets with well-segregated cell populations [photographs: sample tri-laminated hydrogel construct after reaching equilibrium swelling (left), and confocal image demonstrating human MSCs differentially stained with CellTracker Green (bottom) or Orange (top) segregated at an interface between modules]. Three sample types were examined in this study: MSCs in the center module flanked by only one other cell type [co-culture controls: osteoblasts (OMO) or adipocytes (AMA)] or by both cell types (tri-culture; OMA). MSCs, mesenchymal stem cells. Color images available online at www.liebertonline.com/tea
Materials and Methods
Polymer synthesis and characterization
All chemicals were purchased from Sigma-Aldrich unless otherwise noted. Poly(ethylene glycol)-diacrylate (PEG-DA) was prepared as previously described 25 from PEG (Mn=3400 Da). The resultant polymer had a molecular weight Mn=3676±16 Da with a polydispersity of 1.088±0.015 as determined by gel permeation chromatography (Shimadzu), and the presence of conjugated acrylate groups was verified with 1H-NMR.
To allow presentation of adhesive ligands that promote viability of encapsulated cells, fibronectin-derived GRGDS (PeproTech) and laminin-derived YIGSR (Anaspec) adhesion peptides were conjugated to a 3400-Da-molecular-weight Acryl-PEG-succinimidyl valerate spacer (Acryl-PEG-SVA; Laysan Bio) in NaHCO3 buffer (pH 8.5) according to previous protocols, 26 dialyzed (1000 Da MW cutoff), lyophilized, and stored at −20°C until further use.
Cell culture and expansion
All cell culture reagents were obtained from Mediatech unless otherwise specified. Primary hMSCs were obtained from the laboratory of Dr. Darwin Prockop (Texas A&M Health Sciences Center) and expanded according to recommended protocols in minimal essential medium-alpha (αMEM) with 16.5% fetal bovine serum (FBS; Hyclone), 1 g/L glucose, 2 mM L-glutamine, 1% amphotericin B, and 0.1% gentamicin and cultured at 37°C with 5% CO2 in a humidified incubator. 27 Primary human osteoblasts (hObs; Lonza) were expanded to seven or eight population doublings according to the manufacturer's protocol in Dulbecco's modified Eagle's medium (DMEM) with 10% FBS, 1 g/L glucose, 1% amphotericin B, and 0.1% gentamicin. Primary human subcutaneous preadipocytes (Lonza) were expanded to three to four population doublings according to the manufacturer's protocol in DMEM with 10% FBS, 4.5 g/L glucose, 2 mM L-glutamine, 1% amphotericin B, and 0.1% gentamicin. Cultures at 80% confluence were differentiated into human adipocytes (hAds) for 2 weeks in expansion medium with 60 μM indomethacin, 0.5 mM 3-isobutyl-1-methylxanthine (IBMX), 0.5 μM dexamethasone, and 1 μM insulin.
Construct fabrication and long-term 3D co- and tri-culture
Layering devices were fabricated and employed for cell patterning as described in Figure 1. Briefly, 1-mm-thick spacers were cut from cured polydimethylsiloxane (PDMS; Sylgard 184; Dow Corning) and contact-bonded on each side to glass slides (Corning). Each spacer contained a cavity for polymer solution/gels as they were loaded and crosslinked. Assembled devices were sterilized by autoclave prior to use for encapsulation.
Hydrogel precursor solutions were formulated with 10% w/w PEG-DA in phosphate-buffered saline (PBS) containing 0.05% w/w D-2959 photoinitiator (Ciba) and 1 mM Acryl-PEG-GRGDS (for hMSCs) or Acryl-PEG-YIGSR (for hAds). Cell suspensions were prepared from near-confluent cultures using 0.05% trypsin/0.53 mM EDTA and resuspended in their respective gel precursor solutions at a concentration of 15 million cells/mL. These solutions were loaded into layering devices and sequentially photocrosslinked into laminated 1-mm-thick, 1.5-mm-tall hydrogel strips (Fig. 1). After each patterning step, residual noncrosslinked material was rinsed out of the device with fresh 10% w/w PEG-DA solution using a syringe. 13 An opaque photomask was used in subsequent steps to prevent any further UV exposure and crosslinking of the existing gels. Single, laminated constructs were extracted from the device and sectioned with a scalpel perpendicular to the long axis of the laminate to yield twenty-one 1.5-mm-wide co- and tri-culture constructs (Fig. 1).
Co-culture constructs consisted of hMSCs flanked on both sides by human osteoblasts (hObs; OMO) or hAds (AMA), while tri-culture constructs consisted of hMSCs flanked on one side by hObs and hAds on the other (OMA). Constructs were placed in separate wells of six-well tissue culture plates with 4 mL of co-culture medium (DMEM with 10% FBS, 50 μg/mL L-ascorbate-2-phosphate [Sigma], 1 μM insulin, 1% amphotericin B, and 0.1% gentamicin; replenished every 2 days) designed by our lab to maintain each cell type in culture while eliminating traditional exogenous differentiation cues. Constructs containing only hMSCs in each module were fabricated and cultured in osteogenic or adipogenic differentiation medium for 21 days as a positive control to verify that differentiation was possible in this culture system under a standard set of cues. Osteogenic differentiation medium consisted of αMEM with 10% FBS, 50 μM L-ascorbate-2-phosphate, 20 mM β-glycerophosphate (Sigma), 10 nM dexamethasone, 1% amphotericin B, and 0.1% gentamicin. Adipogenic differentiation medium consisted of αMEM with 10% FBS, 0.5 mM IBMX, 60 μM indomethacin, 1 μM insulin, 0.5 μM dexamethasone, 1% amphotericin B, and 0.1% gentamicin.
