
Abstract
Since the first reports of induction of adipose-derived stem cells (ASC) into neuronal and glial cell phenotypes, expectations have increased regarding their use in tissue engineering applications for nerve repair. Cell adhesion to extracellular matrix (ECM) is a basic feature of survival, differentiation, and migration of Schwann cells (SC) during nerve regeneration, and fibronectin and laminin are two key molecules of this process. Interaction between ECM and SC-like differentiated ASC (dASC) could potentially improve the neurotrophic potential of the stem cells. We have investigated the effect of ECM molecules on SC-like dASC in terms of proliferation, adhesion, and cell viability. Fibronectin and laminin did not affect the proliferation of dASC when compared with cell adherent tissue culture plastic, but significantly improved viability and cell attachment when dASC were exposed to apoptotic conditions. To assess the influence of the ECM molecules on dASC neurotrophic activity, dASC were seeded onto ECM-coated culture inserts suspended above dorsal root ganglia (DRG) sensory neurons. Neurite outgrowth of DRG neurons was enhanced when dASC were seeded on fibronectin and laminin when compared with controls. When DRG neurons and dASC were in direct contact on the various surfaces there was significantly enhanced neurite outgrowth and coculture with laminin-conditioned dASC produced the longest neurites. Compared with primary SCs, dASC grown on laminin produced similar levels of neurite outgrowth in the culture insert experiments but neurite length was shorter in the direct contact groups. Anti β1 integrin blocking antibody could inhibit baseline and dASC evoked neurite elongation but had no effect on outgrowth mediated by laminin-conditioned dASC. ECM molecules had no effect on the levels of nerve growth factor and brain-derived neurotrophic factor secretion from dASC. The results of the study suggest that ECM molecules can significantly improve the potential of dASC for nerve regeneration.
Introduction
Tissue engineering studies for nerve repair have utilized combinations of biomaterial scaffolds and various cells types and growth factors. Cell adhesion to extracellular matrix (ECM) is a key feature of survival, differentiation, and migration of cells. 16 After peripheral nerve injury, ECM molecules are rapidly upregulated providing a permissive environment and axon guidance, allowing axons to regrow toward the correct targets.17,18 Signals to both neurons and SC from ECM largely depend on integrins, a family of heterodimeric receptors. Integrins mediate axonal guidance, adhesion, and cell migration and are expressed in growth cones of regenerating axons. 19 Fibronectin mediates SC activation and myelination after injury, 20 and acting through the RGD tripeptide moiety (Arg-Gly-Asp), it binds specific integrins α5 and β6 that are upregulated in the injury milieu, thus facilitating axons and SC regrowth. 21 Laminin, a basal lamina component, regulates SC function during axonal sprouting; disruption of laminin leads to severe hypomyelination and lack of axonal sorting.22,23 SC interact with the matrix and communicate with each other through the ECM molecules, which influence cell shape, fate, metabolism, and behavior, being essential for tissue development. 24 Further, laminin 25 and fibronectin26,27 incorporated into nerve conduits have produced enhanced axonal regrowth.28,29
In this study we have examined the effect of fibronectin and laminin on proliferation and cell viability of dASC, and the influence of these molecules on the neurotrophic effect of dASC when cocultured with neuronal cells.
Materials and Methods
ASC harvest and cultures
All animal procedures were carried out in compliance with the UK Animals (Scientific Procedures) Act 1986. Rat ASC were harvested as described previously 10 and maintained in α-modified Eagle's medium (Invitrogen) containing 10% (v/v) fetal bovine serum (FBS), and 1% (v/v) penicillin/streptomycin solution. The multipotent nature of the cultures was determined as previously described. 10 For induction to a SC phenotype undifferentiated ASC at early passages (P2–P5) were treated with 5 ng/mL platelet-derived growth factor, 10 ng/mL basic fibroblast growth factor (both from PeproTech Ltd.), 14 μM forskolin (Sigma) and 126 ng/mL glial growth factor-2 (GGF-2) (a gift from Acorda Therapeutics, Inc.) as previously described. 10
NG108-15 cultures
The mouse neuroblastoma X rat glioma NG108-15 cell line was purchased from ECACC. Cells were maintained in Dulbecco's modified Eagle's medium (DMEM) plus 10% (v/v) FBS and 1% penicillin/streptomycin and maintained at 37°C, 5% CO2.
SC culture
SCs were isolated from sciatic nerves as previously described, 14 maintained in DMEM containing 10% FBS and 1% penicillin-streptomycin, and supplemented with 14 μM forskolin and 126 ng/mL neuregulin.
Dorsal root ganglia neurons harvest
Dorsal root ganglia (DRG) were harvested from adult male Sprague–Dawley rats using a previously described protocol. 30 Dissociated neurons were suspended in modified Bottenstein and Sato's medium (BSM; F12 medium containing 100 μM putrescine, 30 nM sodium selenite, 20 nM progesterone, 1 mg/mL bovine serum albumin, 0.1 mg/mL transferrin, 0.01 mM cytosine arabinoside, and 10 pM insulin (all from Sigma) and plated under various conditions described below.
Alamar blue proliferation assay
The dASC proliferation on either untreated tissue culture plastic (TCPS) or coated with the ECM molecules (fibronectin or laminin at a concentration of 2 μg/mL) was determined using the Alamar blue assay. Cells were seeded at a density of 1×103/0.25 cm2, and incubated for up to 6 days. Samples were measured every 24 h after addition of 10% (v/v) Alamar blue solution, and proliferation was recorded as the difference between two different absorbance readings, at 570 nm (reduced form) and 595 nm (oxidized form), using a multiwell plate reader.
