
Abstract
Stem cells with broad differentiation potential, such as the recently described germline-derived pluripotent stem cells (gPS cells), are an appealing source for tissue engineering strategies. Biomaterials can inhibit, support, or induce proliferation and differentiation of stem cells. Here we identified (1) polymers that maintain self-renewal and differentiation potential of gPS cells for feeder-free expansion and (2) polymers supporting the cardiomyogenic fate of gPS cells by analyzing a panel of polymers of an established biomaterial bank previously used to assess growth of diverse stem cell types. Identification of cytocompatible gPS cell/biomaterial combinations required analysis of several parameters, including morphology, viability, cytotoxicity, apoptosis, proliferation, and differentiation potential. Pluripotency of gPS cells was visualized by the endogenous Oct4-promoter-driven GFP and by Sox2 and Nanog immunofluorescence. Viability assay, proliferation assay, and flow cytometry showed that gPS cells efficiently adhere and are viable on synthetic polymers, such as Resomer® LR704 (poly(L-lactic-D,L-lactic acid), poly(tetrafluor ethylene) (PTFE), poly(vinylidene fluoride) (PVDF), and on gelatine-coated tissue culture polystyrene. Expansion experiments showed that Resomer LR704 is an alternative substrate for feeder-free gPS cell maintenance. Resomer LR704, PTFE, and PVDF were found to be suitable for gPS cell differentiation. Spontaneous beating in embryoid bodies cultured on Resomer LR704 occurred already on day 8 of differentiation, much earlier compared to the other surfaces. This indicates that Resomer LR704 supports spontaneous cardiomyogenic differentiation of gPS cells, which was also confirmed on molecular, protein and functional level.
Introduction
gPS cells originate from unipotent germline stem cells (GSC), which were defined in Ko et al. by expression of germ cell markers and proof of unipotency. GSC express Oct4 and thus GSC can be induced to acquire pluripotency without exogenous transcription factors by employing specific culture conditions. 10 gPS cells exhibit a gene expression repertoire that is very similar to ES cells and pluripotency of gPS cells was confirmed by in vitro and in vivo differentiation, including germ cell contribution and transmission. 10 gPS cells were so far only obtained from mouse testis and studies on human testis-derived pluripotent stem cells have remained highly controversial and are a challenge for future studies. 12
Applications of biomaterials have become an important field in regenerative medicine. Biomaterials, such as polymers, metals, or ceramics, can serve as scaffolds for cells and can, for example, influence stem cell growth and differentiation. Neuss et al. 13 performed an in-depth study on stem cell/biomaterial interactions for identification of adequate combinations for diverse tissue engineering applications. According to this study, stem cell types behave differently on each material. Thus, gPS cell behavior was analyzed on polymers—based on this standardized test platform 13 —that showed at least one positive feature for stem cell cultivation. Since cultivation of ES cells on gelatine is well established, it was additionally included in our test platform. All used polymers are listed in the biomaterial bank (Fig. 1), which was established in cooperation with the Department of Textile and Macromolecular Chemistry of RWTH Aachen (ITMC). For tissue engineering, biomaterials are used as scaffolds for cells and can, as already mentioned above, influence their growth and proliferation. In addition, specific materials can repress or induce differentiation of stem cells. 14 Some of these polymers have already been described in combination with several stem cell types, dendritic cells, and ES cell-derived cardiomyocytes.13,15–18 Compatible gPS cell/biomaterial combinations were identified and selected for the present study. Here we use poly(tetrafluor ethylene (PTFE), poly(vinylidene fluoride) (PVDF) and Resomer® LR704 because of their low cytotoxic effects on gPS cells and high viability rates of gPS cells.
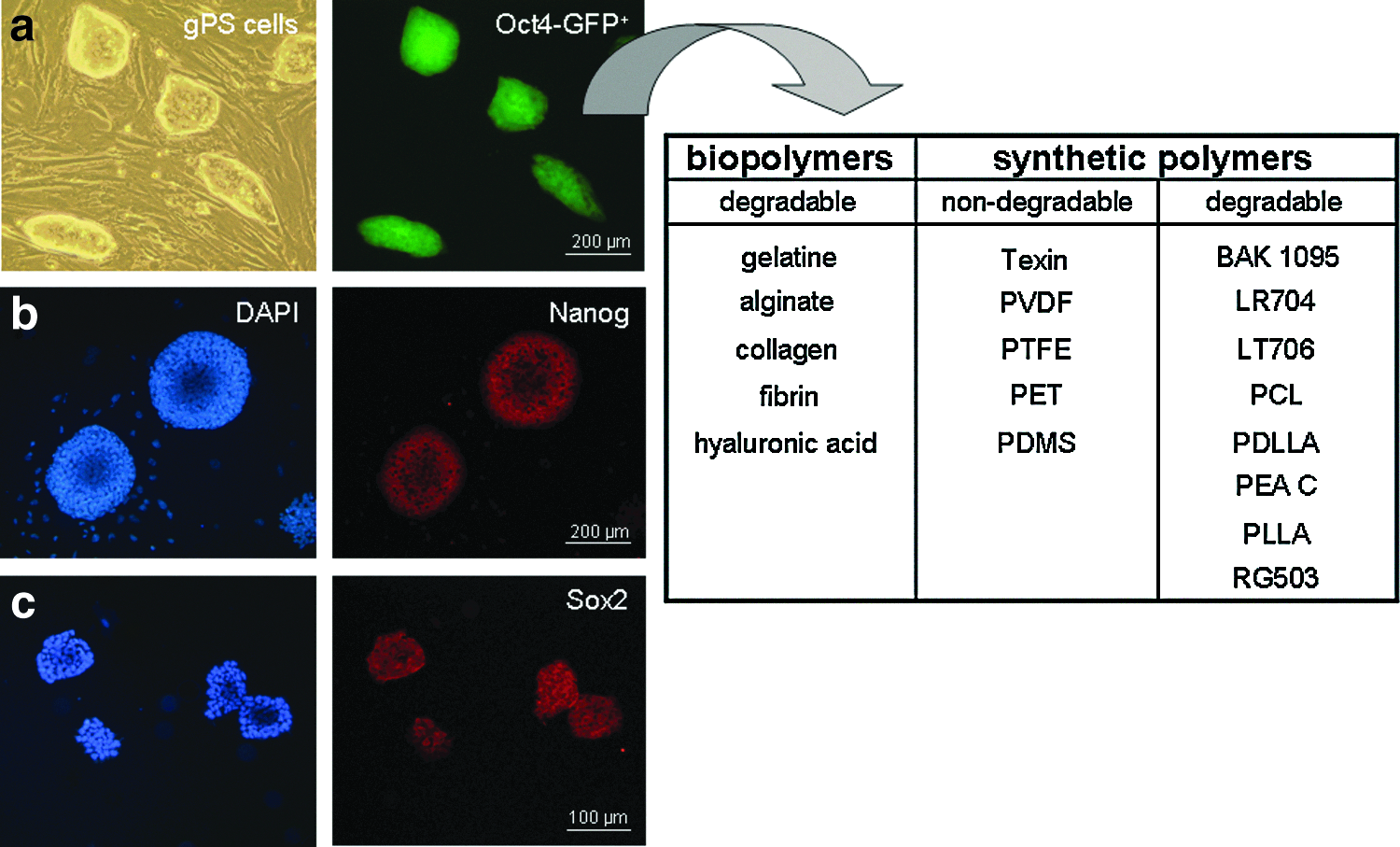
Study design. Germline-derived pluripotent stem cells (gPS cells) were cultivated on polymers of the established biomaterial bank,13 including gelatine, alginate, collagen, fibrin, hyaluronic acid (HA), Texin, poly(vinylidene fluoride) (PVDF), poly(tetrafluor ethylene) (PTFE), poly(ethylene terephthalate) (PET), poly(dimethyl siloxane) (PDMS), polyesteramid type BAK-1095® (BAK), poly(L-lactic-co-D,L-lactic acid) (Resomer® LR704), poly(L-lactic acid-co-trimethylene carbonate) (Resomer LT706), poly(ɛ-caprolactone) (PCL), poly(D,L-lactic acid) (PDLLA), polyesteramid type C (PEA C), poly(L-lactic acid) (PLLA), and poly(D,L-lactic-co-glycol acid) (Resomer RG503). Undifferentiated gPS cells on MEF:
