
Abstract
We have previously shown that mesenchymal stem cells (MSCs) from patients with osteoarthritis (OA) constitutively express type X collagen, a marker of late-stage chondrocyte hypertrophy, osteogenic marker genes, including alkaline phosphatase (ALP), bone sialoprotein (BSP), and osteocalcin (OC), and chondrogenesis marker gene aggrecan (ACAN). As patients with arthritis often take nonsteroidal anti-inflammatory drugs (NSAIDs) and acetaminophen (Acet), the purpose of the study was to assess whether these drugs can affect the gene expression of human MSCs. MSCs isolated from the bone marrow of patients with OA or normal donors were cultured without (control) or with Acet or NSAIDs, which include ibuprofen, diclofenac (Dic), naproxen, and celebrex. After 3 days of culture, the expression of type X collagen alpha 1 (COL10A1), ACAN, COL1A1, as well as ALP, BSP, OC, and Runt-related transcription factor 2 was analyzed by real-time reverse transcription (RT)–polymerase chain reaction. The results showed that COL10A1 and the osteogenic and chondrogenic marker genes can be regulated by NSAIDs and Acet in normal MSCs. In contrast, Acet did not significantly affect COL10A1 expression in OA MSCs, while Dic is the only drug that had no significant effect on all markers in normal MSCs. The upregulation of COL10A1 in normal MCSs by Acet and Npx may explain why stem cells from patients with OA express COL10A1 constitutively. This knowledge may help in designing better strategies for stem cell differentiation into chondrocyte-like cells, from this source, with Dic being a viable option for treating OA pain, with an eye toward preventing the potential to enhance calcification in the repair of cartilage and degenerated intervertebral discs.
Introduction
Mesenchymal stem cells (MSCs) are multipotent stem cells that can be isolated, expanded, and stimulated to differentiate into a variety of cells, including osteoblasts, chondrocytes, myocytes, adipocytes, and beta-pancreatic islets cells.8–11 Recent studies have been directed toward differentiation of MSCs taken from patients with OA, as the stem cells can be employed in biological repair of cartilage or disc lesions.12,13 A major drawback of current cartilage and disc tissue engineering is that human MSCs from patients with OA rapidly express COL X.14–16 COL X, a marker of chondrocyte hypertrophy, is implicated in endochondral ossification.6,17,18 Moreover, these MSCs also express alkaline phosphatase (ALP), bone sialoprotein (BSP), and osteocalcin (OC),19,20 as well as Runt-related transcription factor 2 (RUNX2).21,22 Furthermore, previous studies showed that COL X was expressed before COL II in some cases.23,24 This was surprising since it is well known that stem cells differentiate into chondrocytes before they become hypertrophic. It was not clear whether COL X expression in MSCs is related to drug therapy. 25
Clinical recommendations for the sympathetic treatment of OA include acetaminophen (Acet) and nonsteroidal anti-inflammatory drugs (NSAIDs), including ibuprofen (Ibu), diclofenac (Dic), naproxen (Npx), and celebrex (Cele).26,27 Despite the extensive use of these drugs, a clear understanding of their mechanisms is still lacking. Acet is used as an over-the-counter analgesic (pain reliever) and antipyretic (fever reducer), for the relief of aches and pains associated with arthritis. Although Acet is used to treat inflammatory pain, it is not generally classified as an NSAID, because it exhibits only weak anti-inflammatory activity. To date, its mechanism of action proposed is the inhibition of cyclooxygenase (COX) with high selectivity for COX-2. Ibu is an NSAID used for relief of fever and pain such as in arthritis. It acts as a vasoconstrictor, mainly because it inhibits the vasodilating prostacyclin produced by COX-2 enzymes. Dic is an NSAID taken to reduce inflammation and pain, especially arthritis and rheumatoid arthritis. The exact mechanism of action is not entirely known, but the primary mechanism responsible for its anti-inflammatory, antipyretic, and analgesic action is thought to be inhibition of prostaglandin (PG) synthesis by inhibition of COX. Npx is an NSAID used for the reduction of pain, fever, inflammation, and stiffness caused by arthritis, rheumatoid arthritis, migraine, and ankylosing spondylitis. Npx works by inhibiting COX enzymes. Cele is an NSAID and selective COX-2 inhibitor used in the treatment of OA, rheumatoid arthritis, menstruation, and acute pain. It was originally intended to relieve pain while minimizing the gastrointestinal adverse effects usually seen with conventional NSAIDs. Its primary indication is in patients who need long-term pain relief.
The specific aim of this study was to determine the effects of these drugs on gene expression of MSCs from normal and OA donors.
Materials and Methods
Source and isolation of stem cells
OA human MSCs (OA MSCs) were obtained from the aspirates of the intramedullary canal of donors (60–80 years of age) undergoing total hip replacement for OA, with a protocol approved by the Research Ethics Committee of the Jewish General Hospital. Bone marrow aspirates were processed as previously described.15,28 Normal human MSCs were obtained from Lonza. They were harvested and cultured from normal human bone marrow. Cells were tested for purity by flow cytometry and for their ability to differentiate into osteogenic, chondrogenic, and adipogenic lineages. Cells were positive for CD105, CD166, CD29, and CD44 and were negative for CD14, CD34, and CD45. All cells were expanded in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS), 100 U/mL penicillin, and 100 μg/mL streptomycin and were used within four passages.
Culture of stem cells with drugs
Normal human MSCs were cultured for up to 72 h in the absence (control) or the presence of drugs (all from Sigma-Aldrich). The drugs were dissolved in DMEM with a concentration based on the blood concentrations.29–33 Thus, the concentrations of Npx, Ibu, Dic, Cele, and Acet were 100 μg/mL, 300 μg/mL, 5 μg/mL, 800 ng/mL, and 50 μg/mL, respectively.