RNA isolation and qPCR
Hydrogel constructs were rinsed in PBS and blocks containing individual cell populations were separated from each other using a scalpel for gene expression analysis by qPCR after 1, 7, and 18 days in co- or tri-culture. Gel blocks containing the same cell type were pooled from three constructs of the same culture condition to provide sufficient copies of mRNA for quantification. These blocks were homogenized in microcentrifuge tubes with pellet grinders, after which mRNA was extracted using a QIAshredder tissue homogenizer and RNeasy kit with DNase I digestion (Qiagen). cDNA was generated using SuperScript III Reverse Transcriptase (Invitrogen) with Oligo(dT)15 primers and dNTPs (Promega). Gene expression of each cell type was analyzed for target mesenchymal lineage genes using custom-designed primers (Table 1) with quantitative PCR amplification performed on a StepOnePlus™ Real-Time PCR System (Applied Biosystems) in the presence of SYBR Green/ROX master mix (Applied Biosystems). RPS18 and ACTB were both used as endogenous controls for normalization through geometric averaging,
28
and relative expression (n=4 per culture type and time point) of each target gene was calculated using the formula:
RUNX2, runt-related transcription factor 2; OCN, osteocalcin; PPARG2, peroxisome proliferator-activated receptor γ, isoform 2; LEP, leptin; SOX9, SRY (sex determining region Y)-box 9; MYOD, myogenic differentiation 1; RPS18, ribosomal protein 18S; ACTB, β-actin.
where Ct represents the cycle threshold for amplication. Endogenous controls were evaluated in each cell type to ensure that their expression levels were not altered across time or culture conditions.29,30
Histological analysis for differentiation
Following co- or tri-culture, whole constructs (n=1–2 per culture type and time point; same time points as previously described) were collected for histological staining. Constructs were rinsed in PBS and infiltrated by graded concentrations of sucrose in PBS followed by graded concentrations of optimal cutting temperature compound (OCT; Sakura Finetek) using a technique adapted from the literature, 31 and individually embedded in OCT, flash-frozen in liquid N2, and stored at −80°C until sectioning. Embedded constructs were serially sectioned into 20-μm-thick slices (Microm HM 560 Cryostat; Thermo Scientific), mounted on Superfrost Plus slides (Fisher), and stored at −80°C until staining. Adipogenic differentiation was assessed with an Oil Red O stain for triglyceride accumulation in intracellular storage vesicles using standard protocols 27 and visualized with brightfield microscopy. Osteogenic differentiation was examined by assessing in situ alkaline phosphatase activity (Vector® Red Alkaline Phosphatase Substrate Kit; Vector Labs). Briefly, unfixed sections were reacted with an alkaline phosphatase substrate in 100 mM Tris buffer (pH 8.5) according to the manufacturer's protocol and counterstained with Hoechst 33258 (0.25 μg/mL in PBS for 5 min; Molecular Probes), and visualized with epifluorescence microscopy under Texas Red and DAPI filters, respectively.
Statistical analysis and multivariate modeling
Gene expression results are depicted as mean±standard deviation unless otherwise noted. Prior to statistical analysis, all data were transformed with a Box-Cox transformation. Data were analyzed by multifactor analysis of variance (ANOVA) to identify significant factors (cell type, culture type, and day) and factor interactions (cell type*day and culture type*day) for each target gene assayed. Where significance factors and interactions were identified by ANOVA, Tukey's post hoc test (significance level p<0.05) was used to determine significant differences between individual sample groups.
Multivariate statistical modeling was performed with the overall goal of extracting combinations of time-variant gene expression markers that were the most informative for distinguishing differences among cell types and co- or tri-culture conditions. All Box-Cox-transformed data were mean-centered and scaled to unit variance prior to analysis as a means of normalization to allow all variables to be considered equally scaled in the principal components (PCs) or latent variables.
9
For these analyses, the data set (total of 504 data points) was organized into an N×K matrix
To optimize the quality of PCA and PLS-DA models, several pruning procedures were performed to remove outlying observations (outside 95% confidence interval) and noninfluential variables (weight ∼0 in both components) and enable statistical significance testing of the model and the variables used to generate it.32,34 The quality of each model was summarized by two nondimensional statistical parameters: (1) R2X (for PCA) or R2Y (for PLS-DA), which quantitatively measures the extent to which the model explains the variation in the data matrices and dictates a goodness of fit, and (2) Q2, which quantitatively measures the extent to which the variation of a future experimental data set may be predicted by the model (goodness of prediction). 34 Both of these parameters are analogous to regression statistics, with a value ranging from 0 (poor) to 1 (perfect) fit or predictive capability. The appropriate number of PCs or latent variables was determined by cross-validation.32,34 The results of this procedure were fed into a jack-knifing analysis 32 to calculate the standard errors of the regression coefficients (weights), which were then converted into 95% confidence intervals via the t-distribution to determine which X-variables (genes) of high weight have a statistically significant influence (p<0.05) on each class of observations (Y-variables) in each of the PLS-DA models generated (Figs. 5D and 6D, F).
Results
Gene expression dynamics and histochemical staining
We demonstrate that each of the examined mesenchymal lineage genes (Table 1) exhibited expression dynamics in each cell type with time that appeared to vary depending on the co- or tri-culture condition employed (Figs. 2–4). Certain genes were often uniquely variable only within a particular cell type, with MSCs exhibiting time-variant changes in most of the genes examined (Fig. 4), while adipocytes (Fig. 2) and osteoblasts (Fig. 3) exhibited changes in expression among only two or three of the genes evaluated.
