
Abstract
Three-dimensional (3D) culture systems have been introduced to provide cells with a biomimetic environment that is similar to in vivo conditions. Among the polymeric molecules available, sodium-alginate (Na-alg) salt is a material that is currently employed in different areas of drug delivery and tissue engineering, because it offers biocompatibility and optimal chemical properties, and its gelation with calcium chloride provides calcium-alginate (Ca-alg) scaffolds with mechanical stability and relative permeability. In this work, four different preparations of Ca-alg beads with varying Na-alg viscosity and concentration were used for a human hepatoma cell line (Huh-7) encapsulation. The effects of Ca-alg bead preparation on structural cell organization, liver-specific functions, and the expression of specific receptors implicated in hepatotropic virus permissivity were evaluated. Hepatic cells were cultured in 500 μm diameter Ca-alg beads for 7 days under dynamic conditions. For all culture systems, cell viability reached almost 100% at day 7. Cell proliferation was concomitantly followed by hepatocyte organization in aggregates, which adopted two different morphologies (spheroid aggregates or multicellular channel-like structures), depending on Ca-alg bead preparation. These cellular organizations established a real 3D hepatocyte architecture with cell polarity, cell junctions, and abundant bile canaliculi possessing microvillus-lined channels. The functionality of these 3D cultures was confirmed by the production of albumin and the exhibition of CYP1A activity over culture time, which were variable, according to Ca-alg bead condition. The expression of specific receptors of hepatitis C virus by Huh-7 cells suggests encouraging data for the further development of a new viral culture system in Ca-alg beads. In summary, this 3D hepatic cell culture represents a promising physiologically relevant system for further in vitro studies and demonstrates that an adequate encapsulation condition can be selected for each target application in liver tissue engineering, specifically in viral studies.
Introduction
Three-dimensional (3D) liver cell culture systems have been introduced, as they provide cells with a biomimetic environment that aims at resembling the in vivo conditions.5–7 Among the different 3D systems, cell microencapsulation is one of the most interesting and effective alternatives so far.8–11 The spherical shape of capsules or beads offers support for high cell density growth and presents a significant specific surface-to-volume ratio, which increases the contact surface toward the medium to ensure both metabolite and nutrient transfers. Among the components proposed for 3D matrices, 12 the carbohydrate polymer sodium-alginate (Na-alg) is the most currently employed biomaterial in cell microencapsulation technology for applications in different areas such as drug delivery and tissue engineering.13,14 It is an appealing biomaterial for the construction of “biohybrid organs” and “micro-bioreactors,” thanks to its biocompatibility, chemical and mechanical stability, and permeability for living cells, and its hydrated 3D network allows cells to interact with each other.9,11,15,16 More interestingly, calcium-alginate (Ca-alg) beads hosting cells can be used for further studies on bioreactors in dynamic conditions.14,17–23 The fluid dynamics of these 3D culture systems promote in vivo-like exchanges due to an efficient bi-directional transfer of gas, nutrients, and different metabolites between the cells and the surrounding fluid in the bioreactor while exerting no mechanical stress on cells that are protected by the Ca-alg hydrogel.
Extracted from the cell walls of brown algae such as calcium, magnesium, and sodium salts of alginic acid, alginates are natural linear polysaccharides with 1.4-linked β-D-mannuronate (M) and α-L-guluronate (G) residues arranged as blocks of similar and alternating units. During the gelation process, consecutive blocks of G-, M-, and MG molecules are electrostatically cross-linked with divalent ions such as Ca2+, Ba2+, and Sr2+. 24 The arrangement of these M and G residues, the overall ratio M/G, and the viscosity of extracted Na-alg, all depending on the algal source and processing treatment of the biopolymer, offer a range of different chemical types of Na-alg. These chemical properties and the polymer concentration govern the mechanical and biological behaviors of Ca-alg hydrogels.25–28 The Na-alg concentration could be adapted to change the internal matrix porosity, and then favor the diffusion rates of nutrients and secreted biomolecules. 11 Miranda et al. have compared the effect of Na-alg concentration on hepatocyte functionality using 0.4% and 0.7% of ultrahigh viscosity Na-alg. 14 Finally, the average molecular weight of Na-alg, and, thus, the viscosity, seems to decisively affect the biocompatibility of Ca-alg beads.29,30
All these results demonstrate that the Ca-alg bead preparation condition as the internal matrix porosity plays an important role in elaborating a 3D culture system. Each Ca-alg preparation needs to be then adapted to the target application. The goal of this study is to identify the optimal microencapsulation conditions through the Na-alg concentration and viscosity selection in order to establish a 3D culture model for hepatocytes that is permissible to the hepatotropic virus. We have focused accordingly on the feasibility of encapsulating and culturing a specific hepatic cell line (human hepatocarcinoma-derived cell Huh-7) already recognized to be permissive in 2D to hepatitis C virus (HCV)31,32 in four kinds of Ca-alg beads (low viscosity [LV], medium viscosity [MV], 1%–2%). We evaluated the effect of Ca-alg bead preparation on cell behavior through cell viability, proliferation, specific-liver function, ultrastructural organization, and the expression of HCV receptors by immunocytochemistry during a prolonged period up to 7 days under dynamic conditions.
Materials and Methods
Cell culture in monolayer
A cellular monolayer of human hepatoma-derived cell Huh-7 was maintained at 37°C under an atmosphere of 95% air and 5% CO2 in Dulbecco's modified essential medium (DMEM) high glucose (Institute of Technology Jacques Boy) supplemented with 10% fetal bovine serum provided from the same institute. The medium was changed thrice per week. The cells were cultured to confluence before encapsulation processing.