Total RNA isolation
Cells were harvested at day 3 for gene expression studies. Total RNA was extracted from MSCs using a modification of the method of Chomczynski and Sacchi 34 using Trizol reagent (Invitrogen). Briefly, after centrifugation for 15 min at 12,000 g at 4°C, the aqueous phase was precipitated in one volume of isopropanol, incubated for 45 min at −20°C, and centrifuged again for 15 min at 12,000 g at 4°C. The resulting RNA pellet was washed with 75% ethanol, and then centrifuged and air-dried. Then, the pellets were suspended in 50 μL of diethylpyrocarbonate (DEPC)-treated water and assayed for RNA concentration by measuring A260 and purity by calculating A260/A280.
Reverse transcription
First, 1 μg total RNA isolated from the cells was digested with DNase I according to the Invitrogen protocol. Then, the purified RNA reverse-transcribed using the chemicals from Invitrogen according to the protocol of the company. One microgram RNA was mixed with random primers (final concentration 0.15 μg/μL), dNTP mixture (final concentration 0.5 mM), and DEPC-treated distilled water with a total volume of 12 μL. After the solution was incubated at 65°C for 5 min, it was mixed with a first-strand buffer, Dithiothreitol, RNaseOUT, and Superscript II reverse transcriptase with a final volume of 20 μL. Then, the solution was incubated at 45°C for 50 min to perform the reverse transcription and then in 70°C for 15 min to inactivate the reverse transcriptase.
LightCycler real-time polymerase chain reaction
For LightCycler real-time polymerase chain reaction (PCR), a master mix of the following reaction components was prepared with the final concentrations as 10 μL SYBER PCR master mix (×1; Qiagen), 8 μL distilled water, 0.5 μL forward primer (0.25 μM), and 0.5 μL reverse primer (0.25 μM). To each 19 μL master mix, 1 μL of cDNA was mixed as a PCR template. The sequences of primers are in Table 1. The reaction conditions included one cycle of PCR initial activation step (95°C for 15 min, 20°C/s ramp rate), 45 cycles of amplification and quantification (94°C for 15 s, 57°C for 30 s, 72°C for 30 s), one cycle of melting curve (65°C–95°C with heating rate of 0.1°C/s with a continuous fluorescence measurement), and a final cooling step to 4°C. The crossing points were determined by Light Cycler software 3.3 (Roche Diagnostics) and were measured at a constant fluorescence level.
