
Abstract
Bone-morphogenetic protein 2 (BMP2) is currently the only Food and Drug Administration-approved osteoinductive growth factor used in clinical settings for bone regeneration and repair. However, the use of BMP2 is encumbered by numerous clinical complications, including postoperative inflammation and life-threatening cervical swelling. Thus, methods to prevent BMP2-induced inflammation would have far-reaching clinical implications toward improving current BMP2-based methods for bone regeneration. For the first time, we investigate the potential role of the growth factor Nel-like molecule-1 (NELL-1) in inhibiting BMP2-induced inflammation. Adult rats underwent a femoral bone onlay procedure, treated with either BMP2 protein (4 mg/mL), NELL-1 protein (4 mg/mL), or both proteins combined. Animals were evaluated at 3, 7, and 14 days postoperatively by histology, histomorphometry, immunohistochemistry, and real-time PCR for markers of inflammation (TNFα, IL6). The relative levels of TNFα and IL6 in serum were also detected by ELISA. The mechanism for NELL-1's anti-inflammatory effect was further assessed through examining inflammatory markers and generation of reactive oxygen species (ROS) in the mouse embryonic fibroblast NIH3T3 cells. BMP2 significantly induced local inflammation, including an early and pronounced polymorphonuclear cell infiltration accompanied by increased expression of TNFα and IL6. Treatment with NELL-1 alone elicited no significant inflammatory response. However, NELL-1 significantly attenuated BMP2-induced inflammation by all markers and at all timepoints. These local findings were also confirmed using systemic serum inflammatory biomarkers (TNFα, IL6). In each case, NELL-1 fully reversed BMP2-induced systemic inflammation. Lastly, our findings were recapitulated in vitro, where NELL-1 suppressed BMP2 induced expression of inflammatory markers, as well as NF-κB transcriptional activity and generation of ROS. BMP2-induced inflammation is a serious public health concern with potentially life-threatening complications. In the present study, we observed that the growth factor, NELL-1, significantly attenuates or completely reverses BMP2-induced inflammation. The mechanisms of NELL-1's anti-inflammatory effect are only partially elucidated, and may include reduction of NF-κB transcriptional activity or ROS generation.
Introduction
Many studies have indicated that BMP2-induced inflammation is the major cause of soft tissue swelling, seroma formation, and postoperative radiculitis.7,20,24,25 This inflammation is attributable to higher dosing of BMP2, often over that required for induction of human bone formation.26,27 Animal models have previously demonstrated the dose dependent nature of BMP2 treatment and its inflammatory response.26,28 Moreover, in vitro assays have demonstrated that BMP2-induced inflammation is detectable in various cell types, such as endothelial cells, fibroblasts, 29 and preosteoblasts. 30 On a cellular level, BMP2 is known to be chemotactic for lymphocytes, monocytes, and macrophages. 31 Mechanistically, BMP2's induction of inflammation is known to be mediated through a reactive oxygen species (ROS)-dependent mechanism, by the generation of ROS in endothelial cells and preosteoblasts. 30 Thus and in summary, BMP2-induced inflammation is a well-studied phenomenon and potentially life threatening complication, attributable to an increase in inflammatory cell chemotaxis and an increase in ROS generation among other potential mechanisms.
Nel-like molecule-1 (NELL-1) is a growth factor whose osteoinductive properties were first identified in craniosynostosis patients. Recombinant NELL-1 protein has been observed to induce bone and cartilage formation and suppress adipogenesis in numerous in vitro and in vivo models.32–40 Although the precise NELL-1 signaling pathways are only partially understood, NELL-1 positively regulates Runt-related transcription factor-2 (Runx2) activity and phosphorylation, 41 MAPK signaling, 42 and Hedgehog signaling pathways. 37 In addition, NELL-1 has been described to have combinatorial or synergistic bone-forming effects when combined with BMP2 in various in vivo models, including a nude mouse intramuscular model, rat calvarial defect model, and a rabbit maxillary sinus floor elevation model.43–45 Recently, our laboratory performed preliminary studies geared toward bone regeneration, in which to our surprise we found that NELL-1 also significantly attenuated BMP2-induced inflammation. In this study, we formally investigated whether the growth factor NELL-1 could reduce the inflammation frequently encountered with BMP2 treatment.
