
Abstract
Adipose-derived stem cells (ASCs) have captured growing interests for cartilage regeneration. Although ASCs chondrogenesis can be stimulated by genetic modification, whether genetically engineered ASCs hold promise for the cartilaginous tissue formation remains to be explored. Since baculovirus (an emerging gene delivery vector) effectively transduced ASCs and transforming growth factor β3 (TGF-β3) was recently shown to induce ASCs chondrogenesis more potently than TGF-β1, we constructed a baculoviral vector (Bac-CT3W) to encode TGF-β3. The Bac-CT3W-transduced ASCs expressed TGF-β3 robustly and substantiated the chondrogenesis of ASCs cultured in monolayer and in porous scaffolds. Culture of the transduced cell/scaffold constructs in the rotating-shaft bioreactor (RSB) under hypoxic and perfusion conditions for 2 weeks further augmented the ASCs chondrogenesis and deposition of cartilage-specific collagen II and glycosaminoglycans, leading to the formation of cartilage-like tissues with hyaline appearance and compressive modulus approaching 62% of the native articular cartilage. Intriguingly, prolonged culture to 3 or 4 weeks failed to further augment the construct growth, probably due to the scaffold degradation. Altogether, baculovirus-mediated TGF-β3 expression in ASCs in conjunction with dynamic culture in the RSB for 2 weeks synergistically ameliorated the ASCs chondrogenesis and formation of cartilaginous tissues, representing a novel approach to producing engineered cartilages.
Introduction
Bone marrow-derived mesenchymal stem cells (BMSCs) have become a promising cell source for cartilage engineering. In vivo, chondrogenesis cascade starts with mesenchymal cell recruitment, migration, proliferation, and condensation. Likewise, BMSCs chondrogenesis commences from cell proliferation and aggregation, followed by nodule formation and differentiation into chondrocytes. 4 However, BMSCs isolation necessitates invasive and painful procedures and may result in donor site morbidity. Recently, adipose-derived stem cells (ASCs) have become popular as an alternative cell source because autologous ASCs can be readily and abundantly isolated by liposuction. Similar to BMSCs, ASCs can differentiate into chondrocytes with appropriate environmental and biological cues. Typically, ASCs commitment toward the chondrogenic lineage is guided in vitro by the chondrogenic medium supplemented with exogenous growth factors such as bone morphogenetic protein-6 (BMP-6) 5 and transforming growth factor β1 (TGF-β1), 6 resulting in the formation of compact nodules and subsequent differentiation into chondrocytes. However, recombinant growth factors are expensive and have short half-life, rendering them less cost-effective in the clinical setting. 7
The last decade has witnessed the marriage of stem cell-based therapy and gene therapy, which provides more sustained expression of endogenous chondrogenic factors and obviates the need of exogenous factors. 8 Despite the extensive evaluation of genetically modified BMSCs for bone9,10 and cartilage regeneration,11,12 however, only recently were ASCs genetically engineered with either plasmid for BMP-6 expression 13 or adenovirus for TGF-β2 expression 14 and assessed for cartilage engineering. Moreover, the gene delivery vectors in common use possess their respective drawbacks. For instance, plasmid transfection into stem cells suffers from low efficiency while adenovirus elicits potent immune responses. 8
In contrast to these vectors, baculovirus is an insect virus in nature but can efficiently transduce a broad range of mammalian cells with minimal cytotoxicity. 15 Recombinant baculovirus expressing BMP-2 can transduce and restore the differentiation of partially de-differentiated chondrocytes16,17 and stimulates the formation of engineered articular cartilages after 3-week culture of the transduced chondrocytes/scaffold constructs in the RSB. 18 Implanting the baculovirus-engineered constructs into the critical-size osteochondral defects in rabbit knees heals the defects and results in superior integration of the graft and the host cartilages, 19 thereby demonstrating the potential of baculovirus for cartilage tissue engineering.
Very importantly, baculovirus also transduces adult stem cells such as BMSCs20,21 and ASCs 22 with exceptionally high efficiency (exceeding 95%) and fairly low cytotoxicity. 23 To develop the engineered cartilages from ASCs, in this study we constructed a recombinant baculovirus (Bac-CT3W) to express TGF-β3, which promotes chondrogenic differentiation of BMSCs 24 and ASCs. 5 The rabbit ASCs were transduced with Bac-CT3W and cultured in the porous poly(L-lactide-co-glycolide; PLGA) scaffolds, either statically or dynamically in the RSB. Whether the baculovirus-mediated TGF-β3 expression in ASCs together with the dynamic culture in the RSB ameliorated the properties of the cell/scaffold constructs was evaluated.
