
Abstract
Topographical cell guidance has been utilized as a tissue-engineering technique to produce aligned cellular orientation in the regeneration of tendon- and ligament-like tissues. Other studies have investigated the effects of dynamic culture to achieve the same end. These works have, however, been limited to two-dimensional cultures, with focus given to the effects from the stimuli independently. The understanding of their combined effects in the tenogenic differentiation of mesenchymal stem cells (MSCs) has also been lacking. This study investigated the synergistic effects of mechanical stimulation on aligned MSCs in a three-dimensional (3D) aligned silk fibroin (SF) hybrid scaffold. Enhanced tenogenesis of seeded MSCs was observed in the scaffold group with aligned SF electrospun fibers (AL) under static culture conditions, as evidenced by the upregulation in expression and production of tendon/ligament-related proteins. The intensity and onset of these differentiative markers were increased and advanced, respectively, under dynamic culture conditions, indicative of an accelerated matrix deposition and remodeling process. Consequently, the tensile properties of dynamically cultured AL were significantly improved. We thus propose that the aligned hybrid SF scaffold facilitates mechanoactivity and tenogenic differentiation of MSCs by intensifying the positive effects of mechanical stimulation in a 3D environment.
Introduction
Tendons, such as the knee patellar tendon, the foot Achilles tendon, the hand flexor digitorum profundus tendons, and ligaments, such as the knee collateral and cruciate ligaments, are frequently injured. 5 In the United States, more than 100,000 patients per year undergo surgery to repair tendon or ligament injuries. 6 Unfortunately, these tissues do not heal spontaneously when torn. This is largely due to the lack of vasculature at the tissue, causing inadequate supply of essential reparative factors or agents should the tissue be torn.7,8 Even if the tissues do heal intrinsically, the initial healing matrix formed will consist of randomly arranged collagen, predominantly the smaller and less organized type III collagen (20%–30% of total collagen), and amorphous ground substances.5,9–12 This marked increase in collagen III from 12% in ligaments and 1%–5% in tendons, results in a decrease in collagen type I to type III ratios from 5.9–12.7 in the ligaments and from greater than 19 in the tendons. 1 With the decrease in the collagen I to III ratio, the tissue undergoes a reparative phase, whereby structural integrity between the torn ends is simply restored. This is in contrast to the regeneration of the tissue, whereby the collagen I to III ratio is similar to that of the native tissues, leading to functional recovery. Although gradual remodeling of this healing matrix will then take place eventually, involving an increase of fibril alignment along the tissue length and increase in the proportion of collagen I over collagen III, the process will take longer than a year. Even though there is gradual alignment of collagen fibrils over time, slight disorganization and abnormalities persist due to environment and mechanical factors, with the tissue never to regain its original properties.13–15
With the emergence of tissue engineering as a promising clinical solution, it is important to consider the healing mechanism of the tendon and ligament when devising strategies for regeneration of the tissues for functional recovery. To avoid formation of randomly arranged collagen fibrils, cellular alignment may be a necessary preceding step as it has been shown to have a direct effect on the alignment of the collagen matrix. 16 Contact guidance, by using cellular responsiveness to seeded surface topography, has often been utilized to achieve cellular alignment. In many of the related studies, cells cultured on two-dimensional (2D) microgrooved surfaces were shown to generally elongate in the direction of the groove.17–19 Other studies have preliminarily demonstrated the positive effects of cell culture on aligned electrospun fibers, at the nanometers to the submicron level, in terms of cell morphology and extracellular matrix (ECM) production.20–22 In a study by Lee et al., 20 it was observed that human ligament fibroblasts cultured on electrospun aligned polyurethane (PU) fibers (657±183 nm) were spindle shaped and oriented in the fiber direction. These aligned cells were also observed to secrete significantly more collagen than randomly oriented cells. The tensile strength of the ligament has also been shown to improve with the orientation of fibroblasts along the length. 21
Mechanical stimulation has also been shown to play a significant role in the healing, remodeling, and strengthening of tendons and ligaments. 5 For maintenance of proper in vivo cellular behavior and matrix remodeling, mechanical loading is as important as cellular alignment for tendon and ligament regeneration leading to functional tissues. Cyclic strain has been shown to enhance the expression of ECM proteins and induce the assembly of ECM structures. 23 It has also been shown that tendon and ligament cells respond to mechanical loads by increasing their level of intracellular calcium.24,25 In other studies, tendon cells have been shown to release adenosine triphosphate (ATP) in response to mechanical stimulation. 26 These effects are often enhanced when complemented with contact guidance. As mechanical interactions between cells and the underlying substrate can take several forms, including topographic interactions and surface tensile forces, aligned electrospun fibers have been shown to sensitize these cells for effective mechanotransduction by tensioning cytoskeletal filaments. 27 As a result of the higher level of aligned matrix production, earlier transmission of forces can be effected to the entire regenerating tissue to attain better mechanical properties of the tissue-engineered construct.
In our previous study, we have investigated the effects of a three-dimensional (3D) aligned full silk fibroin (SF) hybrid scaffold system on seeded mesenchymal stem cells (MSCs) for the purpose of ligament tissue engineering. 28 The scaffold system comprised of knitted SF integrated with aligned SF electrospun fibers (SFEFs). It was demonstrated that the aligned SFEFs provided environmental cues for positive cellular responses, including cellular and ECM alignment, expression and production of ligament-related proteins, and enhanced tensile properties. The aligned fibrous scaffold was shown to potentiate differentiation of seeded MSCs toward the ligament fibroblast lineage. Nevertheless, the synergistic effect of mechanical conditioning on the 3D aligned scaffold was not studied. The understanding of the combined effects of these culture conditions in the tenogenic differentiation of MSCs has also been lacking.
The purpose of this study was thus to investigate the biophysical influence of topographical culture environment and mechanical stimuli on tenogenesis of MSCs using the 3D aligned SF hybrid scaffold. This was in contrast to the studies that had investigated the synergy of culture topography and mechanical stimulation primarily in a 2D environment using tendon and ligament fibroblasts.20,29 We hypothesized that tenogenesis of MSCs in terms of the expression of tendon/ligament-related differentiation markers, distinctive protein deposition, histological effect, and the functional biomechanical outcome would be enhanced and accelerated under the synergistic effect of 3D mechanical conditioning and aligned contact guidance.
Materials and Methods
Fabrication of hybrid SF scaffolds
Raw silk knits of dimensions about 40×20 mm (the static culture group) and 60×20 mm (the dynamic culture group) were each fabricated from 3 yarns of raw Bombyx mori silk fibers (80 fibroins/yarn; Silk Innovation Center) using a knitting machine (Silver Reed SK270). Longer knits were used for the dynamic culture group to cater for attachment in the bioreactor chamber.
Both groups of raw silk knits and the silk fibers used for making the electrospinnable SF solution were degummed using the protocol optimized for SF structural and mechanical preservation. 30 Briefly, the process involved mechanical agitation of raw silk knits in the degumming solution consisting of aqueous Na2CO3 and sodium dodecyl sulfate (SDS), 0.25% w/v each (Sigma-Aldrich) at 100°C for 30 min. The degummed silk fibers or SF knits were then rinsed in warmed distilled water and left to air dry for at least 24 h before further processes were conducted.
