
Abstract
Cartilage defects have a limited ability to self-heal. Stem cell treatment is a promising approach; however, replicative senescence is a challenge to acquiring large-quantity and high-quality stem cells for cartilage regeneration. Synovium-derived stem cells (SDSCs) are a tissue-specific stem cell for cartilage regeneration. Our recent findings suggest that decellularized stem cell matrix (DSCM) can rejuvenate expanded SDSCs in cell proliferation and chondrogenic potential. In this study, we were investigating (1) whether transforming growth factor (TGF)-β1 and TGF-β3 played a similar role in chondrogenic induction of SDSCs after expansion on either DSCM or plastic flasks (plastic), and (2) whether DSCM-expanded SDSCs had an enhanced capacity in repairing partial-thickness cartilage defects in a minipig model. SDSCs were isolated from synovium in two 3-month-old pigs and DSCM was prepared using SDSCs. Passage 2 SDSCs were expanded on either DSCM or plastic for one passage. The expanded cells were evaluated for cell morphology, chondrogenic capacity, and related mechanisms. TGF-β1 and TGF-β3 were compared for their role in chondrogenesis of SDSCs after expansion on either DSCM or plastic. The chondrogenic induction medium without TGF-β served as a control. In 13 minipigs, we intraarticularly injected DSCM- or plastic-expanded SDSCs or saline into knee partial-thickness cartilage defects and assessed their repair using histology and immunohistochemistry. We found DSCM-expanded SDSCs were small, had a fibroblast-like shape, and grew quickly in a three-dimensional format with concomitant up-regulation of phosphocyclin D1 and TGF-β receptor II. Plastic-expanded SDSCs exhibited higher mRNA levels of chondrogenic markers when incubated with TGF-β3, while DSCM-expanded SDSCs displayed comparable chondrogenic potential when treated with either TGF-β isotype. In the minipig model, DSCM-expanded SDSCs were better than plastic-expanded SDSCs in enhancing collagen II and sulfated glycosaminoglycan expression in repair of partial-thickness cartilage defects, but both groups were superior to the saline control group. Our observations suggested that DSCM is a promising cell expansion system that can promote cell proliferation and enhance expanded cell chondrogenic potential in vitro and in vivo. Our approach could lead to a tissue-specific cell expansion system providing large-quantity and high-quality stem cells for the treatment of cartilage defects.
Introduction
Repair of partial-thickness defects in articular cartilage can occur as a result of cell recruitment from the synovial membrane. 9 With both self-renewal capacity and an ability to differentiate into a variety of mesenchymal tissues,10–12 adult mesenchymal stem cells (MSCs) are considered a promising candidate cell source for cartilage engineering and regeneration. The hypoimmunogenic nature implies that MSCs can be used in allogeneic cell-based therapy. 13 Allogeneic MSCs allow therapeutic delivery of cells from healthier and younger donors with no donor-site morbidity for the patients. We previously engineered premature cartilage tissue by incubating synovium-derived stem cell (SDSC)-polyglycolic acid constructs in a serum-free chondrogenic medium in a rotating bioreactor system. 14 When we used these constructs to repair knee osteochondral defects in rabbits, we found that 6 months after implantation, cartilage defects were full of smooth hyaline-like cartilage with a high expression of collagen II and sulfated glycosaminoglycans (GAGs), which were also integrated with the surrounding native cartilage. 15
Recently, several studies16–18 suggested that intraarticular injection of bone marrow stromal cells (BMSCs) is a feasible and promising minimally invasive approach for cartilage repair. However, some concerns exist. The first concern is that sufficiency in number of MSCs is a basic requirement for MSC-based tissue regeneration. Unfortunately, the proliferative rate of MSCs usually decreases substantially after repeated passages.19,20 Moreover, repeated passaging seriously impairs MSC multipotent differentiation.19–23 On the other hand, we found decellularized stem cell matrix (DSCM) deposited by SDSCs could provide a three-dimensional (3D) stem cell expansion system in which large-quantity and high-quality stem cells could be produced.19,24–27 The second concern is that BMSCs are a stem cell for endochondral bone formation. 28 In contrast, SDSCs are tissue-specific stem cells for chondrogenesis. 29 A tissue-specific cell expansion system providing large-quantity and high-quality stem cells would be important for treating cartilage defects.
Transforming growth factor (TGF)-β1 and TGF-β3 are two isotypes that are often used in the induction of adult stem cells toward chondrogenic differentiation30–33 ; however, there were no data comparing their chondrogenic induction effect on SDSCs after expansion on either DSCM or plastic flasks (plastic). Despite in vitro data collected in our previous studies demonstrating an enhanced chondrogenic potential of DSCM-expanded SDSCs,19,24,25,34 we proposed that a direct intraarticular injection of SDSCs could repair partial-thickness cartilage defects in a minipig model and that DSCM expansion could enhance the SDSC resurfacing effect. We tested the hypotheses that (1) DSCM expansion enhanced SDSC proliferation and chondrogenic potential and DSCM-expanded SDSCs displayed comparable chondrogenic potential when treated with either TGF-β isotype, and (2) an allogeneic source of SDSCs could repair partial-thickness cartilage defects and DSCM-expanded SDSCs could enhance cartilage defect regeneration.