Materials and Methods
Isolation and culture of ASCs
Cell isolation was performed according to the standard protocols as described.25,26 Briefly, adipose tissues were subcutaneously harvested from the inguinal fat pads surrounding epididymis of 8-week-old New Zealand White (NZW) rabbits, minced and washed with phosphate-buffered saline (PBS, pH 7.4) to remove debris and red blood cells. Tissues were digested in Dulbecco's modified Eagle's medium (DMEM) containing 0.08% type I collagenase, 10% fetal bovine serum (FBS; Hyclone), and 100 U/mL penicillin/streptomycin (Biological Industries) at 37°C for 60 min. The digested tissues were filtered through a 100 μm nylon mesh and centrifuged at 1,200 g for 10 min. The cell pellet was resuspended in high-glucose DMEM supplemented with 10% FBS and 100 U/mL penicillin/streptomycin, and then seeded to tissue culture flasks (50,000 cells/cm2). After 3 days, nonadherent cells were removed, and adherent cells continued to be cultured with medium exchange every 3 days. The cells were passaged by trypsinization upon 80%–85% confluence and ASCs of passage 6 (P6) were used in the following experiments.
Construction of the recombinant baculovirus
To clone the human TGF-β3 gene, the total mRNA of human BMSCs was extracted using the RNeasy Mini Kit (Qiagen) and reverse-transcribed to cDNA using Reverse Transcriptase 1st-Strand cDNA Synthesis Kit (Epicentre Biotechnologies). The cDNA (200 ng) was mixed with the polymerase chain reaction (PCR) mixture consisting of 2 U Phusion High-Fidelity DNA Polymerase (Finnzymes), Phusion HF Buffer, 10 mM dNTP and 10 μM primers (forward primer: 5′-GTCGACATGAAGATGCACTTGCAAAG-3′, and reverse primer: 5′-GGATCCTCAGCTACATTTACAAGAC-3′) in distilled water (total volume=50 μL). The PCR was performed under the condition of 10 s at 98°C, 30 s at 58°C and 30 s at 72°C for 28 cycles. The PCR product (TGF-β3 gene) was subcloned into the baculovirus donor plasmid pBac-CWPRE 22 in between the cytomegalovirus immediate-early promoter (CMV-IE) and woodchuck hepatitis post-transcriptional regulatory element (woodchuck hepatitis post-transcriptional regulatory element [WPRE]). The recombinant baculovirus Bac-CT3W was constructed using this plasmid following the instructions of Bac-To-Bac® systems (Invitrogen). The baculovirus was amplified, stored, and titrated as described. 21
Baculovirus transduction
P6 rabbit ASCs were seeded onto 6-well plates (≈2×105 cells/well) or T-150 flasks (3×106 cells/flask), cultured overnight and washed twice with PBS (pH 7.4) prior to transduction. Depending on the multiplicity of infection (MOI), a certain volume of virus supernatant was diluted with TNM-FH medium and mixed with sodium bicarbonate-free DMEM at a volumetric ratio of 1:4. The transduction was initiated by directly adding the virus solution to the cells (500 μL/well for six-well plates and 5 mL for T-150 flasks) and continued by gentle shaking on a rocking plate at room temperature for 4 h. After the virus transduction, the virus mixture was removed and the cells were cultured at 37°C using the chondrogenic medium (high-glucose DMEM containing 1% FBS, 1% penicillin/streptomycin, 1 mM sodium pyruvate, 0.1 mM ascorbic acid-2-phosphate, 0.1 μM dexamethasone, and 1% ITS+ [insulin–transferrin–selenium]). The medium was completely changed every 2–3 days.
Scaffold preparation and 3D culture under static and dynamic conditions
The PLGA (Purac, Inc.) scaffolds were prepared as described previously. 19 The scaffolds measured ≈5 mm in diameter and ≈3 mm in thickness. The porosity was ≈85% while the pore size ranged between ≈250 and 420 μm. The scaffolds were sterilized with 70% ethanol and prewetted with fresh medium overnight prior to use. One day after transduction on the T-150 flasks, ASCs were trypsinized, seeded into PLGA scaffolds (6×106 cells/scaffold), transferred to 24-well plates, and statically cultured using the chondrogenic medium with medium exchange every 2–3 days.
Some cell/scaffold constructs after 1 week static culture were aseptically positioned to the horizontal shaft in the RSB equipped with the inlets and outlets for continuous medium and gas supply. To mimic the low oxygen concentration in the native cartilage environment, the gas composition was controlled at 5% CO2, 5% O2, and 90% N2. The reactor was operated in a dynamic (rotation speed=10 rpm), perfusion mode (medium flow rate=0.2 mL/min) under the previously optimized condition. 18
Analyses of TGF-β3 expression and viability
The medium from the transduced culture was measured using the TGF-β3 ELISA kit (R&D Systems). The cell viability in the center of the constructs was determined using the Live/Dead Viability Kit (Invitrogen) and visualized by a confocal microscope, by which the green and red fluorescence indicated live- and dead cells, respectively.
Histological and immunohistochemical staining
For monolayer culture, the cell morphology was observed under the microscope and the accumulation of GAGs was stained by Toluidine blue. For 3D culture, the cell/scaffold constructs were fixed using 10% formaldehyde overnight at 4°C, embedded in paraffin, and sectioned (5 μm). The sections were stained with hematoxylin and eosin (H&E) for cell morphology or stained with 1% Alcian blue for the sulfated GAGs according to standard protocols. 27 For immunohistochemical staining, the endogenous peroxidase activity in the sections was inactivated using 1% H2O2 in methanol. Following the treatment with hyaluronidase (0.2% in PBS) at 37°C for 30 min and 2 PBS washes, the sections were blocked with 2% bovine serum albumin at room temperature for 1 h and then incubated with the primary mouse monoclonal antibody against rabbit COL I or II (1:100 dilution; Calbiochem) at room temperature for 2 h. After washing, the sections were incubated with biotinylated goat anti-mouse IgG (1:250 dilution; Kirkegaard & Perry Laboratories) at room temperature for 2 h. Finally, HRP-conjugated streptavidin (Vectastain ABC kit; Vector Laboratories) was added for incubation at room temperature for 30 min. The presence of COL I and II was visualized using the DAB Substrate kit (Vector Laboratories).