Differentiation of ES cells into cardiomyocytes has been described oftentimes.19–24 Since years of research even innovative methods do not ensure derivation of homogenous cell populations from pluripotent stem cells. Whereas quality of ES cell-derived cardiomyocytes seems remarkably good, purity, and quantity are limiting factors. 19 Therefore, differentiation protocols need to be improved for successful tissue engineering. Cardiac tissue engineering is an emerging field. For this reason, it is important to determine if polymers can support differentiation of gPS cells into cardiomyocytes. In the current project we investigated the growth behavior of gPS cells on different biopolymers and synthetic polymers to identify materials for feeder-free expansion and cardiac differentiation of gPS cells. We conclude that the development of biohybrid systems composed of polymers and human gPS cells is the prospective objective for biomedical applications.
Materials and Methods
Materials
Poly(tetrafluor ethylene) and poly(vinylidene fluoride)
PTFE and PVDF foils were bought from GoodFellow Cambridge Ltd., Huntingdon, England. Small blankets in 6-, 24- and 96-well format were tailored. PTFE and PVDF slides were extracted in a Soxhlet apparatus using ethanol and hexane (21:79 v/v) for 2 h. Afterward, foils were first dried at 80°C for 1 h and then for 48 h in the evacuator.
Resomer LR704
Resomer LR704 (poly(L-lactic-co-D,L-lactic acid) ratio 70:30) polymer granules were bought from Boehringer Ingelheim and polymer foils were prepared by a melt press process. The metal plates of the melt press were covered by PTFE foils (9 cm diameter) and 1.2 g of granules was placed at the centre of these foils. The temperature was raised to 200°C and maintained for 3 min, which is the melting temperature of Resomer LR704. A load of 1 ton was applied for 1 min at the same temperature. Foils were cut into a desired size. Afterward, Resomer LR704 foils were cleaned by rinsing in isopropanol and dried by nitrogen stream and finally in the evacuator for 2 h.
The other used materials, like the degradable biopolymers alginate, collagen, fibrin, and hyaluronic acid, the nondegradable synthetic polymers Texin®, poly(ethylene terephthalate) (PET), and poly(dimethyl siloxane) (PDMS), as well as the degradable synthetic polymers polyesteramide type BAK-1095® (BAK), poly(L-lactic acid-co-trimethylene carbonate) (Resomer LT706), poly(ɛ-caprolactone) (PCL), poly(D,L-lactic acid) (PDLLA), polyesteramide type C (PEA C), poly(L-lactic acid) (PLLA), and poly(D,L-lactic-co-glycolic acid) (Resomer RG503) have been prepared following the protocols of Neuss et al. 13
gPS cells
Murine gPS cells originate from Oct4-GFP transgenic (OG2 and OG2 ROSA26, respectively) mice and were described before. 10
Expansion
Expansion of gPS cells (6.3×103 cells/cm2) was performed on gelatine-coated (0.1%) culture dishes on inactivated mouse embryonic fibroblasts (MEF, 2×104 cells/cm2) in Dulbecco's Modified Eagle Medium (DMEM) low glucose (1 g/l, PAA; Cölbe) supplemented with 15% FCS (Lonza), penicillin and streptomycin (100 U/mL), β-mercaptoethanol (100 μM), L-glutamine (2 mM), Hepes (25 mM), and nonessential amino acids (0.1 mM; all Gibco Invitrogen). Leukemia inhibitory factor (ESGRO® LIF; Chemicon) was added freshly to the medium (10 ng/mL). gPS cells were cultured in a 21% O2 and 5% CO2 humidified atmosphere at 37°C. gPS cells were split every 2–3 days.
For expansion of gPS cells on biomaterials a depletion step about 15 min guaranteed separation from MEF for further cultivation.
Embryoid body formation
For differentiation of gPS cells, embyoid body formation that provides cell-cell contacts in suspension was prepared. Trypsinized gPS cells (2×104 cells/cm2) were transferred to a Petri dish after depletion of MEF. Differentiation medium contained DMEM high glucose (4.5 g/L, PAA; Cölbe), 10% FCS (Lonza), penicillin and streptomycin (100 U/mL), β-mercaptoethanol (100 μM), L-glutamine (2 mM), Hepes (25 mM), and nonessential amino acids (0.1 mM; all Gibco Invitrogen). gPS cells were cultured in a 21% O2 and 5% CO2 humidified atmosphere at 37°C. To change medium in the mass culture every second day, embryoid bodies (EBs) were sedimented in a tube with a conical bottom and supernatant was discarded. After 4 days, EBs were transferred to polymers. Single EBs were picked with the pipette and transferred to one 24-well (1 EB/24-well). Medium change was done every 1–2 days.
Experimental procedures
Immunofluorescence
Cells were fixed in 4% PFA (Merck) in phosphate-buffered saline (PBS; Gibco BRL) followed by permeabilization with 3% (v/v) horse serum and 0.5% (v/v) Triton X-100 (Sigma) in 1% bovine serum albumin (BSA; Sigma) in PBS. Nanog (rabbit polyclonal, catalog number LS-B1193; Lifespan Biosciences) and Sox2 antibodies (rabbit polyclonal, catalog number PA1-16968; Thermo Scientific) were diluted 1:50 in 1% BSA in PBS and incubated overnight at room temperature. Cy3-conjugated goat anti-rabbit IgG (1:400; Dianova) was used as a secondary antibody. Samples were covered in DAPI-mounting medium (Dako), observed and photographed with fluorescent microscope (Axiovert 25; Zeiss).