As previously reported, 28 the electrospinnable SF solution was obtained by dissolving dried SF in saturated lithiumthiocyanate (LiSCN, 250% w/v; Sigma-Aldrich) to attain a final concentration of 20% w/v. After residue removal by centrifugation at 3000 rpm for 10 min, the supernatant was dialyzed against distilled water for 3 days using Snakeskin™ pleated dialysis tubing (10,000 MWCO; Thermo Fisher Scientific, Inc.), which was then lyophilized to obtain regenerated SF sponge. The electrospinnable SF solution (9.5% w/v) was obtained by a final dissolution of the regenerated SF sponge in hexafluoro-2-propanol (HFIP, Fluka, Sigma-Aldrich). Two types of hybrid scaffolds with different SFEF arrangements were fabricated. Aligned hybrid scaffolds (AL) were composed of highly aligned SFEF (AL-SFEF) produced using a customized rotational electrospin setup. Random hybrid scaffolds (RD) consisted of randomly arranged SFEF (RD-SFEF) collected from a grounded metallic collector. Both groups of SFEF meshes were collected onto 44×22-mm glass slides and amassed to a comparable mass of ∼12 mg before integrating with the degummed SF knits. It should be noted that the AL-SFEF meshes were integrated with the aligned direction along the length of the SF knits. Integration involved careful peeling of the SFEF meshes from the glass slides and laying them flatly on both sides of the SF knit, before methanol treatment for SFEF crystallization, insolubilization, and binding to the knitted SF was conducted.31–33 The morphologies of the completed SF hybrid scaffolds were observed using phase-contrast microscopy (IX71 Inverted Research Microscope; Olympus). The integrated hybrid SF scaffolds were sterilized by means of formaldehyde (37%) (Mallinckrodt Baker, Inc.) gassing for 24 h prior cell seeding. All other sterile equipment was treated by autoclaving.
Seeding of MSCs onto the hybrid SF scaffolds
MSCs were derived from bone marrow aspirates of New Zealand White rabbits and cultured based on a protocol approved by the Institutional Animal Care and Use Committee, National University of Singapore, using the techniques as reported. 28 Characterization studies have been performed prior and published showing that the derived MSCs retained their multilineage potential. 34 1.5×106 MSCs (P3) were then seeded by simply pipetting onto one side of each sterile hybrid scaffold. All hybrid scaffolds (RD and AL), of both the static and dynamic culture groups, were initially cultured separately in complete culture medium containing the Dulbecco's modified Eagle's medium with high glucose (Gibco, Invitrogen) supplemented with 10% fetal bovine serum (HyClone Logan), L-glutamine (580 mg/L), and penicillin–streptomycin (100 U/mL) in a laminar manner (Fig. 1A) for 3 days before being rolled up with the cell-seeded surface in the inner core (Fig. 1B).

Gross observation of the mesenchymal stem cell (MSC)-seeded hybrid SF scaffold statically cultured in a laminar manner for 3 days
Static and dynamic culture of the hybrid SF scaffolds
For the statically cultured group, rolled-up forms of the AL and RD scaffolds then continued to be cultured separately in customized six-well polycarbonate dishes for another 11 days (total experimental period of 14 days), with the medium being changed twice a week.
For the group undergoing dynamic culture, rolled-up scaffolds (RD and AL) were loaded into steam-autoclaved standalone bioreactor chamber vessels with the ends (10 mm at each end) of the hybrid scaffolds used for anchorage. The middle 40 mm of these scaffolds were seeded, which eventually formed the exposed portion between the clamps after loading in the bioreactor (Fig. 1C). The chamber vessels were then affixed onto the bioreactor main bioreactor vessel stand (Fig. 1D), where dynamic culture conditions would be provided over the next 11 days period. Dedicated medium circulation for each chamber was provided by the environmental control system, which regulated the medium temperature (37°C), pH (7.4) and dissolved the oxygen level (30% saturation level) to ensure that optimal culture conditions were provided to the cultured constructs. Mechanical stimulations of scaffolds were achieved via movements of the center rods of the chamber vessels, which were attached to the motors at the bioreactor vessel stand, providing both a 5% translational strain and a 90° rotational strain to mimic the multidimensional straining environment in vivo. The dynamic culture conditions followed a 12-h daily stimulation regime, whereby the cyclic frequency was set at 0.1 Hz amounting to 4320 cycles per day based on the 12-h daily activity rate.
Based on the different scaffold types and the varied culture conditions provided, four groups of constructs, including RD (static), RD (dynamic), AL (static), and AL (dynamic), would be compared for cell viability, proliferation, collagen deposition, tendon/ligament-related gene expression and protein synthesis, histological profile, and mechanical properties.
Cell viability and proliferation
The different groups of constructs were assayed for cell viability and proliferation using Alamar Blue™ assay following vendor's instructions (BioSource International) at different time points (3, 7, and 14 days postseeding). Briefly, the harvested constructs (n=5) were incubated for 3 h in the complete culture medium supplemented with 10% (v/v) Alamar Blue fluorescent dye. About 200 μL of the medium was then extracted from each sample and measured at 570/600 nm in a microplate reader. As a negative control was necessary for this assay, the cell-free culture medium supplemented with 10% Alamar Blue was used.
Collagen quantification
Collagen production and deposition of the constructs were quantified using a picrosirius red-based colorimetric assay (SirCol™ collagen dye binding assay kit, Biocolor Ltd.) following the vendor's protocol. Briefly, at the various time points (3, 7, and 14 days), the harvested constructs from the different groups were finely cut and digested with 500 μL of the pepsin solution (0.25 mg/mL) under constant mechanical shaking at room temperature (25°C) for 2 h. About 200 μL of the digested solution was then extracted from each sample, and into which, 1 mL of the dye reagent was added and mixed for another 30 min in room temperature. After centrifugation at >10,000 g for 10 min, the dyed collagen was precipitated and redissolved in 1 mL of the releasing reagent. The absorbance of the redissolved dye was measured at 540 nm and the collagen amount was derived by extrapolation from the standard curve.
Histological assessment
Seeded constructs from the respective groups were harvested (n=3 each) for hematoxylin and eosin staining after 7 and 14 days of culture. The specimens were fixed in 10% neutral-buffered formalin, paraffin blocked, and sectioned longitudinally (along the lengthwise axis of the cylindrical scaffolds) at the core as it was of interest to examine the core region for cell morphology and continued viability over the 14-day experimentation. Dehydration of the slides was performed before mounting on glass cover slips.