Live/dead assays
Calcein-AM fluorescein complex and ethidium homodimer-1 (Eth-D1) were used to assess the cell viability (LIVE/DEAD® Viability/Cytotoxicity Kit; Molecular Probes, Invitrogen) after 48 h serum deprivation in culture at 37°C, 5% CO2. Cells were plated at a concentration of 2000/cm2 into 24-well plates either untreated or coated with ECM molecules at a concentration of 2 μg/cm2. After 24 h, differentiation medium was changed to serum-free medium for a further 24 h. Following phosphate-buffered saline (PBS) washings, Calcein-AM and Eth-D1 were added, at a concentration of 0.5 and 2 μL/mL respectively, and incubated for 30 min at room temperature, followed by PBS washes. Images were taken using a fluorescence inverted microscope (Olympus IX51) under 4× magnification, for a total of five fields for each well, representing more than 30% of the well surface. Similar experiments were performed by exposing the cells to PBS only for 2 h, conditions that evoke significant cell detachment or anoikis. Cells were then washed by PBS and the remaining attached cells were stained with the Live/Dead kit as described above. In addition to determining the percentage of viable cells, the number of adherent cells was quantified and expressed as a percentage of the total number of adherent cells compared with control wells (cells maintained in growth medium).
Microscopic fields were chosen following the same geometrical pattern for each treatment. Images captured under both CY3 (dead cells) and fluorescein isothiocyanate (FITC) (live cells) fluorescence filters were merged, and quantification of viable cells was determined using the cell counter tool of ImageJ Software (NIH). The experiment was performed five times with dASC cells derived from different animals.
DRG coculture functional bioassays
Transwell
Twenty-four hours prior to DRG harvest, 40,000 dASC (20,000/cm2) were seeded on uncoated, fibronectin- and laminin-coated 1.0 μm pore size cell inserts (BD Falcon) in differentiation medium. After 24 h incubation (37°C, 5% CO2) the inserts were checked for cell adherence and placed into a 24-well plate (Costar) containing DRG neurons to establish the coculture for further 24 h (37°C, 5% CO2). Prior to the coculture, cells in inserts were washed by HBSS and the medium was changed to a reduced serum (1%) differentiation medium to minimize satellite cells proliferation in the DRG preparations. Additional wells with inserts (uncoated or treated with fibronectin or laminin) containing only differentiation medium with 1% serum were used as controls. Cocultures were maintained for 24 h.
Direct contact
The DRG coculture bioassay was repeated with direct contact with dASC seeded on uncoated or fibronectin- and laminin-coated glass coverslips. Prior to DRG harvest, dASC were seeded at a density of 20,000/cm2 onto coverslips and incubated for 24 h at 37°C, 5% CO2. The surfaces were checked for cell confluence and DRGs were added on top of the cells. The differentiation medium of dASC was changed to a mixed medium (50% BSM and 50% differentiation medium with growth factors with 1% serum) to meet the requirements of the two cell lines. DRGs alone, respectively on uncoated or fibronectin- and laminin-coated coverslips served as controls. Cells were cocultured for 24 h. For integrin antibody blocking experiments, primary neurons were plated as above, and Armenian hamster anti-CD29 antibody (β1 integrin; 1:50; BD Biosciences) or an isotype-matched control antibody (1:50; BD Biosciences) was added to culture media 2 h after DRGs plating, when mixed medium was topped up according to previous reports.31,32
Both transwell and direct contact experiments were also performed using primary SCs seeded under the same conditions as described above.
NG108-15 coculture functional bioassays
Prior to coculture, dASC were seeded (20,000/cm2) on polystyrene slide flasks (Nunc, Fisher Scientific), coated as described above. After 24 h, NG108-15 cells suspended in dASC growth medium were seeded over dASC at 500 cells/cm2 density. After 48 h coculture, the samples were fixed for immunocytochemistry (see below). Uncoated slide flasks seeded first with dASC and then with NG108-15 cells, and slide flask for each condition (uncoated plastic, fibronectin, and laminin coatings) seeded just with NG108-15 cells served as controls. Three independent coculture experiments with different ASC preps were performed.
Neurite outgrowth: immunocytochemistry and quantification
Cultures were fixed in 4% (w/v) paraformaldehyde for 20 min and blocked for 1 h at room temperature in normal horse serum. The cells were incubated overnight (4°C) with monoclonal anti-βIII tubulin antibody (1:500; Sigma). The following day, the cells were incubated (at room temperature in dark, 1 h) with secondary antibodies FITC conjugated horse anti-mouse IgG (Vector Laboratories). The coverslips were mounted with antifading Vectashield solution with DAPI (Vector Laboratories) and examined under a fluorescence microscope (Olympus BX60). The images for anti-βIII tubulin were captured for quantification at 10× magnification using Image-Pro Plus Imaging software (Media Cybernetics).
To assess the effect of ECM molecules and dASC on the neurite outgrowth, measurements of average neurite length and percentage of process bearing-neurons were made. Image capturing was performed using an Olympus DP12 camera and included at least 20 fields for each sample/condition in each experiment. All neurites protruding from each cell body were traced using Image Pro Plus software (Media Cybernetics) to assess their length. Both image capturing and neurite outgrowth measuring was performed by the same examiner, blinded to the groups. Experiments were repeated in at least 4 independent cocultures.
Enzyme-linked immunosorbant assay
At the end of the cocultures the medium from the wells was collected and analyzed by enzyme-linked immunosorbant assay (ELISA) using the chemikine™ brain-derived neurotrophic factor (BDNF) or nerve growth factor (NGF) sandwich ELISA kits (Chemicon) according to the manufacturer's protocol. Medium was also collected from dASC grown as above but in the absence of DRG neurons. All samples were analyzed in triplicate and the absorbance was measured at 450 nm with a Multiskan MC plate reader (Labsystems).
Statistical analysis
One-way analysis of variance with Bonferroni multiple comparison test was used to statistically analyze data (GraphPad Prism 5.00). Significance was determined as *p<0.05, **p<0.01, and ***p<0.001.