35
Every sample was run in duplicate, and the average value was employed in the calculation. The relative ratio of gene transcription was determined by the following equation, and glyceraldehyde 3-phosphate dehydrogenase was applied as the reference gene:
Rationale for genes analyzed
Type X collagen alpha 1 (COL10A1) message levels were examined after drug treatment, because it is a marker of late-stage chondrocyte hypertrophy.36,37 Message levels of ALP were examined, because calcification involves deposition of inorganic calcium phosphates in a crystalline form, 38 and ALP plays an active role in initiating the calcification process.39,40 BSP and OC have been used as bone-specific markers,19,20 while RUNX2 is a key transcription regulator associated with osteoblast differentiation.21,22 ACAN is used as marker of chondrogenesis. 11 COL1A1 is expressed by MSCs. 15
Statistical analysis
Statistical analysis was performed using analysis of variance (ANOVA), followed by Fisher's protected least significant difference post hoc test using Statview (SAS Institute, Inc.). Four samples from four experiments were used in this study and were assessed on 80% power to detect a difference in gene expression. Results are presented as the mean±standard deviation of four experiments. Differences were considered statistically significant with p<0.05.
Results
Baseline expression of all the markers in normal and OA MSCs
The relative ratio of baseline gene expression of all the markers in OA MSCs compared with normal MSCs after they were cultured in the control media for 3 days is shown in Figure 1. ALP and BSP are prominent in OA MSCs, followed by COL10A1, ACAN, and OC, when compared to MSCs from normal donors. Similar expression levels of COL1A1 were observed in OA MSCS when compared to those in normal MSCs, while only RUNX2 gene expression was lower in OA MSCs.
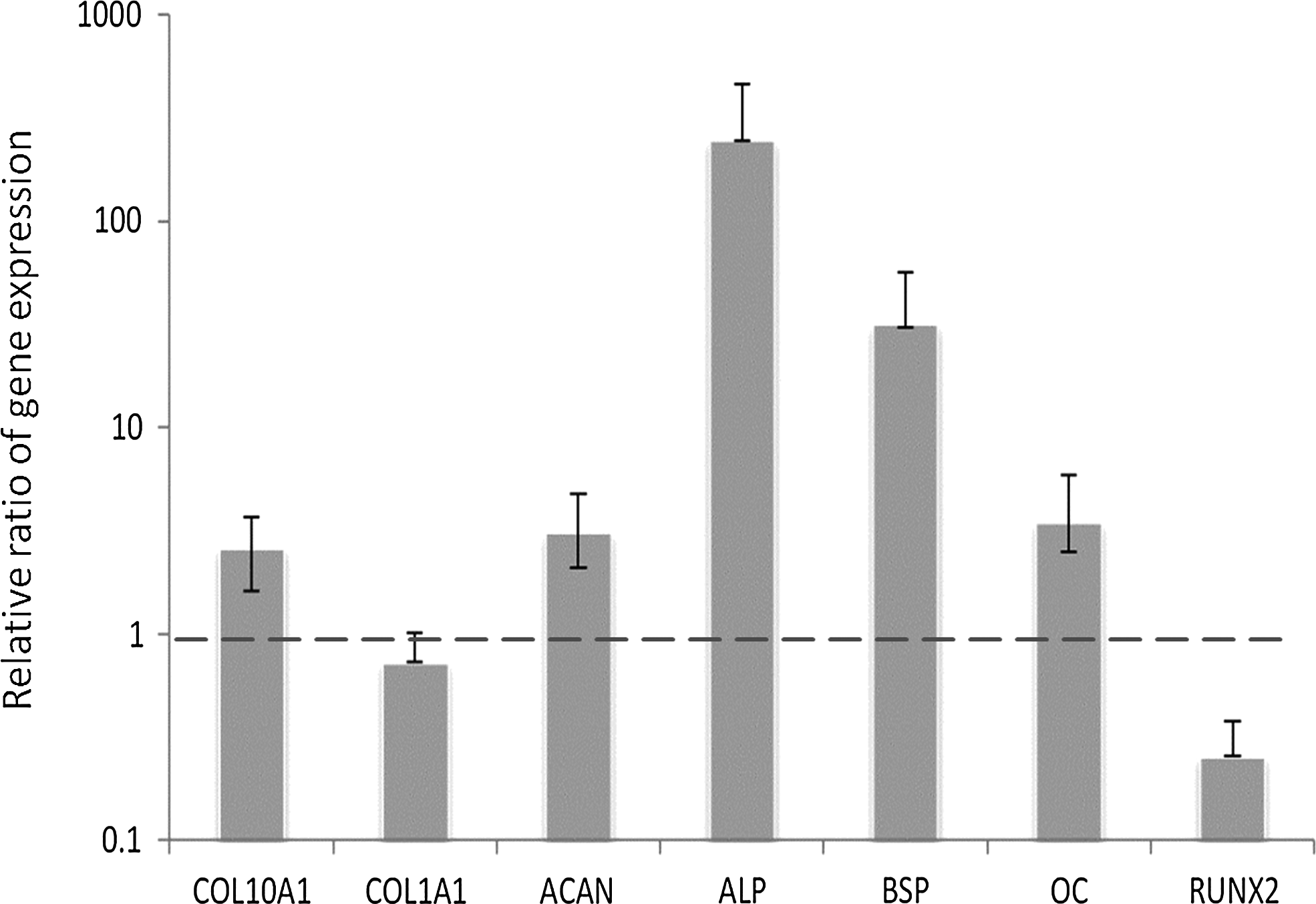
The relative ratio of baseline gene expression of all the markers in osteoarthritic mesenchymal stem cells (OA MSCs) compared with normal MSCs after they were cultured in high-glucose Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum and antibiotics for 3 days. Gene expression was measured by real-time reverse transcription–polymerase chain reaction (RT-PCR). Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as housekeeping gene and served to normalize the results. Values represent the mean±standard deviation of four experiments.
