
Abstract
The fabrication of biomimetic skeletal myocyte constructs continues to present a challenge to functional tissue engineering. The skeletal myogenesis of bone marrow-derived mesenchymal stromal cells (BMSCs) to mimic the native tissue architecture offers great therapeutic promise, but remains particularly difficult. The aim of this study was to examine the possibility of accelerating the skeletal myogenic differentiation of BMSCs with an aligned structure by applying cyclic strain. Mouse BMSCs (mBMSCs) were plated on silicone sheets that were coated with fibronectin and subjected to cyclic 10% uniaxial strain when they reached 80%–90% cell confluency. Cells cultured in a growth medium that were subjected to cyclic strain at a frequency of 0.17 Hz (10 times/min) demonstrated a shift of alignment within 48 h from a completely random orientation to a well-aligned morphology with well-organized actin stress fibers that were parallel to the strain vector. The cyclic strain restricted the motility and proliferation of the aligned mBMSCs in the growth medium, which resulted in tight cellular contact in the cell population. When mBMSCs were subjected to cyclic strain in a myogenic medium, reverse transcription–polymerase chain reaction analysis demonstrated the upregulation of skeletal myogenic marker genes (myogenic factor 5 [Myf5], myogenin, and myogenic regulatory factor 4 [MRF4]), but not smooth muscle marker genes (myocardin and α-smooth muscle actin). In addition, immunocytochemistry showed that the mBMSCs fused to form multinucleated myosin- and myogenin-positive myotubes in the direction of the applied tension within 5 days. These results demonstrate that our simple method of applying of cyclic strain to cells cultured in a myogenic medium greatly accelerates the skeletal myogenic differentiation of mBMSCs with an aligned structure, and they highlight the importance of cellular alignment for creating physiologically relevant environments to study the myogenesis of BMSCs and engineer skeletal muscle.
Introduction
Alternatively, engineered muscle constructs may overcome the problems associated with the autologous transfer of muscle tissue, and provide a valid alternative for tissue replacement in the enhancement of muscle regeneration. Bone marrow-derived mesenchymal stromal cells (BMSCs) are multipotent cells capable of giving rise to cells of mesodermal origin, including bone, cartilage, fat, and muscle, 5 and of nonmesodermal origin, including neurons. 6 BMSCs are easy to isolate and expand rapidly from patients, and are not associated with any major ethical or technical problems. 7 These characteristics of BMSCs make these cells promising candidates for cell-based tissue engineering for the repair of lost or damaged tissues, including skeletal muscle. 8
The practical application of BMSCs to skeletal muscle engineering depends on the ability to efficiently control their differentiation into functional skeletal muscle cells in vitro. In this regard, BMSCs can be induced to express several myocyte markers when treated with 5-azacytidine.9,10 It has also been shown that Notch1 intracellular domain gene transfer leads to efficient myogenic differentiation of BMSCs. 11 However, it is desirable to induce the differentiation of BMSCs without using DNA demethylating drugs or gene delivery to avoid any unexpected affects after transplantation of these cells. On the other hand, Muguruma et al. 12 demonstrated the induction of myocyte formation from BMSCs under physiological conditions only using certain trophic bioactive factors; however, this method still has limitations related to the low efficiency of myocyte induction. Therefore, the potential use of BMSCs for skeletal muscle engineering awaits the development of an efficient protocol that guides BMSCs to a prescribed skeletal myogenic differentiation.
To generate well-differentiated skeletal muscle cells in vitro, cell manipulation that better resembles the physiological conditions would be a promising approach. In terms of the physiological architecture, the prototypical skeletal muscle is composed of striated myotubes that are arrayed in parallel with one another along a common axis. Previous studies have demonstrated that culturing myoblasts on topographically patterned surfaces results in the formation of myotubes, organized in parallel much like the arrangement in muscle tissue.13–15 In addition, it has been shown that uniaxial cyclic strain caused the reorientation of C2C12 myoblasts, leading them to align in a unidirectional manner.16,17
In terms of the physiological environment, skeletal muscle is constantly exposed to biomechanical forces, and changes in phenotype result from differential gene expression in response to mechanical signals.18,19 Resident myogenic stem cells (quiescent satellite cells) are equipped with an inherent capacity to generate, perceive, and respond to biomechanical stimuli, 20 and many investigations have provided evidence that biomechanical forces and signals regulate the development of skeletal muscle.21–23 However, the effect of cyclic strain on the skeletal myogenesis of BMSCs has not yet been evaluated.
The complete control of BMSC differentiation before transplantation for skeletal muscle regeneration therapy is critical to prevent unexpected clinical results, which prompted us to utilize a rodent cell culture model to explore the possibility that appropriate cyclic strain could be used for the in vitro generation of aligned skeletal myocytes from BMSCs. Hence, the aim of this study was to investigate whether the application of cyclic strain could induce efficient skeletal myogenic differentiation of mouse myoblasts and BMSCs with regard to cell alignment.
Materials and Methods
Cell culture
Mouse BMSCs (mBMSCs) were obtained using a protocol approved by the Animal Research Subjects Committee of Osaka University Graduate School of Dentistry (approval number: 19-054-1). Bone marrow cells were collected from the femurs of male 10-week-old C57BL/6 mice, as described previously, 6 and were maintained in the BMSC growth medium consisting of the Dulbecco's modified Eagle's medium (DMEM; Nacalai Tesque, Kyoto, Japan) supplemented with 15% fetal bovine serum (FBS; JRH Biosciences, Lenexa, KS), 100 units/mL penicillin, 60 μg/mL kanamycin, and 250 ng/mL amphotericin B (Nacalai Tesque) at 37°C in an atmosphere of 5% CO2. The nonadherent cell population was removed after 48 h, and the adherent BMSC layer was washed twice with a fresh medium. Cells were then continuously cultured for 1 week.
Clonal cultures of mBMSCs were established because primary mBMSC populations exhibit heterogeneity and contamination by hematopoietic cells.6,24 The whole population of the primary mBMSCs was collected by trypsinization, and was repeatedly passaged over a 6-month culture period. After 30 passages, the cells were spontaneously induced to become immortal, 25 and single-cell clones were then isolated by a limiting dilution method after single-colony selection using a cloning cylinder. Amplified populations of the clonal mBMSCs were subjected to osteogenic, adipogenic, and chondrogenic induction to determine their multipotency.