Materials and Methods
Isolation and culture of porcine SDSCs
Random biopsies of synovial membrane were obtained aseptically from both knees of two 3-month-old pigs from a local slaughterhouse and pooled together for this study. The harvested tissue was digested at 37°C for 30 min in 0.1% trypsin (Roche, Indianapolis, IN), followed by 2 h in 0.1% collagenase P (Roche). Cells were resuspended and plated in culture medium (α-minimum essential medium [Invitrogen, Carlsbad, CA] containing 10% fetal bovine serum [Atlanta Biologicals, Lawrenceville, GA], 100 U/mL penicillin, 100 μg/mL streptomycin, and 0.25 μg/mL fungizone [Invitrogen]) at 37°C in a humidified 5% CO2 and 21% O2 incubator. Nonadherent cells were removed by medium change every 3 days. For negative isolation of SDSCs from primary cultures, 35 the cell suspension was incubated with Dynabeads® M-450 CD14 conjugated with a monoclonal antibody specific for macrophages (Dynal Biotech, Oslo, Norway) at 4°C for 1 h. The conjugated cells and the unbound Dynabeads were collected using the Dynal Magnetic Particle Concentrator® (Dynal Biotech) and the depleted supernatant with SDSCs was saved for further passaging.
Preparation of SDSC-deposited DSCM
The procedure for obtaining cell-free extracellular matrix was described previously.19,25 Plastic flasks were pretreated with a 0.2% gelatin solution (Sigma-Aldrich, St. Louis, MO) at 37°C for 1 h, followed by 1% glutaraldehyde (Sigma-Aldrich) and 1 M ethanolamine (Sigma-Aldrich) for 30 min separately at room temperature. SDSCs were seeded on pretreated plastic flasks at 3000 cells/cm2 in culture medium for 7 days. After reaching 90% confluence, 50 μM
In vitro expansion of SDSCs on plastic and DSCM
To determine whether DSCM could provide an in vitro microenvironment for cell expansion, passage 2 SDSCs were expanded at 3000 cells/cm2 in a culture medium for 7 days under two conditions: conventional plastic and flasks coated with SDSC-deposited DSCM. The medium was changed every 3 days. The expanded cells were evaluated for cell morphology using phase-contrast microscopy (Olympus CK40; Olympus Optical, Tokyo, Japan) and for expression of phosphocyclin D1 (p-cyclin D1) and TGF-β receptor II (TGF-β RII) using western blot. Briefly, radioimmunoprecipitation assay lysis buffer supplemented with halt protease and phosphatase inhibitor cocktail (Thermo Scientific, Milford, MA) was used to extract protein. The samples were denatured and separated by NuPAGE® Novex® Bis-Tris Mini Gels (Invitrogen) at 200 V at 4°C for 45 min and then transferred onto a nitrocellulose membrane (Invitrogen) using XCell II™ blot module (Invitrogen) at 30 V at 4°C for 1 h. The membranes were first blocked in SuperBlock® (Tris-buffered saline) Blocking Buffer (Thermo Scientific) and then incubated with the primary monoclonal antibodies (TGF-β RII; Santa Cruz Biotechnology, Santa Cruz, CA), p-cyclin D1 (Thr286; Cell Signaling, Danvers, MA), and β-actin (Abcam, Cambridge, MA; serving as an internal control) at the recommended dilution at 4°C overnight. The membranes were incubated in the secondary antibody of goat anti-mouse immunoglobulin G (IgG) (H+L) or goat anti-rabbit IgG (H+L) (Thermo Scientific) for 40 min. The exposure used SuperSignal® West Pico Chemiluminescent Substrate (Thermo Scientific) and CL-XPosure™ Film (Thermo Scientific).
In vitro chondrogenic differentiation of SDSCs after expansion on plastic and DSCM
To determine in vitro chondrogenic differentiation potential, 3×105 of SDSCs expanded on either plastic or DSCM were centrifuged to form a pellet in a 15-mL polypropylene tube at 500 g for 5 min. After 24 h, the pellets were cultured in three conditions with 30 pellets in each group: the control group was incubated in chondrogenic medium (high-glucose Dulbecco's Modified Eagle's Medium, 40 μg/mL proline, 100 nM dexamethasone [Sigma-Aldrich], 100 U/mL penicillin, 100 μg/mL streptomycin, 0.1 mM ascorbic acid-2-phosphate, and ITS™ Premix [BD Biosciences, Bedford, MA]); the TGF-β1 group was incubated in chondrogenic medium supplemented with 10 ng/mL TGF-β1 (PeproTech, Inc., Rocky Hill, NJ); and the TGF-β3 group was incubated in chondrogenic medium supplemented with 10 ng/mL TGF-β3 (PeproTech, Inc.). All the groups were cultured for an additional 14 days and pellets at Day 0, 7, and 14 were collected for chondrogenic differentiation analysis, including biochemical analysis for chondrogenic index, real-time polymerase chain reaction (PCR) for chondrogenic marker genes, and histology and immunostaining for chondrogenic markers.
To examine DNA and GAG content in pellets, samples (n=4) were digested at 60°C for 6 h in PBE buffer (100 mM phosphate, 10 mM ethylenediaminetetraacetic acid [EDTA], pH 6.5) containing 125 μg/mL papain and 10 mM cysteine. DNA content was measured using QuantiT™ PicoGreen® dsDNA assay kit (Invitrogen) and CytoFluor® Series 4000 (Applied Biosystems, Foster City, CA) following the manufacturer's protocol. The GAG content was measured using dimethylmethylene blue dye (Sigma-Aldrich) and Spectronic™ BioMate™ 3 Spectrophotometer (Thermo Scientific) with bovine chondroitin sulfate (Sigma-Aldrich) as the standard.