Effect of drug treatments on gene expression in normal and OA MSCs
In normal MSCs, Acet (p=0.02) and Npx (p<0.0001) supplementation led to a significant increase in COL10A1 expression in normal MSCs, when compared to control (Fig. 2). In contrast, treatment with Ibu, Dic, and Cele did not have a significant effect in gene expression of COL10A1. In OA MSCs, only Npx (p=0.0096) supplementation led to a significant increase in COL10A1 expression compared to control. In contrast, treatment with Acet, Ibu, Dic, and Cele did not have a significant effect on gene expression of COL10A1.

Changes of type X collagen alpha 1 (COL10A1) expression in normal and OA MSCs cultured for 3 days in the absence of drugs (Ctl) or presence of drugs. Gene expression was measured by real-time RT-PCR. GAPDH was used as a housekeeping gene and served to normalize the results. Values represent the mean±standard deviation of four experiments. *p<0.05, **p<0.01, ***p<0.001 compared with control.
In normal MSCs, Cele supplementation led to a significant increase in ALP expression (p<0.001) when compared to control (Fig. 3). In contrast, Acet, Ibu, and Npx supplementation led to a significant decrease in ALP expression (p<0.001) when compared to control. Dic had no significant effect (p=0.06). In OA MSCs, Npx supplementation led to a significant decrease in ALP expression (p=0.009) when compared to control. In contrast, Acet, Ibu, Dic, and Cele supplementation had no significant effect when compared to control.
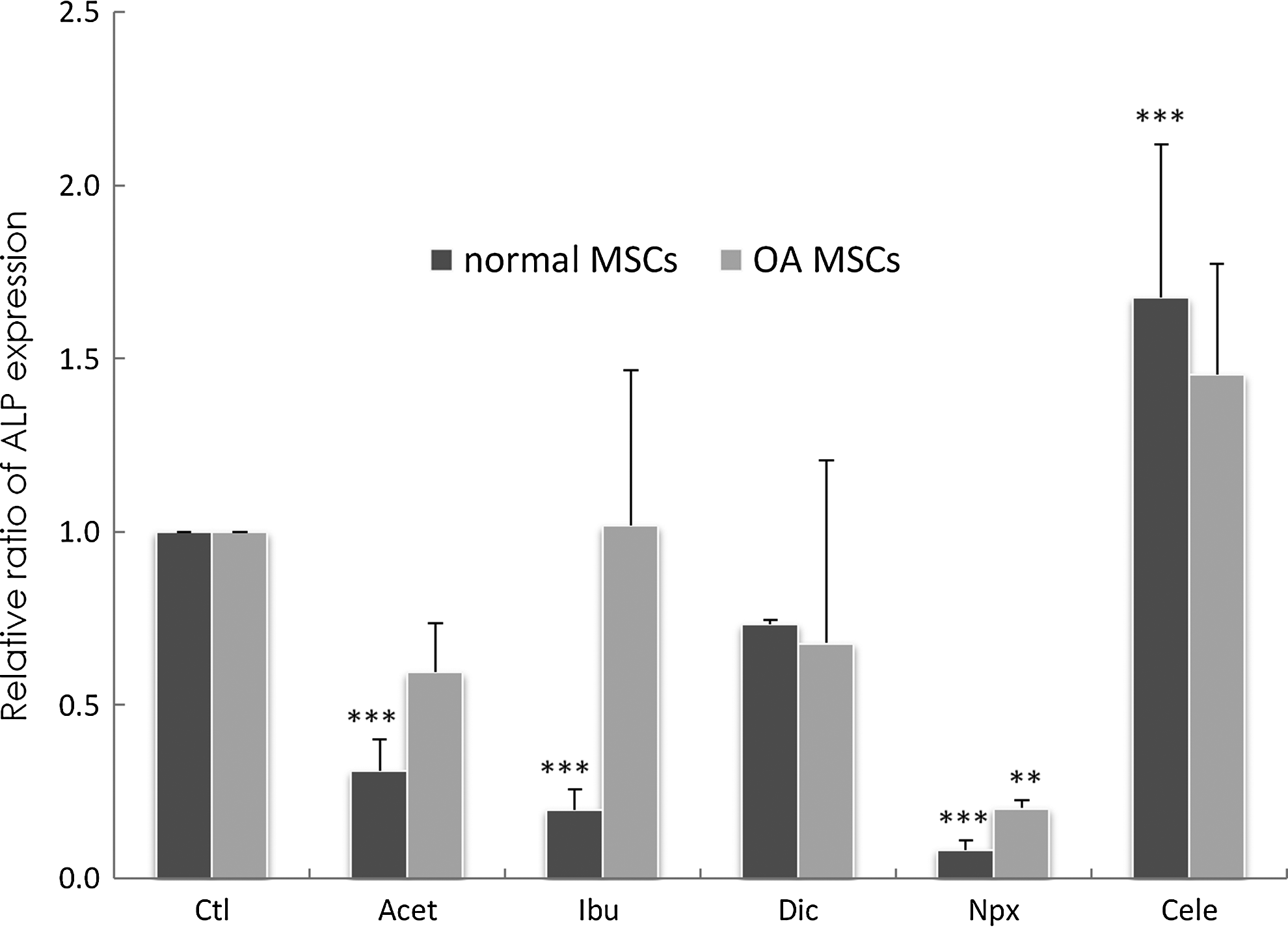
Changes of the alkaline phosphatase (ALP) gene expression in normal and OA MSCs cultured for 3 days in the absence of drugs (Ctl) or presence of drugs. Gene expression was measured by real-time RT PCR. GAPDH was used as a housekeeping gene and served to normalize the results. Values represent the mean±standard deviation of four experiments. **p<0.01, ***p<0.001 compared with control.
In normal MSCs, Acet supplementation decreased the expression of BSP (p=0.04), whereas Ibu, Dic, Npx, and Cele had no significant effects on BSP expression, although Dic had a tendency to enhance BSP expression (Fig. 4). In OA MSCs, Dic supplementation led to a significant decrease in BSP expression (p=0.0062) when compared to control. In contrast, Acet, Ibu, Npx, and Cele supplementation had no significant effects when compared to control.