Ca-alg bead formation and cell culture in 3D
Ca-alg beads were formed using two types of Na-alg extracted from Macrocystis pyrifera: LV (G/M=33/67, viscosity at 2%=200 cP, chain length=from 12,000 to 80,000 Da) and MV alginate sodium salt (G/M=33/67, viscosity at 2%=2000 cP, chain length=from 80,000 to 120,000 Da) (Sigma Aldrich). To prepare different solutions of Na-alg, LV 2%, LV 1.5%, MV 1.5%, and MV 1%, the Na-alg powder was dissolved in a sterile saline solution (154 mM NaCl solution buffered with 10 mM HEPES, pH 7.4). The mixture was subsequently filtered using a 0.2 μm membrane, leading to sterile Na-alg solution. Before the Ca-alg bead formation, Huh-7 cells were removed after trypsin incubation (Trypsin EDTA 1× in phosphate-buffered saline [PBS]; Institute of Technology Jacques Boy), washed with the culture medium, and suspended at 500,000 cells/mL in Na-alg solution. Na-alg droplets were formed using the air-extrusion method previously described.17,18 Parameters of the encapsulation process, including air and Na-alg flows, distance between the needle and the gelation bath surface were experimentally determined to produce spherical beads with the 500 μm required diameter.
Formed beads were left in a CaCl2 bath (115 mM CaCl2, 154 mM NaCl buffered with 10 mM HEPES) for 15 min at room temperature for gelation to occur. The beads were washed with the cell culture medium. After the encapsulation step, 1 mL of beads was transferred to the tissue culture flask (25 cm2) (Grenier Bio-One) combined with 5 mL of complete DMEM on a 3D moving plate. The medium was changed every second day. The diameter of the Ca-alg beads was measured with the aid of pictures taken using light microscopy (Microscope DMI 6000B; Leica). Thirty beads per Ca-alg bead condition were analyzed.
Ca-alg bead porosity studies
High-resolution cryo-scanning electron microscopy
Transmission images of both Ca-alg beads and encapsulated cells were taken under the light microscope. High-resolution cryo-scanning electron microscopy (cryo-SEM; Hitachi S4500 field emission gun SEM equipped with a dedicated Polaron LT 7480 cryopreparation device) was employed to investigate the 3D structure of the Ca-alg bead network at various Na-alg preparations (LV 2%, LV 1.5% and MV 1.5%, MV 1%). Droplets of the samples, deposited on small tubular blocks, were cryofixed by plunge freezing in nitrogen slush at −210°C, then fractured with a cold metal rod. Samples were then transferred into the SEM chamber, where sublimation of the surface ice formed from the interstitial water of the sample was obtained by progressively increasing the temperature up to ∼−70°C. This operation was visually controlled in order to avoid any distortion of the sample structure. The temperature of the sample was then lowered back to about −110°C, allowing a complete stabilization of the sample. The observations were performed at 1 kV without any sample coating. Cryo-SEM has been shown to be a preserving technique that observes the 3D microstructure of polysaccharide hydrogels.33,34
Diffusion of nanoparticles inside alginate beads (without cells)
Polystyrene nanoparticles of different diameters: 33, 75, 103, 161, and 263 nm (Estapor®; Merk Chimie) were submitted to a diffusion test in the Ca-alg beads. Each type of nanoparticles was incubated with Ca-alg beads at a ratio of 1:2 (bead volume/nanoparticle solution volume) under dynamic condition in a 3D stirrer for 24 h at room temperature. Nanoparticle concentrations remaining in the supernatant were then measured on a spectrophotometer (SPECORD) at 254 nm, after a calibration curve had been set.
Assessment of encapsulated Huh-7 cell viability and proliferation
Huh-7 cell viability within Ca-alg beads was assessed by fluorescence staining with propidium iodide and acridine orange using a fluorescence microscope (DMI 6000B; Leica), as previously described.18,23
Cell viability and proliferation were quantified by a modified Alamar blue assay (Invitrogen). Briefly, at days 1, 3, and 7 of cell culture, 5 mL of 10% (v/v) Alamar blue dye in complete DMEM were added to each tissue culture flask (25 cm2). After 1 h of incubation at 37°C, 5% CO2, 100 μL of supernatant were collected from each flask and transferred to 96-well plates that were measured on a fluorescence spectrophotometer (SPECTRAFLUOR Plus; TECAN) using 560EX nm/590EM nm filter settings.
Alternatively, cell mortality was evaluated using the lactate dehydrogenase (LDH) assay (Promega). This assay monitors the conversion of NADH into NAD+ by LDH. The LDH released into the extracellular medium by damaged cells was expressed as a percentage of the total LDH activity in the cells (% LDH released=extracellular LDH/[extracellular LDH+intracellular LDH]). At days 0, 2, and 6 of incubation, the culture medium was changed using 5 mL of DMEM supplemented with 1% fetal bovine serum. After 24 h, 50 μL of culture media were harvested from the wells containing the encapsulated cells and placed in a 96-well microtiter plate. Fifty microliters of the reaction mixture (provided by the LDH Kit) were added to each well. The plate was then incubated for 30 min at room temperature in the dark. The absorbance of the samples was measured at 490 nm using the spectrophotometer. To calculate the relative extent of cell death in the beads, the cells within beads were lysed in 1% Triton X-100 at days 1, 3, and 7, and the intracellular LDH released was then measured.