Cell proliferation and cytotoxicity (multiplex assay)
Stem cells were seeded on polymer slides in 96-well format (1300 cells/cm2) in 200 μL of medium for 24 h. Cell viability test was performed by adding 20 μL CellTiter-Blue® reagent (CellTiter-Blue Cell Viability Assay; Promega) to each well, incubation for 1h at 37°C, and transfer of 100 μL supernatant into a black 96-well plate to measure fluorescent resorufin signals (560 nmEx/590 nmEm), which detects cell metabolism. Proliferation of stem cells was tested by viability assay at day 1, 2, and 3 after cell seeding.
For quantification of cytotoxicity, 100 μL of supernatant was transferred into a black 96-well plate and mixed with cytotoxicity reagent (1:1 v/v; CytoTox-One™ Homogenous Membrane Integrity Assay; Promega). Extracellular lactate-dehydrogenase (LDH) was detected after 10 min by fluorometric resorufin measurement (560 nmEx/590 nmEm). As a positive control for cytotoxicity assay, 2 μL lysis buffer of the kit was added to the supernatant and cytotoxicity reagent to permeabilize cell membranes and thus causes LDH release. Each assay was performed in quadruplicates. Interference of LDH with phenol red in the medium was charged against medium without cells as a blank. Data are presented as the mean±standard error of the mean (SEM). A one-way analysis of variance followed by a Dunnett's multiple comparison test was applied for statistical evaluation. p values of the fluorescence intensity of preparations on biomaterials were evaluated in comparison to control. Statistical significance was defined as p<0.05.
Flow cytometry
gPS cells were seeded on polymers in six-well format (1.4×104 cells/cm2) for 24 h. Cells were trypsinized, washed two times in cold PBS, and resuspended in 100 μL binding buffer, 5 μL APC Annexin V, and 1 μL 7-AAD (BD Biosciences). After incubation for 15 min in the dark at room temperature, 400 μL binding buffer was added. Probes were analyzed within 1 h. Flow cytometry was performed using FACSCanto flow cytometer (BD Biosciences) and FlowJo Flow Cytometry Analysis Software (Tree Star, Inc.,). For elimination of MEF in the control, a depletion step of 15 min on gelatine-coated dishes has been carried out.
Reverse transcription real-time quantitative polymerase chain reaction
Total RNA was extracted using the RNeasy Mini Kit from Qiagen. RNA concentration (A260 nm) and purity (A260/A280 nm>2) was measured by NanoDrop-1000 Spectrophotometer (Peq Lab Biotechnologie). After DNase I digestion (Roche), total RNA (1 μg per 20 μL of reaction volume) was reverse transcribed into cDNA using a High Capacity cDNA Reverse Transcription Kit (Applied Biosystems). Single-stranded cDNA products (1.2 μL in a total reaction volume of 20 μL) were analyzed with real-time TaqMan polymerase chain reaction (PCR) using a master-mix consisting of 6.8 μL diethylpyrocarbonate treated H20 (Sigma), 10 μL SybrGreen (Roche) and 1 μL PCR primers (20 pmol/μL). The level of expression of each target gene was normalized to the reference gene β-actin. Reverse transcription real-time quantitative PCR (RT-qPCR) was performed with the 7300 Real Time PCR System (Applied Biosystems) at 95°C for 10 min followed by 40 cycles of 15 s of denaturation at 95°C, 1 min annealing/elongation at 60°C. 7300 System SDS Software (Applied Biosystems) was used for analysis. Primer sequences were used as follows: β-actin forward 5′-CTACGTCGCCCTGGACTTCGAGC-3′, β-actin reverse 5′-GATGGAGCCGCCGATCCACACGG-3′ (Sigma-Aldrich), α-fetoprotein forward 5′-TCAGCGAGGAGAAATGGTCC-3′, α-fetoprotein reverse 5′-ATGAAAATGTCGGCCATTCC-3′, α-myosin heavy chain (α-MHC) forward 5′-CATGGCTACACTCTTCTCTACCTATGC-3′, α-MHC reverse 5′-TACCACTGTCACCGGTATCAGC-3′, β-myosin heavy chain (β-MHC) forward 5′-GAGAGGGTGGCAAAGTCACTG-3′, β-MHC reverse 5′-CAGTCACCGTCTTGCCATTCT-3′, nestin forward 5′-TGGAGGGTTGCGTCGG-3′ nestin reverse 5′-GCGTCGATTGAGCTCCCA-3′, Oct4 forward 5′-AACCACTCCCGAGGAGTC-3′, Oct4 reverse 5′-CTAGCTCCTTCTGCAGGGCTT-3′ (all Eurofins MWG). The expression level of each target gene was normalized to β-actin as the housekeeping gene and then calculated by the ΔΔCT method. Data are presented as the mean±SEM. A one-way analysis of variance followed by a Dunnett's multiple comparison test was applied for statistical evaluation. P values of the gene expression levels of gPS cells on biomaterials were calculated in comparison to gene expression levels of undifferentiated gPS cells as control. Statistical significance was defined as p<0.05.
Immunohistochemistry
EBs, cultivated for 4 days in mass culture and for another 10 days on polymers, were fixed in 3.7% formaldehyde for 24 h, embedded in paraffin, and cut with a rotating microtome at 3 μm thickness (Leica). Staining of germ layer-specific proteins, desmin (1:20, monoclonal anti-desmin, clone 10519; Progen Biotechnik), and smooth muscle actin (SMA) (1:3000, monoclonal anti-actin, α-smooth muscle antibody, catalog number A2547; Sigma-Aldrich), and cardiac proteins, such as sarcomeric α-actinin (1:100, monoclonal anti-α-actinin (sarcomeric), clone EA-53, catalog number A7732; Sigma-Aldrich) and connexin 43 (1:100, rabbit polyclonal connexin 43/GJA1, catalog number ab47368; Abcam), were performed by Dako REAL™ Detection System Peroxidase/DAB+ K5001 Kit (Dako). Primary antibodies were incubated at room temperature for 25 min. The biotinylated secondary antibody was applied for 15 min followed by color development with streptavidin-peroxidase HRP and 3,3′-diaminobenzidine (DAB+; all Dako). Hematoxylin was used for counterstaining. Slides were dehydrated and mounted in Vitro-Clud (Langenbrinck).
Electron microscopy
Undifferentiated gPS cells were cultured for scanning electron microscopy (SEM) on polymers for 24 h under expansion conditions, fixed in 3% glutaraldehyde for at least 24 h, rinsed with sodium phosphate buffer (0.1 M; Merck), and dehydrated by incubating consecutively in ascending alcohol series (30%, 50%, 70%, 90%) with a final incubation in 100% ethanol for 10 min three times. Probes were critical-point-dried in liquid CO2, and then sputter-coated with a 30-nm gold layer. Samples were analyzed using an environmental scanning electron microscope (ESEM XL 30 FEG, FEI; Philips) in a high vacuum environment.