Real-time quantitative reverse transcriptase–polymerase chain reaction analysis
To assess the extent of tenogenic differentiation of seeded MSCs in the various groups, gene expressions for tendon/ligament-related ECM proteins, such as collagen type I, collagen type III, tenascin-C, and tenomodulin, were analyzed and evaluated. Gene expressions of bone-related proteins, such as osteonectin and osteopontin, were also evaluated to ascertain the level of osteogenic differentiation and for comparison with that of the tendon/ligament-related proteins. After 7 and 14 days of culture, total RNA was extracted from the respective constructs (n=3) using the RNeasy Mini Kit® (Qiagen) according to the vendor's protocol. The RNA concentration was determined via nanodrop (NanoDrop Technologies) and 200 ng of RNA was used for synthesis of cDNA with the Iscript cDNA synthesis kit (Biorad Laboratories). Quantitative reverse transcriptase-mediated polymerase chain reaction (qRT-PCR) was performed using the QuantiTect SYBR-Green PCR kit (Qiagen) to quantify the transcription level of the target markers, with glyceraldehyde 3-phosphate dehydrogenase (GAPDH) as the reference gene. The primer sequences used (Table 1) were obtained from the published literature35–38 and were synthesized by Aitbiotech Pte Ltd. cDNA (1 μL) from each sample was mixed with 10.0 mL of QuantiTect SYBR Green PCR master mix, 0.25 mL of each primer, and 8.50 mL of RNase-free water. Quantitative real-time PCRs were performed and monitored by the Stratagene Mx3000P system. Reaction was done at 95°C for 15 min and amplification was done for 40 cycles, which included a denaturation step at 95°C for 15 s and an extension step at 60°C for 1 min. Duplicate amplifications were performed and the transcription level of the target genes were normalized to GAPDH before analysis using the 2ΔCt formula with reference to undifferentiated MSCs (P3).
Collagen I (α2), Collagen III (α1), and GAPDH sequences obtained from Ref. 34
Tenascin-C sequences obtained from Ref. 35
Tenomodulin sequences obtained from Ref. 36
Osteonectin and osteopontin sequences obtained from Ref. 37
GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
Western blot analysis
After 7 and 14 days of static or dynamic culture, the constructs were digested with pepsin (200 mg/mL in 0.5 N acetic acid; Sigma-Aldrich) for 72 h at 4°C for total protein extraction. Pepsin action was stopped after 10 N NaOH was added in. Upon concentrating the extracted protein via the Microcon 30 centrifugal filter (30,000 Mw cutoff; Millipore Co.), each sample was mixed with the laemmli buffer and 50 mM DTT solution, loaded to a 3%–8% SDS-PAGE, and blotted onto nitrocellulose membranes. Western blot was performed accordingly following the vendor's protocol (Zymed Laboratories, Invitrogen). Briefly, the membranes were first blocked in the blocking buffer for 1 h and incubated at 4°C overnight with the respective diluted (1:500) primary monoclonal antibodies. The specific primary antibodies used were the mouse anti-type I collagen monoclonal antibody (Sigma-Aldrich), the mouse anti-type III collagen monoclonal antibody (Millipore), and the mouse anti-tenascin-C monoclonal antibody (Millipore). Upon washing the membranes with the washing buffer five times, incubation with secondary antibodies diluted to 1:200 in the blocking buffer was performed for 30 min. The membranes were washed and incubated with the enhanced chemiluminescence (ECL) working solution for 5 min. Band signals were detected and relative band intensities (with respect to statically cultured RD at day 7) were obtained and compared among the groups.
Biomechanical test on cultured hybrid scaffolds
The constructs at different time points (7 and 14 days postseeding) were tensile tested to failure using a universal testing machine (Instron 3345 Tester; Instron) at standard environmental conditions (20°C, 60% relative humidity).
For each group, cylindrical specimens (5-mm outer diameter, 20-mm gauge length, n=5) as rolled from laminar form of the hybrid scaffolds along the widths were loaded axially. The specimens were kept moist with the phosphate-buffered saline solution and loaded to failure at a crosshead speed of 10 mm/min without any pretension or preconditioning. The failure loads, linear stiffnesses, and extensions at failure loads were determined from the load–displacement curves. The toe region extent, in relation to displacement, was measured to determine the extent each construct can be stretched before linear extension occurred.