To determine chondrogenic marker gene expression in pellets, the total RNA from pellets (n=4) was extracted using TRIzol® (Invitrogen) and converted to cDNA with High-Capacity cDNA Archive Kit as recommended by the manufacturer (Applied Biosystems). Chondrogenic marker genes (SRY [sex determining region Y]-box 9 [Sox9], aggrecan [AG], and collagen II [Col II]) and hypertrophy marker genes (collagen X [Col X] and alkaline phosphatase [ALP]) were customized by Applied Biosystem as part of the Custom TaqMan® Gene Expression Assays. Custom gene expression assays ordered through the online design tool goes through bioinformatic analysis if done through the custom plus option. Our lab validation included the amplification efficiency check using our custom primer and probe set. Eukaryotic 18S rRNA (Assay ID HS99999901_s1 ABI) was carried out as the endogenous control gene. Real-time PCR was performed with the iCycler iQ™ Multi Color RT-PCR Detection (Bio-Rad Laboratories, Inc., Hercules, CA) and calculated by computer software (Bio-Rad Laboratories, Inc.). Relative transcript levels were calculated as χ=2−ΔΔCt, in which ΔΔCt=ΔE−ΔC, ΔE=Ctexp−Ct18s, and ΔC=Ctct1−Ct18s.
Intraarticular injection of DSCM-expanded SDSCs for cartilage repair
To determine partial-thickness cartilage defect repair using DSCM- or plastic-expanded SDSCs, we used 13 miniature pigs (2-year-old females weighing 38.1–53.7 kg) (Marshall BioResources, North Rose, NY). The minipigs were anesthetized with 30 mg/kg ketamine (Phoenix Pharmaceutical, St. Joseph, MO) and 1.1 mg/kg acepromazine (Phoenix Pharmaceutical) intramuscularly and maintained with isoflurane. The knee joints were approached through medial parapatellar incisions and the articular surfaces were exposed by medial dislocation of the patellae. Partial-thickness cartilage defects, 8 mm diameter×1 mm deep, were created in the medial femoral condyles of both knees using a customized tube osteotome with a depth-limiter. This project was approved by the Institutional Animal Care and Use Committee and conducted in compliance with the National Advisory Committee for Laboratory Animal Research Guidelines.
After 10 days, expanded cells were injected into knees with cartilage defects. Our experimental design included three groups according to injected solution: DSCM-expanded SDSCs in saline (five pigs/10 knees), plastic-expanded SDSCs in saline (five pigs/10 knees), and saline alone (three pigs/six knees). Post-hoc power analyses were performed using a significance level of 0.001 (using Bonferroni correction for multiple testing) and six samples per group, assuming a two-sided alternate hypothesis and signal-to-noise ratio of 6, we could achieve close to 100% power. Before intraarticular injection, expanded cells were prestained by suspending them at a density of 1×106/mL in a serum-free culture medium and mixing them with 5 μL Vybrant® DiI cell labeling solution (1:200; Invitrogen) by gentle pipetting. After incubating at 37°C for 20 min, the mixture was centrifuged at 1500 rpm for 5 min. The cells were ready for use after being washed twice using prewarmed medium. The cells were then resuspended in saline and injected into each knee. To control moderate acute pain, buprenorphine was administered peri-operatively at a dosage of 0.03 mg/kg. Immediately following closure of the incision, 5% Marcaine (bupivacaine) was injected along each incision to alleviate pain; each knee was injected with 3 mL of Marcaine. All minipigs were returned to their cages after the operation and were allowed to move freely. The animals had ketoprofen capsules (75 mg) twice daily for 3 days postoperatively as an analgesic. Three months after implantation, animals were euthanized by an intracardiac puncture of Euthasol® euthanasia solution (Fort Dodge Animal Health, Fort Dodge, IA). The femoral condyles were isolated for histomorphological analysis.
Histology and immunohistochemistry
For in vitro histology, the representative pellets (n=2) were fixed in 4% paraformaldehyde, embedded in paraffin blocks, and cut into 5-μm-thick sections. For in vivo histology, the dissected distal femurs were decalcified at 4°C in 4% EDTA solution on a shaker. Radiographs were used to evaluate the progress of decalcification. After decalcification, the samples were dehydrated with a gradient ethanol series, cleared with xylene, and embedded in paraffin blocks. To detect sulfated GAGs, sections were stained with Safranin O and Alcian blue (Sigma-Aldrich). For immunohistochemistry, the sections were probed with primary antibodies against collagen I (Sigma-Aldrich), collagen II (II-II6B3; Developmental Studies Hybridoma Bank [DSHB], Iowa City, IA), and collagen X (Sigma-Aldrich), followed by the secondary antibody of biotinylated horse anti-mouse IgG (Vector, Burlingame, CA) and detected by using Vectastain ABC reagent (Vector) with 3,3′-diaminobenzidine as a substrate. Hematoxylin (Vector) served as a counterstain. A section from full-thickness normal cartilage with subchondral bone was used as a control. Cartilage served as a positive control for collagen II staining. Subchondral bone stained positively for collagen I. The tidemark generally stained positively for collagen X.
Four of us (M.P., J.T.L., Y.Z., B.J.) evaluated histologic sections of the repaired tissue using a grading system 36 consisting of five categories (cell morphology, matrix staining, surface regularity, cartilage thickness, and integration of donor with host) (Table 1). The scoring was performed in a blinded manner.
Total smooth area of the reparative cartilage compared with the entire area of the cartilage defect.
Average thickness of the reparative cartilage compared with that of the surrounding cartilage.
Statistical analysis
The Mann-Whitney U test was used for pairwise comparison in biochemical analysis (n=4), real-time PCR (n=4), and semi-quantitative histological scoring (n≥6). Statistical analyses were performed with SPSS 13.0 statistical software (SPSS, Inc., Chicago, IL). All data are expressed as means±standard deviation of the mean. p-Values less than 0.05 were considered statistically significant.