Sharp electrode technique
Sharp electrode measurements were performed at the Institute of Neurophysiology, University of Cologne. Intracellular action potentials were recorded in spontaneously beating areas of plated EBs that were perfused with DMEM at 37°C bubbled with 5% CO2 and 95% O2. Measurements were done using pipettes filled with KCl (3 M) at resistances of 30–50 MΩ. Signals were amplified by a SEC10-LX (npi electronic) and converted by an EPC-9 (HEKA). Data were stored with the PULSE software (HEKA) and analyzed with the MiniAnalysis software (Synaptosoft). Data are presented as mean±SEM. Unpaired Student's t test was applied for statistical evaluation, p<0.05 as level of significance.
Microarray assay
Microarray analyses were arranged by Bernd Denecke (Chip-Facility IZKF Aachen, RWTH Aachen University). Total RNA of undifferentiated gPS cells and 12-day-old EBs on gelatine and Resomer LR704 (each n=3) was isolated (see chapter 2.3.4) and the quality assessed using RNA NanoChips with the Agilent 2100 Bioanalyzer (Agilent). Probes for the GeneChip® Mouse Gene 1.0 ST Arrays (Affymetrix) were prepared and hybridized to the arrays according to the Affymetrix GeneChip Whole Transcript (WT) Sense Target labeling Assay Manual. Briefly, for each sample, 300 ng of total RNA was reverse transcribed into cDNA using random hexamer oligonucleotides tagged with a T7 promoter sequence. After second strand synthesis, the double-stranded cDNA was used as the template for amplification with T7 RNA polymerase to create antisense cRNA. Random hexamers were then used to reverse transcribe the cRNA into single stranded sense strand cDNA. The sense cDNA was then fragmented by uracil DNA glycosylase (UDG) and apurinic/apyrimidic endonuclease 1. Fragment size was validated using a Bioanalyzer (Agilent) with optimal fragment size in the 50–200 bp range. Fragmented sense cDNA was biotin-labeled with terminal deoxynucleotidyl transferase. The probes were then hybridized to the GeneChip Mouse 1.0 ST Arrays at 45°C for 16 h, washed and stained on Fluidics Station 450, and finally scanned on a GeneChip Scanner 3000 7G (Affymetrix). The image data were analyzed with AGCC (Affymetrix). Gene expression levels were preprocessed with RMA algorithm. Concluding comparison of transcriptional profiles of undifferentiated gPS cells and differentiated gPS cells on gelatine and Resomer LR704 was performed by principal component analysis (PCA) and hierarchical cluster analysis (heat maps) in cooperation with Qiong Lin (Institute for Biomedical Engineering, Department of Cell Biology, RWTH Aachen University).
Results
Pluripotency of gPS cells
gPS cells grown on MEF formed colonies with an ES cell-like morphology, which expressed GFP under the control of endogenous Oct4 promoter and the pluripotency markers Nanog and Sox2 as visualized by fluorescence microscopy and immunohistochemistry (Fig. 1). These data indicate that gPS cells maintained the phenotype of pluripotent stem cells under these culture conditions.
Vitality and proliferation of gPS cells on polymers