Quantitative biochemical analyses
The constructs (n=5) were sampled from the RSB at different times and the dried constructs were crushed and digested at 60°C for 12 h with 0.05 mg/mL papain in the buffer (55 mM sodium citrate dehydrate, 150 mM sodium chloride, 5 mM L-cysteine HCl, and 5 mM EDTA, pH 6.5). The GAGs content was measured using dimethylmethylene blue dye with bovine trachea chondroitin sulfate as the standard and the optical density at 525 nm was recorded. COL II was quantified using the ELISA-based Native Type II Collagen Detection kit (Arthrogen-CIA®; Chondrex) following the manufacturer's instructions.
Mechanical testing of engineered constructs
The constructs were collected from the RSB (n=5) and measured for the diameters using a caliper before mechanical testing. Mechanical properties were measured in confined compression using an electromechanical materials testing system (ElectroForce® 5210 BioDynamic® test instrument; Bose) as described previously. 28
Statistical analysis
The data are expressed as mean±standard deviations of three independent culture experiments and analyzed using Student's t-test. p-Values less than 0.05 were considered significant.
Results
Baculovirus-mediated TGF-β3 expression substantiated the ASCs chondrogenesis
Albeit a promising cell source, ASCs are difficult to transfect and are inferior to BMSCs in terms of chondrogenic differentiation potential. 29 Given that baculovirus can transduce ASCs at efficiencies approaching 98% (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/tea) and TGF-β3 was recently shown to potently promote the chondrogenic differentiation of ASCs, 30 we constructed a recombinant baculovirus, Bac-CT3W, to express TGF-β3 (Fig. 1A). The NZW rabbit ASCs in the 6-well plates (2×105 cells/well) were transduced with Bac-CT3W at different MOI and cultured using the chondrogenic medium without any growth factor. The culture medium was completely exchanged every 2–3 days and analyzed by ELISA.

Effects of virus dosage on the chondrogenesis of rabbit ASCs.
Figure 1B delineates that MOI 10 conferred low and transient TGF-β3 expression while MOI 50 elevated the maximum TGF-β3 expression level at 5 days post-transduction (dpt) to 178±17 ng/mL. Further raising the MOI to 100 or 150 neither enhanced nor prolonged the expression. Despite the transient expression, the TGF-β3 levels at 10 dpt for MOI 50, 100 and 150 remained higher than 20 ng/mL. Figure 1C reveals that mock-transduction failed to elicit evident signs of cellular differentiation even at 21 dpt. MOI 10 triggered cell condensation at 10 dpt but elicited no nodule formation, a hallmark of chondrogenic differentiation. In contrast, MOI 50 and 100 induced the cell condensation at 7 dpt and formation of compact nodules at 10 dpt, which continued to enlarge with time. At MOI 150, nodule formation was distinguishable at 10 dpt but the cell aggregates shrank after 14 dpt. The Toluidine blue staining (Fig. 1D) illustrated that the GAGs deposition at MOI 50 and 100 was detectable at 7 dpt, continued to condense toward the nodules and was better than that at MOI 150.
Baculovirus-mediated TGF-β3 expression enhanced the formation of engineered cartilages
Since MOI 50 and 100 more effectively promoted the chondrogenic differentiation, ASCs were next transduced with Bac-CT3W at MOI 50 or 100, seeded to porous PLGA scaffolds (6×106 cells/scaffold) and cultured in 24-well plates for 2 weeks using the growth factor-deficient chondrogenic medium (n=5). ELISA analyses (Supplementary Fig. S2) showed that MOI 50 conferred significantly higher TGF-β3 expression than MOI 100; thus, we evaluated the formation of engineered cartilages by ASCs transduced at MOI 50 (n=5). In parallel, the mock-transduced ASCs were seeded to the PLGA scaffolds and cultured in a similar fashion as the control (n=5).
Judging from the gross appearance (Fig. 2A), the transduced constructs grew in size and appeared glassy at week 2 due to the ECM accumulation. The H&E staining of constructs (Fig. 2B) in the surface zone revealed that the mock-transduced ASCs were still dispersed at week 1. In contrast, the transduced cells in the surface zone started to exhibit signs of condensation at week 1 and were already poised to grow interstitially with multiple cell clusters at week 2. The Alcian blue staining (Fig. 2C) and COL II-specific immunostaining (Fig. 2D) performed at week 2 further attested that Bac-CT3W transduction evidently improved the accumulation of both GAGs and COL II. Of note, the nodule formation and cartilage-specific ECM accumulation were more pronounced in the center zone than in the surface zone, regardless of transduction. These data altogether indicated that Bac-CT3W transduction at MOI 50 effectively accelerated and augmented the chondrogenic differentiation of ASCs cultured in the 3D environment.