The mouse C2C12 myoblast cell line was provided by the RIKEN Bioresource Center (Tsukuba, Japan) and maintained in the C2C12 growth medium consisting of DMEM supplemented with 10% FBS, 100 units/mL penicillin, and 100 μg/mL streptomycin (Nacalai Tesque) at 37°C in an atmosphere of 5% CO2.
Multilineage differentiation of clonal mBMSCs
For the osteogenic induction of clonal BMSC cultures, cells that reached 80% confluence were maintained in the BMSC growth medium supplemented with 0.1 μM dexamethasone, 10 mM β-glycerophosphate, and 50 μM ascorbate-2-phosphate (Sigma, St. Louis, MO). 6 The medium was changed every 3 days. The osteoblastic phenotype was assessed by a standard alkaline phosphatase (ALP) assay and von Kossa staining, 6 as well as by reverse transcription–polymerase chain reaction (RT-PCR) analysis for detecting the mRNA expression of osteocalcin and osteopontin. The chondrogenic induction of clonal BMSC cultures was performed by fabricating a three-dimensional (3D) cell construct followed by culture under hypoxic conditions. 26
The adipogenic induction of clonal BMSC cultures was performed when cells reached 80% confluence, and the cells were maintained in the BMSC growth medium supplemented with 10 mM indomethacin, 50 mM 3-isobutyl-1-methylxanthine, 25 mM insulin, and 10−4 M dexamethasone (all from Sigma) with medium exchanges every 3 days. To demonstrate adipogenesis, Oil red O staining was used as previously described. 27 The expression of an adipogenic marker gene, preroxisome proliferator-activated receptor γ (PPARγ), was determined by RT-PCR.
Myogenic medium
To stimulate skeletal myogenesis, mBMSCs were maintained in the BMSC myogenic medium, 12 which consists of DMEM supplemented with 2% horse serum (JRH Biosciences), 2% MCDB-201 medium (Sigma), 1% ITS+ premix (BD Biosciences), 10−6 M dexamethasone (Sigma), 10−2 M ascorbic acid (Sigma), 10 ng/mL human platelet-derived growth factor-AA (Peprotech, London, United Kingdom), 10 ng/mL human insulin-like growth factor-1 (Peprotech), 10 ng/mL human basic fibroblast growth factor (Peprotech), 100 units/mL penicillin, 100 μg/mL streptomycin, and 250 ng/mL amphotericin B (Nacalai Tesque). One-third of the volume of the medium was changed every 2 days. To evaluate the frequency of differentiated cells among the total cell population, the number of myotube-like multinucleated cells in the phase-contrast images was counted.
The myogenic induction of the C2C12 cells was performed using the C2C12 myogenic medium, which consists of DMEM supplemented with 2% horse serum (JRH Biosciences), 100 units/mL penicillin, 100 μg/mL streptomycin, and 250 ng/mL amphotericin B (Nacalai Tesque). The whole medium was changed every 2 days.
Cyclic strain application
A custom-made mechanical strain device 28 was used to apply cyclic strain to the cells (Fig. 1A). In this device, two clamps held the polydimethylsiloxane (PDMS) silicone substrate (Specialty Manufacturing, Saginaw, MI); one clamp was fixed, and a motor controlled the movement of the other to subject the silicone substrate to uniaxial cyclic strain. Cells were seeded at a density of 3×103 cells/cm2 on silicone sheets, which were coated with 2 μg/mL human fibronectin (BD Biosciences, San Jose, CA) at room temperature overnight after UV irradiation for 3 h. Cells were cultured for 24 h to allow them to adhere before applying the cyclic strain. The silicone substrate was incubated in 35 mL of medium for the duration of the experiment. The entire device was maintained in an incubator at 37°C in an atmosphere of 5% CO2 during the course of the experiments, and cells were subjected to cyclic strain at a frequency of 0.08–0.50 Hz and an amplitude of 10%.
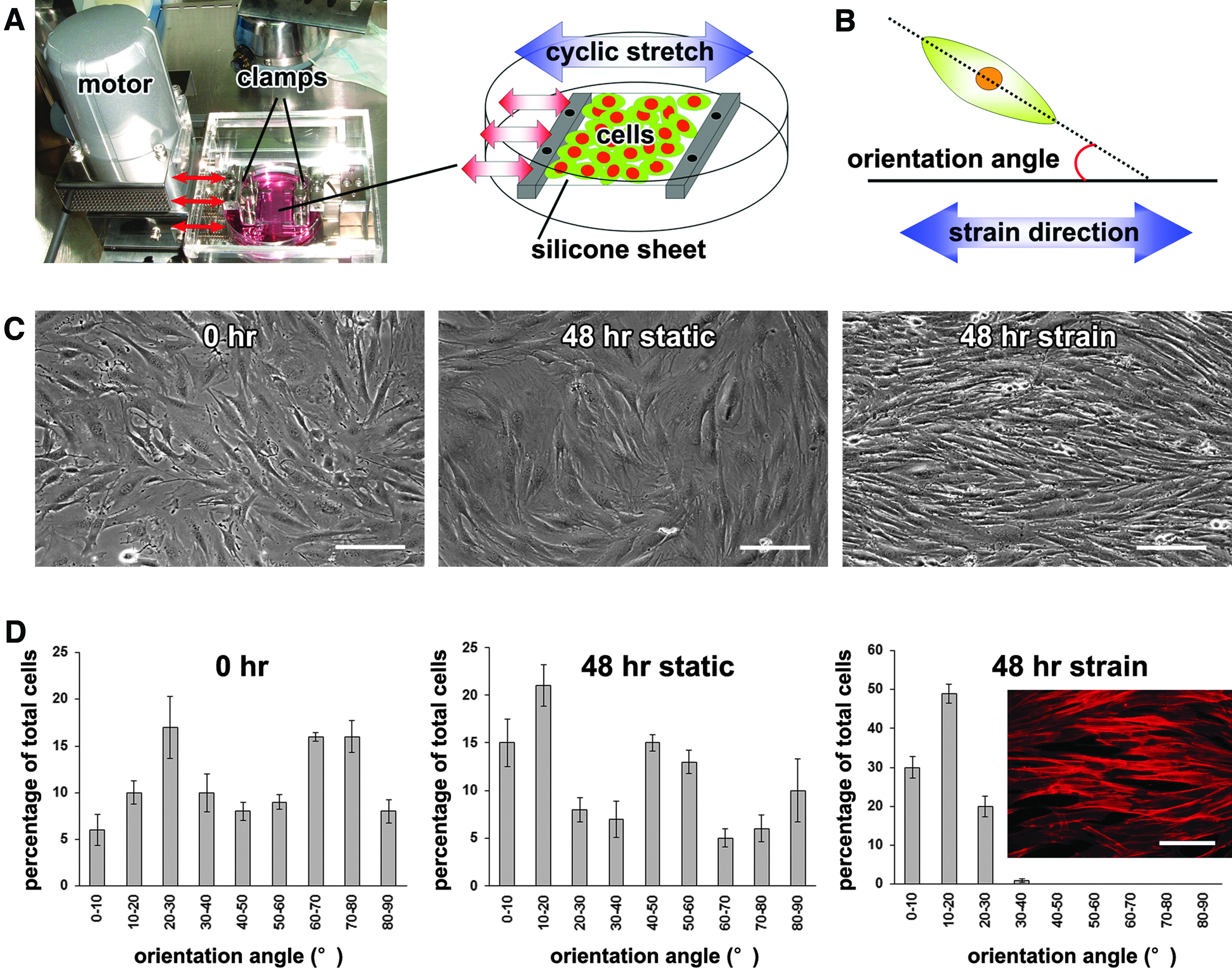
The effects of cyclic strain on mouse bone marrow-derived mesenchymal stromal cell (mBMSC) alignment.