Materials and Methods
Antibodies and reagents
Human recombinant NELL-1 was purified by Aragen Bioscience (Purity >98%). Human BMP2 (INFUSE Bone Graft) was purchased from Medtronic. Primary antibodies used in this study were anti-TNFα (AF-510-NA; R&D) and anti-IL6 (AF506; R&D). NIH3T3 fibroblasts, Human endothelial cells (HUVEC), C2C12 myoblasts, and J774A.1 macrophages were purchased from American Type Culture Collection (ATCC). NIH3T3, C2C12, and J774A.1 cells were cultured in DMEM+10% FBS. HUVEC were grown in EBM2 MV Bulletkit (Lonza). All the cells were sustained in an incubator that maintained an environment with a temperature of 37°C and 5% CO2.
Surgical procedure and study groups
All animal procedures were conducted in accordance with the Guide for the Care and Use of Laboratory Animals of the University of California and approved by the Chancellor's Animal Research Committee. Surgical procedure was conducted as previously described. 26 A concentration of 4 mg/mL of BMP2 was used in this study as that concentration is proven to consistently induce inflammation. 26 The ratio of volume of reagent to sponge area was kept constant between all test groups and was based on the ratio recommended in the manufacturer's manual (INFUSE Bone Graft, Medtronic). Briefly, 74 male Lewis rats aged 12–14 weeks were randomized into four experimental groups according to sacrifice timepoints of 3, 7, or 14 days postoperative (Table 1). Helistat absorbable collagen sponges (ACS; thickness 1 mm, porosity 88.2%; Integra, Plainsboro, NJ) were cut into 1×2 cm strips. Sponges were saturated by dropwise application of 0.2 mL of either: (1) phosphate buffered saline (PBS), (2) BMP2 (total dose: 0.8 mg), (3) NELL-1 (total dose: 0.8 mg), or (4) NELL-1+BMP2 combined (total dose of BMP2: 0.8 mg; total dose of NELL-1: 0.8 mg), and allowed to soak for 15 min before implantation. Human BMP2 and NELL-1 were dissolved in PBS.
BMP2, bone-morphogenetic protein 2; NELL-1, Nel-like molecule-1; PBS, phosphate buffered saline.
Animals were anesthetized by isoflurane inhalation. After anesthesia, the hind limbs were shaved, aseptically scrubbed and a transverse incision was made at the level of the midpoint of the femur. The vastus lateralis and biceps femoris were elevated through an anterolateral approach with care to keep the periosteum intact along the surface of the bone. ACS with or without growth factors was placed over the femoral surface, adjacent to the muscle and fascia. Each layer was closed with 4–0 Vicryl absorbable suture. After surgery, all animals were housed in separate cages and allowed to eat and drink ad libitum. Weight bearing was started immediately postoperatively, with daily monitoring. Buprenorphine was administered for 2 days as an analgesic, and trimethoprim/sulfamethoxazole was administered for 10 days as an antibiotic. Animals were sacrificed at 3, 7, and 14 days postoperative.
Serum TNFα and IL6 measurements
Blood (∼1 mL) was extracted at 3, 7, and 14 days postoperative. After collection of whole blood, it remained at room temperature for 20 min, allowing it to clot. Clots were removed by centrifuging at 2000 g for 10 min in a refrigerated centrifuge. Serum TNFα and IL6 was measured using rat TNFα single analyte ELISArray kit (SER06411A; SABiosciences) and rat IL6 single analyte ELISArray kit (SER06481A; SABiosciences) as per manufacturers' instructions. The serum TNFα and IL6 concentrations were calculated based on standard calibration curves.