Results
Influence of ECM molecules on cell proliferation
The Alamar blue proliferation assay was used to determine dASC proliferation rates on fibronectin and laminin (2 μg/cm2) compared with TCPS. Proliferation was measured for 6 days until cells were confluent. There was no significant difference in proliferation rate of dASCs seeded on various surfaces (Fig. 1). Varying the concentration of fibronectin and laminin (0.4, 2, 10 μg/cm2) also did not show any significant differences between groups (data not shown).

Effect of extracellular matrix (ECM) molecules on cell proliferation. Tissue culture plastic (TCPS) was precoated with fibronectin or laminin. Proliferation of differentiated adipose-derived stem cells (dASC) was quantified using the Alamar blue reduction assay and compared with control cultures grown on uncoated TCPS. No significant difference in proliferation between groups was recorded over the 6-day period, when cells reached saturation. Data represent n=3 different cell preparations and are expressed as mean±SEM.
Influence of ECM molecules on cell viability
Cells transplanted into nerve conduits are subjected to stress including growth factor and oxygen deprivation. Increasing cell resistance to stress conditions could potentially enhance efficacy of cell delivery. Viability of dASC seeded on different ECM substrates and subjected to serum deprivation was assessed by measuring the number of live and dead cells (Fig. 2, left column). A significant increase in cell viability was observed when cells were seeded on fibronectin and laminin surfaces compared with uncoated TCPS (Fig. 3A).

Influence of ECM molecules on cell viability. Calcein-AM fluorescein complex, labeling viable cells (green staining), and ethidium homodimer-1 (Eth-D1), labeling dead cells (red staining) were used to assess cell viability and survival. Left column: dASC after 48 h of serum-free medium inducing apoptosis. Right column: dASC after 2 h of treatment by phosphate-buffered saline (PBS) inducing cell detachment and anoikis. Representative images of dASC seeded on ECM molecules-coated surfaces showing a better survival and superior surface adhesion when compared with uncoated cell adherent plastic (TCPS). Scale bar=100 μm. Color images available online at www.liebertpub.com/tea
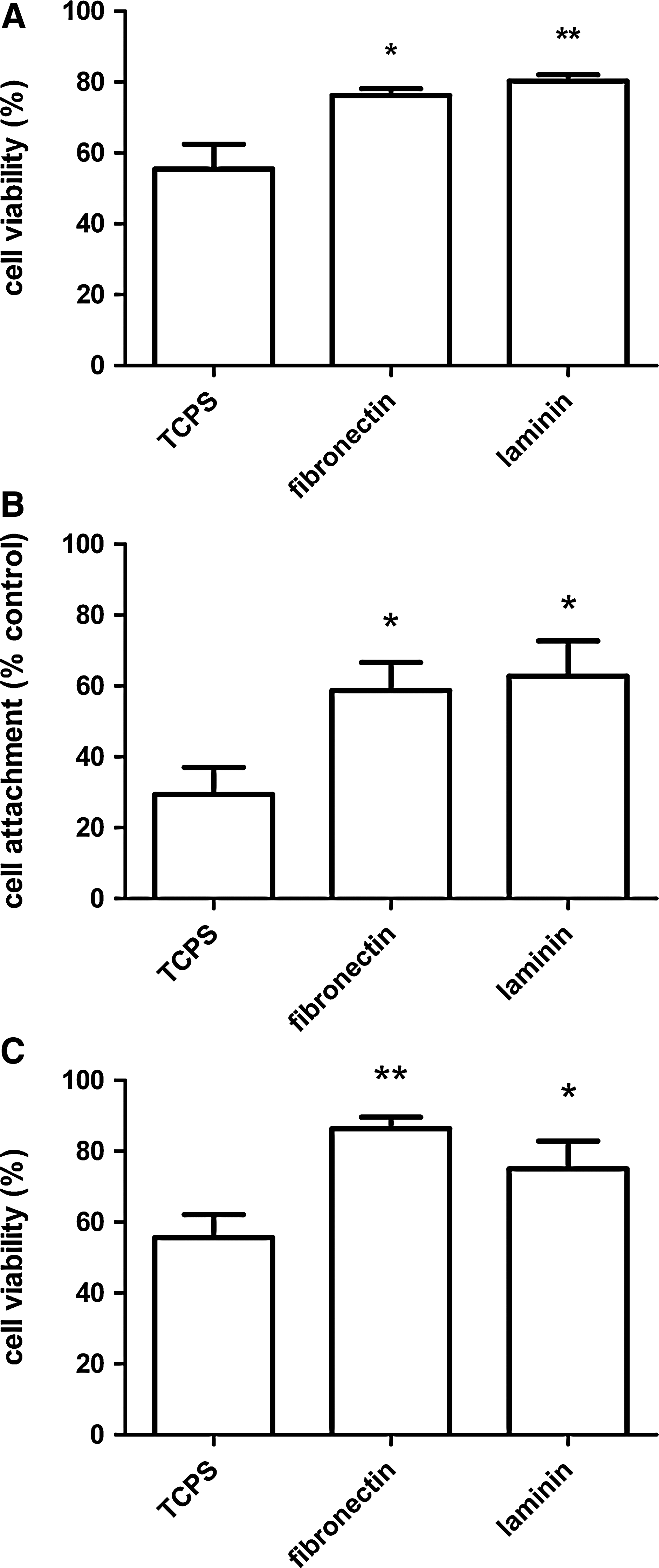
Quantitative analysis of cell viability. Quantitative analysis of live/dead images was performed from the microscopic captured fields.
Anoikis is the apoptosis triggered when cells loose contact with supportive matrix. An assessment of attachment and viability of cells that remained adherent to ECM coating was determined following dASC treatment with PBS (Fig. 2, right column). dASC seeded on fibronectin and laminin showed a significantly improved adhesion when compared with cells on uncoated TCPS (Fig. 3B). When the attached cells were assessed for viability, ECM molecules confirmed their effect on cell survival under stress condition, significantly improving cell viability when compared with TCPS (Fig. 3C).