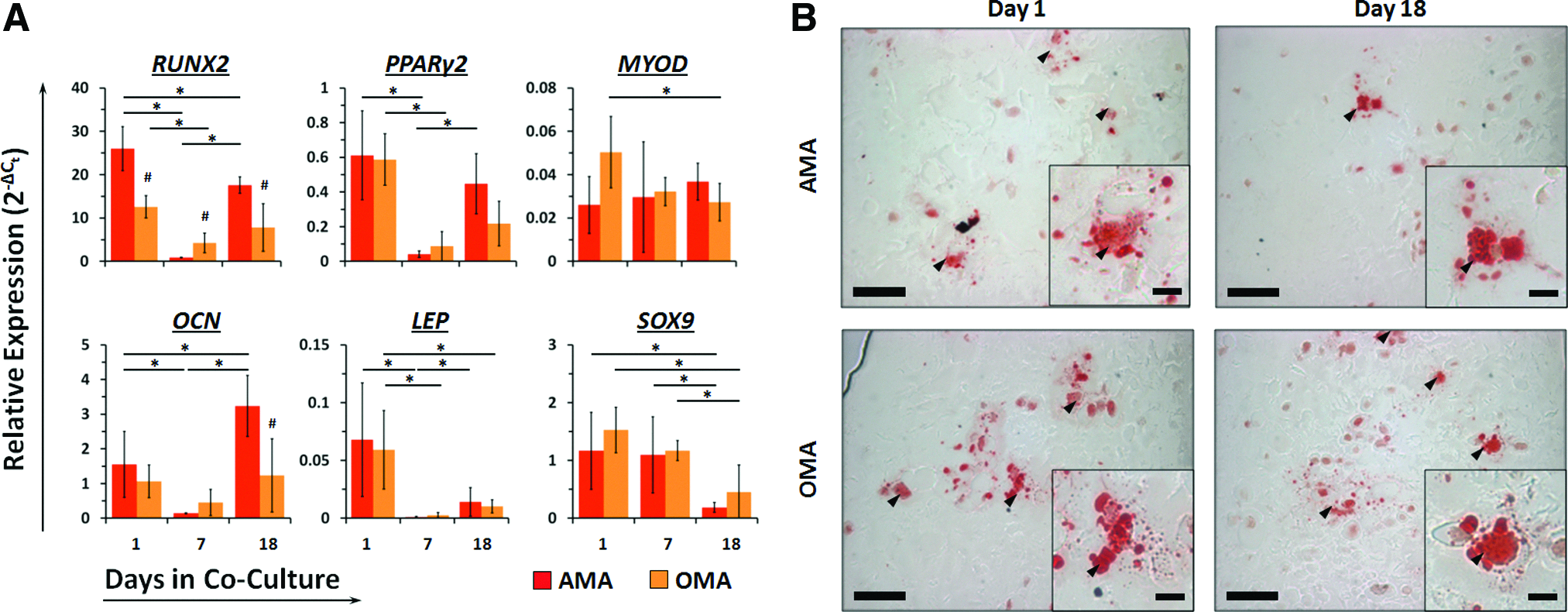
Co-culture and tri-culture differentially affect expression dynamics of osteoblastic (RUNX2 and Osteocalcin) and adipocytic (PPARγ2 and Leptin) genes in adipocytes but does not affect triglyceride storage with time.

Co-culture and tri-culture differentially affect expression dynamics of osteoblastic (RUNX2 and Osteocalcin) and chondrogenic (SOX9) genes in osteoblasts, in addition to persistence of alkaline phosphatase activity over time.
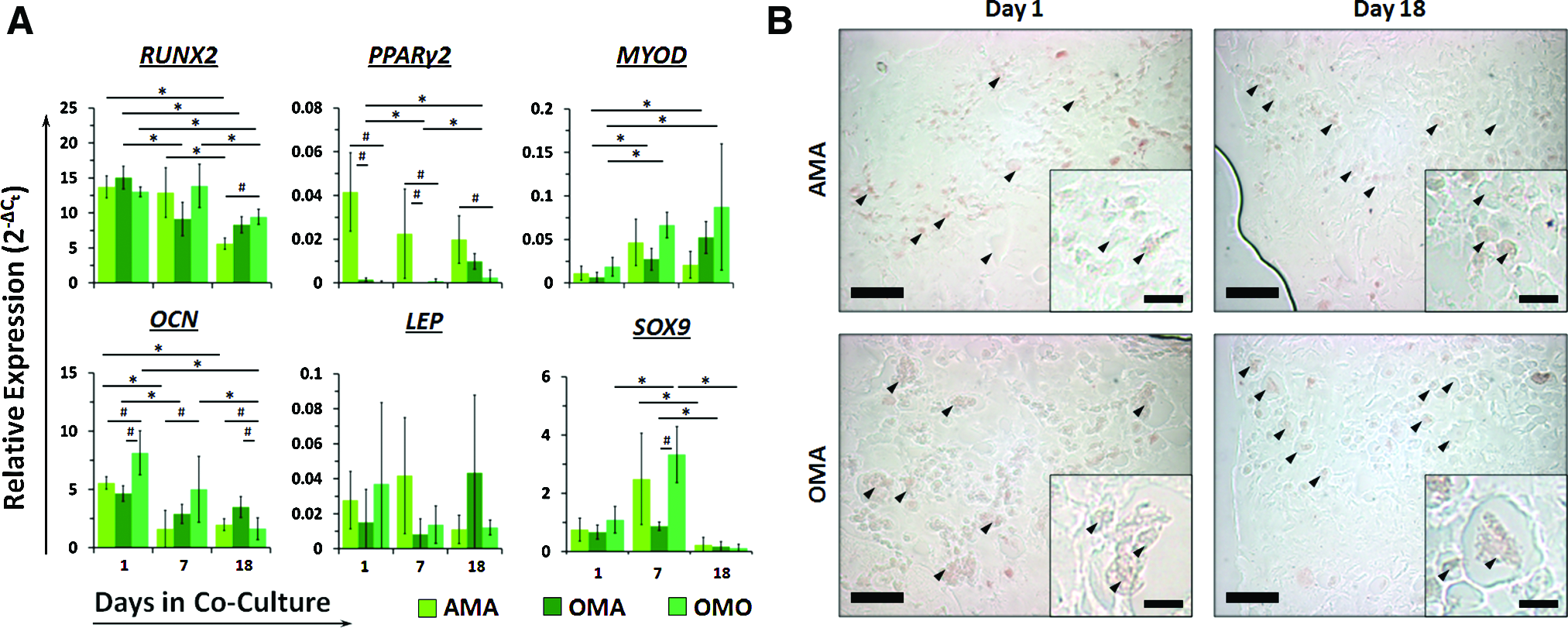
Co- and tri-culture differentially affect expression levels and dynamics of several lineage-specific transcription factors (but not terminal differentiation markers) in MSCs, while only causing scant and transient alkaline phosphatase expression in MSCs from osteoblast-containing cultures.