Cellular alignment
Cells were cultured on the silicone sheets in the growth medium under static or cyclic strain conditions, and were monitored for 48 h using an inverted microscope system with a digital camera (Eclipse TE2000-U; Nikon, Tokyo, Japan). Phase-contrast images of each cell culture were taken with the visual field set in the center of each silicone sheet, and a total of 100 cells within the triplicate sample images for each condition were randomly chosen. The cellular orientation angles (the long axis angle of selected cells relative to the direction of strain application; Fig. 1B) were analyzed using the Photoshop 7.0 (Adobe, San Jose, CA) and ImageJ (National Institutes of Health, Bethesda, MD) software.
Cellular motility and proliferation
The mBMSCs were seeded at a density of 3×103 cells/cm2 on a silicone sheet in a 5 mm in diameter circular adhesive region created by placing a noncoated silicone sheet containing a 5-mm hole onto the bottom sheet before cell seeding, and then seeding cells into the resultant well formed by the top sheet. 28 After a 12-h incubation to allow the cells to attach on the sheet, the upper silicone sheet was removed to allow the cells to migrate. The cells seeded on the silicone sheet in the circle were then subjected to cyclic strain in the growth medium. Under static or cyclic strain conditions, phase-contrast images were taken at 24, 48, and 72 h of culture, and the cell motility was determined by measurement of the expanded cellular area using the Photoshop 7.0 and ImageJ software programs.
For the analysis of cell proliferation, mBMSCs were seeded at a density of 1×103 cells/cm2 on a fibronectin-coated silicone sheet, and were cultured under static or cyclic strain conditions for 72 h. All cells on the silicone sheets at 0, 24, 48, and 72 h were collected, and the amount of cellular DNA from each sample was measured using a Fluorescent DNA Quantitation Kit (Bio-Rad, Hercules, CA) with a fluorescent microplate reader (Twinkle LB907; Berthold Technologies, Bad Wildbad, Germany).
RT-PCR analysis
Total RNA was isolated using an RNeasy Mini Kit (QIAGEN, Hilden, Germany). After DNase I treatment (Ambion, Austin, TX), cDNA was synthesized from 1 μg of total RNA using Super Script III reverse transcriptase (Invitogen, Carlsbad, CA). The cDNA target was amplified by PCR using Taq DNA polymerase (Promega, Madison, WI) following the manufacturer's recommendations. The PCR conditions and primer pairs used are given in Table 1. PCR products were subjected to 1.5% agarose gel electrophoresis with ethidium bromide staining, and were visualized under ultraviolet light illumination. The expression of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA was used as an internal control.
Fw, forward; Rv, reverse; Ann Temp, annealing temperature; HS, hot start.
A quantitative TaqMan® real-time PCR analysis for the expression of MyoD, Myf5, myogenin, and MRF4 was performed using an ABI 7300 real-time PCR system (Applied Biosystems, Foster City, CA). The expression of GAPDH mRNA was used as an internal control. The TaqMan® primer and probe sets used were Mm00440387_m1 (MyoD), Mm00435125_m1 (Myf5), Mm00446194_m1 (myogenin), Mm00435126_m1 (MRF4), and 4352339E (GAPDH).
Fluorescent staining
To determine the actin fiber patterns, mBMSCs incubated under static or cyclic strain conditions in the growth medium for 48 h were fixed in 10% buffered formalin phosphate (Wako, Osaka, Japan). Cells were incubated in 1% bovine serum albumin (BSA; Sigma) and 0.1% triton-X100 (Sigma) in phosphate-buffered saline (PBS) for 20 min. After washing, the cells were incubated in 2 units/mL rhodamine-phalloidin (Molecular Probes, Eugene, OR) in PBS for 30 min.
For the immunocytochemical analysis, the mBMSCs or C2C12 cells were cultured on a silicone sheet for 4 to 5 days under static or cyclic strain conditions in the myogenic medium. Cells were then fixed in 10% buffered formalin phosphate and incubated in 1% BSA and 0.1% Triton-X100 in PBS for 20 min. After washes in PBS, the cells were incubated with a mouse anti-rabbit skeletal myosin monoclonal antibody (1:1000; clone MY-32, Sigma), a rabbit anti-rat myogenin polyclonal antibody (1:200; Santa Cruz Biotechnology, Santa Cruz, CA), or control IgG (1:500; mouse or rabbit IgG whole molecules: Santa Cruz Biotechnology) overnight at 4°C. The cells were then washed and incubated for 30 min at 37°C with Alexa 488 conjugated to goat anti-mouse or anti-rabbit IgG (1:500; Molecular Probes), followed by 4 μM ethidium homodimer-1 (Molecular Probes) nuclear staining.
Statistical analyses
A one-way analysis of variance with the Tukey or Dunnett post hoc tests was used for comparisons of more than two groups. The Student's t-test was used for comparisons of two groups. A significant difference was defined when p<0.05.