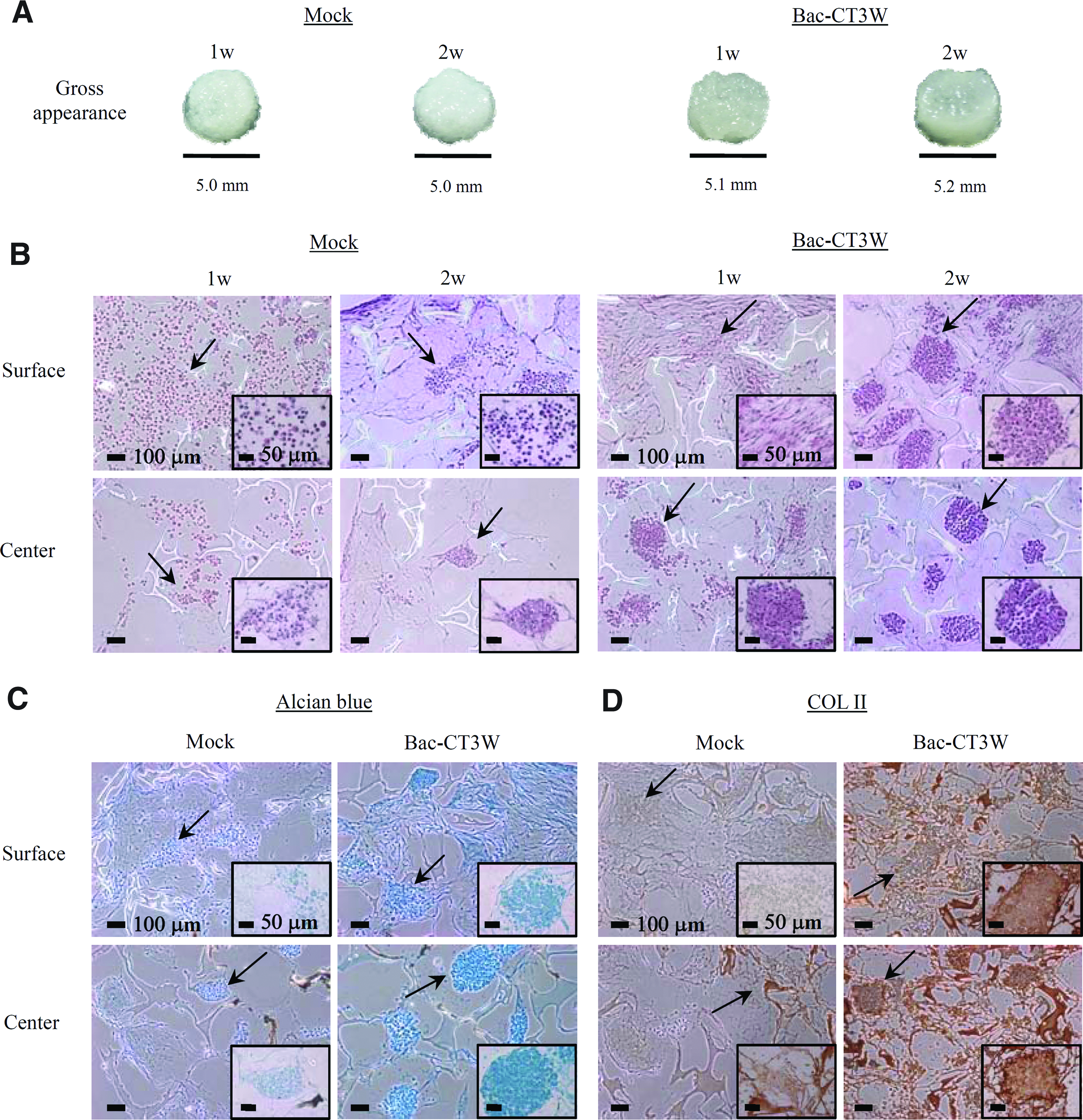
Effects of Bac-CT3W transduction on the ASCs chondrogenesis and cartilage formation.
RSB and Bac-CT3W transduction synergistically ameliorated the formation of cartilaginous tissues
Formation of engineered cartilages is stimulated by dynamic culture in the bioreactor, 2 yet static preculture is crucial for the chondrogenic differentiation of BMSCs/ASCs because immediate dynamic culture suppresses the chondrogenesis.27,31 Therefore, mock-transduced and transduced ASCs were seeded to the PLGA scaffolds, statically cultured in the 24-well plates for 1 week as in Figure 2, and then transferred to the RSB (Fig. 3A) for dynamic culture. Since chondrogenic differentiation of ASCs is potentiated by low oxygen concentration, 32 the reactor was operated with 5% O2 (5% CO2 and 90% N2) in a perfusion mode.