Changes of the bone sialoprotein (BSP) gene expression in normal and OA MSCs cultured for 3 days in the absence of drugs (Ctl) or presence of drugs. Gene expression was measured by real-time RT-PCR. GAPDH was used as a housekeeping gene and served to normalize the results. Values represent the mean±standard deviation of four experiments. *p<0.05, **p<0.01 compared with control.
In normal MSCs, Acet (p<0.001) and Npx (p=0.0025) supplementation led to a significant increase in OC expression when compared to control (Fig. 5). In contrast, addition of Ibu, Dic, and Cele had no significant effect on OC gene expression. In OA MSCs, Acet (p=0.013) and Npx (p=0.025) supplementation led to a significant increase in OC expression when compared to control. Ibu, Dic, and Cele had no significant effects on OC when compared to control.
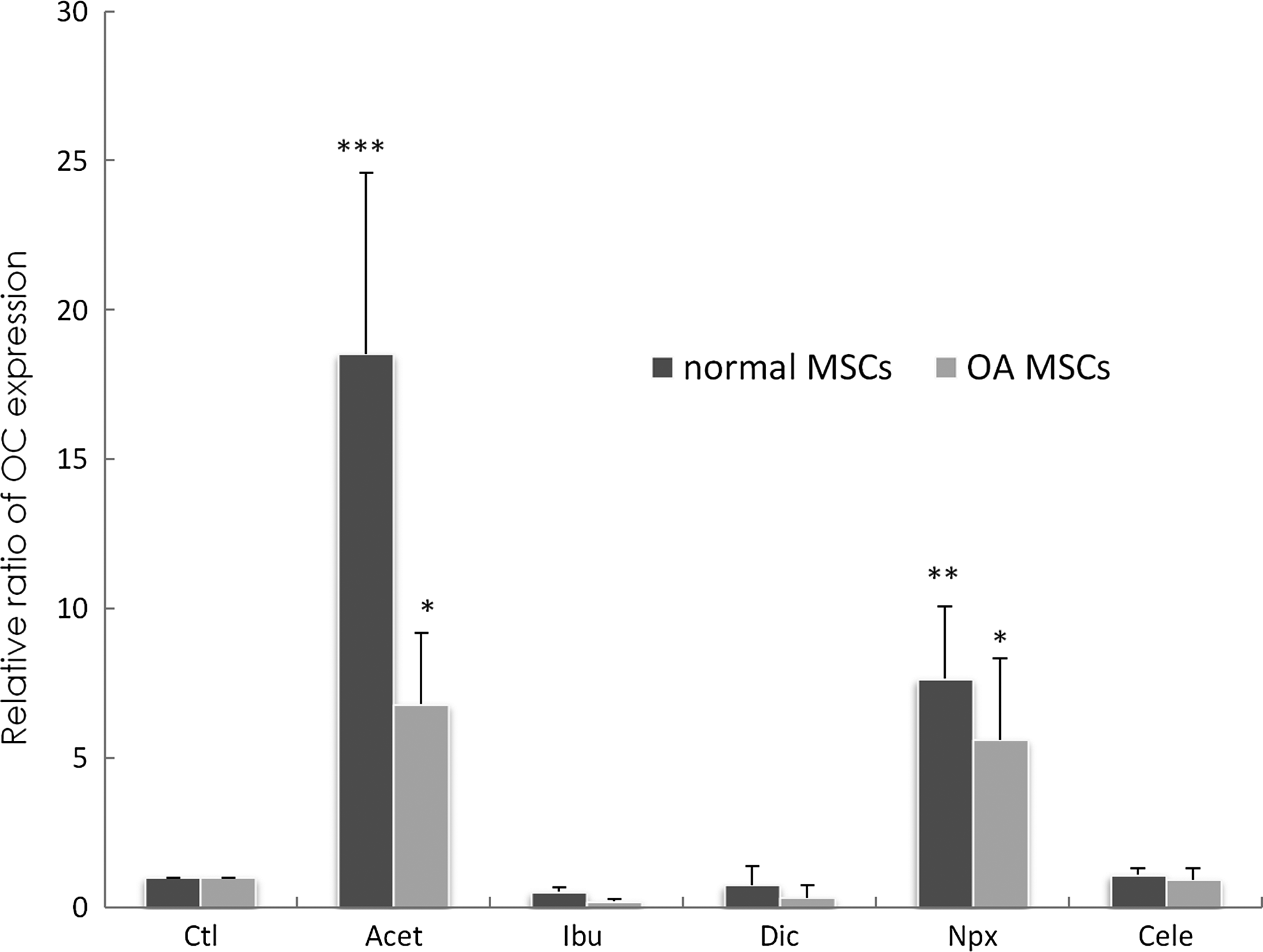
Changes of osteocalcin (OC) gene expression in normal and OA MSCs cultured for 3 days in the absence of drugs (Ctl) or presence of drugs. Gene expression was measured by real-time RT-PCR. GAPDH was used as a housekeeping gene and served to normalize the results. Values represent the mean±standard deviation of four experiments. *p<0.05, **p<0.01, ***p<0.001 compared with control.
In normal MSCs, the addition of Npx led to a significant increase in RUNX2 expression (p=0.0003) when compared to control (Fig. 6). In contrast, addition of Acet, Ibu, Dic, and Cele had no significant effect on RUNX2 gene expression after 3 days of exposure. In OA MSCs, Npx supplementation led to a significant increase in RUNX2 expression (p=0.035) when compared to control. In contrast, Ibu (p=0.036) and Dic (p=0.047) supplementation decreased RUNX2 expression significantly. Acet and Cele had no significant effects on RUNX2 gene expression after 3 days of exposure.
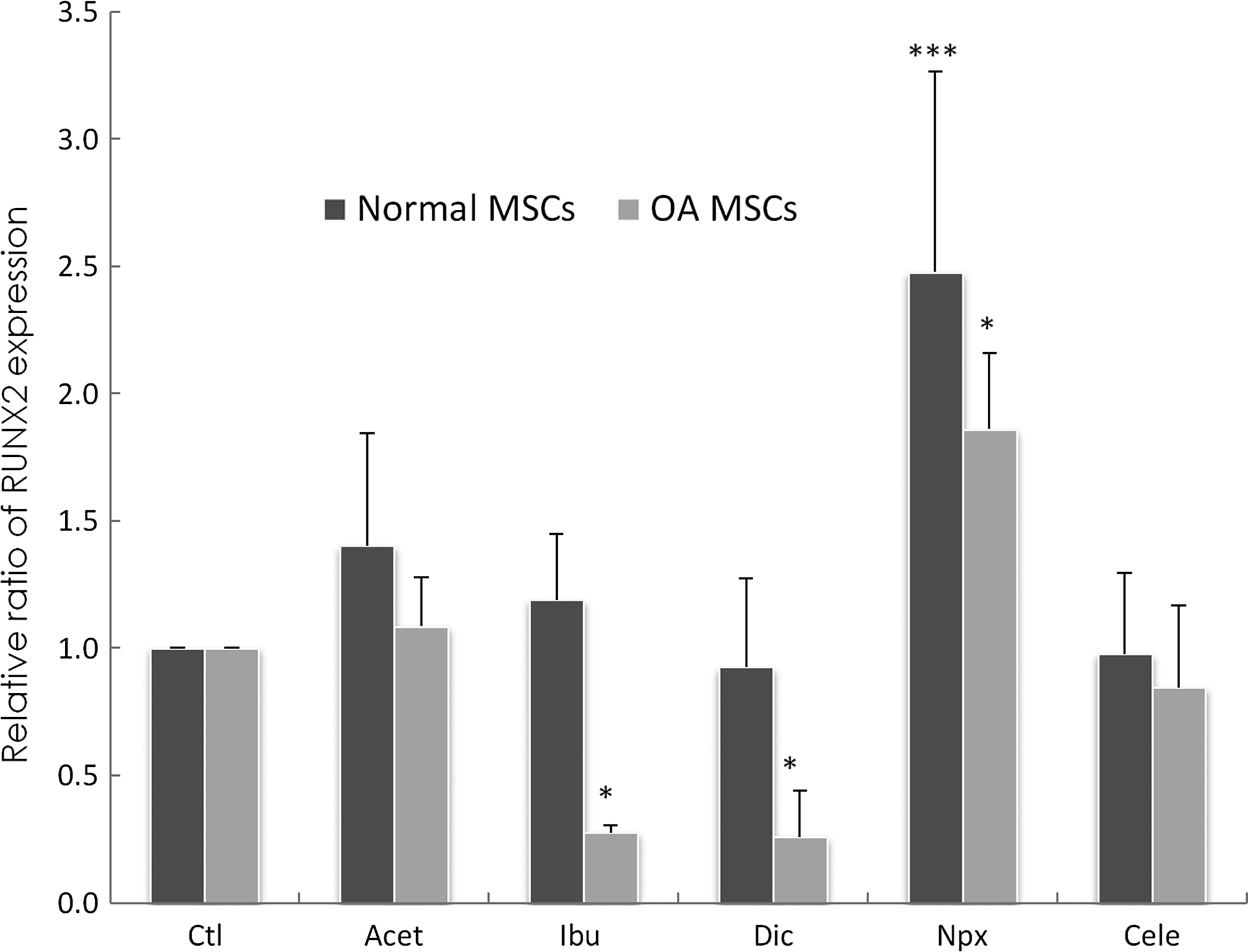
Changes of runt-related transcription factor 2 (RUNX2) gene expression in normal MSCs cultured for 3 days in the absence of drugs (Ctl) or presence of drugs. Gene expression was measured by real-time RT-PCR. GAPDH was used as a housekeeping gene and served to normalize the results. Values represent the mean±standard deviation of 4 experiments. *p<0.05, ***p<0.001 compared with control.