Vitality and proliferation of gPS cells on biomaterials were measured on the basis of metabolic activity. Figure 2 illustrates the fluorescence intensity of cellular resorufin, which is produced by reduction of resazurin of viable cells, at day 1, 2, and 3 after cell seeding. gPS cells that were cultured under expansion conditions with LIF showed stronger cell proliferation than gPS cells growing in differentiation medium without LIF (Fig. 2a). gPS cells are viable on gelatine and on the synthetic polymers Resomer LR704 and PVDF, but no material was supporting gPS cell proliferation as good as MEF. gPS cells showed a constant and strong cell proliferation rate with a relative fluorescence intensity up to 65,000 arbr. units on gelatine, 50,900 arbr. units on PLLA and 46,000 arbr. units on Resomer LR704 after 3 days under expansion conditions. Most materials did not even count 35,000 arbr. units and Fibrin, BAK-1095 and PEA C showed the lowest proliferation rates lower than 19400 arbr. units after 3 days (Fig. 2a). Fluorescence intensity data were proved by statistical analysis. gPS cells on other polymers showed significantly less cell proliferation than control cells on MEF (*p≤0.01).
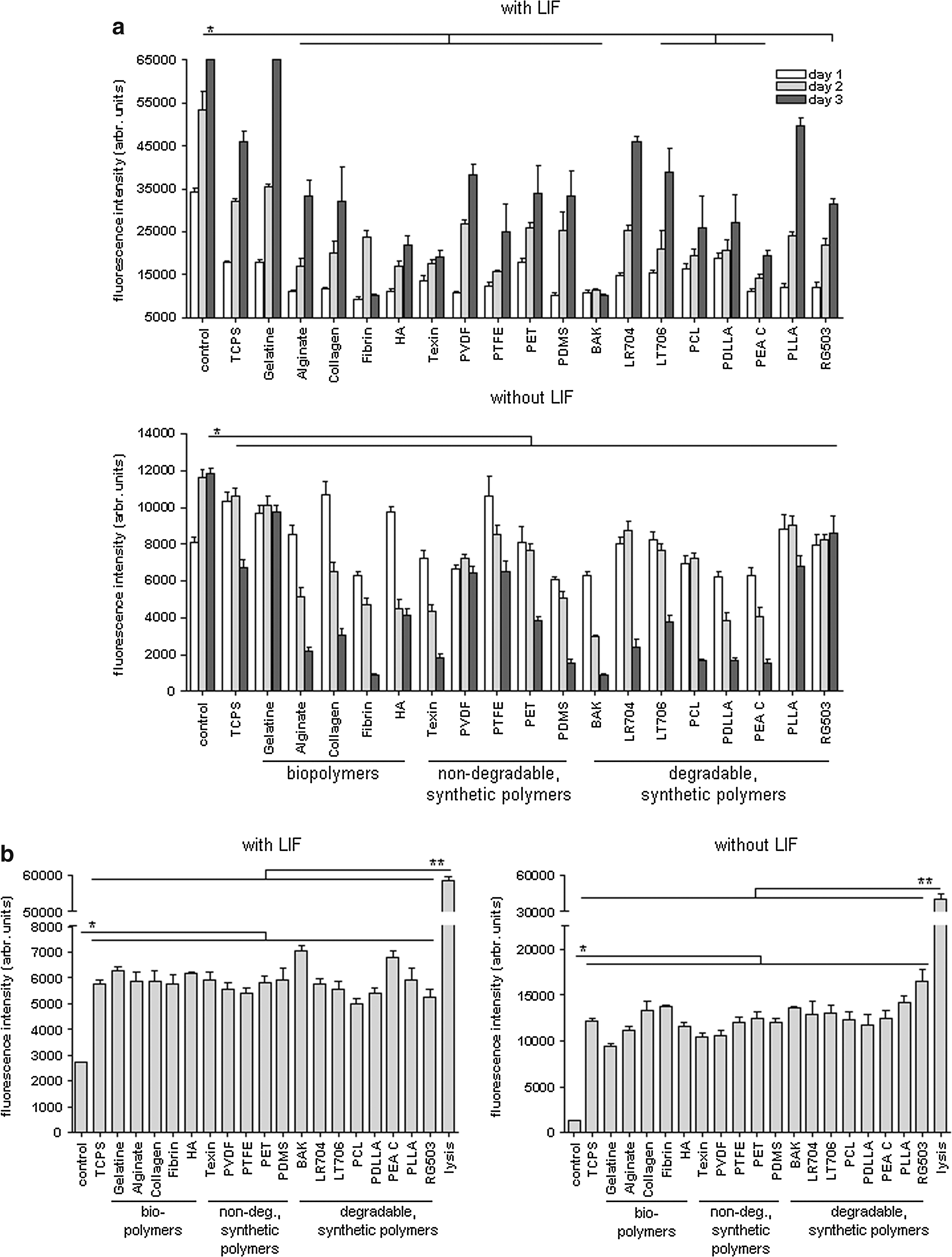
Cytotoxicity of polymers on gPS cells
CytoTox-One Homogenous Membrane Integrity Assay measures release of LDH, an indicator of cytotoxicity. The fluorescence signal of resorufin produced by gPS cells grown on polymers, which is proportional to LDH content, is shown in Figure 2b. The LDH released by gPS cells grown on polymers was significantly lower than the lysis control (**p≤0.001), but 2–3 times or up to 7–12 times higher in gPS cells grown on polymers, respectively, in the presence or absence of LIF as compared to the LDH released by control cells cultured on MEF (*p≤0.01). BAK-1095 and PEA C showed higher cytotoxic effect on undifferentiated gPS cells than other materials (Fig. 2b). Fluorescence intensity of resorufin was lowest on PCL, PTFE, and Resomer RG503 under expansion conditions with LIF and on gelatine, Texin, and PVDF under differentiation conditions without LIF (Fig. 2b).
Quantification of viable and apoptotic gPS cells on polymers
Quadrant analysis of flow cytometry shows exemplarily the distribution of viable (7AAD−, Annexin V−), apoptotic (7AAD−, Annexin V+), and necrotic (7AAD+, Annexin V+) gPS cells on PDMS and Resomer LR704 in comparison to MEF under expansion and differentiation conditions (Fig. 3a). Control gPS cells on MEF showed more than 93% viable cells in undifferentiated state. gPS cells on PDMS are an example of an inappropriate material because of the high amount of apoptotic (more than 45%) and necrotic cells (more than 30%) under expansion conditions with LIF (Fig. 3a). Resomer LR704 showed a high amount of viable cells under both culture conditions. Over 75% of gPS cells on Resomer LR704 were viable, while the percentage of dead cells is around 13%, indicating a normal physiological cell turnover but not enhanced apoptosis. This data suggest Resomer LR704 as a cytocompatible polymer for gPS cells. The heat map depicted in Figure 3b illustrates flow cytometry data, calculated in percent of viable, apoptotic, and necrotic gPS cells on polymers. PDMS showed a higher amount of dead cells under expansion conditions than under differentiation conditions without LIF. On gelatine, tissue culture polystyrene (TCPS), alginate, Texin, PET, Resomer LT706, PLLA, and Resomer RG503, we also found a higher amount of dead cells under expansion conditions after 24 h (Fig. 3b). Most dead gPS cells were detected on Fibrin and PDMS. Resomer LR704 and PVDF are characterized by the highest amount of viable gPS cells that is close to that observed for control gPS cells on MEF. In summary, these results provide the evaluation of compatible polymers for gPS cell cultivation and indicate that gelatine, and Resomer LR704, PTFE, and PVDF represent promising surfaces that support the proliferation and viability of gPS cells.
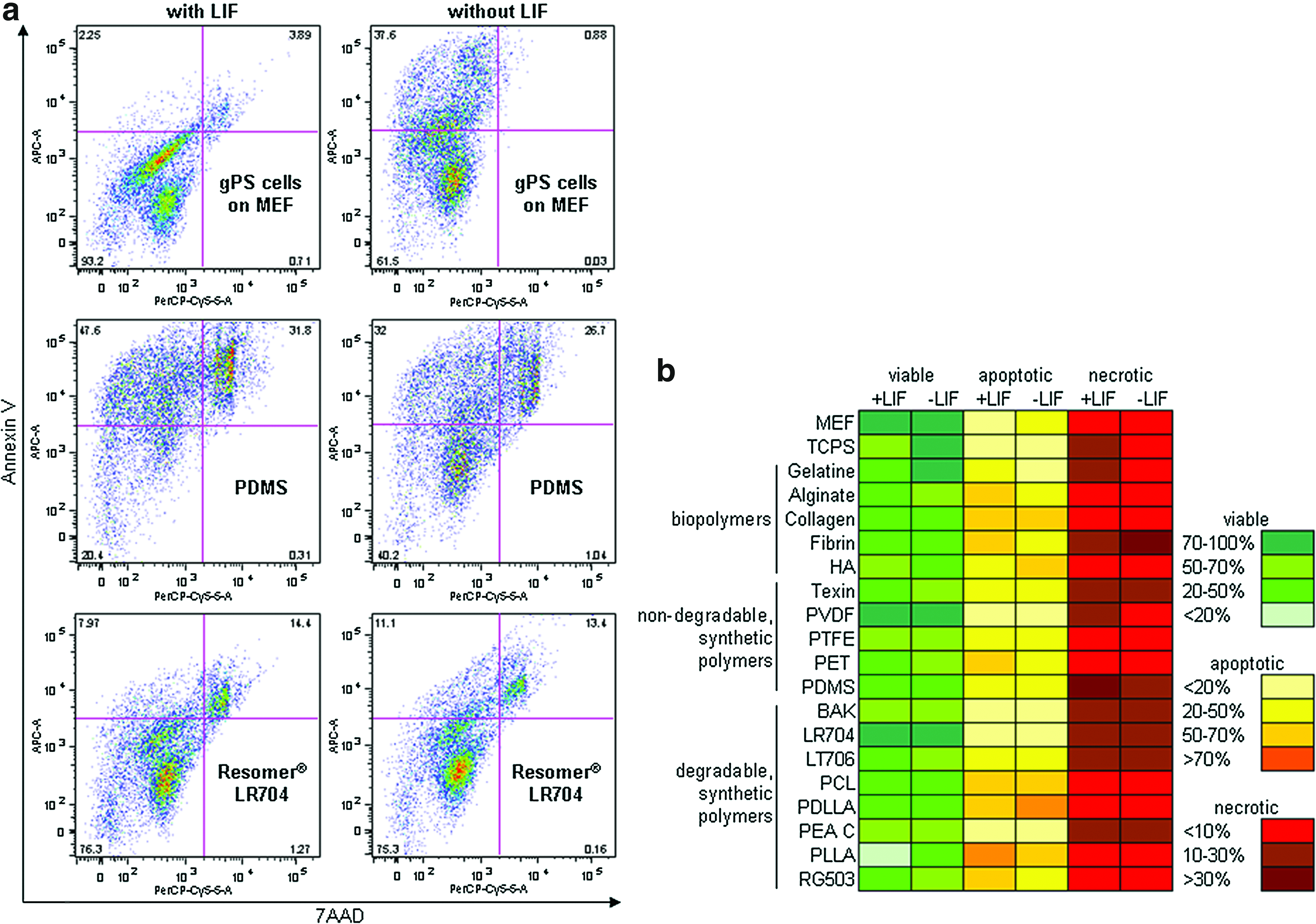
Quantitation of cell viability using flow cytometry.