Statistical analysis
To assess statistical significance of multiple comparisons, single-factor analysis of variance technique and post hoc Tukey tests were used. For pairwise comparisons, two-tailed, unpaired Student's t-tests were used. GraphPad Prism ver. 5 (GraphPad Software, Inc.) was used to implement the analysis. All data are expressed as means±standard deviation and p<0.05 was considered statistically significant.
Results
Hybrid SF scaffold morphology
As we have reported earlier, 28 hybrid SF scaffolds produced using the methods described were highly porous (Fig. 2A, B). The pores were uniformly distributed and were interconnected, with pore sizes ranging from 1–60 μm in the equiaxed pores of RD and 1–20 μm (minor axis) and 10–100 μm (major axis) for the elliptical pores of AL. There was also firm binding between the SFEF meshes and the knitted SFs, resulting from the two-step integration process that utilizes the contractile forces from methanol-treated SFEF.

Phase-contrast images illustrating scaffold morphology of hybrid SF scaffolds: RD
Cell viability and proliferation
Cell viability was shown to be significantly higher in the AL group compared to the RD group for both culture conditions (static and dynamic) after 7 days of culture (Fig. 3). For the groups cultured in the dynamic conditions, AL hybrid scaffolds had 23% (day 7) and 27% (day 14) more viable cells when compared to RD hybrid scaffolds (p<0.01). Within each type of hybrid scaffold, a significant difference was only seen on day 14 between AL hybrid scaffolds cultured in static and dynamic conditions (p<0.05), while no significant difference was observed in the RD hybrid scaffolds of the two culture conditions during this 14-day period (p>0.05).
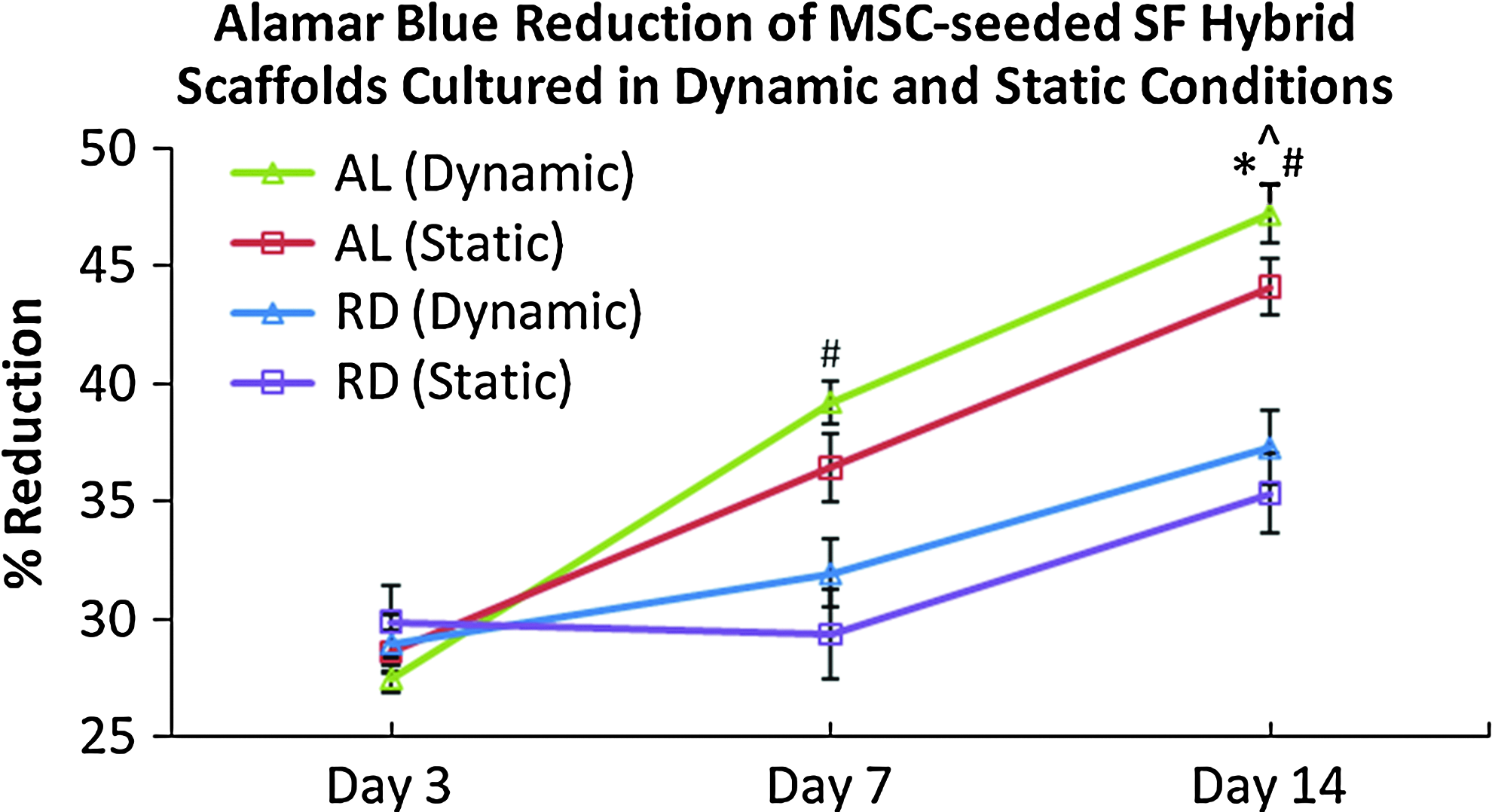
Alamar Blue™ assay illustrating consistent and significantly more viable cells in the AL groups (both static and dynamic) compared to the RD group of similar culture condition from day 7 onward (#p<0.01, the Student's t-test, n=5) and AL (dynamic) having more viable cells than AL (static) on day 14 (*p<0.05, the Student's t-test, n=5). Significant proliferation (^p<0.05, analysis of variance and post hoc Tukey tests, n=5) was observed in AL (dynamic), AL (static), and RD (dynamic) through the 14-day culture. Color images available online at www.liebertpub.com/tea
From Figure 3, it was also clear that there was consistent proliferation in both the AL groups (static and dynamic) and also the dynamically cultured RD group through the 14-day culture period (p<0.05). However, there was no significant proliferation in the statically cultured RD group (p>0.05) within the same 14-day culture period.
Collagen synthesis
To give an indication on the extent of ECM formation, insoluble collagen assays were performed to determine the amount of deposited collagen in the various cultured constructs (Fig. 4). As was previously reported, 28 statically cultured AL had 33.5% more collagen deposited than statically cultured RD at day 14 (p<0.01). With dynamic conditioning, significant differences in collagen deposition was found in the AL groups only after 14 days of culture, whereby dynamically cultured AL hybrid scaffolds had 24.9% more collagen deposited than the statically cultured counterpart (p<0.01). No significant difference was found for the RD groups cultured using the two different conditions through the 14 days (p>0.05). Between the two construct types that were cultured dynamically, AL constructs demonstrated significantly more collagen deposition (41.3%) by day 14 as compared to the RD type (p<0.01).
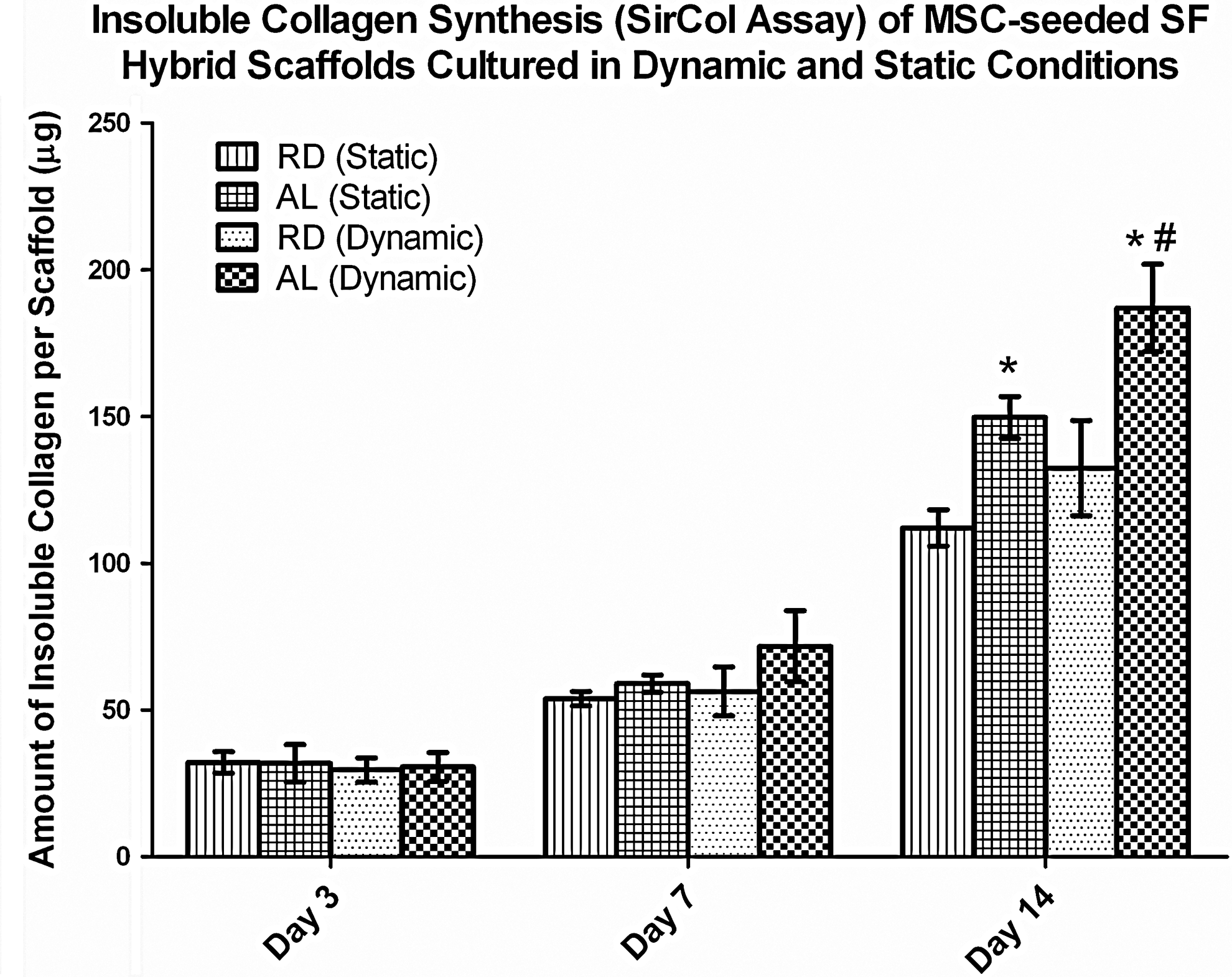
SirCol™ assay for amount of collagen deposited per cultured construct. Significant increase in collagen deposition was observed in AL relative to RD under each culture condition at day 14 (*p<0.01, the Student's t-test, n=3). Significantly more collagen was deposited in the AL (dynamic) group as compared to the AL (static) group at day 14 (#p<0.01, the Student's t-test, n=3).