After culturing normal and OA MSCs for 3 days in the absence of drugs, ACAN expression could be detected in normal MSCs (Fig. 7). With Acet, Ibu, and Npx supplementation, ACAN message levels were decreased significantly (p<0.001). Addition of Cele increased ACAN gene expression significantly (p<0.001) after 3 days of exposure, whereas Dic showed no significant effect. In OA MSCs, with Acet (p=0.01), Dic (p=0.0005), and Npx (p=0.002) supplementation, ACAN message levels were decreased significantly. Cele increased ACAN expression significantly (p=0.04). Ibu had no significant effect while a tendency toward a decrease was observed.
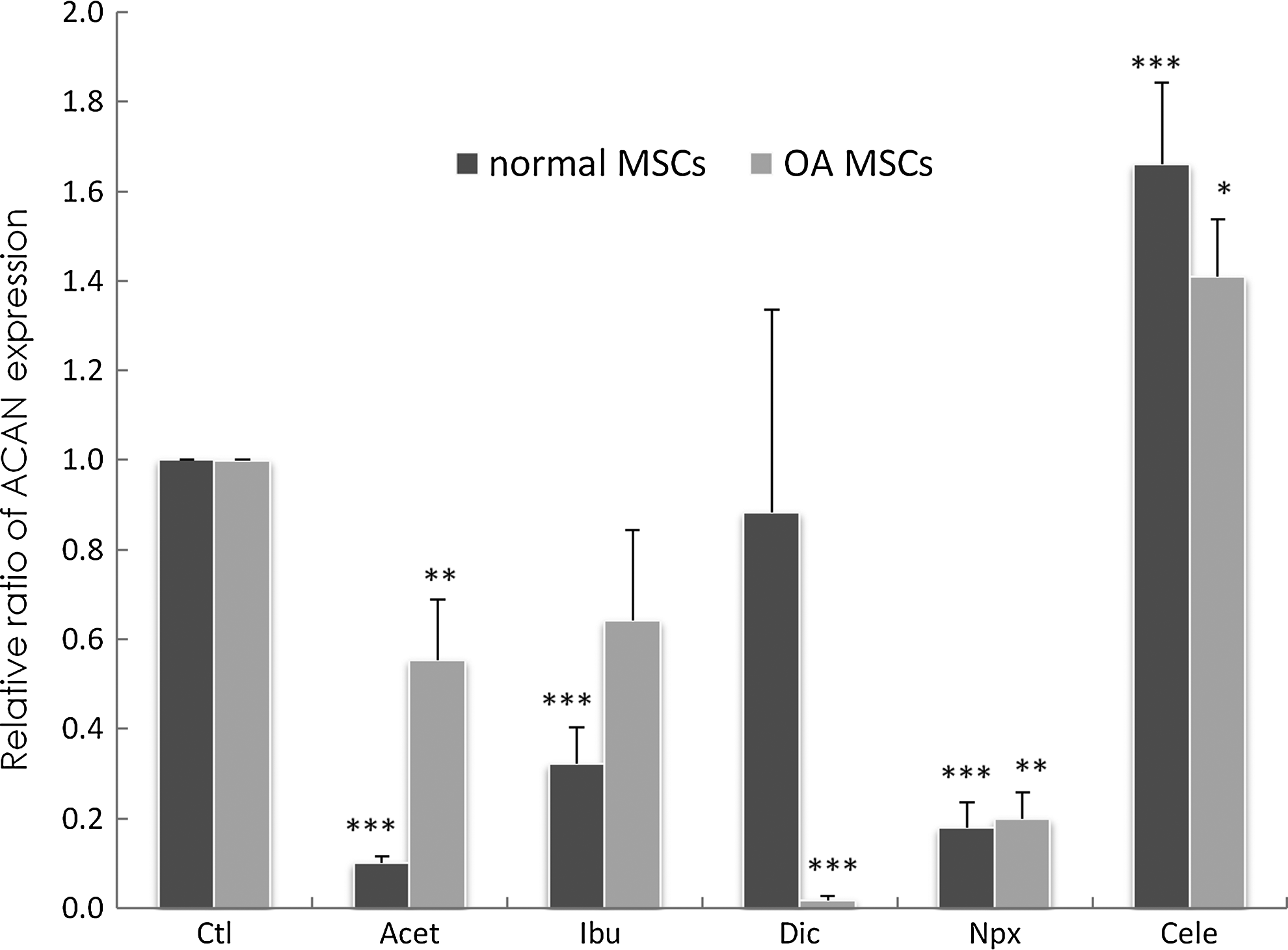
Changes of aggrecan (ACAN) gene expression in normal and OA MSCs cultured for 3 days in the absence of drugs (Ctl) or presence of drugs. Gene expression was measured by real-time RT-PCR. GAPDH was used as a housekeeping gene and served to normalize the results. Values represent the mean±standard deviation of four experiments. *p<0.05, **p<0.01, ***p<0.001 compared with control.