Influence of ECM molecules on dASC neurotrophic effect
Previous experiments showed the effect of ECM molecules on SCs cocultured with neuronal cells.33–35 Since dASC exhibited glial cell potential when cocultured with NG108-15, 10 we determined whether these effects might be enhanced by ECM molecules.
dASC-DRG neuron cocultures: soluble molecules
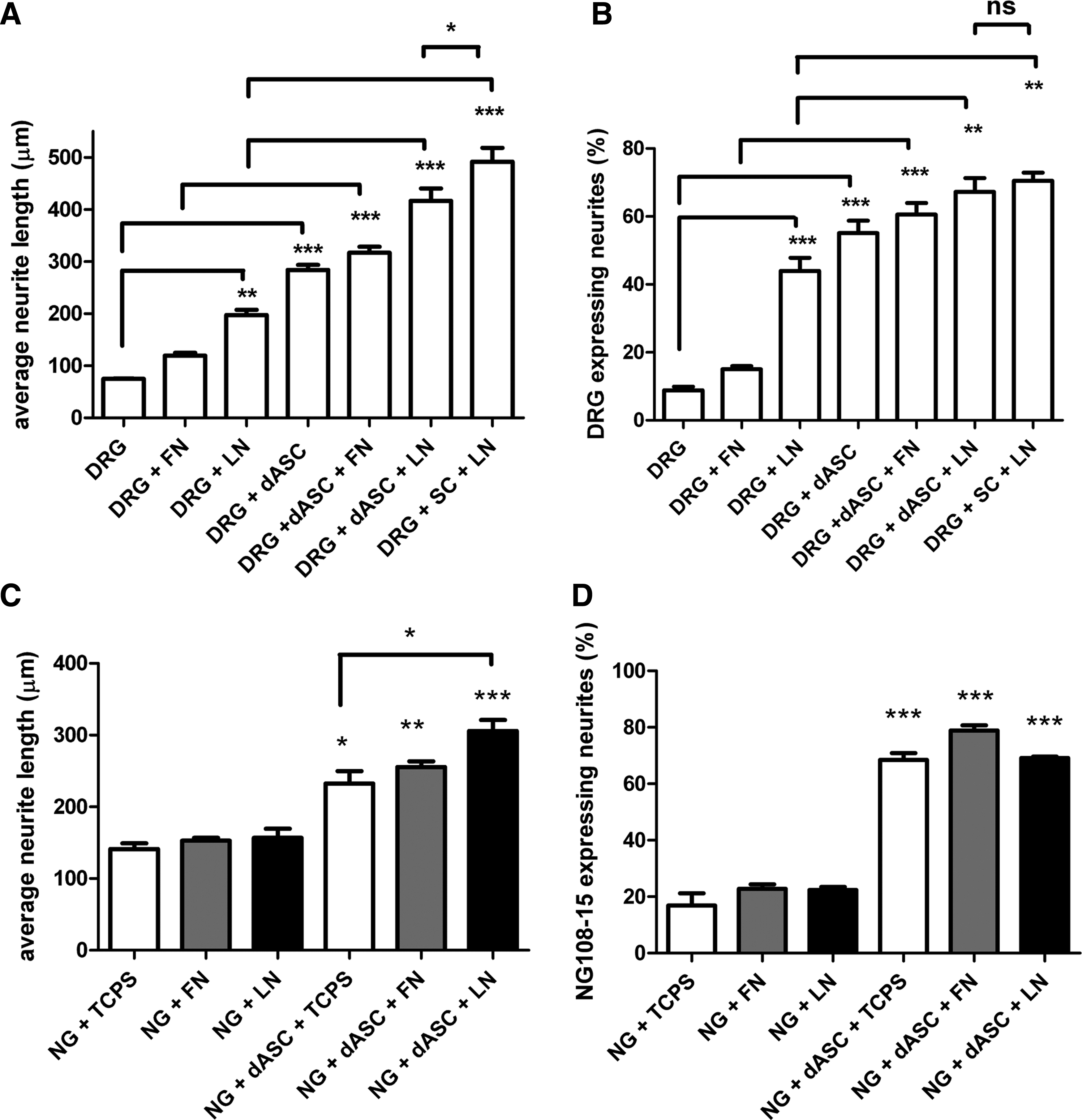
The beneficial effects of transplanted stem cells result from released soluble neurotrophic factors and cytokines. We investigated the effect of ECM molecules on the ability of dASC to modulate neurite outgrowth from neurons as a result of growth factor secretion using noncontact transwell cocultures. DRG neurite outgrowth visualized with βIII-tubulin staining was enhanced by the presence of dASC (Fig. 4A). When compared with their respective controls, the DRG neurons average neurite length was significantly increased when dASC were seeded on ECM-coated inserts (*p<0.05 for fibronectin and **p<0.01 for laminin; Fig. 4B). The neurite length evoked by dASC grown on culture inserts coated with laminin was similar to that produced by primary SCs grown on laminin (Fig. 4B). dASC seeded on uncoated and ECM-coated inserts showed a significant increase of neurons bearing neurites when compared with controls (Fig. 4C). The effect of coating of inserts with ECM molecules alone did not significantly alter neurite outgrowth from the DRG neurons seeded below (Fig. 4B, C).