For DNA quantification, cells were treated with a fluorescent DNA-binding dye (Bisbenzimidazole derivative Hoechst 33258; Invitrogen). The assay was used to count the cells in Ca-alg beads in the range of ∼10,000–500,000 per well on the basis of DNA content for proliferation studies by a comparison with a standard curve. One milliliter of Ca-alg beads was dissolved by incubating with 2 mL of a solution containing 50 mM sodium citrate (Sigma) and 55 mM EDTA (Sigma) at pH 7 at 37°C for 15 min. The released cells were recovered, rinsed after centrifugation (200 g, 5 min), and suspended in 1 mL of distilled water. One hundred microliters of final cell solution was transferred into microplate wells (96 wells) to perform the assay. Fluorescence was measured using excitation and emission filters that were centered, respectively, at 360 and 460 nm. To quantify the DNA from viable cell populations, the values obtained were calculated by taking into account the viability (%) measured by the LDH test.
In order to monitor the cell expansion within the Ca-alg bead matrix, diameter of cell clusters (spheroid aggregate) and size (length and thickness) of multicellular channel-like structures were quantified and recorded from light microscopy transmission images. The test was performed using 10 clusters per bead and 10 beads per Ca-alg bead condition and time point.
Assessment of albumin production
Samples of the cell culture medium in days 1, 3, and 7 were taken and frozen for a subsequent analysis of albumin concentration. Albumin concentration was determined by a sandwich enzyme-linked immunosorbent assay (ELISA) as previously described. 35 Goat anti-human albumin antibodies were purchased from Cappel Laboratories (Cochrainville). The results were presented as the amount of albumin released within 24 h, normalized with 106 cells. The cell number was obtained using a DNA quantification test.
Cytochrome P4501A activity measurement
The cytochrome P4501A (CYP1A) enzyme activity was determined using EROD assay. 3-methylcholanthrene (3-MC, 5 μM) served as inducer of CYP1A (treatment of 24 h). The EROD activity was determined using 10 μM of ethoxyresorufin in DMEM as the substrate of CYP1A enzyme in the presence of 1.5 mM of salicylamide to inhibit conjugating enzymes. After incubation for 1 h at 37°C, the fluorescence was measured using a fluorescence spectrophotometer with 535EX nm/595EM nm filter settings. The test was performed in triplicate for each point. A resorufin standard curve was used to convert fluorescence to μmol of resorufin formed.
Assessment of membrane markers by immunofluorescence
At day 7 in culture, encapsulated Huh-7 cell aggregates were fixed with 100% cold methanol and rinsed with PBS. Three-dimensional cell aggregates were stained with antibodies that were specific for MRP2 (Sigma), CD26 (Invitrogen), occluding (Invitrogen), claudin-1 (Invitrogen), CD81 (BD Biosciences), and SR-B1(BD Transduction Laboratories™) overnight at 4°C, followed by incubation with a 1:100 dilution of an appropriate Alexa555-conjugated secondary antibody (Invitrogen) or Alexa488-conjugated secondary antibody (Invitrogen) for 1 h at room temperature. Cell nuclei were counterstained by Hoechst dye. Bound antibodies were visualized via confocal microscopy (Zeiss).
Transmitting electron microscopy
Encapsulated 3D Huh-7 cell aggregates from day 7 in culture were fixed, prepared in ultrathin sections, and stained with uranyl acetate (Merck) and lead citrate (Agar). Images were observed using a JEOL JEM-1400 transmission electron microscope operating at 120 kV (JEOL).
Data analysis
Data are expressed as mean±standard error of mean values. Each experiment was repeated independently on four separate occasions. Statistical analysis was performed using the nonparametric Kruskal–Wallis test in order to compare the differences between groups. p<0.05 was considered significant.
Results
Morphological characterization and porosity of Ca-alg beads
Four different Na-alg preparations (LV 2%, LV 1.5%, MV 1.5%, and MV 1%) were used for cell encapsulation. The permeability of this kind of Ca-alg beads was analyzed in a previous paper. 18 Ca-alg bead diameter can be controlled by modifying the parameters of the encapsulation system (airflow, alginate flow, and distance between the tip of needle and the bath gelation surface). Applying the developed fabrication method, spherical Ca-alg beads were formed with diameters of ∼500 μm. The internal structure of the beads was assessed by cryo-SEM, which is specifically useful for unstable wet organic samples.33,34 This technique then allowed observation of the Ca-alg bead samples in their “natural” hydrated state, preserving the porous microstructure of the matrix (Fig. 1a). The pore size appeared to be dependent on the type and the concentration of the Na-alg used in the preparation and varied from around 6–10 μm.

Diffusion studies were conducted for beads LV 2%, LV 1.5%, MV 1.5%, and MV 1% by applying different polystyrene nanoparticle sizes. The bead (Vb) and nanoparticle (Vnp) volumes were identical irrespective of the Ca-alg preparation. Nanoparticle concentration decreases after 24 h of incubation in dynamic condition were due to transfer from the supernatant to Ca-alg beads. Ci, Cf were the initial and final (after 24 h of incubation) concentrations of nanoparticles, respectively.