Results
Multilineage differentiation potential of clonal mBMSCs
When the primary mBMSCs were cultured over 6 months (30 passages), the cells were spontaneously induced to become immortal, showing a dominant spindle-shaped cell population. Clonal mBMSC lines (mBMSC-2, −4, and −7) were established from the immortalized mBMSCs. When these clonal cell cultures were maintained in the osteogenic medium for 21 days, they showed strong staining for ALP and calcium deposition (Supplementary Fig. S1A; Supplementary Data are available online at www.liebertpub.com/tea). On the other hand, when these clonal cells were maintained in the adipogenic medium for 14 days, they were strongly stained red by Oil Red O. The RT-PCR analysis showed strong expression of osteopontin and osteocalcin, or PPARγ, genes in mBMSC-2 when the cells were maintained in the osteogenic or adipogenic induction medium, respectively (Supplementary Fig. S1B). In addition, we previously demonstrated that a large 3D construct comprising clonal mBMSCs exhibited region-specific chondrogenic differentiation and mineralization in vitro, representing the initial stage of endochondral ossification. 26
These results confirmed that the established clonal mBMSCs have the ability to differentiate specifically into osteoblast, adipocyte, and chondrocyte lineages, demonstrating their multipotency as mBMSCs. We used mainly mBMSC-2 for the cyclic strain application in this study.
Effects of cyclic strain on mBMSC alignment
For the cyclic strain application, we first examined the effects of different frequencies of the cyclic application (0.08, 0.17, 0.33, and 0.50 Hz) on mBMSC attachment to the silicone sheet. When a frequency of 0.50 or 0.33 Hz was applied to mBMSCs in the growth medium, the cells were detached from the silicone sheet at 6 h after the cyclic strain application. In contrast, a frequency of 0.08 Hz cyclic strain did not affect the attachment of the cells, but also did not significantly alter the cellular orientation within 48 h. On the other hand, cyclic strain at a frequency of 0.17 Hz (10 times/min) resulted in a cellular alignment shift from a completely random orientation to a well-aligned morphology parallel to the strain vector within 48 h (Fig. 1C). The histomorphometric analysis of the cultures revealed that the average orientation angle was around 10–20° for cells under cyclic strain, whereas cells in the static controls had no prevailing orientation (Fig. 1D). Fluorescence observation of phalloidin-stained mBMSCs showed parallel F-actin distribution and elongated cellular form under the cyclic strain condition for 48 h (Fig. 1D). Other mBMSC lines (mBMSC-4 and −7) also aligned predominantly parallel to the strain direction.
Cell type-dependent alignment under the cyclic strain
We next examined the effects of cyclic strain on the alignment of several other cell types with fibroblastic morphology that were cultured in the growth medium, such as rat BMSCs (rBMSCs), 29 human gingival fibroblasts, 30 and mouse C2C12 myoblasts. Under the cyclic strain condition at a frequency of 0.17 Hz, these types of cells aligned predominantly perpendicular to the strain direction within 48 h (Supplementary Fig. S2A–C), which is vertically different from the alignment direction of mBMSCs (Supplementary Fig. S2D).
Sequential observation of cell alignment under the cyclic strain
Sequential observation by phase-contrast microscopy of the morphological changes of rBMSCs in the growth medium (Supplementary Fig. S2E–H) revealed that cells that were randomly oriented before strain application became rounded and smaller because of cell retraction within 1 h of the application of cyclic strain. The rounded cells then gradually changed their morphology to a spindle shape vertical to the strain direction within 6 h. After that, the cells continued gradually elongating their morphology vertical to the strain direction, and the percentage of aligned cells gradually increased during cyclic strain application. The cells completed their alignment within 24 h. Fluorescence observation of phalloidin-stained rBMSCs showed cell elongation and well-organized actin stress fiber alignment vertical to the strain direction in cells subjected to the cyclic strain condition for 24 h (Supplementary Fig. S2I). In contrast, cells did not elongate or orient in any specific direction under the static culture condition over experimental course of 24 h (Supplementary Fig. S2J).
Effects of cyclic strain on mBMSC motility and proliferation
Under the static condition in the growth medium, the mBMSCs migrated toward the outside from the circular region they were originally located in, and as a result, the area covered by the cells gradually increased over the course of the culture period (Fig. 2A, B). In contrast, cells exposed to the cyclic strain exhibited inhibited motility, so that the cell location under cyclic strain did not change significantly. Magnified observation of the center area of the cell population under cyclic strain showed aligned and dense cell aggregates, in which cells seemed to tightly contact each other within 72 h (Fig. 2C, D).
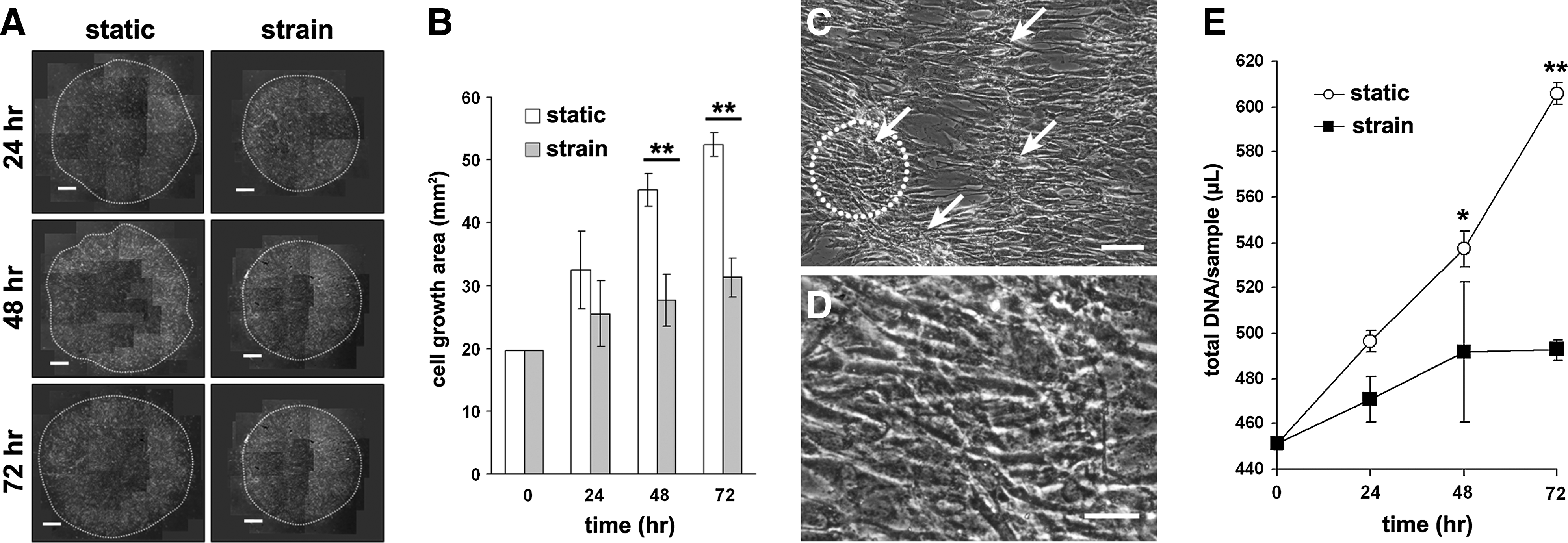
The effects of cyclic strain on cellular motility and proliferation. Cyclic strain was applied to the cell population of mBMSCs, originally localized to a 5 mm in diameter circular region of the substrate.