Histological and immunohistochemical analyses
Hematoxylin and Eosin (H&E) and immunohistochemical (IHC) stains were performed as previously described. 43 Briefly, tissue from the biceps femoris, vastus lateralis, and the femur and its overlying periosteum adjacent to and inclusive of the collagen sponge were harvested after sacrifice, fixed in formalin, and decalcified using Cal-Ex solution (Fisher Scientific) for 7 days. All staining was performed on frozen sections. Primary antibodies were used at a dilution of 1:100. Appropriate biotinylated secondary antibodies (Dako) were used at a dilution of 1:200. ABC complex (Vectastain Elite ABC Kit; Vector Laboratories, Inc., catalog no. PK-6100) incubation was followed by development with AEC chromagen (Dako; catalog no. K346911-2). Photomicrographs were acquired using Olympus BX51 (100× and 1000× magnification lens, UPLanFL; Olympus). The intensity of staining was analyzed using commercial software Image-Pro Plus 6 and quantified by the mean optical density of staining signal x per percent area positively stained ×100. 46 The software analyzed and averaged the results from six random fields per sample. For cell counting, eight fields (1000× magnification) from each sample of each group were randomly selected to evaluate for the presence of polymorphonuclear and mononuclear cells based on the morphology. Results were reported as the mean number of inflammatory cells per high power field.
Real-time PCR
To determine the local expression of TNFα and IL6 in rats, 1.5×2×1 cm tissue (4 rats/group) around the implanted sponges was dissected and homogenized in Trizol (Invitrogen). Total RNA extraction and real-time PCR were performed as previously described. 47 Real-time PCR was performed using the 7300 real-time PCR System instrument (Applied Biosystems). The reactions were incubated in 96-well optical plates at 95°C for 10 min, followed by 40 cycles at 95°C for 15 s, and at 60° for 10 s. The threshold cycle (Ct) data was determinate using default threshold settings. The Ct was defined as the fractional cycle number at which the fluorescence passes the fixed threshold. Mix primer sequences of mouse TNFα were sense: ATGAGCACAGAAAGCATGATC and anti-sense: TACAGGCTTGTCACTCGAATT; those of mouse IL6 were sense: ATGGATGCTACCAAACTGGAT and anti-sense: TGAAGGACTCTGGCTTTGTCT; those of rat TNFα were sense: TACTGAACTTCGGGGTGATCG and anti-sense: CCTTGTCCCTTGAAGAGAACC; those of rat IL6 were sense: AAATCTGCTCTGGTCTTCTGG and anti-sense: TTAGATACCCATCGACAGG; and primers for human TNFα were sense: TGAGGCCAAGCCCTGGTAT and anti-sense: CGAGATAGTCGGGCCGATT.
The relative expression level of the transcripts was compared to GAPDH. The primer sequences of the mouse GAPDH control were sense: CTCCCACTCTTCCACCTTCG and anti-sense: CCACCACCCTGTTGCTGTAG. The primer sequences of rat GADPH control were sense GCCTCGTCTCATAGACAAGATGG and anti-sense: TGCTCCTGCTACTTTAGACTCCG. The primer sequences of the human GAPDH control were sense: TGCACCACCAACTGCTTAGC and anti-sense: GGCATGGACTGTGGTCATGAG.
Transfection and luciferase assays
NIH3T3 and J774A.1 cells were plated at a density of 5×104 cells/well on 24-well plate. Transient transfection was performed using lipofectamine 2000 (Invitrogen) according to the instructions provided by the manufacturer. Cells were transfected using 4 μg of NFκB luciferase reporter vector and 0.5 μg of renilla luciferase reporter vector as a transfection control. Thus, the total amount of DNA for each transfection was kept constant. Luciferase activity was assessed using Dual-luciferase Reporter Assay System (Promega).
Measurement of ROS
The total ROS was detected by Total ROS Detection Kit (Enzo) and performed as per manufacturer's descriptions. Briefly, NIH3T3 cells were seeded at 2×105 cells/well on six-well plates in DMEM+10% FBS and starved overnight with DMEM+0.1% FBS. Cells were treated with BMP2 and NELL-1 at the indicated dose for 4 h before staining for ROS. Flow cytometry was used to detect the intensity of ROS and percentage of positive stained cells. 10,000 events were collected per sample. Samples not stained for ROS were used to quantify the average percentages of positively stained cells, serving as our negative control. DIVA version 6.1.3 was the software used to collect and analyze the data.