At equilibrium: Ci*Vnp=Cf*(Vnp+Vb)
Hence, Cf/Ci=Vnp/(Vnp+Vb) with Vnp=2Vb
If Ca-alg matrix was fully permeable to nanoparticles, the theoretical ratio at equilibrium was 2/3=66.67%. The data (Fig. 1b) demonstrated that for all types of Ca-alg bead preparations, the smallest nanoparticle (33 nm) can diffuse within the beads. For LV 1.5%, MV 1.5% Ca-alg beads, most of the bigger-sized polystyrene nanoparticles (75, 103 nm) can penetrate within the beads, while the diffusion of these particles are hindered for other types. Nanoparticules of 161 and 263 nm did not penetrate at all within the Ca-alg beads.
Hepatic cell growth
Cell viability and proliferation
During the encapsulation process, we previously noted a cellular stress generated by air extrusion. 18 To quantify the importance of this mechanical stress on cell viability, the LDH assay was applied to our culture models. 36 At day 1, cell viability was about 50%, without a significant difference between Ca-alg bead conditions (Fig. 2a). A reduction in cell death was quantified over the next few days in culture: At day 3, viability values increased to about 80%, and at day 7, viability reached almost 100% for all types of Ca-alg beads.

Cell viability.
Figure 2b shows microscope pictures of cells in beads after staining with acridine orange and propidium iodide to label viable and dead cells, respectively, after 1 day of incubation at 37°C 5% CO2. The proportion of red cells compared with green ones was similar, irrespective of the Ca-alg bead conditions, and is homogeneously distributed among bead volume.
Cell proliferation within Ca-alg beads was assessed using Alamar blue assay (Fig. 3a). There was a significant increase in fluorescence intensity from day 1 to 7, from about 400 units to almost 17,000 units, indicating cell growth. A slightly higher metabolic activity on day 7 of cells cultured in the Ca-alg bead MV 1.5% condition was observed (p<0.05).
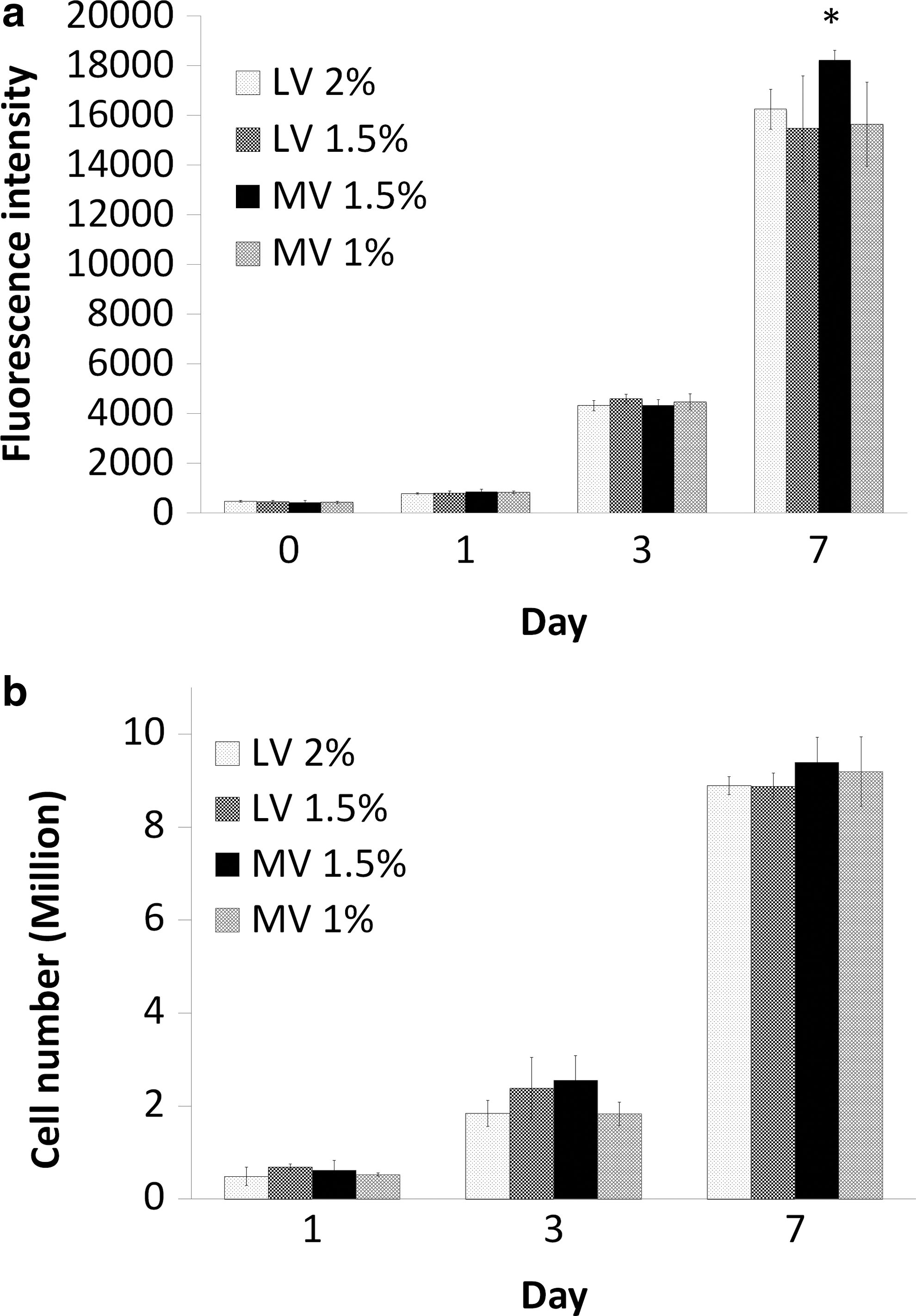
Cell proliferation analysis.