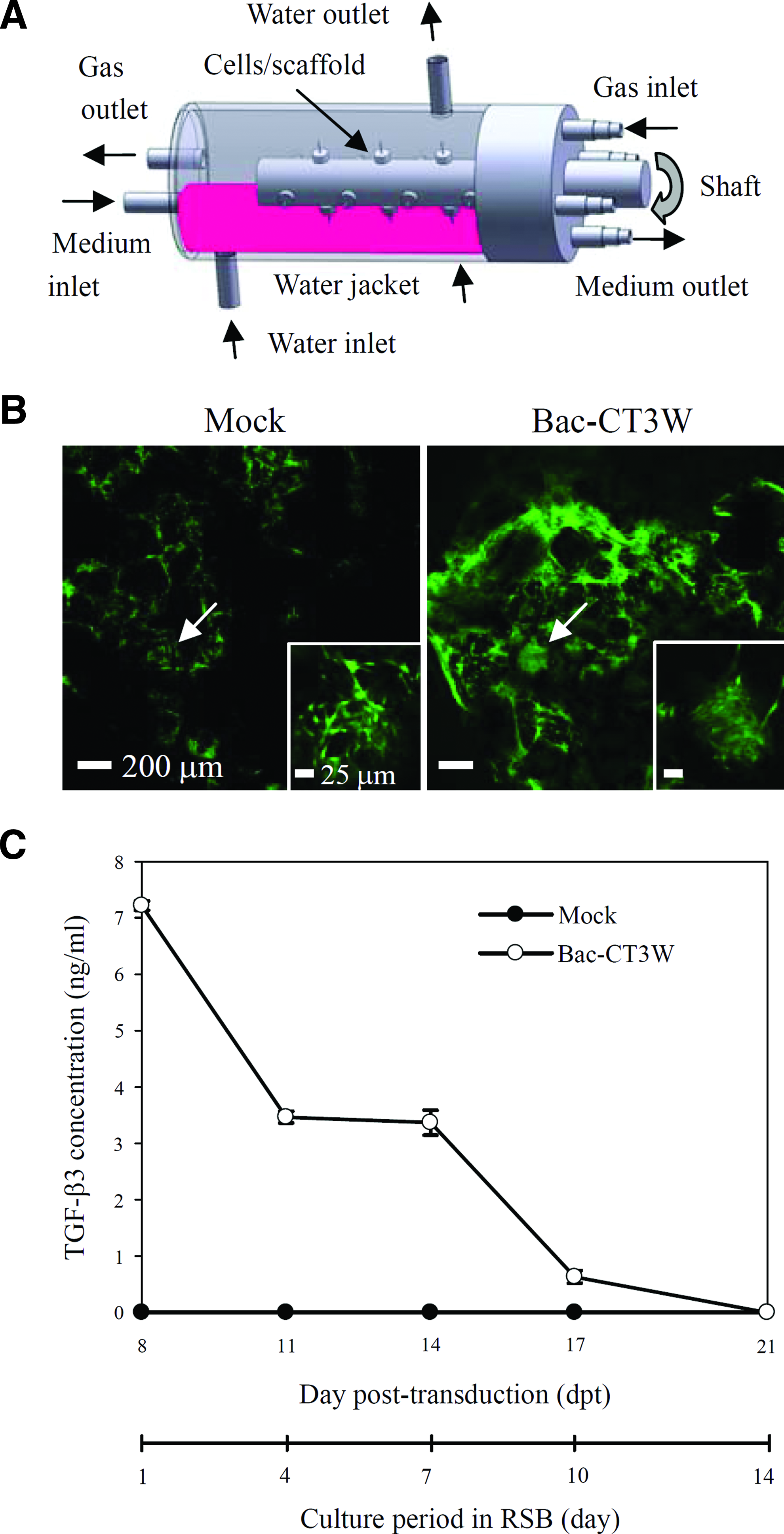
Culture of ASCs/PLGA constructs in the RSB.
Two weeks after the RSB culture, the viability was analyzed using the Live/Dead kit. The representative confocal microscopy images (Fig. 3B) illustrated green-fluorescing (live) cells without discernable red-fluorescing (dead) cells in the center of the transduced and mock-transduced constructs (Fig. 3B), attesting that the cells within the nodules remained viable. ELISA analyses of the culture supernatant (Fig. 3C) confirmed that the viable, transduced cells continued to express TGF-β3 in the RSB although the TGF-β3 expression decreased with time and extinguished at 21 dpt (day 14 in the RSB culture). Such transient expression is common for the baculovirus vector as baculovirus does not replicate in the mammalian cells and the virus genome is degraded within the cells. 33
With TGF-β3 expression, the diameter of the transduced constructs (n=5) evidently grew from 5.4±0.1 mm at week 1 to 6.2±0.1 mm at week 2 thanks to the ECM accumulation (Fig. 4), yet the diameter remained near 6.2±0.1 mm thereafter. At week 1 the cells in the center of the transduced constructs already formed cell nodules (Fig. 5A), deposited GAGs (Fig. 5B), and accumulated COL II (Fig. 5C), but did not produce the de-differentiation marker COL I (Fig. 5D). At week 2 the cell nodules and ECM accumulation in the transduced constructs became even more prominent and were remarkably superior to those in the mock-transduced constructs (Fig. 5A–C). In particular, at week 2 the lacuna and round cells (Supplementary Fig. S3) were observable while COL I remained undetectable (Fig. 5D) in the transduced constructs. Further, chondrocyte-specific genes (e.g., COL II and aggrecan) were significantly upregulated at week 2 (Supplementary Fig. S4). These data indicated that the transduced ASCs had differentiated into mature chondrocytes. In contrast, the cells in the mock-transduced constructs remained fibroblast-like at week 2 (Supplementary Fig. S3) and expressed COL I (Fig. 5D) instead of COL II (Fig. 5C).