Results
DSCM expansion enhanced SDSC proliferation and chondrogenic potential
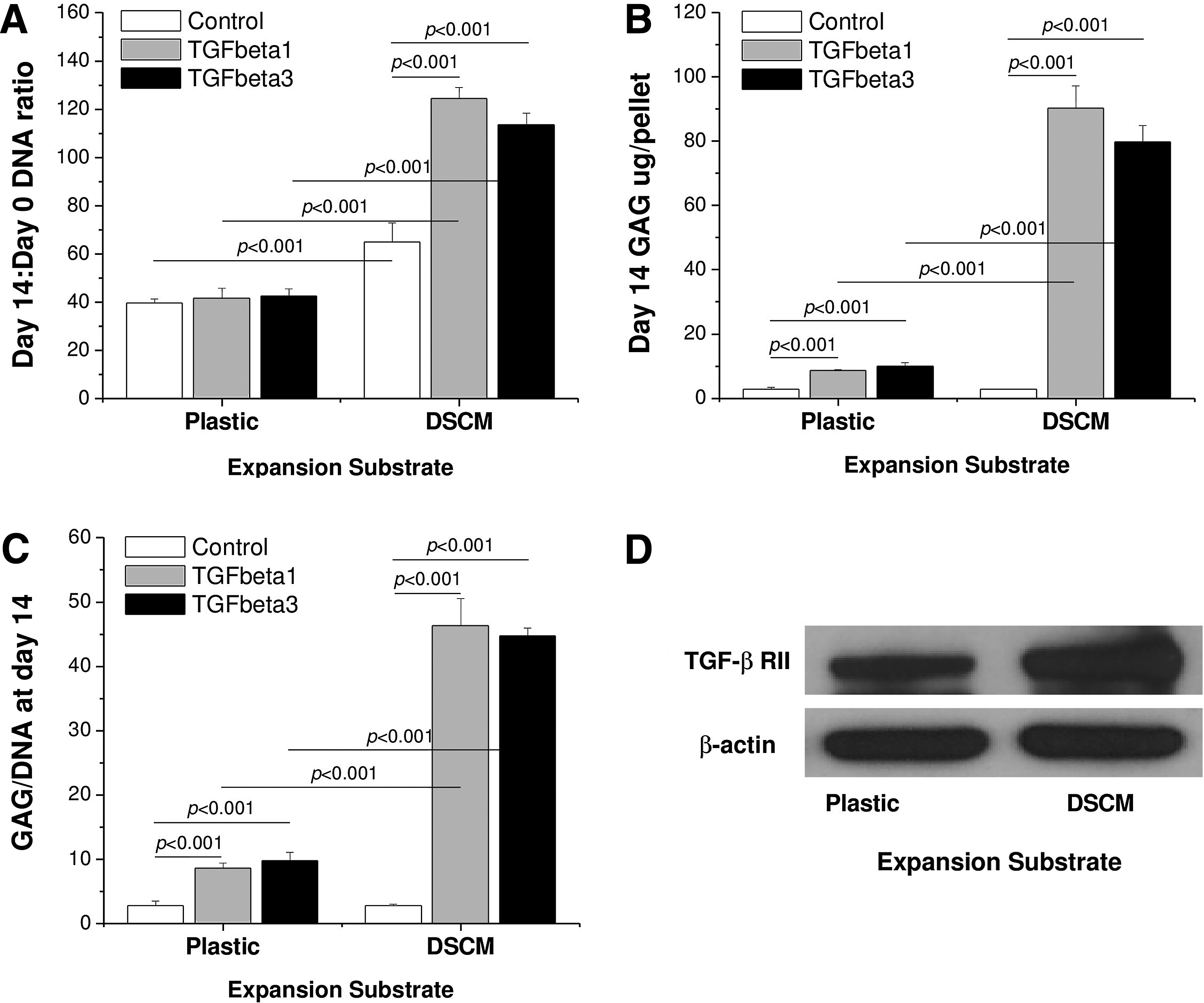
DSCM expansion enhanced SDSC proliferation and chondrogenic potential, and DSCM-expanded SDSCs displayed comparable chondrogenic potential when treated with either TGF-β isotype. Despite the initial cell density being similar in both groups, cells expanded on DSCM grew more by overlapping in a 3D format with a small fibroblast-like shape (Fig. 1A). p-cyclin D1 was nondetectable in plastic-expanded cells, whereas p-cyclin D was up-regulated in DSCM-expanded cells (Fig. 1B). DSCM expansion enhanced SDSC chondrogenic potential, as evidenced by intensified staining of collagen II and sulfated GAGs in the larger pellets from DSCM-expanded cells in chondrogenic medium supplemented with either TGF-β1 or TGF-β3 (Fig. 2). Collagen I staining was intensified in the pellets from plastic-expanded SDSCs, indicating DSCM-expanded SDSCs were more sensitive to chondrogenic induction and became mature earlier than plastic-expanded SDSCs (Fig. 2). The histology data (Fig. 2) also suggested that both TGF-β isotypes exhibited similar chondrogenic induction in pellets from both expanded SDSCs, which was also confirmed by our biochemical analysis data (Fig. 3A). DSCM expansion enhanced SDSC survival in pellets cultured in a serum-free chondrogenic medium. However, if incubated in the absence of TGF-β, DSCM-expanded SDSCs yielded pellets with low GAG content and chondrogenic index, comparable to those from plastic-expanded SDSCs. In contrast, if incubated in the presence of TGF-β, DSCM-expanded SDSCs yielded pellets with enhanced GAG content and chondrogenic index compared to those from plastic-expanded SDSCs. Interestingly, DSCM-expanded SDSCs exhibited an up-regulation of TGF-β RII (Fig. 3B). Chondrogenic markers, including Sox9 (Fig. 4A), Col II (Fig. 4B), and AG (Fig. 4C), were also up-regulated in the pellets from DSCM-expanded SDSCs after incubation in the chondrogenic medium with TGF-β, although there was no difference between the two TGF-β isotypes. Intriguingly, chondrogenic induction with TGF-β3 favored plastic-expanded SDSCs toward chondrogenesis at the mRNA level (Fig. 4A–C) despite no difference being found at the protein level (Figs. 2 and 3C). Col I mRNA (Fig. 4D), an early expression gene in chondrogenesis, 37 was down-regulated in the pellets from DSCM-expanded SDSCs when incubated in TGF-β-containing medium for 14 days, which is consistent with our histology data. The addition of TGF-β in chondrogenic medium down-regulated Col X mRNA levels, a hypertrophic marker, 38 in SDSCs after expansion on either DSCM or plastic (Fig. 4E). While there was no difference in the induction of chondrogenic differentiation of plastic-expanded SDSCs by the two TGF-β isotypes, TGF-β1 induced decreased Col X (Fig. 4E) and ALP (Fig. 4F) mRNAs compared to TGF-β3 in DSCM-expanded SDSCs, suggesting that TGF-β-mediated chondrogenic induction could lower hypertrophic marker levels in SDSCs, especially in those from DSCM expansion.