Statistical analyses
Means and standard deviations were calculated from numerical data. To allow multiple comparisons between groups, statistical analyses were performed using appropriate ANOVA followed by post-hoc Tukey's range test to directly test two different treatment groups.
Results
NELL-1 inhibits BMP2-induced local inflammation
The femoral onlay procedure was performed using a collagen sponge implant (Fig. 1). The procedure was relatively atraumatic, and did not result in perceptible swelling upon gross examination of soft tissues at site of implantation. This was in agreement with our previous observations. 48 Next, we examined the histology of implant sites treated with control carrier, BMP2 alone, NELL-1 alone, or a combination of NELL-1+BMP2 (see Table 1 for a detailed breakdown of treatment groups). Histological analysis was performed at 3, 7, and 14 days postimplantation to assess for local inflammation.

Schematic of femoral onlay model. Black rectangle shows the orientation of the histological images. Mu, muscle; Fs, femoral shaft. Color images available online at www.liebertpub.com/tea
H&E staining showed that control treatment (sponge with PBS) resulted in minimal inflammation with rare scattered inflammatory cells (Fig. 2A–F). In striking contrast, diffuse inflammation was observed with BMP2 treatment. At 3 days postimplantation, a dense polymorphonuclear infiltrate was observed in sponge and surrounding edematous soft tissue (Fig. 2G, H). At 7 days postimplantation, BMP2 treated samples showed a dense, mixed mononuclear and polymorphonuclear infiltrate in a background of exudative fibrous soft tissue (Fig. 2I, J). A persistent mononuclear infiltrate existed 14 days postimplanation with predominant macrophages and lymphocytes with scattered neutrophils was observed, on a background of newly formed osteoid (Fig. 2K, L).
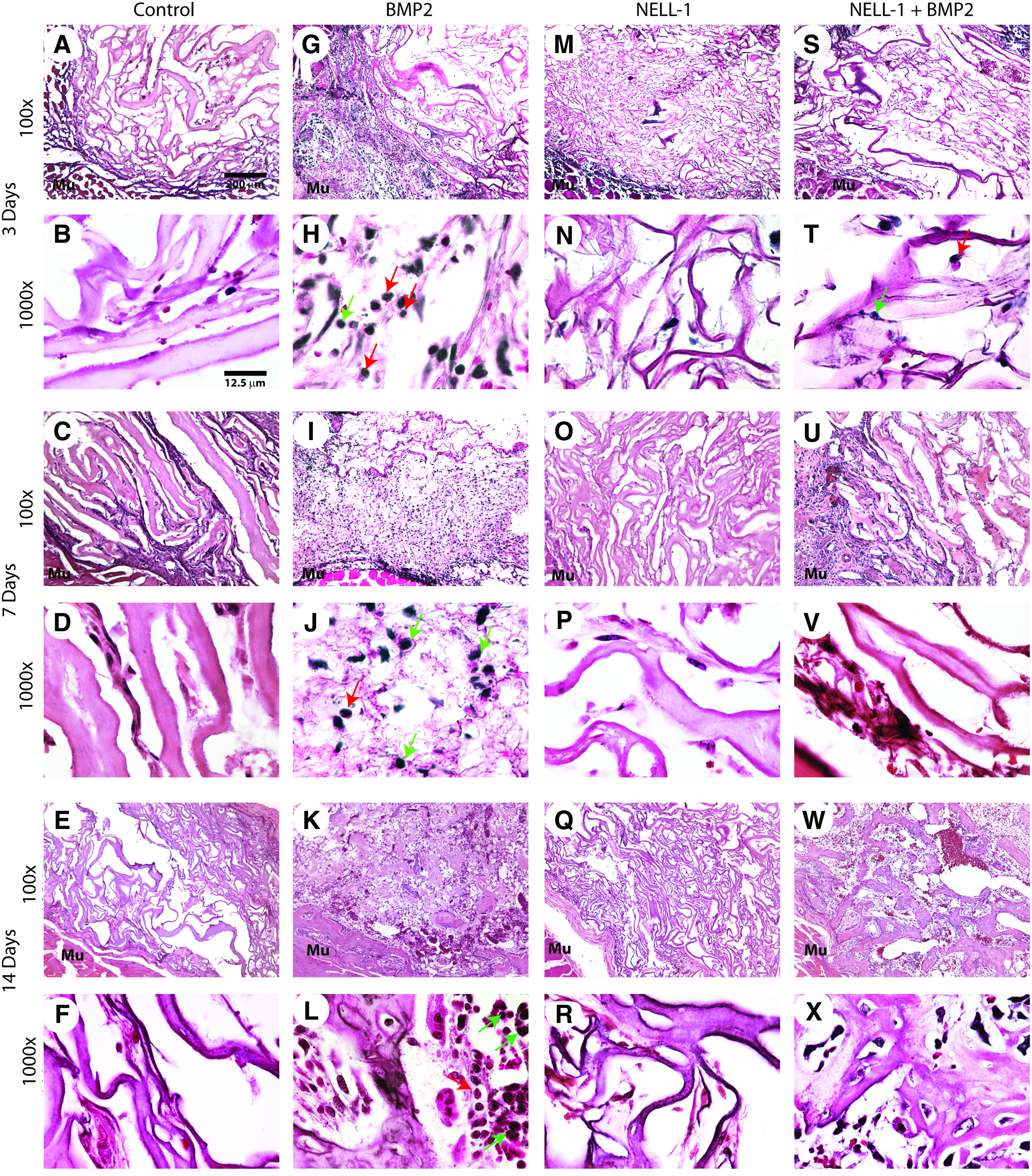
H&E staining of femoral onlay samples at 3, 7, and 14 days postoperative. The doses of BMP2 and NELL-1, and the number of animals in each group are shown in Table 1. At all time points studied, minimal inflammatory cell infiltration, and exudates were observed in control