Gross appearance of the engineered cartilages cultured in the RSB. The constructs were cultured in the RSB as in Figure 3 and n=5 for both the transduced and mock-transduced constructs in each time point. Color images available online at www.liebertpub.com/tea

Histochemical and immunohistochemical staining of engineered cartilages cultured in the RSB.
At weeks 3 and 4, we surprisingly observed progressive regression of ECM on the surface (Fig. 4), loss of GAGs and cells, and increased COL I deposition (Fig. 5A–D and Supplementary Fig. S3) in the mock-transduced constructs, suggesting the construct degeneration. Breakdown of GAGs, COL II, and scaffold was similarly observed in the transduced constructs.
Concurrently, the total GAGs (6.6±0.2 mg/construct, Fig. 6A) and COL II (354.2±43.8 μg/construct, Fig. 6B) contents of the transduced constructs (n=5) culminated at week 2, which corresponded to≈52% and ≈40% that of the native articular cartilages of the same size, respectively, and declined after week 2. In agreement with the ECM content analyses, the biomechanical testing (Fig. 7) depicted that the compressive modulus of the transduced constructs (n=5) peaked at week 2 (3.0±0.6 MPa), which corresponded to 62% that of the native cartilages (4.8±0.3 MPa). In contrast, the compressive moduli of the mock-transduced constructs at week 2 (1.4±0.6 MPa) and the blank PLGA scaffold (0.3±0.1 MPa) were only ≈30% and ≈5% that of the native cartilages, respectively. Concomitant with the decreasing ECM content, the compressive moduli of the transduced and mock-transduced constructs also declined with time.
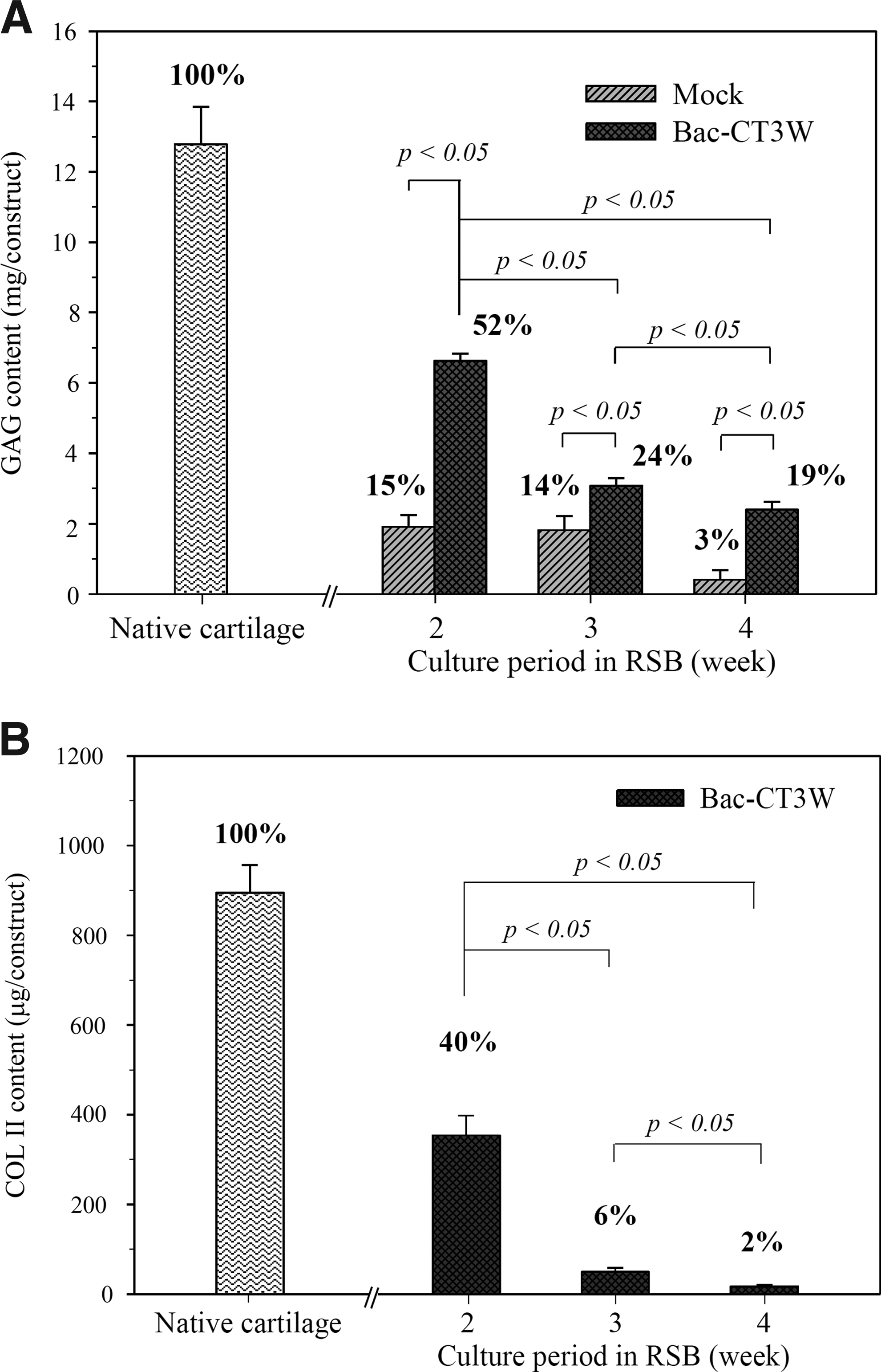
Effects of in vitro culture time on the

Effects of in vitro culture time on the mechanical property. The numbers above the bars represent the percentages of the compressive moduli when compared with the native cartilages of the same size.