Decellularized stem cell matrix (DSCM)-mediated synovium-derived stem cell (SDSC) expansion through the cyclin D pathway is shown. Passage 2 SDSCs were plated on either DSCM or plastic for 7 days.
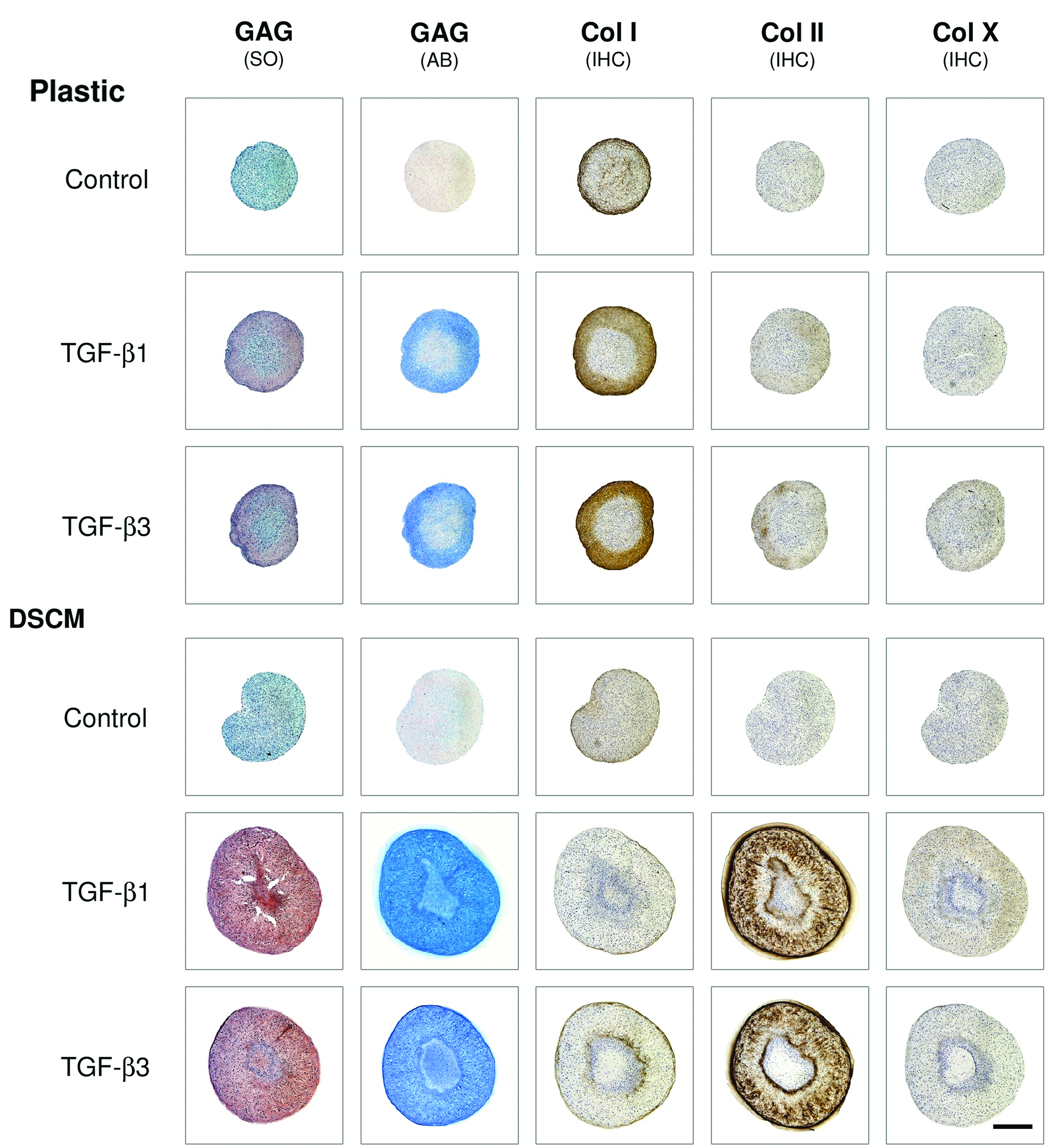
Under the treatment of a serum-free chondrogenic medium with either transforming growth factor (TGF)-β1 or TGF-β3 for 14 days, DSCM-expanded SDSCs yielded pellets with intensified staining of chondrogenic markers compared to those from plastic-expanded SDSCs. Safranin O (SO) and Alcian blue (AB) were used to stain sulfated glycosaminoglycans (GAGs); immunohistochemistry (IHC) was used to stain collagen I (early marker of chondrogenesis), collagen II (chondrogenic marker), and collagen X (hypertrophic marker). Scale bar is 800 μm. Color images available online at www.liebertpub.com/tea
DSCM-expanded SDSCs yielded 14-day pellets with higher

DSCM-expanded SDSCs yielded 14-day pellets with higher mRNA levels of chondrogenic marker genes [
DSCM expansion enhances SDSC in vivo cartilage regeneration
An allogeneic source of SDSCs could repair partial-thickness cartilage defects and DSCM-expanded SDSCs could enhance cartilage defect regeneration. Compared to the saline control, cartilage defects treated with plastic- or DSCM-expanded SDSCs were resurfaced with a whitish tissue (Fig. 5). In the saline control group, tissue in the cartilage defect area was barely detectable by immunostaining; in contrast, new tissues covered the cartilage defect area in both SDSC groups. Compared to the newly formed tissues from the group injected with plastic-expanded SDSCs, DSCM-expanded SDSC injection appeared to produce new tissue with intensified staining of GAG and collagen II but no obvious staining of collagen I. We did not include the Safranin O staining for in vivo samples because the in vivo sections could not be stained by this dye, possibly due to decalcification. Therefore, we used Alcian blue staining instead. Although both SDSC implantation groups were superior to the saline group (1.50±1.29) (p<0.001), histological scoring of cartilage repair was higher (p=0.004) for implantation of DSCM-expanded SDSCs (12.75±0.96) than for implantation of plastic-expanded SDSCs (9.75±0.96) (Fig. 6), indicating that DSCM pretreatment could enhance SDSC in vivo cartilage regeneration compared to immature resurfaced cartilage tissue from the implantation of plastic-expanded SDSCs.