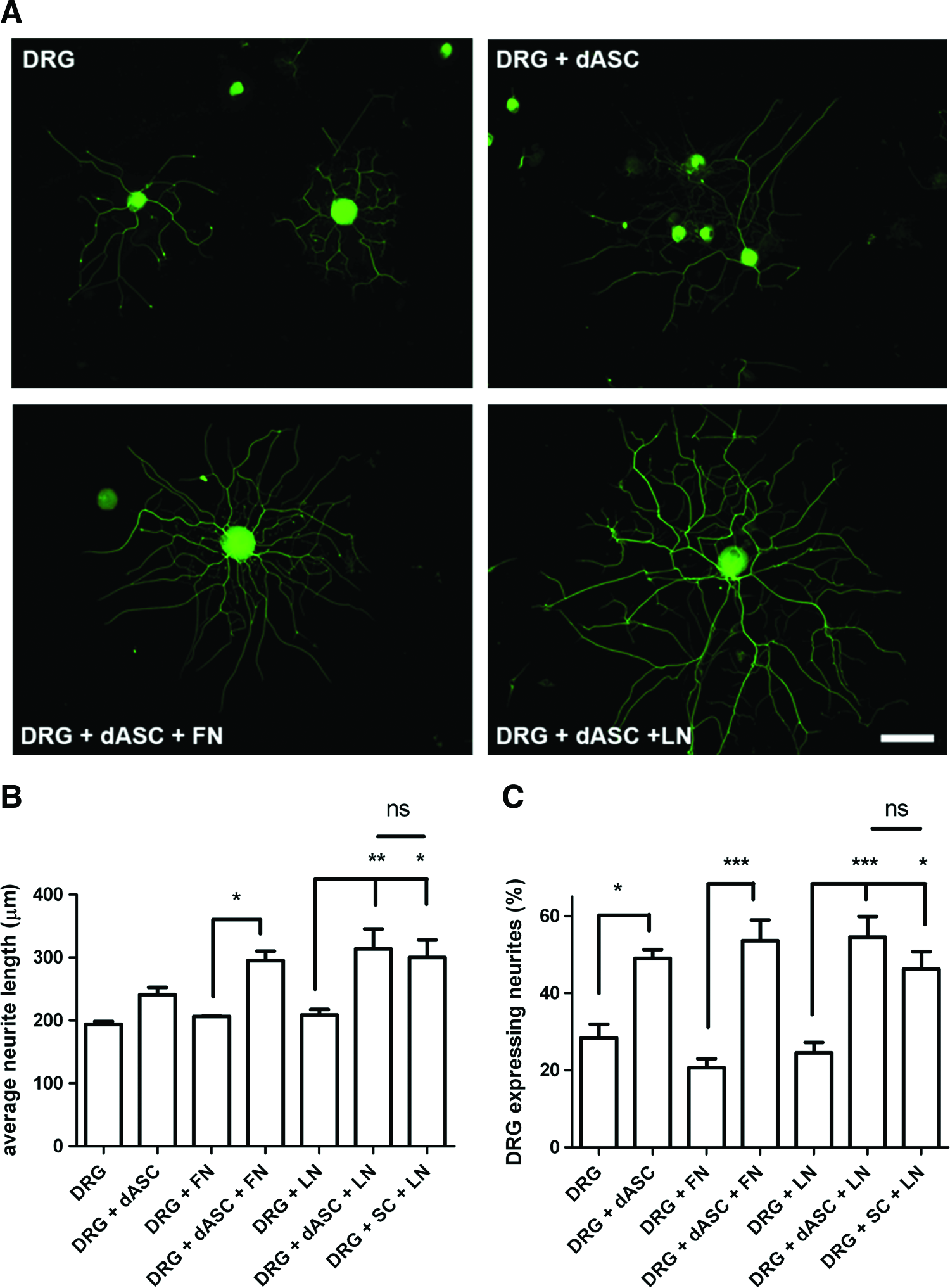
The neurotrophic effect of ECM-treated dASC on dorsal root ganglia (DRG) neurite outgrowth: soluble molecules.
dASC-DRG neuron cocultures: direct contact
We also analyzed the influence of ECM molecules on dASC ability to promote neurite outgrowth when in direct contact with DRG neurons, which were seeded on uncoated or ECM-coated coverslips in the absence or presence of dASC (Fig. 5). Neurite length was significantly longer in coculture with dASC compared with respective controls (Fig. 6A). DRG neurons in contact with dASC plated on laminin displayed the longest neurites when compared with other dASC groups but this was significantly less than that evoked by primary SCs cultured on laminin (Fig. 6A). The percentage of DRG neurons expressing neurites was also significantly increased when cocultured with dASC (Fig. 6B), although there were no significant differences between the effects of dASC seeded on uncoated or ECM-coated coverslips. Primary SCs and dASC seeded on laminin evoked similar levels of DRG neurons to express neurites (Fig. 6B).

The neurotrophic effect of ECM-treated dASC on DRG neurite outgrowth: direct contact effects. Immunocytochemistry for βIII tubulin (green) of neurite outgrowth from adult rat sensory neurons cocultured in direct contact with dASC. DRG were seeded on uncoated or ECM-coated coverslips in the absence (left column) or presence of dASC (right column). DAPI staining (blue) shows monolayer of dASC. Scale bar=100 μm. Color images available online at www.liebertpub.com/tea
The neurotrophic effect of ECM-treated dASC on DRG and NG108-15 neurite outgrowth: analysis of direct contact effects.
dASC- NG108-15 cocultures: direct contact
To confirm the effect of ECM on neurotrophic activity of dASC, the stem cells were cocultured with NG108-15 cells. Seeding NG108-15 cells directly on either fibronectin or laminin had no effect on average neurite length or percentage of cells expressing neurites (Fig. 6C, D). NG108-15 cells seeded on top of dASC plated on either TCPS or ECM molecules showed significantly increased neurite outgrowth compared with the absence of dASC. Importantly, dASC seeded on laminin showed significantly enhanced neurotrophic ability compared with dASC seeded on TCPS (Fig. 6C).