Discussion
ASCs have sparked growing interests for cartilage engineering in recent years. 29 Since chondrogenic preinduction of tissue-engineered cartilage and implanting into the defect a construct capable of withstanding the mechanical loads are imperative to prevent construct deformation in vivo 34 and improve cartilage repair,19,35 it is desired to initiate the ASCs chondrogenesis and induce deposition of cartilage-specific ECM and formation of cartilage-like tissues in vitro. To date, ASCs commitment into the chondrogenic pathway is commonly triggered by culturing in the chondrogenic medium containing growth factors such as BMP-65 and TGF-β1. 6 When induced with TGF-β1,36–38 however, ASCs undergo chondrogenesis to a lesser extent than BMSCs presumably because ASCs have fewer TGF-β receptors. 30 Nonetheless, it was recently discovered that TGF-β3 is more potent than TGF-β1 in promoting ASCs chondrogenesis 5 and continued TGF-β3 supply in the medium ameliorates chondrogenic differentiation of ASCs, 39 suppresses the BMSCs hypertrophy 27 , and improves the mechanical properties of the engineered cartilages. 40 Therefore, TGF-β3 was selected as a novel transgene to promote the ASCs chondrogenesis.
We unraveled that Bac-CT3W transduction of ASCs at MOI 50 gave rise to the optimal TGF-β3 expression level either in monolayer (Fig. 1B) or in 3D culture (Supplementary Fig. S2). Thanks to the efficient baculovirus transduction, the expression level amounted to ≈178 ng/mL at 5 dpt and far exceeded the TGF-β3 concentration (10 ng/mL) generally supplemented to stimulate the chondrogenesis of BMSCs 27 and ASCs 5 for at least 12 days (Fig. 1B). Since 10–50 ng/mL of exogenous TGF-β3 combined with dexamethasone was insufficient to induce chondrogenesis in ASCs as efficiently as in BMSCs under the same conditions,30,41 higher concentrations of growth factors were required for ASCs to match the chondrogenic capacity of BMSCs. 38 As such, the robust and prolonged TGF-β3 expression mediated by Bac-CT3W was crucial to warrant the ASCs chondrogenesis. In accord, MOI 50 led to rapid cell condensation, nodule formation (Fig. 1C), GAGs deposition (Fig. 1D), and was superior to MOI 0 (i.e., mock-transduction) and other MOI. Such vector dose dependency was probably due to the relatively higher cytotoxicity at higher MOI (Supplementary Fig. S5) and was similarly observed in BMSCs transduced with adenovirus 42 and in chondrocytes transduced with baculovirus. 16 In the static culture within the PLGA scaffold (Fig. 2), Bac-CT3W transduction further enhanced the cellular differentiation and ECM deposition, and excelled when compared with mock-transduction.
One step further, we combined the baculovirus-mediated TGF-β3 expression and RSB culture, hoping that multiple stimuli could act in concert to promote the chondrogenesis. Indeed, the cells cultured for 2 weeks remained viable and able to express TGF-β3 (Fig. 3B, C), indicating that RSB conferred sufficient nutrient and gas transfer to maintain viability even under the hypoxic conditions. Orchestrated stimulation with these biological (TGF-β3) and physical (bioreactor and hypoxia) factors remarkably enhanced the construct growth (Fig. 4), accelerated the chondrogenic differentiation (Fig. 5A), and ameliorated the cartilage-specific ECM accumulation in 2 weeks (Figs. 5B, C and 6), while obviating the production of de-differentiation marker (Fig. 5D). Concurrently, the compressive modulus of the transduced constructs at week 2 was significantly elevated to ≈62% that of the native cartilages (Fig. 7). The cartilaginous tissue formation significantly excelled in comparison with the Bac-CT3W-transduced constructs in static culture (Fig. 2), attesting the beneficial effects imparted by the dynamic, perfusion culture in the RSB. Conversely, the mock-transduced constructs produced less cartilage-specific ECM and expressed COL I (Figs. 5C, D and 6B), suggesting that, without Bac-CT3W transduction, dynamic culture using the chondrogenic medium was insufficient to induce robust ASCs chondrogenesis and correct ECM expression, leading to the poor compressive moduli of the mock-transduced constructs. Although after completion of this work a recent publication demonstrated the transfection of TGF-β3 into precartilaginous stem cells, the transfection only induced the cells to differentiate into chondroblasts in 2D culture. 43 This less-than satisfactory result was likely due to the poor plasmid transfection efficiency into stem cells. In contrast, baculovirus is very efficient in transducing ASCs (transduction efficiency exceeds 95%). Our data further confirmed that Bac-CT3W transduction in conjunction with RSB culture synergistically directed the ASCs differentiation to chondrocytes and promoted the formation of cartilaginous tissues from ASCs, thus representing a significant progress.