To confirm the cell ability to grow in our culture model, independently of their metabolic activities, cell proliferation was quantified using DNA content. Due to the fact that the quantification of DNA could not distinguish between the viable and dead cells, DNA results have been recalculated to take in account the viable cell population alone, using the viability percentage obtained by the LDH assay. Our data show that Huh-7 cells proliferated within all Ca-alg bead types, increasing the initial cell density by a factor of 20 over 7 days (Fig. 3b), independently of the Ca-alg bead type.
Cellular aggregate organization
Proliferation was associated with Huh-7 cell organization in multicellular structures. The number of aggregate spheroids (cluster) and cluster size increased over time (Fig. 4a). In Ca-alg bead of MV, Huh-7 cells formed multicellular channel-like structures and round spheroids; whereas in Ca-alg bead of LV, cells were organized in round spheroids only. In order to monitor the cell expansion within Ca-alg bead matrix, diameters of cell spheroid clusters were measured (Fig. 4b). Multicellular channel-like structures increased and reached a mean of ∼120 μm length and 40–50 μm thickness.
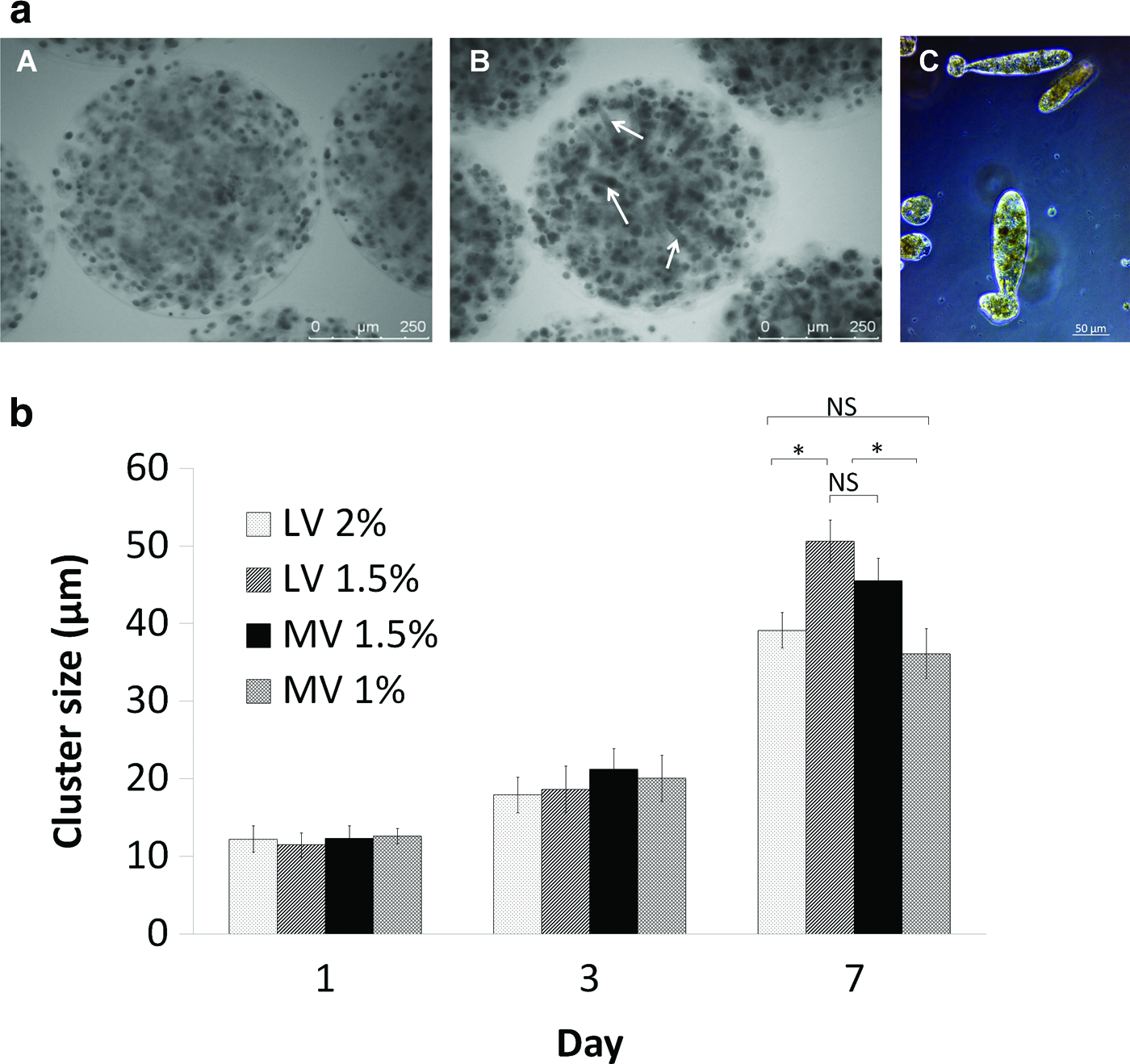
Huh-7 cell multicellular structures.