Adipocytes co-cultured only with MSCs (AMA configuration) exhibited significant decreases in expression of RUNX2 (29-fold), PPARγ2 (15-fold), OCN (11-fold), and LEP (64-fold) at day 7 (Fig. 2A). Relative to day 1, RUNX2 expression at day 18 was still significantly lower (1.5-fold), OCN expression was significantly higher (2.1-fold), and PPARγ2 and LEP were unchanged. Adipocytes from tri-culture (OMA) showed a significant decrease in expression of RUNX2 (3.0-fold), PPARγ2 (6.8-fold), and LEP (22-fold) at day 7 that persisted through day 18. Relative to AMA adipocytes, expression of RUNX2 was significantly lower at each time point and OCN was expressed in higher quantities at day 18. Compared with the osteoblastic and adipocytic genes, expression of MYOD and SOX9 remained relatively stable in each cell type over time. Over the entire culture period, examination of Oil Red O staining revealed no qualitative differences in triglyceride storage vesicles in adipocytes from either culture condition (Fig. 2B).
Osteoblasts co-cultured only with MSCs (OMO) exhibited significant decreases in expression of RUNX2 (2.0-fold), OCN (2.1-fold), and SOX9 (7.9-fold) at day 7, and RUNX2 decreased further by day 18 (2.7-fold relative to day 1; Fig. 3A). Osteoblasts from tri-culture (OMA) also exhibited significant progressive declines in RUNX2 (1.6- and 3.8-fold at day 7 and day 18, respectively, relative to day 1) and OCN (1.5- and 2.8-fold) through day 18. While RUNX2 and OCN expression was higher in OMA than OMO osteoblasts at day 7, both genes exhibited the same expression levels in each culture setting by day 18. Expression of SOX9 in OMA osteoblasts, while much lower than OMO osteoblasts on day 1, remained the same by day 7 and significantly decreased (48-fold) by day 18. Alkaline phosphatase activity appeared similar in osteoblasts from both culture conditions on day 1 (Fig. 3B). This activity persisted longer in osteoblasts from OMO than OMA constructs by day 7. By day 18, no alkaline phosphatase activity could be observed in either of the sample types (Supplementary Fig. S1; Supplementary Data are available online at www.liebertonline.com/tea).
Lineage markers in MSCs exhibited much more diverse and complex expression dynamics dependent on their co- or tri-culture setting (Fig. 4A). RUNX2 expression levels persisted through day 7 in MSCs from co-culture settings while declining significantly in tri-cultured MSCs by day 7 (1.6-fold). By day 18, RUNX2 expression was graded in MSCs depending on the relative amount of osteoblasts present in the co- or tri-culture construct. Conversely, PPARγ2 expression was dramatically higher in MSCs co-cultured with adipocytes (AMA) as early as day 1 and persisted throughout the culture period, while OMO MSCs exhibited consistently low levels that did not change. Tri-cultured MSCs (OMA) exhibited a 14-fold reduction in PPARγ2 expression at day 7, but this was upregulated 6.6-fold relative to day 1 after 18 days to an intermediate level between AMA and OMO MSCs. Expression of MYOD increased only in MSCs from OMO and OMA culture conditions by day 7 (3.6- and 4.1-fold, respectively), and was sustained through 18 days in culture. SOX9 expression was only upregulated in MSCs co-cultured with osteoblasts at day 7 (3.0-fold) before declining to day 1 levels. Genetic markers of terminal differentiation toward osteoblasts (OCN) and adipocytes (LEP) were not significantly upregulated over time in MSCs from any of the co- or tri-culture conditions tested, and this was reflected in our histological analysis. No triglyceride storage occurred in MSCs from any culture condition over the entire length of the co-culture period (Fig. 4B). Transiently increased, relatively low alkaline phosphatase activity was observed only in a small number of MSCs from constructs containing osteoblasts (OMO and OMA, day 7), which was absent on day 18 (Fig. 4C and Supplementary Fig. S2). Positive control gels containing only MSCs and cultured in osteogenic or adipogenic medium exhibited visible alkaline phosphatase activity and triglyceride storage, respectively (Supplementary Fig. S3).