Biomaterials as feeder substitutes
gPS cells were expanded on biomaterials in LIF-supplemented medium over eight passages to investigate their self-renewal potential on gelatine, Resomer LR704, PTFE, and PVDF. Multiplex assay (see 3.2 and 3.3) and flow cytometry results (see 3.4) pointed these biomaterials because of high amounts of viable gPS cells and low amounts of dead gPS cells as promising surfaces for further experiments. SEM and photographic documentation via light and fluorescence microscopy show gPS cell colony morphology at the end of this expansion period (Fig. 4). gPS cells on gelatine and TCPS formed ES cell-like colonies that were similar in size and shape to control gPS cells on MEF. gPS cell colonies on Resomer LR704, PTFE, and PVDF were much smaller than control colonies on MEF (Fig. 4a).

Morphology of gPS cells on MEF, gelatine, tissue culture polystyrene (TCPS), Resomer LR704, PTFE, and PVDF.
In passage 9, gPS cells were re-transferred from polymers onto MEF to observe if gPS cell colonies still possess their typical morphology and pluripotency. gPS cells on PTFE adhered loosely and did not proliferate, so that expansion was not possible. The sparse gPS cells on PVDF lost the Oct4 promoter-driven GFP-fluorescence, indicating the loss of pluripotency. gPS cells passaged on gelatine and TCPS were still green fluorescent. After re-transfer on MEF, gPS cells maintained their typical ES cell-like morphology and were GFP-positive. gPS cell colonies that were obtained on Resomer LR704 under expansion conditions were GFP-positive and remained so after re-seeding on MEF, indicating the suitability of Resomer LR704 for long-term gPS cell expansion.
Expression of lineage-specific markers in gPS cells cultured on polymers
Gene expression profile of gPS cells were tested by RT-qPCR after 14 days of cultivation under differentiation conditions on gelatine, Resomer LR704, PTFE, and PVDF (Fig. 5a). Oct4 gene expression was significantly downregulated in gPS cells differentiated on these biomaterials as compared to undifferentiated cells (Fig. 5a). The expression of lineage-specific transcripts α- and β-MHC (mesoderm) and α-fetoprotein (endoderm) was strongly upregulated under all these conditions compared to undifferentiated cells. Nestin is an ectodermal marker and its transcripts were not upregulated upon differentiation (Fig. 5a). On Resomer LR704, gPS cells showed the strongest upregulation of cardiac α- and β-MHC transcripts demonstrating their stronger cardiogenic propensity on this material than on gelatine, PTFE, and PVDF.
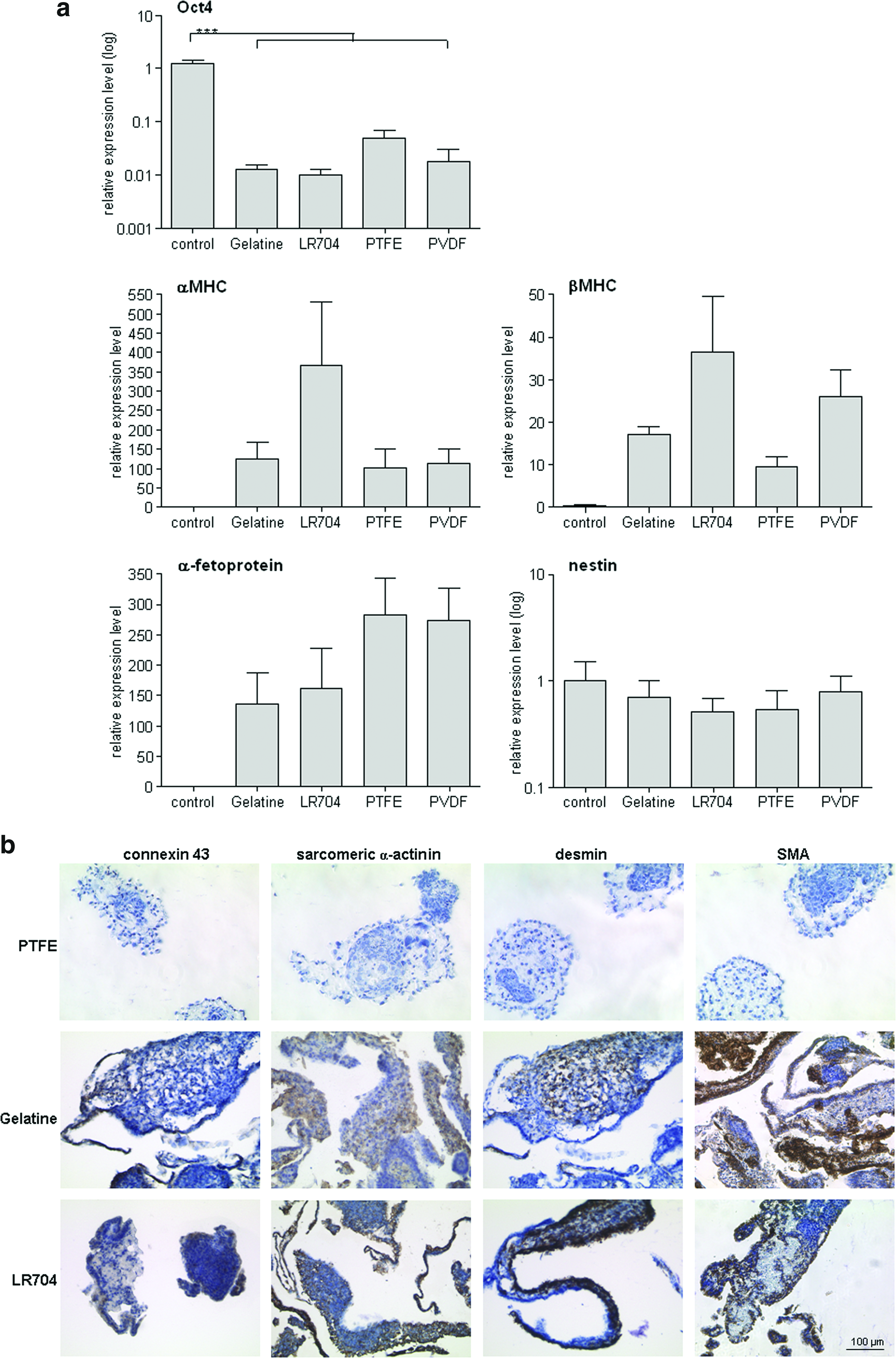
Since no beating areas were found on PTFE (see chapter 3.6, Fig. 6a), immunohistochemistry was performed to check if EBs on this material showed cardiomyogenic differentiation and none of the tested lineage-specific markers were found to be positive in these cells. SMA and desmin were used as mesodermal markers in immunohistochemistry. Strong representation of SMA and desmin was indicative of mesodermal differentiation in EBs cultured on gelatine and Resomer LR704 (Fig. 5b). Positive connexin 43- and sarcomeric α-actinin-stainings of gPS cells on gelatine and Resomer LR704 proved differentiation into cardiomyogenic direction on protein level (Fig. 5b).