Mechanisms of neurite outgrowth
β1 integrin-mediated DRG neurite outgrowth
Sensory neurons have been shown to express both α4β1 and α5β1, which are implicated in neurite outgrowth.19,36 Since the neurotrophic effect on DRG neurons was more robust when dASC were plated on laminin, we determined whether β1 integrin was involved in enhanced neurite outgrowth. DRG neurons were seeded on dASC plated with or without laminin coating, and DRG neurons on laminin were used as control. The effect of β1 integrin function-blocking antibodies was compared with untreated neurons and to the neurite outgrowth observed in the presence of an isotype-matched control antibody. In DRG neurons seeded alone on laminin and seeded on dASC, blocking β1 integrin significantly reduced (p<0.001) neurite length when compared with both untreated and isotype control antibody (Fig. 7A). Interestingly, blocking β1 did not significantly reduce the neurite length when DRGs were seeded on dASC plated on laminin (Fig. 7A). The percentage of neurons expressing neurites was also significantly reduced by β1 blocking of DRG seeded on laminin or dASC alone (Fig. 7B). In contrast to measurements of neurite length, the blocking antibody significantly reduced the percentage of neurons expressing neurites in the presence of dASC plated on laminin, but the isotype control had a similar effect (Fig. 7B).
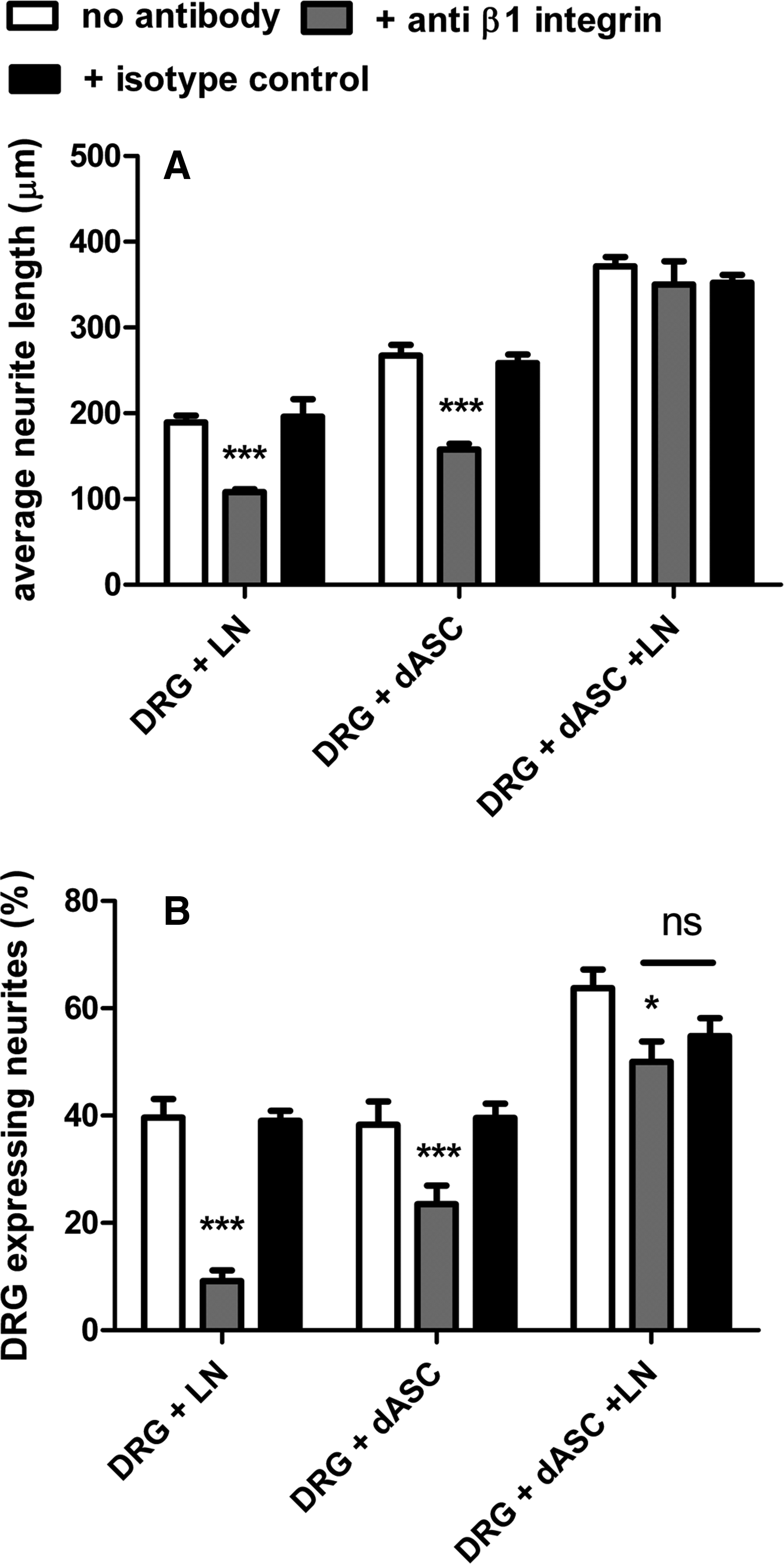
β1 integrin-mediated DRG neurite outgrowth. The effect of function-blocking antibodies against β1 integrin was compared with the untreated neurons and to the neurite outgrowth observed in presence of an isotype-matched controlled antibody.
NGF and BDNF protein levels
Medium from dASC grown on ECM molecules in the absence or presence of DRG neurons was analyzed by NGF and BDNF ELISAs to assess whether differences in the neurotrophic effect of dASC were related to the secretion of these molecules (Fig. 8). The dASC secreted detectable levels of NGF (Fig. 8A) and BDNF (Fig. 8B), which were not significantly changed by plating on either fibronectin or laminin. When dASC were present in direct coculture with DRG neurons there were significantly enhanced levels of both NGF (Fig. 8A) and BDNF (Fig. 8B) compared with DRG medium alone. However, the levels were not significantly different between dASC plated on uncoated or ECM molecule-coated surfaces.