The cell proliferation assay showed that the proliferation rate of mBMSCs under the static condition was significantly higher than that under the cyclic strain condition over the course of the experiment (Fig. 2E).
Effects of cyclic strain on the differentiation of C2C12 myoblasts
We first investigated whether the cyclic strain condition can be used for myogenic induction using C2C12 myoblasts. When cells were cultured in the C2C12 myogenic medium under the cyclic strain condition, the cells aligned in the vertical direction of the strain vector within 48 h. After 4 days of cyclic strain, the cells showed myotube morphology, and remained in the vertical alignment. These aligned multinucleated cells were positively stained for myosin (Fig. 3A). In contrast, cells cultured under the static condition did not show a unidirectional alignment (Fig. 3B) The number of myosin-positive myotubes from cultures cultivated under the cyclic strain condition was significantly higher compared with cells cultured under the static condition (Fig. 3C).

The effects of cyclic strain on the differentiation of C2C12 myoblasts. Fluorescence images overlaid with the phase-contrast images of C2C12 cells cultured in myogenic medium under cyclic strain
The real-time RT-PCR analysis demonstrated that the expression levels of myogenic marker genes, MyoD, Myf5, and MRF4, in the cyclic strained cells on day 4 were higher compared with cells cultured under the static condition (Fig. 3D). These results suggest that the cyclic strain applied by the bioreactor can efficiently promote the myogenic differentiation of C2C12 cells with cellular alignment.
Effects of cyclic strain on the myogenesis of mBMSCs
We next investigated the effects of cyclic strain on the skeletal myogenesis of BMSCs. Under the cyclic strain condition in the myogenic medium, the mBMSCs aligned in a unidirectional manner and fused to form multinucleated myotube-like cells (Fig. 4A) with nuclei that were aligned in the direction of tension application only after 5 days (Fig. 4B). On the other hand, when mBMSCs were cultured in the myogenic medium under the static condition, a small number of cells showed myotube-like morphology after 14 days, but the multinucleation of these cells was incomplete (Fig. 4D). The number of myotube-like multinucleated cells among the total cell population under the cyclic strain condition increased over the time course of the experiment (Fig. 4E). These multinucleated cells were positively stained for the myogenin and myosin proteins (Figs. 4F, H). The mBMSCs cultured in the same medium under the static condition showed neither myotube-like morphology (Fig. 4C), nor were stained for the myogenin and myosin proteins within 5 days (Fig. 4G, I).

The effects of cyclic strain on the differentiation of mBMSCs. (A-C) Phase-contrast images of mBMSCs cultured in myogenic medium under cyclic strain
Effects of cyclic strain on gene expression during the mBMSC myogenesis
When mBMSCs were cultured in the growth medium, they did not express the Myf5, myogenin, and MRF4 genes under the static and cyclic strain conditions (data not shown). In contrast, mBMSCs cultured in the myogenic medium under the cyclic condition demonstrated clearly increased mRNA expression of Myf5 and myogenin on day 5, and MRF4 on day 7 (Fig. 5A, B). The expression of MyoD was not detected in cells grown under either culture condition by the primer sets, including the TaqMan® primer and probe system used in this study (Fig. 5B). The expression of myocardin, which is a cardiac- and smooth muscle-specific transcriptional factor, was not detected at the mRNA level in mBMSCs cultured under either the static or cyclic strain conditions (Fig. 5B). The gene expression of α-smooth muscle actin (α-SMA), a differentiation marker of smooth muscle cells, was inhibited by the cyclic strain (Fig. 5B). The mRNA expression of tension-induced proteins (TIP)-1 and immunoglobulin heavy chain-binding protein/glucose-regulated protein 78 kDa (BIP/GRP78; also known as heat shock 70-kDa protein [5HSPA5]) was induced by the cyclic strain, while the expression of tension-inhibited proteins (TIP)-3 was inhibited on day 7 (Fig. 5B).