COL1A1 message was also present in the absence of drugs (Fig. 8). In normal MSCs, in the presence of Acet (p=0.025), Ibu (p=0.01), and Npx (p=0.002), significant decreased COL1A1 message levels were observed, whereas Cele significantly increased COL1A1 expression (p=0.0035), and Dic supplementation had no effect (Fig. 8). In the presence of Acet (p=0.035), Ibu (p=0.047), Dic (p=0.0001), and Npx (p=0.004), decreased COL1A1 message levels were observed while Cele increased COL1A1 expression significantly (p=0.03).
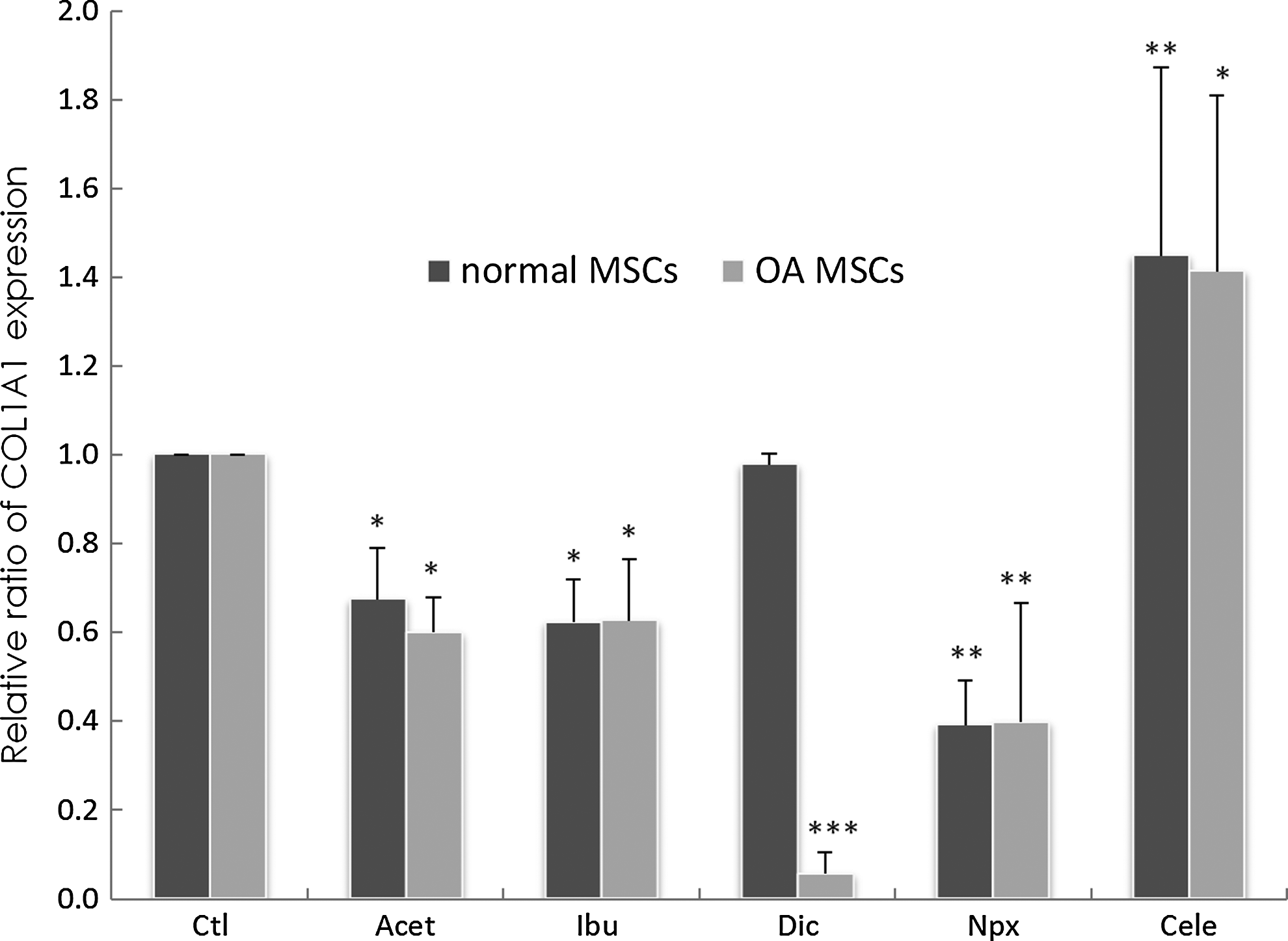
Changes of type I collagen (COL1A1) gene expression in normal MSCs cultured for 3 days in the absence of drugs (Ctl) or presence of drugs. Gene expression was measured by real-time RT-PCR. GAPDH was used as a housekeeping gene and served to normalize the results. Values represent the mean±standard deviation of four experiments. *p<0.05, **p<0.01, ***p<0.001 compared with control.
Discussion
Previous studies have shown that ACAN is constitutively expressed by MSCs from patients with OA, and COL X is expressed as an early event. 25 Further, we showed that MSCs from patients with OA express not only COL X but also osteogenic differentiation markers such as ALP, BSP, OC, and the transcription regulator RUNX2. 15 The present data indicate that Acet and NSAIDs can affect the expression of COL X, ALP, BSP, OC, COL1A1, and ACAN in cultured normal MSCs. In contrast, Acet did not significantly affect COL10A1 expression in OA MSCs. Dic is the only drug that had no significant effect on all markers in normal MSCs, while it affected BSP, RUNX2, ACAN, and COL1A1 in OA MSCs. Although the reason for this is unclear, Dic effects in normal MSCs are features needed for any agent designed to relieve pain, but not have an effect on MSCs. Therefore in principle, Dic could be a viable option for treating OA pain to prevent the potential to enhance calcification in the repair of cartilage and degenerated IVDs.