Histological analysis
Increased cell density and distribution were observed at the core of statically cultured AL hybrid scaffolds (Fig. 5E, F), while no observable increase in cellular distribution was found at the core of statically cultured RD (Fig. 5A, B). AL scaffolds were thus shown to support uniform proliferation in the core during static culture, while for statically cultured RD, the individual cell colonies were not interconnected. However, with dynamic culture, more uniform cell distribution was observed for RD (Fig. 5C, D), especially at day 14. This could be attributed to the improved medium uptake into the scaffold cores with dynamic culture.
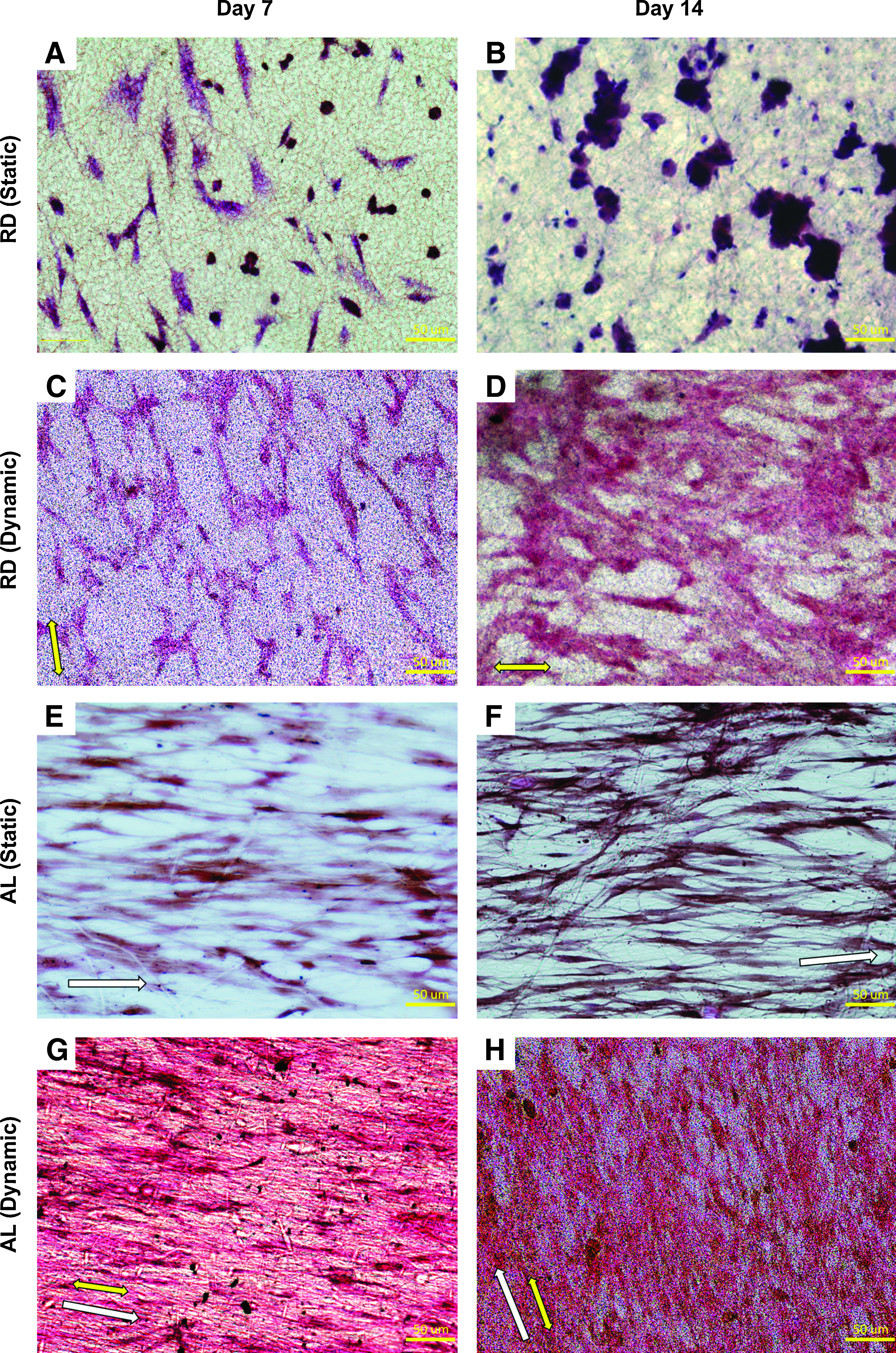
Histological evaluation of MSC-seeded statically cultured RD
The effect of dynamic conditioning on cellular morphology and ECM deposition was apparent in both the RD and AL constructs. Compared to the statically cultured counterpart, dynamically cultured RD exhibited improved cellular elongation and ECM deposition along the direction of the mechanical strain. Nevertheless, the effect was more prominent in AL. Increased cellular density and ECM deposition were also observed qualitatively in the dynamically stimulated AL group (Fig. 5G, H) as compared to both the statically cultured AL group (Fig. 5E, F) and the dynamically cultured RD group (Fig. 5C, D), demonstrating the synergistic effect of the mechanoactive AL hybrid scaffold with mechanical cues from dynamic culture in accelerating construct growth.
Gene expression of tendon/ligament-related ECM proteins using real-time qRT-PCR
The expression of tendon/ligament-related and osteogenic genes in RD (static and dynamic cultures) and AL (static and dynamic cultures) were evaluated via real-time qRT-PCR (Fig. 6). It was observed that gene expression for all targeted tendon/ligament-related genes were significantly upregulated in both dynamically cultured groups (RD and AL) by day 14, as compared to their respective statically cultured counterparts at day 14 (p<0.05). Nevertheless, significantly higher expression for collagen I, tenascin-C, and tenomodulin was observed in the dynamically cultured AL group as early as day 7, compared to the static equivalent. This was, however, not the case for the RD groups, as there was no significant difference in tendon/ligament-related gene expression between statically and dynamically cultured RD groups until day 14. In terms of osteogenic gene expression, both osteonectin and osteopontin were generally downregulated in constructs with mechanical stimulation, with significant downregulation of osteopontin observed in dynamically cultured AL compared to statically cultured AL at day 14 (p<0.05). With dynamic stimulation, all targeted tendon/ligament-related genes were upregulated in the AL group as compared to the RD groups (static and dynamic cultures) by day 14, indicating that dynamic conditioning did synergistically complement the favorable topographical cues presented by the AL hybrid scaffolds in accelerating tenogenic differentiation.

Gene expression for MSCs, statically and dynamically cultured in RD and AL for 7 and 14 days.