The structures and ultrastructures of cell aggregates formed in two types of Na-alg viscosity were characterized using transmitting electron microscopy (TEM). Huh-7 cell multicellular ultrastructures within Ca-alg beads of MV 1.5% and LV 1.5% conditions on day 7 of culture showed cell 3D arrangement with a tight junction between adjacent cells (Fig. 5). The distinct nuclei, abundant mitochondria, lipid droplets, and endoplasmic reticulum indicated that Huh-7 cell aggregates within Ca-alg bead were metabolically active. Furthermore, the cells displayed polarity as judged by the presence of bile canaliculi possessing microvillus-lined channels, bordered by tight junctions. These ultrastructural characteristics were similar in two types of cell aggregates, spheroid or channel-like structures.
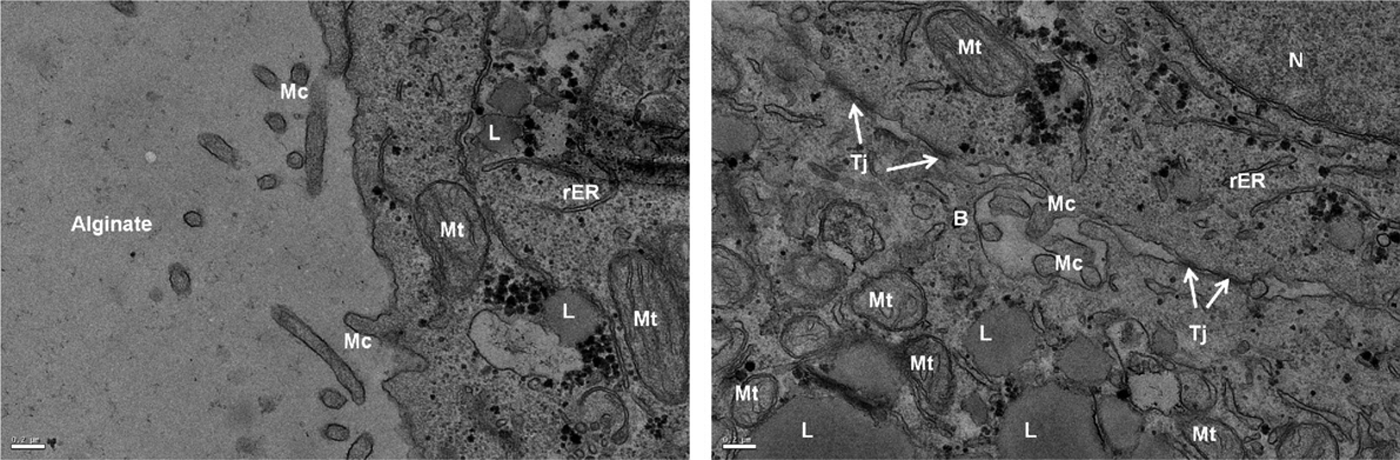
Huh-7 cell multicellular ultrastructures of Ca-alg beads MV 1.5% conditions by transmitting electron microscopy at day 7 of culture. Nuclei (N), tight junctions (Tj) mitochondria (Mt), lipid droplets (L), endoplasmic reticulum (rER), microvillus-lined channels (Mc), and bile canaliculi (B). Scale bars 200 nm.
Impact of Ca-alg bead preparation on Huh-7 cell functionality
Albumin production
Albumin secretion in the supernatant was determined by ELISA assay as an indicator of hepatocyte functionality of Huh-7 cells in 3D culture. As shown in Figure 6a no significant difference of albumin production between the fourth types of Ca-alg beads was observed at day 1. The average secretion rates varied from 2±0.76 μg/24h/million to 2.82±0.56 μg/24h/million of cells. If prolonged cultivation of Huh-7 cells within the MV 1.5% Ca-alg bead maintained albumin production at about 2 μg/24h/million of cells, a significant decrease was observed in the Ca-alg bead of LV conditions, with a more pronounced influence of LV 2% Ca-alg beads (p<0.05). At day 7, a significantly higher production of albumin was achieved in MV 1.5% Ca-alg beads compared with other conditions (p<0.05).

Impact of Ca-alg composition on Huh-7 cell functionality.
Cytochrome P450A1 activity
The activity of major drug-metabolizing enzyme (CYP1A) was analyzed during 7 days of cell culture. The level of CYP1A activity on Huh-7 cells was assessed in untreated (control condition) or treated cells with 5 μM of 3-MC, a marginal inducer of CYP1A, using the EROD assay. As shown in Figure 6b, our data revealed that Huh-7 cells encapsulated in Ca-alg beads exhibited CYP1A activity and at day 7 of culture, a significantly higher CYP1A activity in the MV 1.5% condition was observed (p<0.05).
Immunolocalization of HCV receptors
Since these factors are essential for HCV entry in hepatocytes, expression and localization of cellular tight junction proteins (claudin-1 and occludin) and polarity markers (CD26, MRP-2) were assessed in 3D Huh-7 cell aggregates at day 7 using immunofluorescence analysis (Fig. 7). Three-dimensional Huh-7 cell channel-like structures in Ca-alg bead of MV 1.5% condition exhibited a higher expression of cellular tight junction proteins and MRP-2 marker in comparison with 3D spheroid structures in Ca-alg bead of LV 1.5% condition, especially on the basolateral cell membranes. A similar expression of CD26 marker was observed in both conditions. By comparison, no expression was observed for MRP-2 and CD26 markers in 2D conventional culture on polystyrene substrate (data not shown).