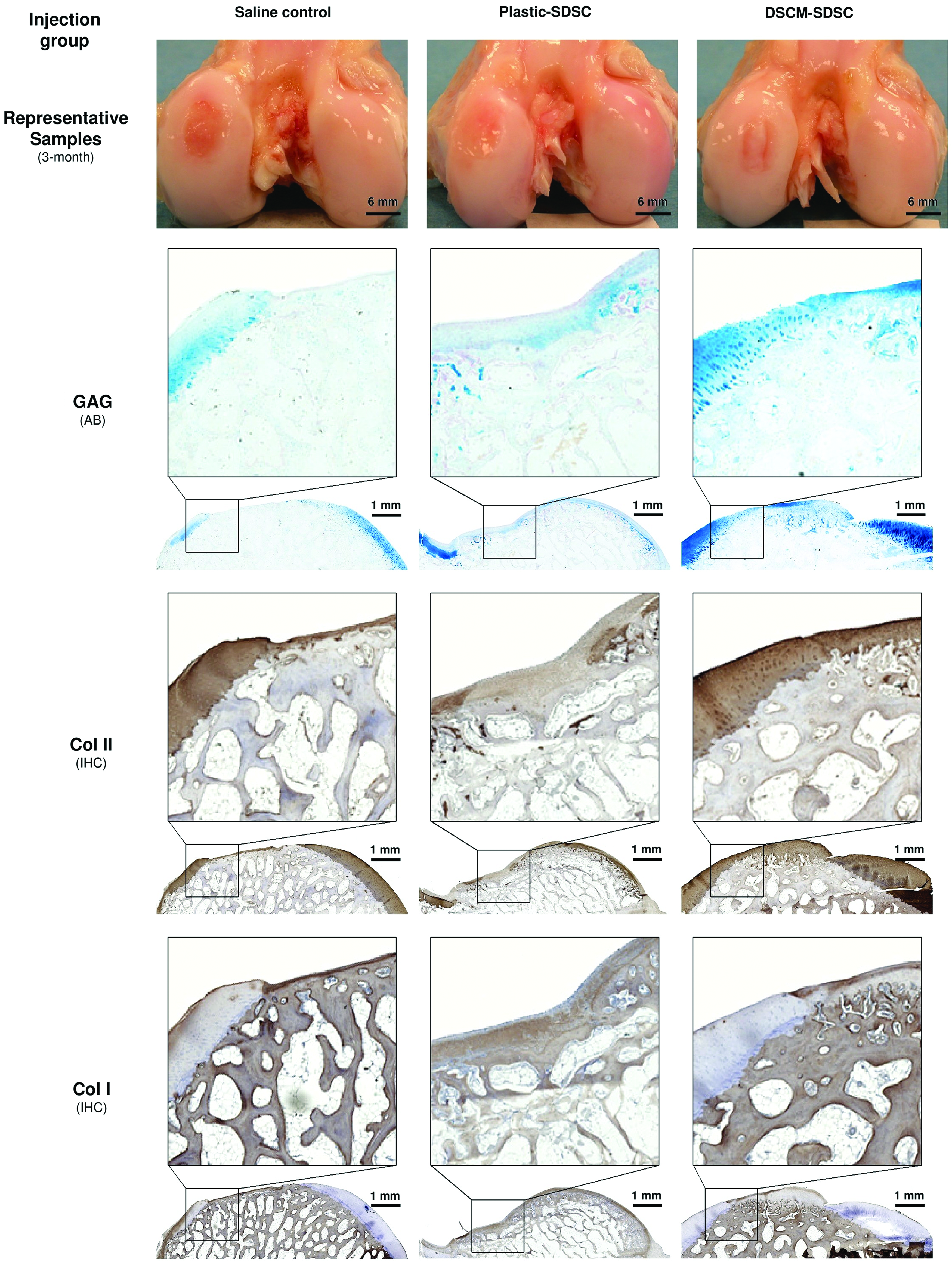
Three months after implantation, the DSCM-expanded SDSC (DSCM-SDSC) implantation group was compared to the plastic-expanded SDSC (plastic-SDSC) implantation group and the saline control group. AB staining was used for sulfated GAGs and IHC staining was for protein expression of collagens I and II. Color images available online at www.liebertpub.com/tea
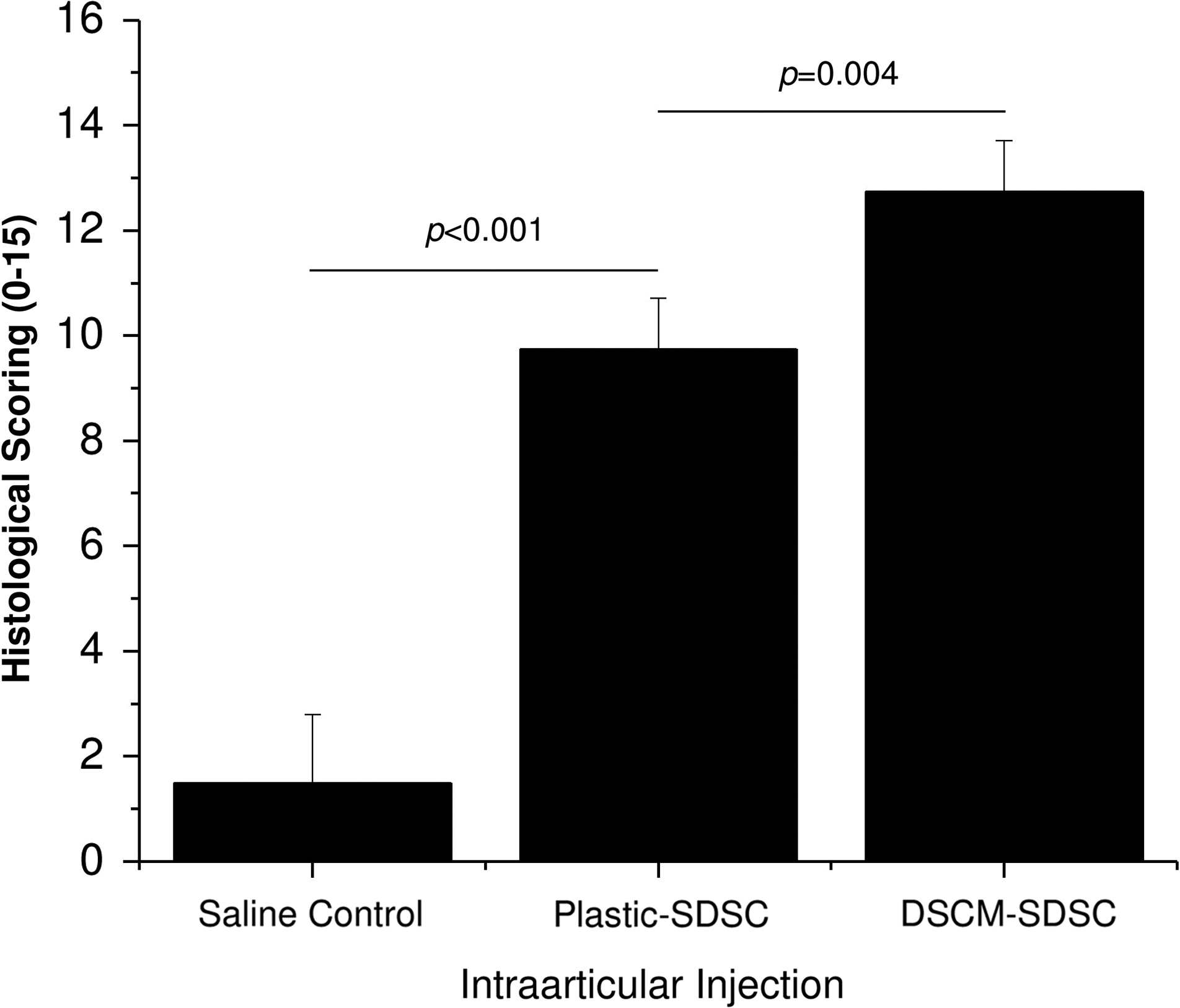
The histological scoring system was used to evaluate the regeneration of repair tissue and integration with the surrounding native tissue. Data are shown as average±SD for n=6–10.
Discussion
Our findings suggest expansion on DSCM enhanced SDSC proliferation, possibly through the up-regulation of cyclin D1, which is a reliable index for proliferation rate 39 ; DSCM-expanded SDSCs exhibited enhanced chondrogenic capacity in a pellet culture system, as evidenced by histology, immunostaining, biochemistry, and real-time PCR, possibly through the up-regulation of TGF-β RII, which has a direct relationship with stem cell chondrogenesis. 