Western blot analysis
Protein expressions for collagen I, collagen III, and tenascin-C were detected for dynamically and statically cultured RD and AL groups after 7 and 14 days. Analysis was performed on densitometric data obtained from the optical intensity of each lane expressed in the western blot membranes, which was normalized to statically cultured RD at day 7. It was clear that the matrix of all the cultured constructs composed of collagen I predominantly as its expression was consistently higher across the various groups when compared to collagen III and tenascin-C (Fig. 7). Significant increase over the experimental duration (from day 7 to 14) was found for all of the three proteins tested in the dynamically cultured groups, for both RD (collagen I: 80.7% more, collagen III: 33.1% more, tenascin-C: 50.5% more; p<0.05) and AL (collagen I: 81.5% more, collagen III: 64.1% more, tenascin-C: 70.3% more; p<0.05). Between the RD and AL groups that were dynamically cultured, AL hybrid scaffolds could stimulate increased type I collagen production since day 7 compared to the RD types (day 7: 37.7% more, day 14: 38.3% more; p<0.05). Similar to static culture, significantly more type III collagen and tenascin-C were found for the dynamically cultured AL group relative to the dynamically cultured RD group on day 14 only (collagen III: 40.7% more and tenascin-C: 43.4% more; p<0.05). With mechanical conditioning, both the RD and AL constructs composed of significantly more collagen I from day 7 onward (For RD, day 7: 90.9% more, day 14: 26.7% more, and for AL, day 7: 60.0% more, day 14: 28.7% more; p<0.05), and significantly more collagen III and tenascin-C on day 14 only (For RD, collagen III: 45.0% more and tenascin-C: 73.4% more, and for AL, collagen III: 24.7% more and tenascin-C: 19.7% more; p<0.05). These results were indicative of the differentiative stimulatory effects of mechanical stimulation on the hybrid scaffolds, especially the aligned hybrid SF scaffolds.
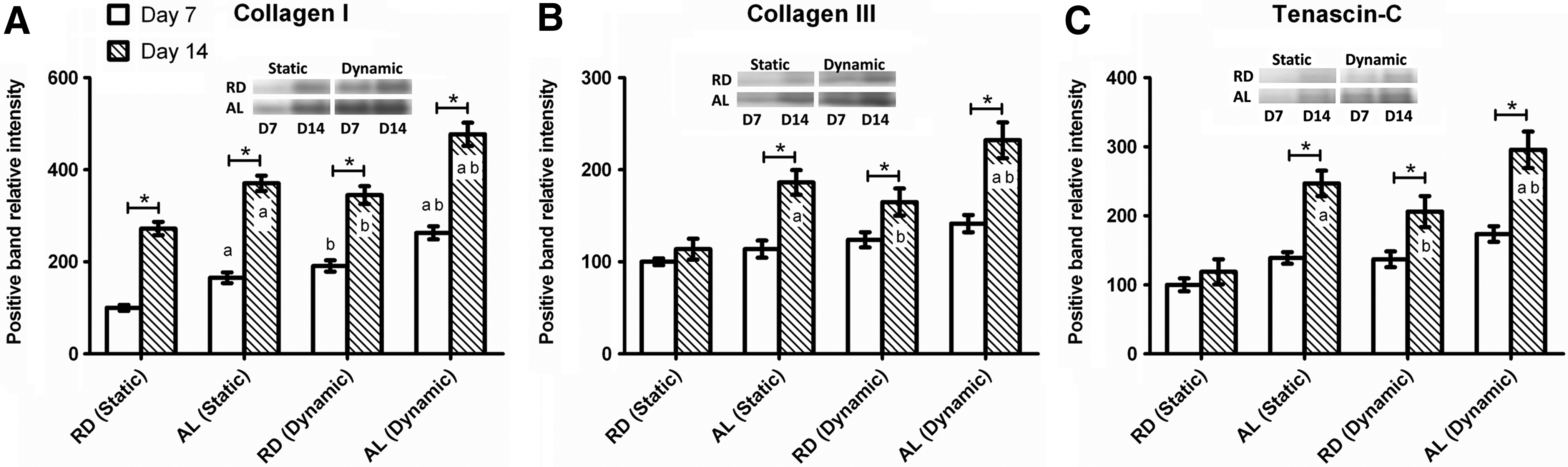
Western blot analysis of tendon/ligament-related extracellular matrix proteins,
Tensile properties of cultured hybrid scaffolds
Statically and dynamically cultured rolled-up constructs (RD and AL at day 7 and 14) were tested for their tensile properties as tabulated in Table 2. The samples were tested to failure, which was initiated from the central region of the entire gauge length. Typical regions of the load–displacement curves for native tendons and ligaments, which included the toe region, linear region, microfailure region, and failure region, were similarly observed in the various cultured constructs (Fig. 8). The microfailures observed were largely due to the failure of the knitted SF microfibers. With mechanical stimulation, both RD and AL groups at day 14 had increased the maximum load and stiffness than that at day 7 (For RD, maximum load: 13.3% increase, stiffness: 32.0% increase and for AL, maximum load: 19.1% increase, stiffness: 10.7% increase; p<0.05). This was not the case for RD when it was subjected to static culture, as there was no significant increase in the maximum load and stiffness from day 7 to 14 (p>0.05). Similar to the statically cultured groups, dynamically cultured AL were also significantly stronger and stiffer than the dynamically cultured RD at both day 7 and 14 (p<0.05). When compared to the statically cultured groups, dynamically cultured AL was significantly stronger than the statically cultured AL by day 14 (8.9% stronger, p<0.05). This was not the case when dynamically and statically cultured RD constructs were compared within this experimental duration. This indicated that the dynamic conditioning did have an effect in strengthening the AL constructs, but had limited effects in the RD type. Nevertheless, no significant increase was found for stiffness between the two culture conditions for AL (p>0.05). This could be attributed to the freshly deposited collagen being not as stiff and required longer culture period for maturation and remodeling. There was no significant difference in the extent of toe regions measured between the groups cultured dynamically and when cross comparisons were made with the statically cultured counterparts (p>0.05). A longer culture period might be necessary before significant changes in the extent of the toe region could be observed.