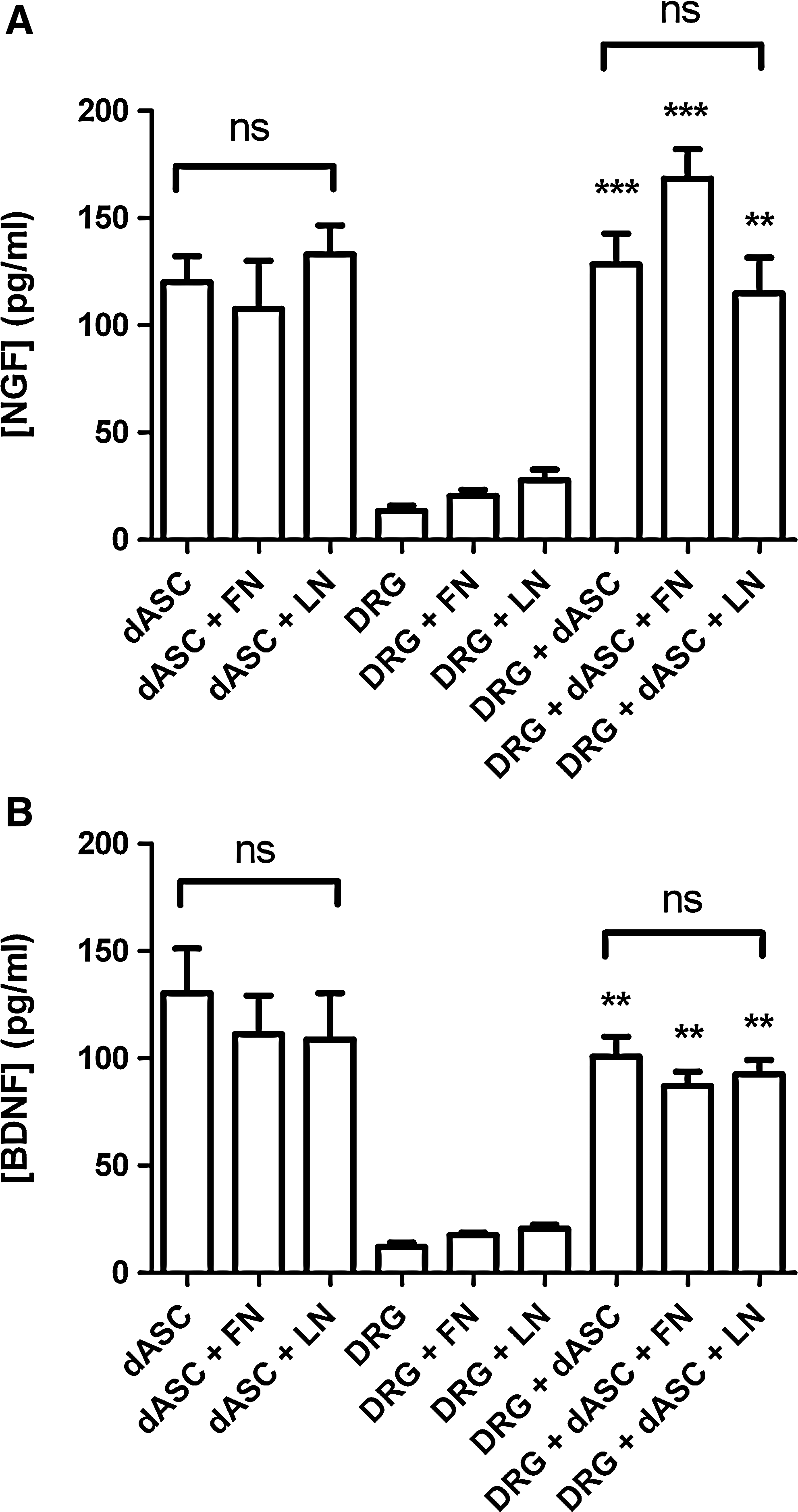
The dASC release neurotrophic factors. After 24 h of dASC culture alone or in direct coculture with DRG neurons on uncoated or ECM molecule-coated surfaces, the medium from the wells was collected and analyzed by ELISA for detection of neurotrophic factors
Discussion
ASC show differentiation along several mesodermal37,38 and nonmesodermal lineages.7,38 Recently, ASC have been differentiated into a SC-like phenotype 10 with capacity to produce myelin in vitro.11,12 Hence, adipose tissue is considered an abundant source of stem cells for regenerative medicine and for nerve repair. The potential of dASC to stimulate axonal growth has also been confirmed. 15 In this study, the interactions between dASC and fibronectin and laminin, and ECM molecules known to play a key role in peripheral nerve regeneration,17,21 were analyzed.