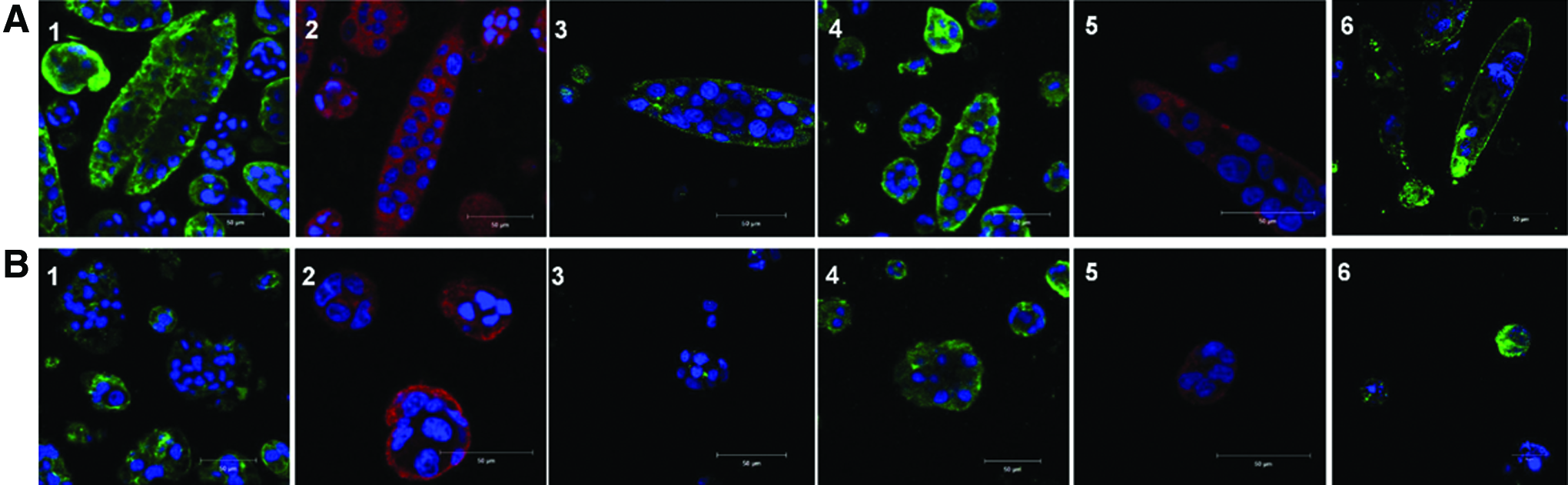
Immunolocalization of membrane markers. Organization and localization of cell polarity and tight junction protein in three-dimensional Huh-7 cell aggregates in
CD81 and scavenger receptor B1 (SR-B1), which are specific cellular receptors for HCV, were assessed using indirect immunofluorescence after 7 days of culture in Ca-alg beads (Fig. 7). Huh-7 cells expressed SR-B1 receptor and CD81 with a high level in channel-like structures in MV 1.5% Ca-alg beads and in spheroid ones in LV 1.5% Ca-alg beads.
Discussion
Liver cell microencapsulation in Ca-alg beads has been applied in tissue engineering for adult or embryonic hepatocyte transplantation and in biomedical applications such as extracorporeal bio-artificial liver.14,37–39 Since each of these purposes required appropriate specifications to respond to pathophysiological conditions, culture models of encapsulated hepatocytes have been adapted through various experimental approaches. Our objective here was to establish a hepatocyte culture system that is suitable for future viral studies. Therefore, we compared the influence of Na-alg viscosities (LV vs. MV) and percentages (1%, 1.5%, and 2% [w/v]) on hepatocyte behavior and viral receptor expression in a 3D encapsulated hepatocyte model. Na-alg concentration range was chosen to respond to three constraints: (1) to produce beads with optimal sphericity and stable mechanical properties; (2) to obtain a Ca-alg bead structure that is compatible with cell survival and proliferation; and (3) to allow the diffusion of HCV particles between the cells and the external bead microenvironment. Ca-alg bead of LV 1% condition was not retained, because it generated irregular bead shapes with fragments of Ca-alg gel. 18 This was supported by Wang et al., who noted that a low percentage of Na-alg was associated with mechanical instability which was incompatible with long-time culture, and generated matrix fractures, resulting in the release of encapsulated cells. Alternatively, the stringent properties of MV 2% Ca-alg beads did not support cell proliferation. 11
The extrusion with the coaxial air flow method was reproducible and easy to implement; it also allowed excellent control of the bead diameter but compromised cell viability. 18 In fact, the recovery of dead cells from the beads 24 h after encapsulation was ∼50% according to LDH assay and was homogeneously distributed all over the bead volume, as shown by the acridine orange/propidium iodide staining. In comparison with other methods of Ca-alg encapsulation, higher viability has been reported,11,15,40 whereby immediately after encapsulation, about 75%–90% of live cells have been detected. However, the cell viability 7 days after the encapsulation in their cultures decreased. On the contrary, the assessment of cell viability in the present study indicated about 100% of the live cells in day 7, indicating that the four Ca-alg bead preparations cultivated in a dynamic system avoided cell death in the bead center and provided a compatible environment for hepatic cell culture.