Documentation of beating areas.
Cardiomyogenic differentiation of gPS cells cultured on polymers
Suspension culture of gPS cells in bacterial dishes and lack of LIF causes EB formation. Four-day-old single EBs with a diameter of 350–450 μm were seeded on gelatine, Resomer LR704, PTFE, and PVDF in one well of a 24-well plate. Cell morphology and behavior were documented by light microscopy (Fig. 6a). No beating areas could be observed on PTFE after 11 days (Fig. 6a) and even after 21 days (data not shown). gPS cells remained in aggregates and adhered hardly on PTFE as expansion experiments already have shown (see chapter 3.4, Fig. 4b). Only few beating areas were detected on PVDF after 11 days. On gelatine, beating areas were found at day 9 of differentiation and an increasing percentage of beating areas was observed in the following days reaching maximum at around day 16 (Fig. 6a and Supplementary Movies S1 and S2; Supplementary Data are available online at www.liebertpub.com/tea). Rhythmic contractions were detected at the earliest time point (day 8) only in EBs cultured on Resomer LR704 and they increased with the similar kinetics as those observed in EBs on gelatine (Fig. 6a).
Electrophysiological measurements confirmed the cardiomyogenic differentiation of gPS cells on the functional level. Quantity and quality of beating areas on gelatine and Resomer LR704 were evaluated. Since successful cardiomyogenic differentiation of ES cells on gelatine is well established, gelatine was estimated here as control and as standard of comparison for gPS cell contraction. Amplitude, maximal diastolic potential, spontaneous beating frequency, and action potential duration (APD) were quantified (Fig. 6b). Typical cardiac action potentials were observed and general action potential morphology was similar on gelatine and Resomer LR704. However, the statistical analysis showed slight but significant differences of APD90 of gPS cell-derived cardiomyocytes on Resomer LR704 compared to gPS cells on gelatine.
Cardiomyogenic differentiation of gPS cells on molecular level
PCA of differentially expressed genes (with a two-fold change) between differentiated gPS cells on gelatine and Resomer LR704 and undifferentiated gPS cells revealed successful differentiation of gPS cells on gelatine and Resomer LR704 on transcript level, because differentiated gPS cells on both materials clustered together and were distinguished from undifferentiated pluripotent gPS cells (Fig. 7).

Microarray analysis.
Genealogical tree of the heat map of a gene subset of typical pluripotency genes and genes that are expressed in cardiomyocytes also demonstrates clearly the difference between undifferentiated gPS cells and differentiated gPS cells on gelatine and Resomer LR704 (Fig. 7). Characteristic pluripotency-associated genes, such as Oct4 (here as Pou5fl), Sox2, undifferentiated embryonic cell transcription factor 1 (UTF1), and Nanog, were downregulated in gPS cells on gelatine and Resomer LR704, whereas cardiac-specific genes, like cardiac ryanodine receptors (Ryr2), troponin I (Tnni1, Tnni3), troponin T (Tnnt2), and cardiac α-actinin (Actc1), were upregulated. These genes and other cardiac-specific genes showed stronger upregulation in EBs cultivated on Resomer LR704 than on gelatine.
Discussion
Generation of murine gPS cells via specific cultivation procedure and without the introduction of any virally expressed transcription factors, as used, for example, for the generation of iPS cells, has been reported recently.9–11,25,26 These cells have been confirmed to be pluripotent and possess ES cell characteristics as demonstrated by alkaline phosphatase staining, SSEA1 expression, chimera assay, differentiation into somatic cells of the three germ layers in vitro and teratoma formation in vivo. 10 This is a crucial progress in generating patient-specific pluripotent stem cells for medical requirements. The possibility of deriving human gPS cells from adult testis without major ethical concerns compared to human ES cells and the potential for autologous application (in male) characterize them as an interesting tool for regenerative medicine and tissue engineering for myocardial repair. Biomaterials could serve as scaffolds in 2D- and 3D format to promote a niche or simply influence differentiation fate. Findings of the present study indicate that gPS cell-Resomer LR704 combination support cardiomyogenic differentiation forming the basis of those approaches.
Immunofluorescence stainings of undifferentiated gPS cells showed positive signals for the characteristic pluripotency-associated markers, such as Nanog and Sox2. Same cells formed ES cell-like colonies and express Oct4 promoter-driven GFP fluorescence (Fig. 1). Oct4 was described as master gene for pluripotency and is a valid important transcription factor.27,28 Thus, pluripotency of gPS cells was proved by ES cell-like morphology, Oct4-visualization, and positive immunofluorescence staining of Nanog and Sox2 on protein level and confirmed their pluripotent properties described before. 10
To identify capable polymers for expansion and differentiation of gPS cells, viability assay, cytotoxicity test, and flow cytometry for apoptosis quantification were performed. Proliferation of gPS cells varied on polymers although initial cell seeding number was identical. Similar observations were described in proliferation studies of Neuss et al. 13 and Itthichaisri et al. 29 Some materials support proliferation and growth of cells, others have inhibitory effects. 13 According to this, gPS cells proliferated well on Resomer LR704, whereas BAK-1095 showed inhibitory effects. Neuss et al. 13 advised against combination of murine ES cells (R1 cell line) and BAK-1095. The two polyesteramides BAK-1095 and PEA C showed the lowest proliferation rates and highest cytotoxic effects under expansion conditions with LIF (Fig. 2a, b). In contrast, highest proliferation of gPS cells were quantified on TCPS, gelatine, PLLA, and Resomer LR704 (Fig. 2a). Same materials, in addition to PTFE and PVDF, provided appropriate microenvironment for cultivation of gPS cells, resulting in low apoptosis rates and high viability (Fig. 3b). Publications of Itthichaisri et al. 29 and Neuss et al. 13 as well as results of the present study support the theory that the same cell type behaves differently on different materials. Moreover, present results compare cultivation of stem cells on polymers under expansion and differentiation conditions for the first time. That means that not only the material influences the behavior of cells but also cultivation conditions in combination with the same material have a crucial effect. As vitality and cytotoxicity assay and flow cytometry showed, gPS cells behave indeed differently depending on culture conditions on the same biomaterial.