Multivariate modeling with PCA
Given the seemingly complex gene expression responses among different cell types placed under different co- and tri-culture conditions, we thought that taking a more global view of these factors using multivariate modeling would provide additional information; from this analysis, one could potentially extract and examine clusters of cell types that exhibit similar responses in gene expression to their culture conditions based on covariance of more than one gene. Therefore, we performed PCA to reduce the dimensionality of all the summarized gene expression data based on the sources of maximum variance (Fig. 5A). Plotting samples in the first two PCs, which captured ∼69% of the variability in the data set, indicated that MSCs and osteoblasts cluster apart from adipocytes (first PC; 51.1% of data set variability). The second PC captured an additional 17.8% of the variance in the data and separated MSCs co-cultured with adipocytes (AMA) from osteoblasts and MSCs from other culture conditions (OMO and OMA). Plotting the variable loadings in the first two PCs indicated that each of the genes evaluated was highly influential in the model at one or more time points since each has large weight on one or both PCs (Fig. 5B; variables with 0 weight were removed during pruning). In distinguishing the different clusters of observations from the score plot, adipocytes from OMA and AMA constructs were most distinguishable by PPARγ2 expression, MSCs from AMA constructs were most correlated with high PPARγ2 and LEP expression, and osteoblasts and MSCs from OMO and OMA constructs were highly correlated with RUNX2 and OCN expression.
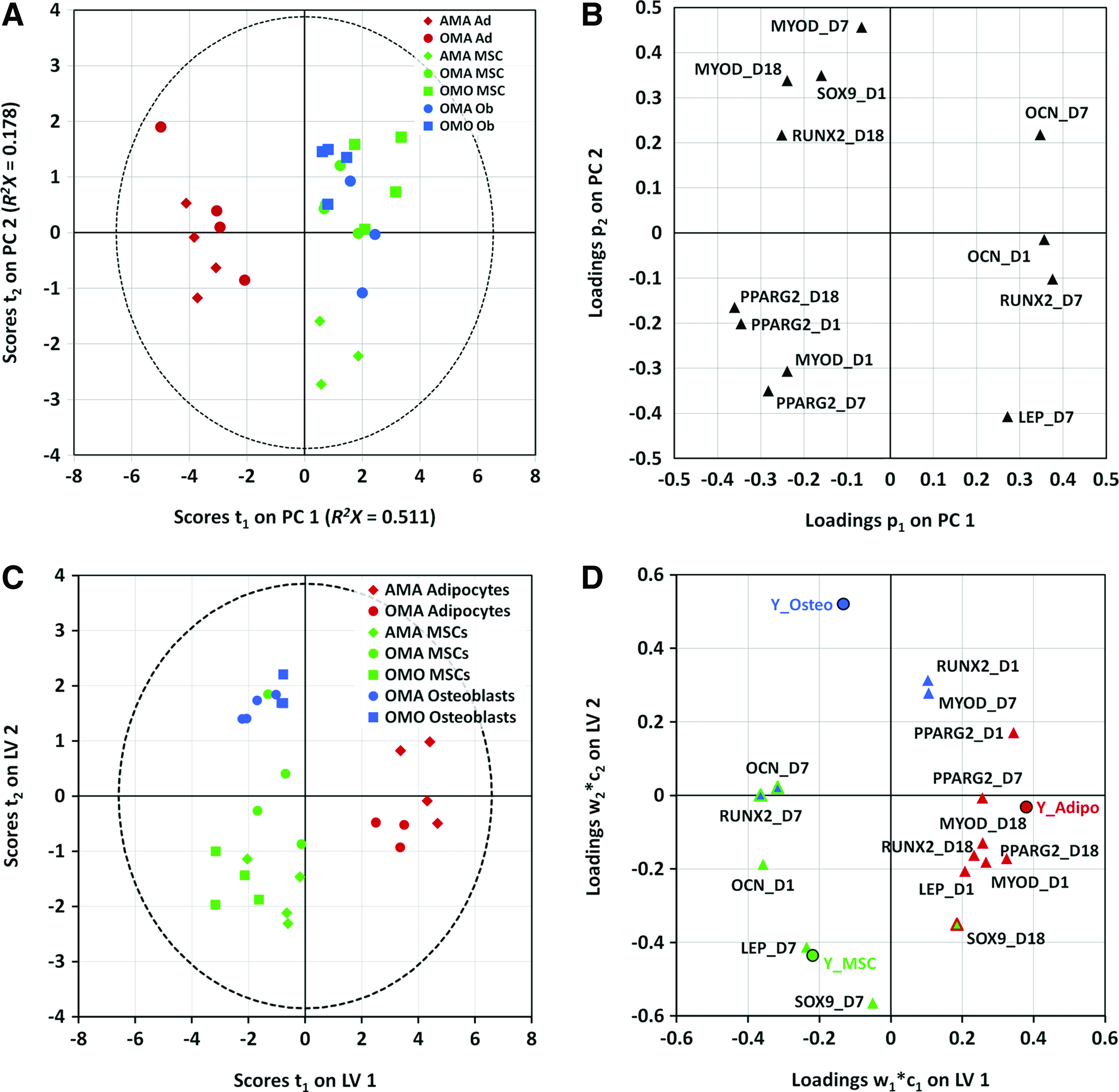
Statistical modeling based on covariance of the expression of several mesenchymal lineage genes yields two latent variables that are able to distinguish MSCs from adipocytes and osteoblasts, respectively, and elucidates the correlation structure of the gene expression at various time points with each cell type present.