40 The data were in accordance with our recent reports.19,24–26 In this study, we also found no difference between TGF-β1 and TGF-β3 in chondrogenic induction of SDSCs after expansion on DSCM. TGF-β is a key component in serum-free chondrogenic medium. However, there was controversy concerning the effect of TGF-β isotypes on MSC chondrogenic differentiation. Of the two commonly used isotypes, TGF-β3 reportedly had a higher chondrogenic induction in human BMSCs than TGF-β1. 30 Another study indicated chondrogenic conditioning with both TGF-β isotypes similarly induced human BMSC hypertrophy and allowed the enhancement of the hypertrophic chondrocyte phenotype by hypertrophic medium. 33 A third study suggested TGF-β1 treatment led to an increase in chondrogenic-specific gene expression as compared with moderate effects of TGF-β3 in mouse suture-derived mesenchymal cells 31 and human amniotic fluid-derived stem cells. 32 Despite comparable data in histology, cell viability, and chondrogenic index of SDSCs after expansion on either DSCM or plastic, our data also indicated that, at the mRNA level, TGF-β3 was better than TGF-β1 in chondrogenic induction of plastic-expanded SDSCs. DSCM-expanded SDSCs showed comparable chondrogenic differentiation under the treatment of TGF-β isotypes at both the protein and mRNA levels.