Cultivation of undifferentiated ES cells requires MEF feeders to retain self-renewal potential and remain pluripotency. 30 Such cells would be unsuitable for clinical applications because of contact with potentially immunogenic and infectious animal material. For this reason, it is essential to create a feeder- and serum-free environment for expansion of pluripotent stem cells. These conditions would also be beneficial, because they would allow for a more simple and timesaving cultivation of different types of pluripotent stem cells. Cultivation of ES cells on PVDF has not been reported yet. PTFE, also known as teflon, is a very hydrophobic and inert material that impedes cell adhesion and proliferation. 31 In the present study gPS cells did not proliferate and lost their undifferentiated state on PTFE and PVDF, which excludes these materials as suitable surfaces for gPS cell expansion. In contrast, gelatine and TCPS supported the expansion of pluripotent gPS cells over 9 passages as demonstrated by their ability to form and maintain their ES cell-like colony morphology and the endogenous Oct4 promoter activity. However, it still needs to be determined whether full pluripotency can be maintained after long-term culture on these surfaces, for example, by examination of global gene expression profiles. Although some ES cell lines can be adapted for permanent expansion on gelatine, 32 gPS cells are preferentially cultured on MEF to keep their capacity of continuous proliferation and to stabilize their undifferentiated state.9–11,25,26 Proliferation rate of gPS cells on Resomer LR704 was lower than that of gPS cells on MEF. However, this polymer supported the typical ES cell-like morphology and stable expression of Oct4-GFP fluorescence over passages. Therefore, Resomer LR704 can be regarded as an alternative to feeder-based cultivation of gPS cells. Cultivation of stem cell types, like murine and human mesenchymal stem cells (MSC), human dental pulp stem cells, human preadipocytes, human endothelial progenitor cells (EPC), murine hematopoietic stem cells, and murine ES cells, on Resomer LR704 has been reported by Neuss et al. 13 and cytocompatibility of constituents of this co-polymer, namely, PLLA and PDLLA, has been described for several cell types, like human hepatocytes and MSCs.33,34
Differentiation of gPS cells into cardiomyocytes has been demonstrated by detection of expression of cardio-specific markers at the transcript and protein level in EBs cultured on gelatine and Resomer LR704. At the same time, the expression of the pluripotency gene Oct4 was strongly down regulated upon differentiation in EBs on all tested materials, as described in the literature. 11
Mohr et al. reported that EB size regulates cardiac differentiation of human ES cells. 35 To rule out different initiation of contracting EBs because of variable body size or EB interaction, only one EB with the same size (350–450 μm diameters) was seeded per well in a 24-well plate in the present study. EBs could not adhere and spread on PTFE and did not exhibit enhanced cardiomyogenic differentiation on this material. Furthermore, gene expression profile showed lower expression of α- and β-MHC in EBs on PTFE (Fig. 5a). In contrast, expression of these two cardiac genes that are essential for structure and functionality of cardiomyocytes was higher in gPS cells on Resomer LR704 (Fig. 5a). This polymer also seemed to have accelerated cardiogenic differentiation of gPS cells because, among all tested polymers, earliest beating areas were observed on Resomer LR704. Ko et al. described first contractions of gPS cells after 12 days 10 and EBs cultured on gelatine in this study showed spontaneous beating activity on day 9 of differentiation. In addition, detection of cardiac proteins, such as SMA, desmin, connexin 43, and sarcomeric α-actinin, attested differentiation of gPS cells into cardiomyocytes on gelatine and Resomer LR704 (Fig. 5b). Therefore, Resomer LR704 seems to support cardiomyogenic differentiation of gPS cells at similar or better efficiency as gelatine.
Cardiomyocytes that are generated from ES cells or gPS cells in vitro should possess cardiac properties on molecular, structural, and functional level. Functionality of gPS cell-derived cardiomyocytes on Resomer LR704 has been documented by rhythmically beating areas and was assessed by sharp electrodes technique (Fig. 6a, b). Amplitude, maximal diastolic potential, frequency, and APD50/APD90 were similar under both differentiation conditions, pointing to a comparable subtype differentiation in general. However, beating areas of gPS cells on Resomer LR704 showed slightly shorter APDs than on gelatine, indicating a more mature cardiomyogenic differentiation level. 36 gPS cell-derived cardiomyocytes resemble functionally their ES cell-derived counterparts as reported before. 37
Microarray analysis (Fig. 7) illustrates successful differentiation of gPS cells by the clear shift from undifferentiated gPS cells. Upregulation of typical cardiac genes showed robust cardiac differentiation. Ryr2 are calcium channels located in the sarcoplasmatic reticulum membrane that are expressed strongly by ES cell-derived cardiomyocytes 20 and were found to be highly expressed in gPS cell-derived EBs. In addition, cardiac troponines, like Tnnt2, Tnni1, and Tnni3, are essential mediators of muscle contractions 38 and they were upregulated in gPS cells upon differentiation on gelatine and especially on Resomer LR704.
Expression of lineage-specific markers in gPS cells cultured on polymers and cardiomyogenic differentiation of gPS cells proved on RNA and protein level on Resomer LR704 as well as by functionality are meaningful results with regard to stem cell-related tissue engineering in particular for myocardial applications.
Conclusion
gPS cells are generated from adult testis and their isolation is not related to ethical concern. Polymers can influence both, stem cell growth or differentiation. Resomer LR704 is a suitable polymer for gPS cell cultivation supporting cell adhesion and viability. Expansion of gPS cells on Resomer LR704 maintains their undifferentiated state and additionally their broad differentiation potential. Here we demonstrate that on defined conditions Resomer LR704 supports cardiomyogenic differentiation of gPS cells to a similar or better extent than gelatine as assessed by marker expression on RNA and protein level as well as by functionality.
Thus, Resomer LR704 should be further examined as a promising polymer for derivation of therapeutically relevant cells, especially cardiomyocytes.
Footnotes
Acknowledgments
Supported by a grant from the Interdisciplinary Centre for Clinical Research within the Faculty of Medicine at the RWTH Aachen University (IZKF 520813), by the grant from the Stem Cell Network North-Rhine Westphalia to S.N. and T.Š. and by the German Ministry for Education and Research to T.Š. (grant number 01GN0937), and by the National Research Foundation of Korea Grant funded by the Korean Government (MEST) (NRF-2012-0005133). We thank Qiong Lin, M.Sc., Institute for Biomedical Engineering, Department of Cell Biology, RWTH Aachen, for PCA and hierarchical cluster analysis (heat maps), and Nadin Lange (Institute for Neurophysiology, Cologne) for technical assistance.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.