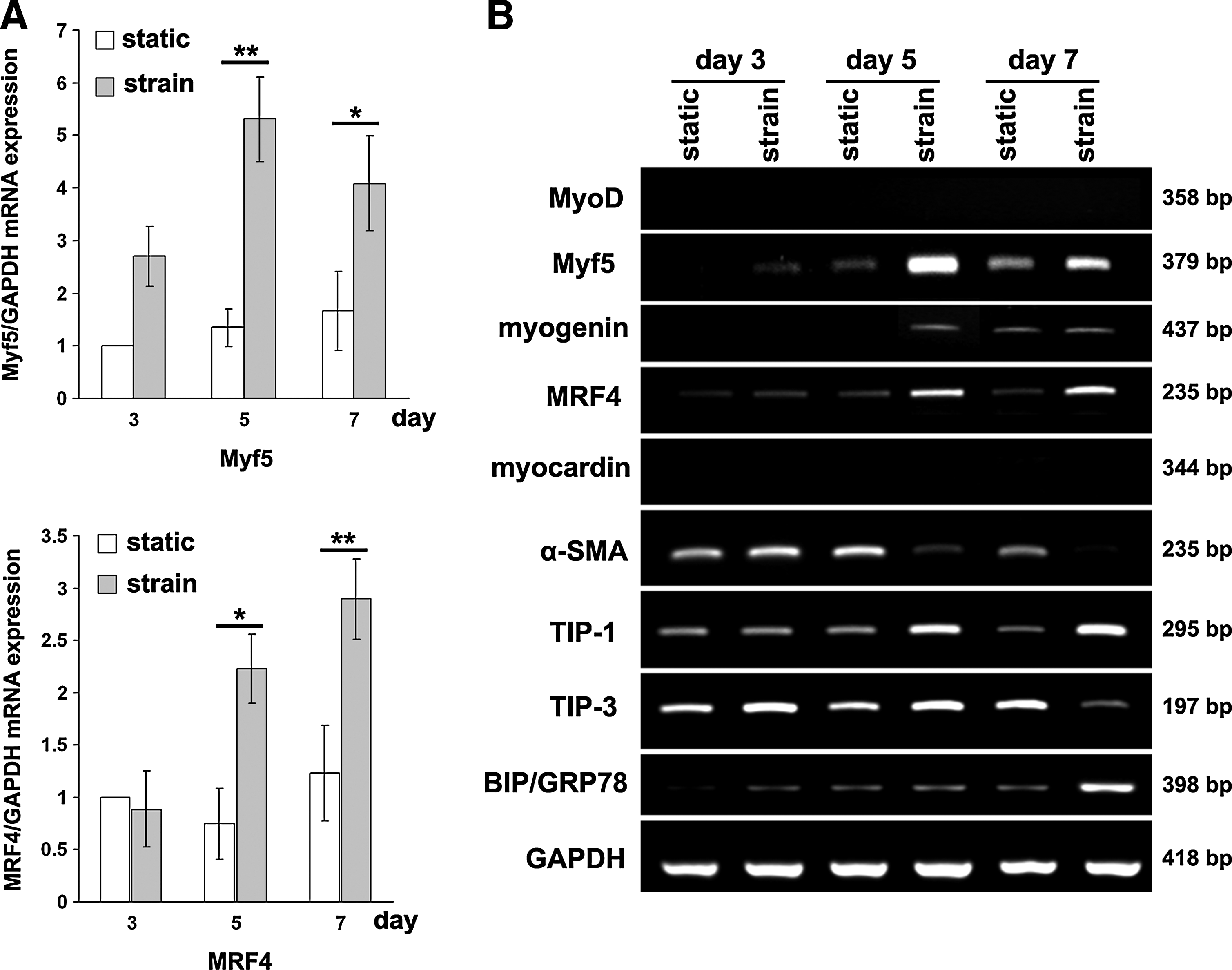
The effects of cyclic strain on the expression of myogenic genes during the myogenesis of mBMSCs. The mBMSC were cultured in myogenic medium under static or cyclic strain conditions.
Discussion
Our results show that mouse myoblasts and BMSCs respond to cyclic strain by changing their orientation to align in a unidirectional manner within 48 h. Previously, Pennisi et al. 17 demonstrated using a finite element model of a uniaxial cyclic strain system that a rectangular area in the center part of the stretching substrate is subjected to the highest strain force. Therefore, we investigated the cellular orientation at the center of each silicone sheet. It is known that actin fibers adjust their length and orientation to an equilibrium, where extension is minimized when cells are extended beyond a certain threshold. 17 A sequential observation indicated that BMSCs cultured under the cyclic strain condition became rounded and smaller at the initial stage of the cellular alignment process, suggesting that the force of the cyclic strain initially led to a gradual loss of cell attachment to the silicone surface at focal adhesion sites. The rounded cells were then gradually elongated, with alignment of the F-actin cytoskeleton, and the cell long axis also became aligned. These results suggest that the cells reorganized their cytoskeletal network according to the strain force and vector, which led to the formation of aligned cytoplasmic actin-containing stress fibers that terminated in adhesion complexes 19 on the stretching silicone surface.
At the molecular level, previous reports suggested that cytoskeletal signaling involving focal adhesion kinase (pp125FAK), 31 paxillin, 32 and stretch-activated Ca2+ channels 33 regulates cell alignment. Several studies that have examined the effects of cyclic strain on cell orientation showed perpendicular alignment of rBMSCs, 34 human BMSCs, 35 rat adipose-derived stem cells (ASCs), 36 C2C12 cells, 17 and human aortic endothelial cells 37 to the direction of the cyclic strain. On the other hand, mBMSCs cultured in type I collagen gels, 38 rBMSCs cultured in a 3D fibrin matrix, 39 and human umbilical vein endothelial cells 40 aligned parallel to the direction of the cyclic strain. It should be noted that the cellular responses to mechanical stress can differ according to the magnitude and duration of the applied mechanical strain, and the frequency at which the strain is applied. 41 Interestingly, our results showed that although the same cyclic strain condition was applied, the alignment direction of rBMSCs, C2C12 cells, and human gingival fibroblasts cultured in the growth medium was vertically different from that of mBMSCs. The differences in the direction of the cellular alignment among the cell types may partly depend on the cell type-specific expression pattern of cell adhesion molecules, 42 which govern the cytoskeletal signaling as mechanosensitive receptors. 43 However, how they are acting directly on the cell orientation remains to be determined.
The fact that geometric stimuli influence myoblast differentiation is now well accepted. 16 Yamamoto et al. 13 demonstrated that culturing C2C12 cells on micropatterned glass results in the formation of myotubes, organized in parallel, much like the arrangement in muscle tissue. Wang et al. 15 showed that C2C12 cells were aligned along the direction of submicron grooves, which enhanced myotube formation. On the other hand, C2C12 cells also sense and respond to cyclic strain stimuli to induce GTP-binding proteins, which play a central role in signal transduction. 44 To this end, we have applied the cyclic strain to align C2C12 myoblasts to enhance skeletal myogenesis. Besides producing cell alignment, the cyclic strain enhanced the myogenic phenotype of C2C12 cells. These results are consistent with the findings from a recent report showing that uniaxial cyclic strain drives the assembly of C2C12 cells and increases the percentage of myosin-positive myotubes. 17 Generally, myogenesis is initiated by the expression of the MRFs (Myf5, MyoD, MRF4, and myogenin), a group of four related basic helix-loop-helix transcription factors. 45 In the present study, we provided the first evidence that enhanced gene expression of these MRFs in C2C12 cells was induced by the cyclic strain stimuli, which supports the usefulness of the cyclic strain for inducing the physiological differentiation of skeletal myocytes in vitro.