For the first time, we demonstrated DSCM-expanded SDSCs also exhibited an enhanced in vivo cartilage regeneration capacity compared to those grown on plastic, evidenced by repairing partial-thickness cartilage defects through intraarticular injection in a minipig model. We found that intraarticular injection of plastic-expanded SDSCs promoted cartilage regeneration in partial-thickness cartilage defects in minipigs compared to nontreated controls. More importantly, we found intraarticular injection of DSCM-expanded SDSCs yielded the most improved cartilage resurfacing, evidenced by intensified staining of hyaline cartilage markers and comparable depth of regenerated cartilage with surrounding host cartilage tissue. Despite a better resurfacing cartilage in the implant group with plastic-expanded SDSCs than in the saline control, the existence of collagen I in the regenerated cartilage suggested fibrocartilage, which was nondetectable in the resurfacing cartilage in the group implanted with DSCM-expanded SDSCs. This outcome might result from the harsh environment in minipig knees where the partial-thickness cartilage defects were created. The harsh environment, including inflammatory cytokines, metalloproteinases, nitric oxide, serum deprivation, or mechanical forces, might cause implanted cell necrosis or apoptosis. 41 Inflammatory stress reportedly causes dedifferentiation of chondrogenic cells to fibroblast-like cells, resulting in the defects being filled with a mix of fibrous and cartilaginous tissues. 42 Our in vitro data demonstrated DSCM-expanded SDSCs could better survive a serum-free chondrogenic induction in a pellet culture system. Implantation with DSCM-expanded SDSCs exhibited hyaline cartilage resurfacing, indicating that DSCM pretreatment could not only enhance the chondrogenic capacity of SDSCs but also may promote the ability to resist inflammation, which needs to be further validated.
Some limitations in this study deserve further discussion. First, we used DiI to prestain SDSCs to trace implanted cells, but we did not observe a fluorescent signal in histology samples 3 months after injection. One possibility is that an organic reagent, such as xylene, applied in the paraffin embedding process, might have dissolved the DiI. Cell labeling with green fluorescent protein may provide a better alternative in a future study. Another possibility could be that the injected cells might not be directly repairing cartilage defects but are causing paracrine signaling effects that promote cartilage repair. To elucidate a mechanism of repair, we tried to collect synovial fluid samples from the injected knees after we euthanized the minipigs. Unfortunately, in most of the minipigs, we did not get sufficient samples for biomarkers of inflammatory mediators, which might provide another approach for comprehensive analysis of OA and cartilage regeneration. We will include blood collection in a future study. This information would help us to understand the difference in cell behavior and the repair process among groups. Second, the regenerated cartilage exhibited intensified staining with hyaline cartilage markers in the defects injected with DSCM-expanded SDSCs. This finding suggests that it is possible that DSCM expansion enhances the ability of SDSCs to resist inflammation and free radicals caused by the surgical procedure, such as intraarticular bleeding and stimulation of the synovial membrane that interfere with the viability, maturation, and integration of a cell-based graft. This phenomenon deserves further investigation. In addition, 3 months for cartilage regeneration seems short, which might be responsible for the gap between regenerated cartilage and host cartilage in the group treated with DSCM-expanded SDSCs. Fourth, our previous studies also suggest that predifferentiation of allogeneic SDSCs after in vitro induction are more active in generating and organizing hyaline-like cartilage without immune rejection. 15 In the future, predifferentiated SDSCs will be assessed for their capacity to survive and differentiate in this harsh environment.
Clinical benefits abound for SDSC use in cartilage engineering and regeneration. Many of the properties that set SDSCs apart from other MSCs are particularly appealing, but ease of access and removal of synovium for cartilage defect repair results in less morbidity for the patient. Synovium can be obtained with few complications in a minimally invasive fashion during arthroscopy. A small punch biopsy plus our DSCM approach would be sufficient for growth of a clinically useful quantity of cells. Additionally, the regenerative capacity of synovium after surgical and chemical synovectomy has been demonstrated.43,44 It is important to note that two subsets of SDSCs can be obtained from humans: fibrous SDSCs and adipose SDSCs. 45 Fibrous SDSCs (used in this study) can be harvested from such places as the lateral part of the knee joint capsule that overlays the noncartilaginous area of the lateral condyle of the femur. Adipose SDSCs are located in the infrapatellar fat pad overlying the patellar tendon. Both SDSCs were superior in their chondrogenic potential. 24 Although adipose SDSCs may not have all the qualities of fibrous SDSCs, these cells may be a better choice for use in patients with OA since fewer inflammatory changes are associated with the infrapatellar fat pad. 46
Taken together, the findings from this study suggested that DSCM enhanced SDSC expansion and chondrogenic potential. There was no significant difference between TGF-β1 and TGF-β3 in chondrogenic induction of SDSCs after expansion on either DSCM or plastic. Intraarticular injection of plastic-expanded SDSCs enhanced partial-thickness cartilage defect resurfacing in our minipig model; more importantly, we found that DSCM-expanded SDSCs were superior to plastic-expanded SDSCs in the allogeneic repair of partial-thickness cartilage defects. We believe that our DSCM approach would also benefit autologous SDSCs for the treatment of cartilage defects with the advantage of there being no disease transmission and immune rejection issues. Our study, for the first time, demonstrated that DSCM is a novel and promising cell expansion system that not only promoted cell proliferation but also enhanced expanded cell chondrogenic potential, evidenced by both in vitro and in vivo studies.
Footnotes
Acknowledgments
We thank Suzanne Smith for editing the manuscript. We thank Brendan Jones (B.J.) and Ying Zhang (Y.Z.) for their help in histologic scoring evaluation and also thank Nina Clovis, Suzanne Smith, and Vincent Kish for their technical support in animal anesthesia and postoperative care. This study was supported by the AO Research Fund of the AO Foundation (Project No. S-08-67P) and a Peer-Reviewed Research Grant from the Musculoskeletal Transplant Foundation.
Disclosure Statement
No competing financial interests exist.