Our data showed an excellent proliferation in 3D culture with an exponential profile up to 7 days. Similar results have been reported for the 3D culture of other hepatocarcinoma cell lines in Ca-alg beads.9,21,40,41 Nevertheless, HHY41, HepG2 human cells grew only after a delay of few days after the encapsulation, compared with Huh-7 cells, which seemed to quickly adapt in Ca-alg bead matrix. The Na-alg condition viscosity/concentration MV 1.5% gave better microenvironmental properties than other Ca-alg bead conditions, as demonstrated here by Alamar blue assay (p<0.05). The Huh-7 cell proliferation was associated with spatial organization in compacted multicellular structures, characterized by an influence of Ca-alg bead condition on the aggregate shape. The MV allowed the development of multicellular channel-like structures, whereas only spheroids were observed in LV Na-alg, in the concentration conditions used in our model. The channel-like structures were described by Shirai et al., who investigated the conditions of channel formation, such as Ca-alg bead preparation and gelation salt impact. 42 Their data suggested that the channel-like structures originated in a vacant space formed during the gelation stage of the encapsulation process, although their development was linked to the cellular presence in the Na-alg solution. 35 In the present study, the cell aggregate shape was controlled via the selection of Ca-alg bead preparation and culture delay to optimize the microenvironment of hepatocytes.
Hepatocytes form in vivo a polarized epithelium where their basal membranes are oriented toward the sinusoidal endothelial cells and their apical membranes are lined by the biliary cells. This polarized tissue structure is essential to maintain the liver-specific functionality but disappears in conventional 2D cell culture on plastic substrate. Here, Huh-7 cells in 3D culture formed cohesive configurations, in the core of which canaliculi-like structures were observed by TEM. The bile canaliculi were characterized by projections of a few microvillosities in the lumen and by tight junctions lining the lateral surface. This ultrastructural cell organization was found in all aggregates examined, irrespective of their shape, spheroid or tubular, from Ca-alg beads of MV 1.5% and LV 1.5% conditions. This complex organization was previously described in Ca-alg scaffolds, where primary or fetal hepatocytes were cultivated,13,43 and seemed to occur early after the establishment of perfusion in a bioreactor. 19 The recovery of cell polarity was confirmed by immunostaining the proteins MRP2 and CD26 which are reliable markers for the canalicular membrane. 44
We confirmed the functionality of the encapsulated cells by testing the production of albumin and CYP1A activity. The stability of albumin secretion in MV Ca-alg beads over the 7 day culture, while decreasing in LV condition, might result from an improvement in hepatic function. The slight difference in Ca-alg bead porosity could influence the diffusion of secreted albumin out of the beads to the supernatant. The presence of multicellular channel-like structures in MV Ca-alg beads might also be linked to a greater albumin secretion at day 7. This pattern of albumin secretion correlated well with previously published studies of 3D culture of hepatic cells. As shown by Khalil et al., hepatic HepG2 cell cultivation as cohesive spheroids up-regulated their synthetic and detoxification functions, 9 with a noticeable influence of spheroid size on albumin production. 45 CYP1A is one of the major drug metabolizing CYP enzymes.46,47 Here, using our 3D culture model, Huh-7 cells exhibited significantly greater activity in MV 1.5% over 7 days of culture for albumin secretion and CYP1A activity compared with other Ca-alg bead preparations. Hepatocellular detoxification functions were maintained in 3D culture, in agreement with other studies.12,48 We hypothesize that higher activity in 3D culture was due to significantly tight cell-to-cell interactions. In fact, the culture of isolated hepatocytes was followed by a rapid loss of CYP P450 expression after a short time in 2D culture.49,50
A relevant model culture for the study of the entire HCV life cycle should allow entry, replication, and production of infectious particles. 51 In this context, Ca-alg bead matrices should support the viral exchanges between hepatocytes and culture fluid. The Ca-alg bead porosity of the both types of beads that were more specifically analyzed (MV 1.5% and LV 1.5% conditions) was largely superior to the HCV particle size, which is 50–60 nm in diameter.31,52
HCV infection depends on the expression and organization of numerous cellular receptors that coordinate the viral entry in liver cells. These receptors belong to a multistep mechanism where CD81, SR-B1, and Niemann Pick C1-like 1 cholesterol absorption receptors act as specific receptors for HCV particules, and claudin-1 and occludin act as coreceptors that are required for a late step in viral entry.53–56 The expression of these factors in Huh-7 cells organized in channel-like aggregates suggests that MV 1.5% Ca-alg bead hosting Huh-7 cells is a new and promising model culture that could be submitted to HCV replication and viral production experiments.
Conclusion
In summary, we succeeded in reconstituting a hepatic-like structure formation from Huh-7 cells using a Ca-alg encapsulation model. The 3D culture of beads provided multicellular structures in vitro, presenting some of the major liver functions, such as albumin production and CYP1A activity, depending on the preparation of Ca-alg beads. By selecting the appropriate percentage and viscosity of Na-alg, these 3D hepatic cells represent a promising physiologically relevant system for further in vitro studies for different applications in tissue engineering, especially for applications in viral studies.
Footnotes
Acknowledgments
This work was supported financially by the FEDER Program and the Picardie Region, France. The authors thank B. Satiat-Jeunemaitre and D. Jaillard, Science Institute of Plants, CNRS, Gif sur Yvette, France, for scientific and technical assistance, respectively, on ultrastructural preparations and analysis by TEM, and A. Richard (Electronic Microscopy Center, University Orléans, France) for help in cryo-SEM observations. The authors are grateful to M. Jaffrin (UMR CNRS 7338, University of Technology of Compiègne) for corrections made in the article.
Disclosure Statement
The authors state that no competing financial interests exist.