Cell proliferation and differentiation are generally alternative processes that mutually affect the determination of the stem cell phenotype. 46 Our results show that cyclic strain terminated the proliferation of mBMSCs cultured in the growth medium, which was consistent with the findings from a previous study. 34 In addition, we observed that the motility of cells cultured in the growth medium was markedly restricted under the cyclic strain condition during cellular orientation, which resulted in tight contact among cells. The tight contact of myoblasts provides remodeling of the extracellular matrix environment and allows for cellular fusion. Therefore, it is possible that the mBMSCs cultured under cyclic deformation switched off the expression of somatic genes associated with cell division, and the cells compressed each other during their alignment. However, the cyclic strain itself appears not to result in the lineage commitment of BMSCs to skeletal myogenic cells because it did not induce mBMSCs cultured in the growth medium to express skeletal myogenic marker genes. Therefore, trophic factors provided by a myogenic medium are necessary to initiate the skeletal myogenesis of BMSCs subjected to cyclic strain. Once these trophic factors induce the mBMSCs to commit to a skeletal myogenic lineage, the tight contact of each cell provided by appropriate cellular alignment should facilitate cellular fusion to promote skeletal myogenesis.
Indeed, Bayati et al. 36 showed that cyclic uniaxial strain coupled with a myogenesis-inducing demethylating chemical factor (5-azacytidine) improved the myogenic properties of rat ASCs; however, the resulting myogenic induction was not sufficient to achieve cell fusion. In the present study, mBMSCs cultured in the myogenic medium that were subjected to cyclic strain formed apparent aligned, myotube-like structures. Enhanced skeletal myogenesis was evidenced by a significant increase in the number of multinucleated cells and in the expression of skeletal myogenic marker proteins, such as myogenin and myosin. Our results showed that the use of only the skeletal myogenic medium resulted in the mBMSCs exhibiting myotube-like morphology within 14 days; whereas the additional application of cyclic strain greatly shortened the induction period to 5 days. Therefore, the combined use of the induction medium and the cyclic strain could be a simple, but powerful tool to induce the skeletal myogenesis of BMSCs. However, sarcomeric actin was not observed in the myotube-like structures of mBMSCs, whereas it appeared in some of the C2C12 myotubes under the cyclic strain condition. Therefore, the skeletal myogenesis of mBMSCs was immature when compared with that of C2C12 myoblasts subjected to the same induction method, possibly because mBMSCs are uncommitted stem cells, whereas C2C12 cells are already committed to a myoblast lineage and can, thus, more easily develop to mature myotubes. It should be also noted that although the skeletal myogenic induction of mBMSCs subjected to cyclic strain seemed to mimic some aspects of conventional skeletal muscle development in that the early expression of Myf5 and myogenin and the subsequent expression of MRF4 were detected, the expression of MyoD was not detected by the primer sets used in this study. It has recently been reported that MRFs can be bypassed altogether in some muscle stem cells. 47 Because the characteristics of the BMSCs used in this induction method are different from those of conventional myogenic progenitor cells, such as C2C12 cells, it is possible that some of the underlying differentiation mechanisms may differ, especially in the initial step in which BMSCs are committed to become myogenic progenitor cells. Although our data demonstrate that cyclic strain promotes the skeletal myogenesis of mBMSCs, a definitive method to commit all BMSCs to the skeletal myogenic lineage at the initial induction stage is necessary.
Our induction method specifically guided the differentiation of mBMSCs into skeletal myogenic cells, but not into smooth muscle cells, as demonstrated by the lack of expression of myocardin and the decreased expression of α-SMA during differentiation. However, it has also been shown that cyclic strain induces vascular smooth muscle cell differentiation in human endothelial cells 40 and mouse embryonic stem cells. 48 In addition, BMSCs have been reported to show smooth muscle cell-like properties induced by cyclic tensile strain,34,35 cyclic contractile force in type I collagen gels, 38 or shear and compressive stress. 49 In these studies, mechanical stress was applied to cells in the general growth medium or the smooth muscle growth medium. In contrast, in the present study, we used a skeletal myogenic medium 12 during the application of cyclic strain, which may explain the discrepancy between our findings and those of previous studies.
In the present induction method, mBMSCs were seeded at a density of 3×103 cells/cm2 and demonstrated 80%–90% confluency when the cyclic strain was applied. When the mBMSCs were seeded at a lower density (1.5×103 cells/cm2), they did not demonstrate apparent myotube formation upon induction (data not shown), which suggests that a high seeding density, which permits the cellular alignment to provide tight contact among cells, is necessary for the cells to undergo cellular fusion to promote myogenesis. Therefore, to maximize the skeletal myogenic potential of BMSCs, we consider that the application of cyclic strain to BMSCs cultured in the myogenic medium should be performed when the cells reach 80%–90% confluence on the silicone sheet.
Although the tight contact among cells aligned by the cyclic strain was likely the main factor underlying the enhanced myogenesis of mBMSCs in our method, several reports have demonstrated the ability of mechanical signals to influence BMSC fate.50–52 Jakkaraju et al. 53 previously identified TIPs as stretch-responsive factors during myogenesis of lung embryonic mesenchymal cells. They found that TIP-1 is induced by stretch and stimulates myogenesis, whereas TIP-3 stimulates adipogenesis and is suppressed by stretch. In mBMSCs, the expression of TIP-1 was induced by cyclic strain, while the expression of TIP-3 was inhibited. These results support the results of Jakkaraju et al., and imply that these stretch-responsive factors may be involved in the myogenic differentiation of BMSCs under cyclic strain conditions. On the other hand, other recent reports suggested that the application of mechanical stress to cells triggers endoplasmic reticulum (ER) stress,54,55 which is defined as the accumulation of unfolded or misfolded proteins in the ER, resulting in a coordinated adaptive program. 56 Nakanishi et al. 57 reported that ER stress signaling exerts a positive effect on myogenic differentiation. In this study, the application of cyclic strain induced the expression of an ER stress marker, BIP/GRP78, 58 in the mBMSCs. Therefore, it is possible that the cyclic strain stimulated ER stress signaling in BMSCs, resulting in the coordination of gene transcription and/or transcriptional controls to promote skeletal myogenic differentiation. These results imply that such stretch-responsive factors may, at least in part, be involved in the myogenic differentiation of mBMSCs under cyclic strain conditions. Further studies are required to determine the possible role that cyclic strain directly plays in the enhancement of BMSC skeletal myogenesis.