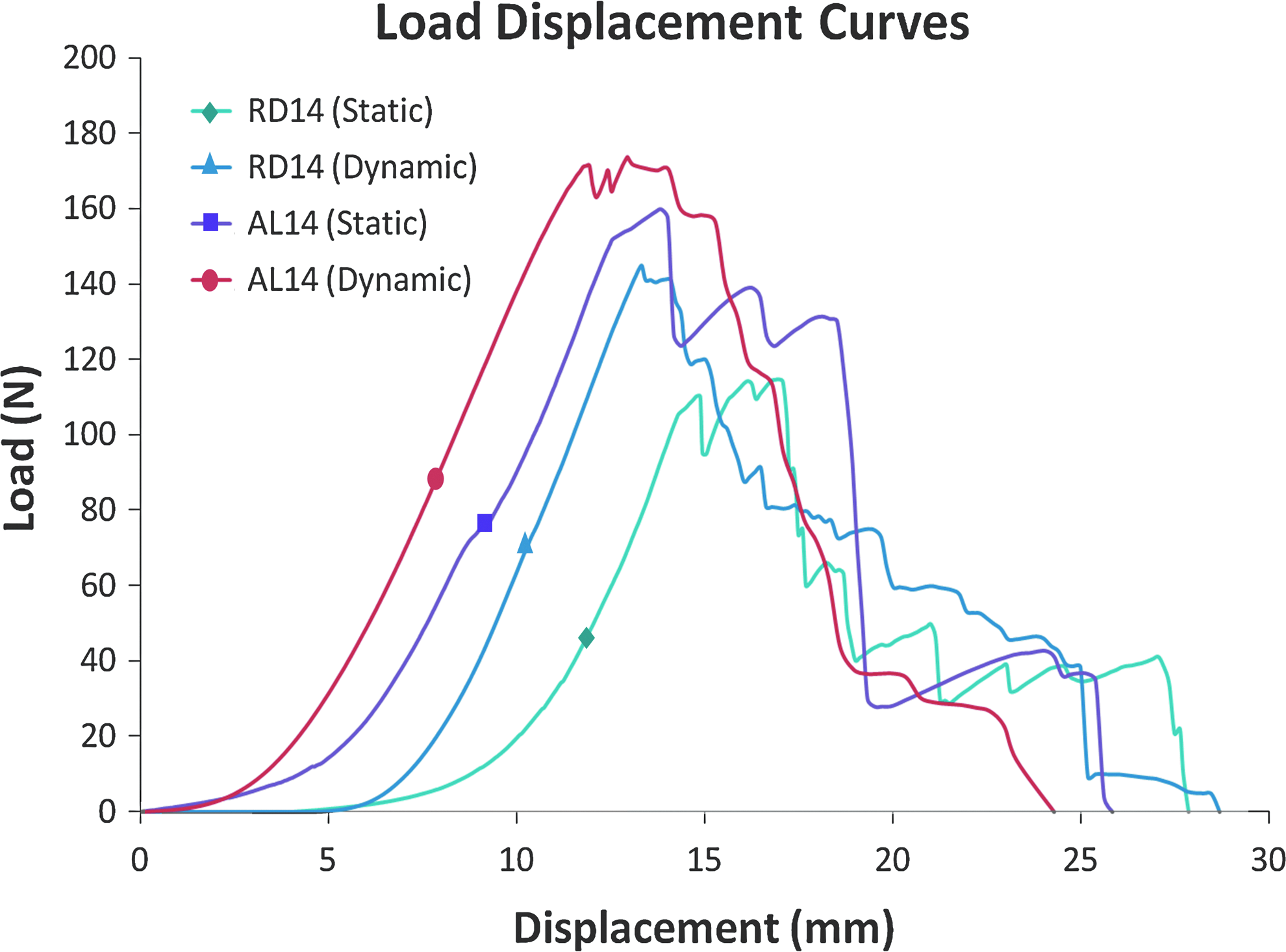
Representative load–displacement curves for statically and dynamically cultured RD and AL constructs at day 14. Color images available online at www.liebertpub.com/tea
n=5, data: mean±standard deviation.
p<0.05 when compared to cultured RD constructs at each time point of the same culture condition (for static and dynamic cultures, respectively).
p<0.05 when dynamically cultured constructs were compared to the statically cultured equivalent of the same time point.
AL, aligned hybrid scaffolds; RD, random hybrid scaffolds.
Discussion
This work is focused on the biophysical influence of topographical culture, environment, and mechanical stimuli on tenogenesis of MSCs in relation to the expression of tendon/ligament-related differentiation markers, distinctive protein deposition, histological effect, and the functional biomechanical outcome. In contrast to the studies that have investigated the synergy of culture topography and mechanical stimulation in a 2D environment,20,29 we have studied this synergy spatiotemporally in a 3D environment with significant temporal consideration and evaluated the functional measures for tendon and ligament regeneration.
Over the duration of this study, significant disparity was observed between the cellular behavior of MSCs seeded on the two types of construct (RD and AL) in the presence of mechanical stimuli. The mechanically functional full silk scaffold with customizable aligned topographical cues could effectively mechanotranslate mechanical cues to accelerate the tenogenic differentiation of the MSCs. This was apparent as enhanced cell proliferation, cell viability, and ECM production were observed in the dynamically cultured AL constructs when compared with the RD type similarly cultured. Under both static and dynamic conditions over the experimental period, the seeded cells in the AL group had also retained their directionality in cellular elongation along the axis of the 3D cylindrical tendon/ligament analogues. The dynamically cultured AL constructs stimulated a general upregulation of gene expression for functional tendon/ligament-related ECM proteins as early as day 7. Expression of these functional markers (collagen I, tenascin-C, and tenomodulin) consequently induced an augmented deposition of aligned collagen and tendon/ligament-related ECM components, which subsequently translated to improving the overall mechanical properties of the dynamically cultured AL constructs. Amplification of these effects, by the presence of mechanical cues, was apparent when statically and dynamically cultured AL constructs were compared.
It is noteworthy that collagen I was expressed and deposited with significant differences between the AL static and dynamic culture groups (more in AL dynamic) from day 7, while collagen III was expressed and deposited with significant differences between the same groups (more in AL dynamic) at a later time point from day 14. Between AL and RD cultured dynamically, collagen I was expressed and deposited significantly more in AL from day 7 and collagen III from day 14. Collagen I was expressed and deposited earlier and more prominently in the AL dynamically cultured group as compared to collagen III. This indicated that the combined effects of dynamic culture and aligned topography triggered regenerative cues from the cultured cells instead of the reparative type. Moreover, tenomodulin, which is responsible for tendon and ligament fibril maturation, 39 was upregulated in the dynamically cultured AL from day 7. This was translated to improved strength as observed for dynamically cultured AL when compared with the statically cultured counterpart after 14 days of culture. These findings demonstrated that the AL scaffold types were suitable for tendon/ligament functional tissue engineering, and that the presence of dynamic conditioning would help accelerate the process of attaining viable regenerated tendon/ligament for tissue reconstruction applications.
Although much of the studies that have looked at the effects of mechanical stimuli and topographical cell guidance were focused on 2D cultures,20,23,29 functional development of the tendon and ligament tissues essentially depends on the presence of a 3D spatial environment. As compared to 2D cultures, 3D cultures can provide environmental cues that mimic the native tissue to stimulate an activation of cell-surface receptors and adhesion sites responsible for regulating cell cycle and expression of key tendon/ligament ECM components.1,40–42 The advantageous topography of the AL hybrid SF scaffold further enhanced these responses as previously reported. 28 In that study, it was concluded that the AL hybrid SF scaffold could support MSC proliferation and was topographically favorable for cellular and ECM alignment. Coupled with the suitable surface chemistry provided by the SF material, MSCs were stimulated to produce elevated amounts of tendon/ligament-related proteins that were indicative of tendon/ligament fibroblast differentiation. These effects were further amplified and accelerated when mechanical stimulation was provided, whereby increased differentiative activities were observed from the increased production of tendon/ligament-related genes and proteins in dynamically cultured AL hybrid scaffolds. This indicated that not only did the SF-aligned topography mimic the native ECM in terms of its surface chemistry and topography; it also worked to effectively transfer axial strains to the attached aligned MSCs. This was supported by works conducted by Lee et al., 20 whereby effective mechanotransduction leading to enhanced ECM production was observed for human ligament fibroblasts cultured on 2D aligned electrospun PU fibers.