Multivariate modeling with PLS-DA
Several clusters of data by cell type and/or culture type were discernible by the PCA results, though they overlapped to an extent. These findings motivated further supervised analysis to deconvolve this complex data set into a meaningful set of variables that adequately describe the patterns of samples and their gene covariance in the overall data set. We first classified the observations into three groups by cell type and generated a two-latent variable PLS-DA model with quality parameters R2Y=0.75 and Q2=0.681. The model distinctly classified the scores with the first latent variable describing the differences of adipocytes from the other cell types present, and the second latent variable describing the differences between MSCs and osteoblasts (Fig. 5C). Osteoblasts and adipocytes constitute smaller clusters on this score plot, while MSCs constitute a much larger cluster, owing to differences in heterogeneity between the three cell types. With respect to the gene expression dynamics that constitute the latent variables, several genes from different mesenchymal lineages overlap in their contribution to discrimination of the different cell types. This can be visualized by their proximities to one another on the weight plots in the latent variable space (Fig. 5D). PPARγ2 (days 1, 7, and 18), MYOD (days 1 and 18), LEP (day 1), and RUNX2 (day 18) significantly correlated with adipocytes; SOX9 (day 7), LEP (day 7), and OCN (day 1) significantly correlated with MSCs; and RUNX2 (day 1) and MYOD (day 7) significantly correlated with osteoblasts. In addition, SOX9 (day 18) is significantly correlated with both MSCs and adipocytes, and RUNX2 (day 7) and OCN (day 7) are significantly correlated with both MSCs and osteoblasts.
Within the larger clusters of cell types in our first PLS-DA model (Fig. 5C), we noticed that several observations appeared to also cluster by co- or tri-culture setting. After generating several preliminary PCA models, we were able to verify that much of the variation within these clusters (>80%) could be accounted for in PCs that separated the observations by co- or tri-culture conditions (Supplementary Fig. S4). This motivated the development of a separate PLS-DA model for each cell type (Fig. 6) to further classify the observations by culture condition and determine gene expression variables at specific times that are highly determinative of cells from each condition. A single-latent variable model of adipocytes revealed that expression dynamics of adipocytic (PPARγ2), osteoblastic (RUNX2 and OCN), and myogenic (MYOD) genes are important for discriminating the cell populations from AMA and OMA culture conditions (Fig. 6A, D). A single-latent variable model of osteoblasts discriminates OMO and OMA populations largely by their expression dynamics of osteogenic (RUNX2 and OCN) and chondrogenic (SOX9) markers (Fig. 6B, E). Further, a two-latent variable model of MSCs discriminates cell populations from all three culture conditions (Fig. 6C, F) on the basis of several mesenchymal lineage markers unique to each population. AMA MSCs are most significantly correlated with expression of adipogenic marker PPARγ2 on days 1 and 7, while OMO MSCs are characterized by osteogenic markers RUNX2 (day 18) and OCN (day 1). Interestingly, we observed that unlike co-cultured MSCs, tri-cultured MSCs (OMA) correlated strongly with osteogenic (RUNX2, day 1; OCN, day 18), adipogenic (LEP, day 18), and myogenic (MYOD, days 7 and 18) markers.
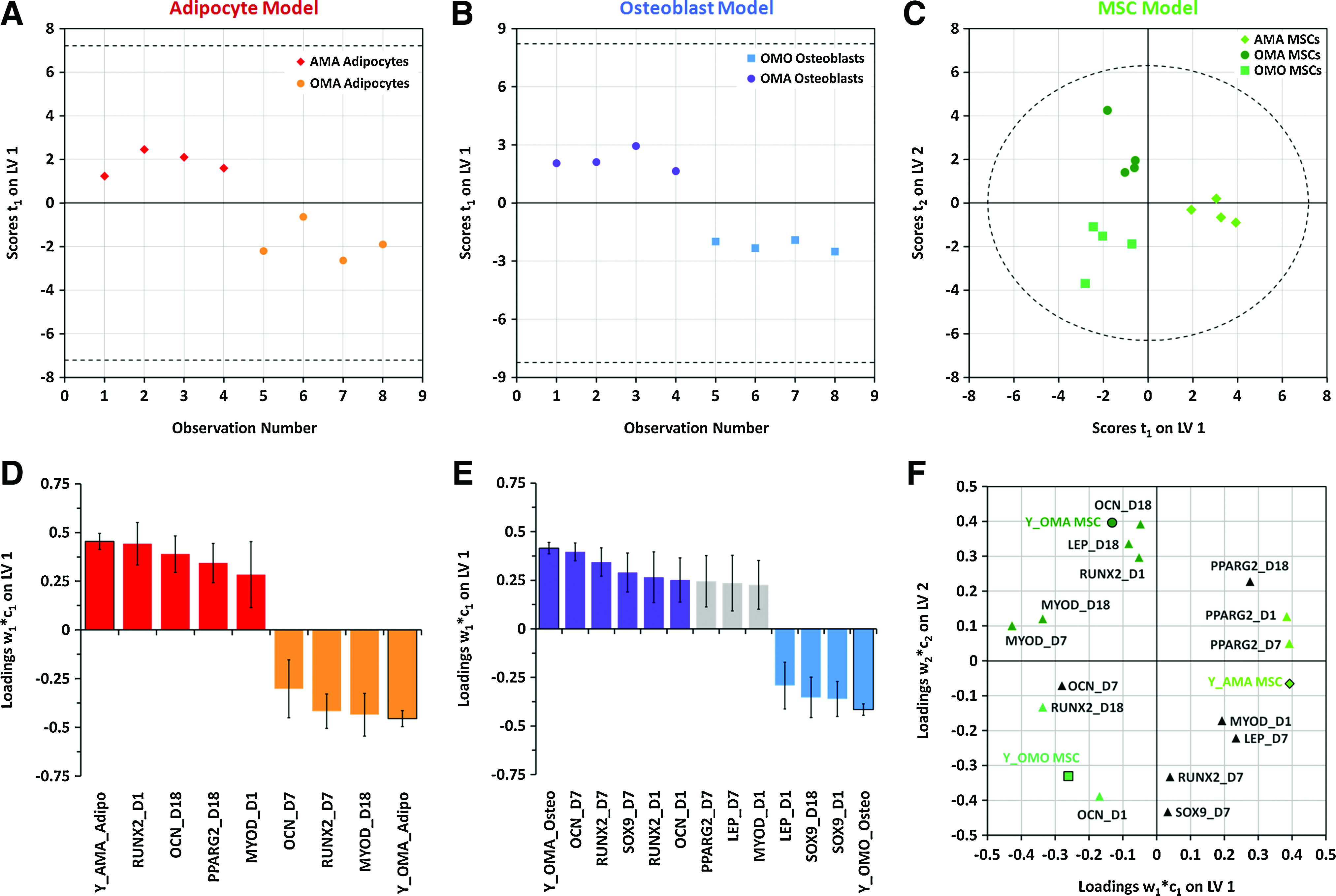
PLS-DA models of single cell types can robustly separate cell samples derived from different culture conditions and describe the important gene expression variables that correlate with each response to co- or tri-culture.