It should be noted that the stiffness of the matrix on which the cells are grown is also an important factor for the lineage specification of stem cells. Engler et al. 59 demonstrated that a matrix with a stiffness similar to that of striated muscle (i.e., elastic modulus of 8–17 kPa), but not softer or stiffer matrices, induced the skeletal myogenic differentiation of human mesenchymal stem cells (MSCs). In our study, skeletal myogenic induction by the matrix is thus expected to be minimal because the PDMS silicone sheet was considerably stiffer (Young's modulus >600 Kp) 60 than actual muscle tissue. In this regard, Quinlan et al. 61 recently developed a method for culturing cells under controllable substrate stiffness and dynamic stretch conditions and suggested that similar mechanotransduction mechanisms exist for sensing stiffness and stretching. Therefore, simultaneous manipulation of the cyclic strain and matrix stiffness may provide a greater control over the guided skeletal myogenesis of BMSCs than the application of cyclic strain alone.
Notably, Choi et al. 62 demonstrated that human ASCs had a superior sensitivity and capacity for skeletal myogenic differentiation on matrices that mimicked skeletal muscle elasticity when compared with BMSCs, possibly because the ASCs formed mechanosensing focal adhesion assemblies earlier than the BMSCs. Choi et al. thus suggested that specific differences in mechanosensor expression (e.g., that of integrins and vinculin in focal adhesion signaling pathways) and force transduction, which may depend on the source used to derive MSCs, also affect mechanically induced skeletal myogenesis. Therefore, a defined cell expansion protocol to reinforce the mechanosensor expression of BMSCs before a cyclic strain application may hold promise for optimal skeletal myogenesis in BMSCs. Additionally, signals mechanically activated by cyclic strain and/or substrate stiffness could be integrated with growth factor signaling within the cell, which should be examined in future studies.
In conclusion, the present study demonstrated that the application of cyclic strain induced neat alignment of mBMSCs and greatly enhanced the cellular skeletal myogenic differentiation (Fig. 6). To our knowledge, this is the first report to show that cyclic strain can be utilized to promote the skeletal myogenesis of BMSCs, although potential differences in the response to the cyclic strain between human and mouse BMSCs remain to be elucidated. For skeletal muscle tissue engineering, aligned myogenic cells should be beneficial for promoting enhanced function. Our simple method of applying cyclic strain to cells cultured in the myogenic medium may represent a promising approach for future clinical applications and provide an important step toward creating more physiologically analogous replacement tissue.
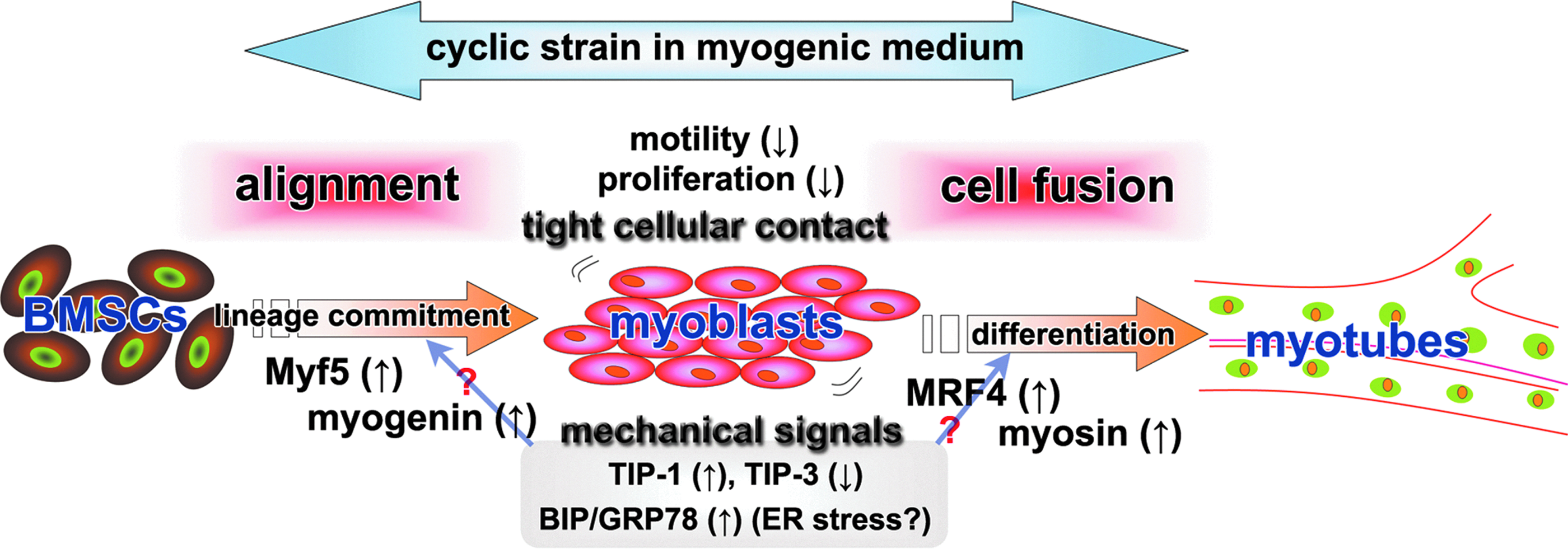
A schematic diagram depicting the enhancement of mBMSC skeletal myogenic differentiation by our method. Uniaxial cyclic strain applied to mBMSCs cultured in a myogenic medium aligns the cells and restricts their motility and proliferation, which leads to tight cellular contact in the cell population. Once the trophic factors in the myogenic medium induce the mBMSCs to commit to a skeletal myogenic lineage, the tight contact among cells provided by the appropriate cellular alignment facilitates cellular fusion to promote skeletal myogenesis. Additionally, the cyclic strain stimulates the gene expression of a tension-induced protein (TIP-1) and an ER stress marker (BIP/GRP78), while reducing the expression of a tension-inhibited protein (TIP-3), which may also facilitate myogenic differentiation. Color images available online at www.liebertpub.com/tea
Footnotes
Acknowledgments
This investigation was supported by a Grant-in-Aid for Exploratory Research from Japan Society for the Promotion of Science (19659505, H.E.) and Grant-in-Aid for Young Scientists (B: 22791879, M.K.) from the Ministry of Education, Culture, Sports, Science and Technology, Japan. Support was also received from the Cosmetology Research Foundation, Japan (H.E.).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.