Previously, we have shown that COL I, but not COL II, was detected in MSCs from patients with OA.18,37 Similar findings were observed in this study. It is important to note that COL II was not investigated in this study as a marker of chondrogenesis, because it is not expressed by MSCs from patients with OA without transforming growth factor β1 or β3 supplementation. 25
All NSAIDs can be found in synovial fluid after repeated dosing. NSAIDs with short half-lives tend to stay for much longer than predicted by their half-lives, while NSAIDs with longer half-lives disappear from synovial fluid at a rate proportional to their half-lives. 41 Thus, it is possible that these drugs can affect MSC characteristics in patients with OA, which leads to different gene expression levels in OA MSCs.
The anti-inflammatory activity of NSAIDs is mediated chiefly through inhibition of COX enzyme activity, which diminishes the biosynthesis of PGs. 42 Ibu, Npx, and Dic are nonselective COX inhibitors. Cele is a selective COX-2 inhibitor and has about 375 times more selectivity for COX-2 than for COX-1.32,41 Acet is a weak inhibitor of COX activity with weak anti-inflammatory activity. 43 It is not clear if this may explain why Acet and Npx supplementation enhanced COL X gene expression in normal MSCs, while only Npx stimulated COL X expression in MSCs from patients with OA.
Although they differ in their modes of action, as discussed above, both Acet and Npx are COX inhibitors. While Npx and Ibu inhibit both COX-1 and COX-2, Acet only inhibits COX-2. This is contrary to what was found in a previous study using the COX inhibitor 5-Nitro-2-(3-phenylpropylamino) benzoic acid (NPPB). 44 In the previous study, human MSCs were cultured in a standard medium supplemented with the COX inhibitor NPPB. The results indicated that COL X expression was inhibited in OA MSCs. Similarly, the specific COX-1 (COX-1: SC-560) and 5-lipoxygenase (5-LOX: MK-866) inhibitors suppressed COL X expression. However, the same study implied that neither the COX-2 nor 12-LOX (baicalein) inhibitors affected COL X expression. Thus, other signaling mechanisms are likely to be involved.
Recently, we reported that COL X expression was upregulated in MSCs from rabbits in a surgical instability model of OA (anterior cruciate ligament transection) when compared to control healthy rabbit MSCs. 45 Thus, COL X expression in OA is a common phenomenon that can be due to the disease process itself and can be upregulated by Acet and Npx supplementation. It is therefore critical to understand the MSC phenotype in patients with OA, as these cells are clinically important for biological repair of cartilage lesions using autologous stem cells.
Of special interest here was that Acet, Ibu, and Npx supplementation led to a significant decrease in ALP expression when compared to control in normal MSCs. However, only Acet and Npx supplementation led to a significant decrease in ALP expression in MSCs from patients with OA. Although only ALP gene expression was measured in these studies, other work has demonstrated that these cells express ALP activity.
The present work indicates that Acet and Npx can promote OC expression in MSCs from patients with OA. While statistical significance is achieved in some expression levels, the biological significance of these small changes in gene expression may await further analysis. The fact that there is a statistically significant difference indicates only that the difference is unlikely to have occurred by chance. Thus, since only COL X, OC, and RUNX2 were upregulated by Acet and Npx, but not ALP or BSP, in normal MSCs, it raises the question whether these markers have a serendipitous expression, but with no functional consequence.
Previously, it was found that some NSAIDs had no significant effect on net synthesis of either glycosaminoglycans or protein in organ cultures of femoral condylar cartilage, and did not increase the proportion of newly synthesized glycosaminoglycans recovered from the culture medium, suggesting that they had no direct effect on the integrity of the extracellular matrix. 46 However, in this study, Acet and NSAIDs affected ACAN expression in MSCs. As gene expression was only studied on MSCs cultured for 3 days, the effects of drugs on MSCs for a longer term need further research.
One of the limitations of this study is that while we provided baseline gene expression for MSCs from patients with OA and the effect of drug therapy on MSCs from normal and OA, a more complete analysis for other variables such as age, gender, isolation sites, and disease status (osteoarthritis/rheumatoid arthritis) will await further studies. Further, we did not perform group comparisons between the two cell types. Ideally, a two-way ANOVA would be employed for examining the effects of disease condition (OA vs. Normal) and the drug treatments. However, we could not obtain the medical records of the patients with OA, as this information is in local clinics across the country with various general practitioners and are only referred to the hospital for surgery. Thus, we could not determine if there is a significant interaction effect between the drugs. Finally, the signaling pathways activated by these drugs and other methods such as microarrays to determine what other genes are activated are under study. In this work, only real-time reverse transcription-polymerase chain reaction (RT-PCR) was used.
Conclusions
The results showed that NSAIDs and Acet in normal MSCs could regulate COL10A1 and the osteogenic and chondrogenic marker genes. The upregulation of COL10A1 by Acet and Npx may explain why stem cells from patients with OA express COL X constitutively. Thus, this knowledge may help in designing better strategies for stem cell differentiation into chondrocyte-like cells, for use in tissue engineering from this source.
Authors' Contributions
A.A. performed the experiments, acquired data, and performed statistical analysis, as well as wrote the manuscript. H.T.W. designed the primers, developed the real-time PCR conditions, and revised the manuscript. O.C. has organized the figures. S.A.L.R. and S.R. contributed to the cell isolation and culture. J.A. made substantial contributions to the study design and revised the manuscript. F.M. conceived and supervised the whole study and finished writing the manuscript. All authors read and approved the final manuscript.
Footnotes
Disclosure Statement
The authors declare that they have no competing interests.