In fact, we propose that the presence of aligned cellular substratum topography may be a necessary element, rather than an accessorial enhancement, when it comes to providing effective cyclic straining. This is so as cells respond to mechanical stimuli by altering their orientation. Such mechanoresponse arises as a form of adaptation to applied force, which involves local regulation of dynamic cytoskeletal elements, based on the regulation of Rho family GTPases. 43 In the case when the uniaxial constant stretch is applied to 2D cultures, the actin stress fiber and focal adhesion align parallel to the applied force, leading to the favorable effect of adhesion strengthening. 43 On the contrary, cyclic straining tend to induce cellular alignment perpendicular to the applied force, 44 depending on the subjected frequency. 45 Based on a mathematical model, such cellular alignment is dependent on the rate at which the forces are applied relative to the characteristic rates of remodeling in load-bearing subcellular structures. 46 Cellular alignment perpendicular to the strain direction occurs to minimize stretching of these elements under a higher cyclic load frequency. On the other hand, cells can internally remodel and align in the direction of applied stress when the cyclic load frequency is lower than the remodeling rate. 47 Therefore, with cellular and ECM alignment predominantly depicted by contact guidance, the concerns for cellular and ECM orientation shift from the strain direction and reduced responsiveness to mechanical stimulation over culture duration can be minimal. As demonstrated in this study, cellular alignment persisted in the direction of scaffold topography and strain orientation through the culture period.
Under static culture conditions, the aligned architecture of the AL construct aided medium uptake into the construct's core through active capillary action, as evidenced from the increased cell densities and colony distribution in the core of the AL group when compared to the RD group. With mechanical conditioning, however, the medium intake was further facilitated by the active strain actions rendered to the loaded constructs, generating a pumping action to bring about effective mass transfer within the constructs. This explained why improved cell density and morphology was observed in the core of both the AL and RD scaffolds when mechanical stimulation was present.
It was clear that the AL scaffolds could promote proliferation to result in high cell density of aligned spindle-shaped cells that eventually contributed to the formation of extensive aligned ECM network. Although this formed the foundation for regeneration of functional tendons and ligaments, it was necessary for the ECM network to form 3D collagen fibers to achieve functional tissues with mechanical integrity. Under static culture, gene expression for tenomodulin was significantly upregulated at day 14 from day 7 of the AL group, which was indicative of pending tenomodulin expression to facilitate the increase of collagen fibril diameter. 39 With mechanical stimulation, the upregulation in gene expression of tenomodulin was brought forward in the dynamically cultured AL group to as early as day 7. Coupled with the increase in tendon/ligament-related ECM proteins detected in the dynamically cultured AL group, it was apparent that mechanically stimulated AL promoted accelerated tendon/ligament tissue formation and maturation, with a close structural similarity to the native tissue. From this study, it can be concluded that superior mechanical properties resulted from the aligned ECM structure of the AL constructs. Under both static and dynamic culture conditions, MSC-seeded AL constructs were observed to be significantly stronger and stiffer after being cultured for 7 and 14 days compared to the RD constructs. For both static and dynamic culture, AL and RD constructs both demonstrated significantly extended toe regions, indicated by the nonlinear increase in load displacement, after 14 days of culture when compared to the respective blank scaffolds. This could be attributed to the presence of immature and discontinuous ECM structures in these constructs, which were loaded before the scaffold. Such loading patterns were typical of native tendons and ligaments, whereby the crimp morphology of the fibrils within the matrix allowed tandem loading of the fibrils to buffer for low strains without overall fibrous damage48,49 and to absorb shock loads. 50 Although the implementation of mechanical stimulation did not significantly increase the extent of the toe region measured from the statically cultured counterparts, equivalently higher values were observed compared to the blank scaffolds. It might be necessary for a longer duration of mechanical conditioning be applied on the hybrid scaffolds before significant changes in this aspect could be seen.
As the hybrid SF scaffold could be customized to cater for different in vivo loading requirements, it was designed to fail at similar loads as the rabbit native ACL tissue in this study. On the other hand, to facilitate effective loading of the seeded MSCs, the tensile stiffness of knitted SF was designed to be not more than half that of the native ACL tissue, measured to be 47.07±14.84 N/mm for rabbit. 2 As such, stress shielding of the attached cells from mechanical stimuli could be prevented, thereby enhancing the differentiative potential of the seeded MSCs in mechanically conditioned constructs either in vitro or in vivo. In this case, in vitro mechanical conditioning had been shown to be effective and had synergistically complemented the positive stiffness and mechanotransduction property of the AL-type hybrid scaffold to promote functional regeneration of the tendon and ligament tissue.
Moving forward, the enhancements in tenogenesis brought by combined dynamic culture with aligned scaffold topography have practical significances in several areas. First, the tissue culture process from seeding to implantation of a viable construct for tendon/ligament repair can be accelerated. With shorter time necessary for MSC differentiation down the tenogenic lineage, a tissue-engineered tendon/ligament construct can be implanted into patients in reduced lead-time. This is especially significant if the MSCs are autologous. With a shorter lead-time to patients, it is expected that less resources will be necessary for construct culture and conditioning, which will lead to cost reduction. Second, the combined effects of aligned topography and mechanical stimulation will improve lineage commitment of the MSCs toward the tenogenic lineage as shown in this study. With improved lineage certainity, production consistency can be improved, making the tissue-engineered tendon/ligament construct safer as a product. Third, with cellular and ECM alignment induced by the aligned topography of the scaffold, effective mechanotransduction can be provided to the cells and construct as a whole, leading to mechanically viable constructs that can be applied in patients more readily. With improved tissue-engineered graft integrity, clinicians and patients will accept such tissue-engineered products more readily. Furthermore, since this approach focuses on functional tissue regeneration that arises from ECM alignment and mechanical conditioning, instead of a simple structural repair, this tissue-engineered graft is not as likely to fail over time after implantation.
Future studies should look into optimization of mechanical conditions necessary for accelerated tenogenic differentiation of MSCs, increased ECM deposition, collagen fiber formation, and construct remodeling. Prolonged mechanical fatigue analysis should also be carried out to understand the mechanically conditioned constructs' response to physiologically relevant cyclic stresses. It would also be interesting to look into the fabrication of multiphasic aligned SF scaffolds for the enhanced tissue-engineered graft to host tissue regeneration, via encapsulation methods such as coaxial electrospinning techniques. Preclinical in vivo studies would be necessary to determine the efficacy of the mechanically conditioned aligned constructs under biological and physiological stresses.
Conclusion
This study demonstrated that mechanical loading of MSCs within a 3D aligned environment promoted tenogenic differentiation and tissue maturation in vitro. Using a custom-made rotating wire frame collector, we could obtain highly aligned AL-SFEFs and showed that the corresponding hybrid SF scaffold could provide favorable topographical cues and surface chemistries for cellular and ECM alignment. While the aligned topography potentiated the expression of tenogenic markers in MSCs and stimulated elevated production of tendon/ligament-related proteins, the introduction of mechanical stimulus intensified and accelerated the tenogenic differentiation process. With mechanical stimulation, the cultured AL constructs were eventually significantly stronger and stiffer, displaying similar biomechanical characteristics as the native tendon and ligament tissues. Based on our results, we propose that the AL mechanoactive hybrid SF scaffold can intensify the positive effects of mechanical stimulation and synergistically combine these effects with those provided by the aligned topography of the scaffold, leading to a tissue-engineered construct that shows promise for tendon and ligament reconstruction. To our knowledge, this is the first study that reports the synergistic effect of mechanical stimuli with aligned topography in a 3D environment to both intensify and accelerate the tenogenic differentiation of MSCs.
Footnotes
Acknowledgments
This study was supported by the Faculty of Engineering, the National University of Singapore and the Agency for Science, Technology and Research (A*STAR).
Disclosure Statement
No competing financial interests exist.