Discussion
This work presents a simple platform for patterning multiple types of cells into tissue-scale 3D hydrogel constructs that are permissive for multidirectional paracrine signal communication and produce distinct responses in each cell type dependent on the co- or tri-culture environment surrounding it. Devices used to generate these platforms (Fig. 1) are easily constructed from inexpensive, sterilizable, and reusable materials, making them appropriate and readily adaptable for use in any laboratory environment. The size of the PDMS cavity can be readily adapted to contain modules of different sizes and thicknesses with some limitations depending on the materials used (e.g., light penetration through the entire thickness and efficiency of the crosslinking reaction). 13 The modularity of this hydrogel system—enabled by the sequential crosslinking and lamination of these synthetic, chemically tailorable hydrogels—allows for flexibility in the configuration of patterned cells and also provides a means for tailoring the microenvironmental niche of each cell type (e.g., biomaterials, material stiffness, biochemical moieties, and cell density) independently.2,35 This feature stems from the ability to use a mask for preventing further crosslinking, UV exposure of cells, or other modification of a gel module after each step.
The data acquired from these proof-of-principle experiments suggest that the differential effects of each co- and tri-culture environment on each cell type can result from paracrine signaling that occurs between each cell type in the absence of proliferation, migration, cell spreading, and direct cell–cell contact. Our hydrogel platform is specifically designed to isolate these effects due to its sufficiently small mesh size to prevent cell migration and proliferation while allowing the diffusion of soluble cues from the small molecule to protein scale. 36 While cell spreading is permissive and promotes osteogenesis, 18 we chose to decouple the effects of cell spreading from soluble signals in these experiments to isolate the effects of signaling crosstalk alone.
Fibronectin- and laminin-derived peptides (RGDS and YIGSR, respectively) were employed here to promote cell viability of encapsulated hMSCs and adipocytes during the culture as this may be adhesion dependent.37,38 During co- and tri-culture, cells are coupled via these peptides to the polymer network of the hydrogel and may additionally deposit extracellular matrix (ECM). Consequently, we cannot definitively rule out the possibility of intercellular communication via mechanical coupling between the cells and the polymer/ECM. However, all of the cells sense the same bulk mechanical properties of the gel modules since they are composed of the same gel material. The encapsulated cells are likely unable to generate large traction forces (due to lack of cell spreading),18,39 and mechanical signals would have to propagate over relatively long distances for cells from different modules to communicate. Further, the synthetic polymer matrix of high crosslink density used here cannot be remodeled to aid in mechanical signal propagation (e.g., compared with collagen, 40 hyaluronic acid, 20 and ionically crosslinked alginate 19 ), and much of the matrix deposition in these synthetic, nondegradable gels likely remains confined to a pericellular location. 26 Together, these considerations support the likelihood that much of the intercellular crosstalk in this system is dominated by soluble paracrine factors.
Traditional mono- and co-culture experiments with MSCs and their differentiated counterparts use exogenous factors to drive and maintain their differentiation and terminal function, 27 or employ a mixture of differentiation media as a method for providing a permissive environment for differentiation while minimizing bias.19,20 We formulated our co-culture medium without any exogenous differentiation cues, relying instead on the cells themselves to produce the soluble signals necessary to drive each other's responses. While terminal differentiation was not observed, environment-dependent responses occurred in each cell type. This suggests that the encapsulated cells are both producing and responding to soluble signals during the 18-day culture period evaluated in this study. Although these signals were not directly measured in these experiments, characterizing their roles will be included as a part of future studies with this platform.