ECM does not influence proliferation of dASC
The proliferation rate of dASC on ECM molecules-coated surfaces showed no significant differences between fibronectin or laminin coating and uncoated TCPS used as a control. This result could be explained by the high affinity of stem cells for TCPS.39,40 Similar studies showed no significant proliferation difference between TCPS and fibronectin coating. 40 Hence, dASC behave differently from primary SC, which show enhanced proliferation on fibronectin and laminin.34,41
ECM molecules protect dASC from induced apoptosis and anoikis
Cell adhesion to ECM is important for survival, differentiation, and migration of cells, 16 and cell attachment to biomaterials and survival is important for regenerative cell therapy. Indeed, cells often encounter an hostile microenvironment when transplanted in the host tissue, due to low oxygen tension and poor nutrient supply,42,43 and high peri-transplantation cell death is an important issue. 44 To assess the potential protective role of ECM molecules against cell death, dASC were seeded on fibronectin- and laminin-coated surfaces and then deprived of serum growth factors. Serum deprivation has been described as an effective model to induce stem cell apoptosis in vitro.43,44 After 48 h of serum deprivation, we evaluated cell viability by live/dead staining, and dASC displayed significantly stronger survival capacity when seeded on either ECM molecules-coated surfaces. Following a 2 h PBS treatment, attachment assays of dASC confirmed a better survival on ECM substrates, possibly related to stronger mechanical stability of the cells, essential for cell migration and controls of signaling environment.16,45 Loss of survival signals from matrix attachments occurring during cell detachment might impair dASC survival.16,44 A specific type of apoptosis, called anoikis, is triggered when cells loose contact with the supportive matrix. 46 A stable attachment to ECM and the presence of growth factors normally synergize to suppress pro-apoptotic factors such as Bmf and Bim, which stimulate Bax (leading to apoptosis) and inhibit Bcl-2 (anti-apoptotic). 46
A previous study showed that ASC have optimal adhesion on poly-caprolactone functionalized with IKVAV laminin-peptide sequence. 47 Protection, or at least delay of the apoptotic cascade, would lead to better cell survival when delivered into a neural guide and, possibly, to better regeneration outcomes. SCs showed improvements in nerve regeneration and viability in vivo when combined with alginate matrix treated with fibronectin. 48 Neural stem cells seeded on laminin and fibronectin tissue-engineered scaffolds showed a slower rate of cell death rate and improved survival post-transplant. 42 These observations indicate that fibronectin and laminin may provide adhesive support to transplanted cells, improving cells microenvironment and mediating subsequent cell signaling events. 42
The ECM regulates the neurotrophic behavior of dASC
Trans-well cocultures showed that ECM-preconditioned dASC could significantly enhance neurite outgrowth of DRG although no significant differences were detected between groups. These results indicate that dASC exert neurotrophic activity via secretion of soluble molecules. In direct contact cocultures, dASC grown on laminin enhanced neurite length outgrowth significantly better than in other groups, a result not observed in the presence of diffusible factors alone, and possibly due to dASC activation by laminin. As significantly higher concentrations of secreted NGF and BDNF were found regardless of ECM coating, neurite elongation may be due to a different mechanism from that due to neurotrophic stimulation by NGF and BDNF. These results confirm the differential sensitivity of neurons to fibronectin and laminin, 19 with a higher percentage of DRGs expressing neurites on laminin when compared with fibronectin. Also, contact between dASC and neurons potentiates neurite elongation once neurite growth has been initiated in response to diffusible factors. 34 When compared with primary SCs, dASC grown on laminin evoked similar levels of neurite outgrowth in the trans-well cocultures but shorter neurites when in direct contact. This indicates that the dASC produce significant levels of soluble neurotrophic growth factors but perhaps lack some of the SC expressed surface cell adhesion molecules that contribute to axon elongation.
It has been shown that laminin can synergistically signal with the growth cones and SCs influencing regrowth.21,49 ECM signals to neurons and SCs largely depend on integrins, a family of heterodimeric receptors that modulate axonal guidance, adhesion, and migration.19,50 Several laminin receptors are present in mature SCs, but the β1 subfamily of integrins have been shown to play a key role in nerve regeneration, being necessary for SCs-axons interactions and myelination.51–54 Consistent with previous studies on the primary role the β1 subfamily of integrin in axonal sorting,51,52,54 blocking of β1 integrins did not significantly affect adhesion of neither stem cells nor DRGs, but dramatically reduced neurite outgrowth of DRGs alone on laminin, or of DRG in the presence of dASC without laminin coating. However, this effect was reversed when dASC were seeded on laminin, suggesting that laminin preconditioning of dASC can enhance the neurotrophic effect of dASC and also over-ride β1 mediated effects on neurite outgrowth.
SCs development and commitment is driven by ECM via integrins, and integrin signaling regulates SC function.23,49 Similarly, laminin might interact with different integrins on the dASC cell surface, which could result in activation of different pathways and amplification of neurotrophic effect. ECM can modulate cell surface expression and localization of integrins. 19 Laminin preconditioning of dASC may overcome the functional block of β1 integrin receptors by increasing these receptors, possibly changing their conformational shape or finding alternative integrins to allow cell–cell- and cell-axonal/neurite interaction. These mechanisms of redundancy and compensation were detected in β1-null SCs, 55 suggesting how other laminin receptors like dystroglycans, α6β4, α6β1, and α7β1 could possibly cooperate to myelinate axons.55,56 Upregulation of β1 or of other laminin- and fibronectin-binding integrins was also shown after induced injury promoting neuronal repair.19,57 Integrin trafficking in response to laminin could, in this way, improve the contacts between dASC and the growing axons, partially bypassing the inhibited contact between neurons and ECM. Also, increased integrin signaling may lead to increased secretion of neurotrophic factors or upregulation of other adhesion molecules like L-1 and N-cadherin, finally improving neurite extension.
Taken altogether, these data show how the ECM molecules can significantly improve the potential of dASC for nerve regeneration. ECM-preconditioning of dASC resulted in improved survival and adhesion in stress conditions, Moreover, ECM molecules and laminin substrate could improve the neurotrophic activity of dASC, an effect we suggest to be independent of β1 integrin function alone. Importantly, it can be concluded that, in general, the in vitro neurotrophic effects of laminin-conditioned dASC are comparable with primary SCs.
Footnotes
Acknowledgments
The authors are thankful to Dr Cristina Mantovani (Blond McIndoe Research Laboratories, University of Manchester, United Kingdom) for help and assistance throughout this study and to Dr Alessandro Faroni (Blond McIndoe Research Laboratories, University of Manchester, United Kingdom) for critical advices and helpful discussions in experimental planning and methodology. The authors are grateful to the Swiss Society of Plastic Surgery and to the Swiss National Fund (Fonds National Suisse de la Recherche Scientifique) for the scholarships that supported the work of P.G. di Summa. This study was also supported by funds from the Swedish Medical Research Council, European Union, Umeå University, County of Västerbotten, Åke Wiberg Stiftelse, Magnus Bergvalls Stiftelse, Clas Groschinskys Minnesfond and the Gunvor and Josef Aner Foundation. Finally, we would like to thank Acorda Therapeutics for the generous supply of GGF-2.
Disclosure Statement
No competing financial interests exist.