Strikingly, extended culture past 2 weeks did not impart beneficial effects to cell differentiation and cartilage growth. Instead, ECM breakdown (Figs. 4–6), loss of cells (Supplementary Fig. S3), and impaired mechanical property (Fig. 7) occurred at weeks 3 and 4, which concurred with the degradation of ASCs/PGA (polyglycolic acid) constructs induced with such growth factors as BMP-6 and TGF-β1 after 2-week culture. 31 In another recent study, ASCs were seeded to the composite PGA/PLA (polylactic acid) scaffold and cultured in the chondrogenic medium containing 10 ng/mL of TGF-β1 and 100 ng/mL of IGF-1 (insulin-like growth factor) for 5 weeks. 35 Likewise, the amounts of GAGs and cells significantly elevated until week 2 but ceased to rise in the following 3 weeks. The stagnant construct growth and even construct breakdown were attributed to the failure of ASCs to differentiate into mature chondrocytes fast enough to synthesize sufficient ECM molecules to replace the degraded scaffolds. 31 Similarly, the constructs deterioration observed at weeks 3 and 4 probably arose from the hydrolysis of the PLGA polymer (Fig. 5A–C) and reorganization of the microstructure, 44 leading to the constructs shrinkage and degradation.
The deterioration of the constructs comprising ASCs suggests the importance of fabricating a proper scaffold material with controllable and desirable degradation rate. This also underscores the necessity to expedite the ASCs differentiation and augment ECM accumulation before scaffold degradation. In this regard, hyaluronan (HA)-coated PLGA scaffolds can enhance the ASCs chondrogenesis, 45 thus the ASCs chondrogenesis may be further improved by the use of HA-PLGA scaffold in combination with baculovirus-mediated genetic modification. Conversely, an FLP/Frt-based hybrid baculovirus vector enabled prolonged maintenance of the transgene in the episomal form. 20 This hybrid vector enhanced and extended the BMP-2 and vascular endothelial growth factor expression in rabbit ASCs, thereby augmenting the osteogenesis and accelerating the repair of critical-size segmental defects at the femora of NZW rabbits after implantation. 22 As such, use of the FLP/Frt-based hybrid vector for prolonged TGF-β3 expression (e.g., for more than 2 weeks) at high levels, instead of transient stimulation as in this study, might further improve the ASCs chondrogenesis and the formation of engineered cartilages.
Taken together, although ASCs are attractive for tissue regeneration, whether genetically modified ASCs hold promise for cartilage engineering remains to be established. We demonstrated that baculovirus-mediated engineering of ASCs and subsequent TGF-β3 expression, in combination with dynamic culture in the RSB for 2 weeks, synergistically improved the ASCs chondrogenesis and formation of cartilaginous tissues. Recombinant baculovirus is easy to construct and produced high titers simply by infecting insect cells. Baculovirus transduction is simple and straightforward without signs of viral replication and random integration. 15 Baculovirus transduction of stem cells neither disrupts the differentiation potentials nor alters the immunopriviliged properties,46,47 and implantation of baculovirus-transduced stem cells only elicits mild and transient inflammatory responses, 46 with no signs of tumorigenesis 23 even after a 6 month follow-up (unpublished data). These properties render baculovirus a highly attractive vector for the ASCs engineering and production of engineered cartilages. The compressive moduli of the transduced constructs approached 62% that of the native cartilage at week 2. Such mechanical property was satisfactory when compared with the compressive modulus of ASCs/PGA constructs, which reached only ≈51% that of the normal cartilage after 2-week induction in vitro and 3 months after implantation but were still able to repair full-thickness cartilage defects. 35 Also, extended periods of cultivation of the cartilaginous constructs may result in poor integration between the graft and host cartilages. 48 Therefore, the Bac-CT3W-transduced ASCs/PLGA constructs cultured in the RSB for 2 weeks hold great promise for cartilage repair.
Footnotes
Acknowledgments
The authors thank Prof. Shu Wang (National University of Singapore) for providing the plasmid containing the WPRE sequence and Prof. Jyp-Ping Chen of Chang Gung University for the biomechanical testing. The authors also acknowledge the financial support from the National Tsing Hua University (Toward World-Class University Project, 100N2050E1, Booster Program 99N2544E1), CGMH-NTHU Joint Research Program (99N2419E1, 100N7753E1, 101N2753E1, CMRPG3B0431, CMRPG361041), CGMH Intramural Project (CMRPG300131, CMRPG300161), and National Science Council (NSC 99-2221-E-007-025-MY3), Taiwan.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.