As a direct consequence of being able to readily separate each cell type after the co- or tri-culture period by simple gel sectioning, we were able to observe the effects of paracrine crosstalk on each cell type independently at the individual gene level (Figs. 2–4 and Supplementary Fig. S1–S2) and more globally through multivariate analysis (Figs. 5–6 and Supplementary Fig. S4). While controls containing only one encapsulated cell type were not examined in this study, our analysis does demonstrate gene expression dynamics and potential histological changes that vary with co- and tri-culture conditions and are independent of the effects of time in culture. While these culture-dependent effects could be inferred from close examination of the data on a gene-by-gene basis (Figs. 2–4), we were able to leverage the power of multivariate modeling to examine how distinct our observations were from each other given the covariance that existed across the multiple genes we examined. Our high-quality PLS models (R2≥0.75 and Q 2 ≥0.68) 32 confirmed many of the findings present in our preliminary inspection of the data, they are as follows: (1) adipocytes could be clearly distinguished from the other cell types, (2) AMA MSCs could be represented as a distinct cluster from other MSCs, (3) similarities in osteoblasts and MSCs from OMO and OMA samples could be mapped, and (4) a reduced set of genes that dynamically contribute to different emerging phenotypes of each cell type from the different co- and tri-culture conditions could be clearly distinguished.
Our analysis showed that adipocytes in co- and tri-culture differentially expressed the adipogenic genes PPARγ2 and LEP and (surprisingly) osteogenic genes RUNX2 and OCN, though this did not appear to affect their triglyceride storage (Figs. 2 and 6D). The presence of MSCs (with or without adipocytes) affected osteoblastic (RUNX2 and OCN) and chondrogenic (SOX9) markers in osteoblasts (Figs. 3 and 6B, E). Interestingly, alkaline phosphatase activity appeared to persist longer in co-cultured than in tri-cultured osteoblasts (Fig. 3B). Taken together, our results suggest that adipocytes and osteoblasts in co- and tri-cultures maintain some of their major functions or phenotypic markers. However, there may be subtle and previously unreported differences in the functional capacities of adipocytes and osteoblasts depending on neighboring cell types, based on the differential expression of genes regulating each lineage. Conversely, these findings may suggest that the culture environment (soluble cues, the presence of MSCs, or the hydrogel scaffold) or the preculture of primary cells may need further optimization to promote sustained functions associated with terminal differentiation. These potentially novel phenotypes and their determinants, including intercellular signals and potential artifacts of the in vitro microenvironment, could be more closely evaluated further in future studies.
The analytical techniques applied here were particularly useful in clearly discerning population-level differences among MSCs from different co- and tri-culture settings. We selected transcription factors for multiple MSC lineages, since their plasticity and level of lineage commitment are attributable to the simultaneous expression of these markers in their undifferentiated state that are progressively downregulated as the cells commit. 41 All lineage-specific transcription factors in these cells were affected to some extent by the co- and tri-culture settings. For example, AMA MSCs, the most distinct population of MSCs (Fig. 5A), were most highly correlated with increased expression levels of PPARγ2 (Figs. 4A and 6F) and were the only MSCs that failed to exhibit alkaline phosphatase activity during the co-culture period (Supplementary Fig. S2). Conversely, OMO MSCs correlated with higher expression levels of RUNX2 and OCN (Figs. 4A and 6F), clustered with osteoblasts (PCA; Fig. 5A, B), and appeared to exhibit alkaline phosphatase expression in some cells at day 7 (Fig. 4B). In contrast, MSCs cultured alone in the same hydrogel formulation, but using standard differentiation media, exhibited marked histological signs of differentiation (Supplementary Fig. S3). This finding suggests that the lack of significant staining in co- or tri-culture for either differentiation marker in MSCs (as well as other cell types) may not be an artifact of the culture system, but rather a sign that the signaling cues from neighboring cell types are not sufficient to induce terminal differentiation. Taken together, this suggests that co-cultured MSCs, while not terminally differentiated, may be biased toward differentiation toward the cell type with which they were co-cultured, in agreement with previously published data from other groups.22–24 Tri-cultured (OMA) MSCs, while closely related to osteoblasts and OMO MSCs in their gene expression and apparent alkaline phosphatase expression (Figs. 4A, B and 5A, B), correlated with a broader set of mesenchymal lineage genes, including MYOD (Fig. 6F). This might suggest a much more heterogeneous population of cells that emerged from tri-culture or a different bias in lineage commitment, each of which merit further study given the paucity of literature describing the combined effects of osteoblast- and adipocyte-derived soluble signals on MSCs. Additionally, this validates the relevance of this tri-culture system for future efforts to re-capitulate the bone marrow niche and to model bone marrow pathologies. 16
Conclusion
In this study, we develop a simply fabricated, photopatternable 3D culture system that enabled observation of distinct gene expression dynamics resulting from dynamic paracrine interactions between MSCs, osteoblasts, and adipocytes. This may result in distinct phenotypes for both the stem and terminally differentiated cells, representing changes in their lineage plasticity and physiological function, respectively. Importantly, our versatile platform can be applied in the collection of rich data that could not have been elucidated with traditional co-culture systems that examine interactions between only two cell types. These findings suggest that this system provides a powerful platform to study the cell fate and potential healing functions of MSCs and how they are affected by the types of cells surrounding them. Similarly, the system allows assessment of the effects of MSCs on cells from their surrounding niche in mediating normal physiological functions, tissue repair, and regeneration in orthopedic and other settings.
Footnotes
Acknowledgments
The authors acknowledge funding from the NIH (R21EB009153 to H.L. and J.S.T. and T32GM008169 to T.M.H.), and the Whitaker Foundation to J.S.T. The authors thank Nathaniel C. Bloodworth and Torri E. Rinker for experimental assistance and Dr. Brani Vidakovic (BME) for assistance with statistical analysis. Primary hMSCs employed in this work were provided by the Texas A&M Health Science Center College of Medicine, Institute for Regenerative Medicine at Scott & White Healthcare through a grant from NCRR of the NIH (P40RR017447).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.