Similar to the control treatment, treatment with NELL-1 alone resulted in minimal inflammation (Fig. 2M–R). After observing the group treated with NELL-1 14 days postoperative, it was concluded that NELL-1 did not induce ectopic bone formation (Fig. 2Q, R). This is consistent with the findings of previous studies,43,48 where the direct delivery of adenoviral NELL-1 to muscle did not cause ectopic bone formation, unlike the effects of BMP2. Additionally, in vivo ectopic bone formation was quantified using microCT (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/tea). Analysis of bone mineral density showed no significant difference across all groups (Supplementary Fig. S1A). Analysis of bone volume showed that treatment groups, including BMP2 significantly induced ectopic bone formation compared to control, while revealing no significant difference between BMP2 only and NELL-1+BMP2 groups (Supplementary Fig. S1B). Next, the inflammatory response from combination treatment of NELL-1+BMP2 was analyzed (Fig. 2S–X). At 3 days postimplantation, a striking reduction of inflammation was apparent in comparison to BMP2 treatment, with few neutrophils observed at high powered fields (Fig. 2S, T in comparison to Fig. 2G, H). This reduction in inflammation was observed throughout the 14 day study period. Abundant bone matrix was present at 14 days, and with only rare inflammatory cells present (Fig. 2W, X).
The qualitative observations of inflammation were next quantified, by counting the number of the inflammatory cells per high power field (Fig. 3). First, the presence of a polymorphonuclear infiltrate was analyzed and showed a significant increase in BMP2-treated samples at both 3 and 7 days postoperative (Fig. 3, left). This increase in acute inflammation was completely reversed by NELL-1. Next, the presence of a mononuclear inflammatory infiltrate was analyzed, and showed a statistically significant increase in BMP2-treated samples at 7 and 14 days (Fig. 3, right). This chronic inflammation was either completely or partially reversed by NELL-1 combination treatment. Thus and in summary, combination treatment with NELL-1+BMP2 was observed to mitigate the proinflammatory effects of BMP2 in the femoral onlay model.
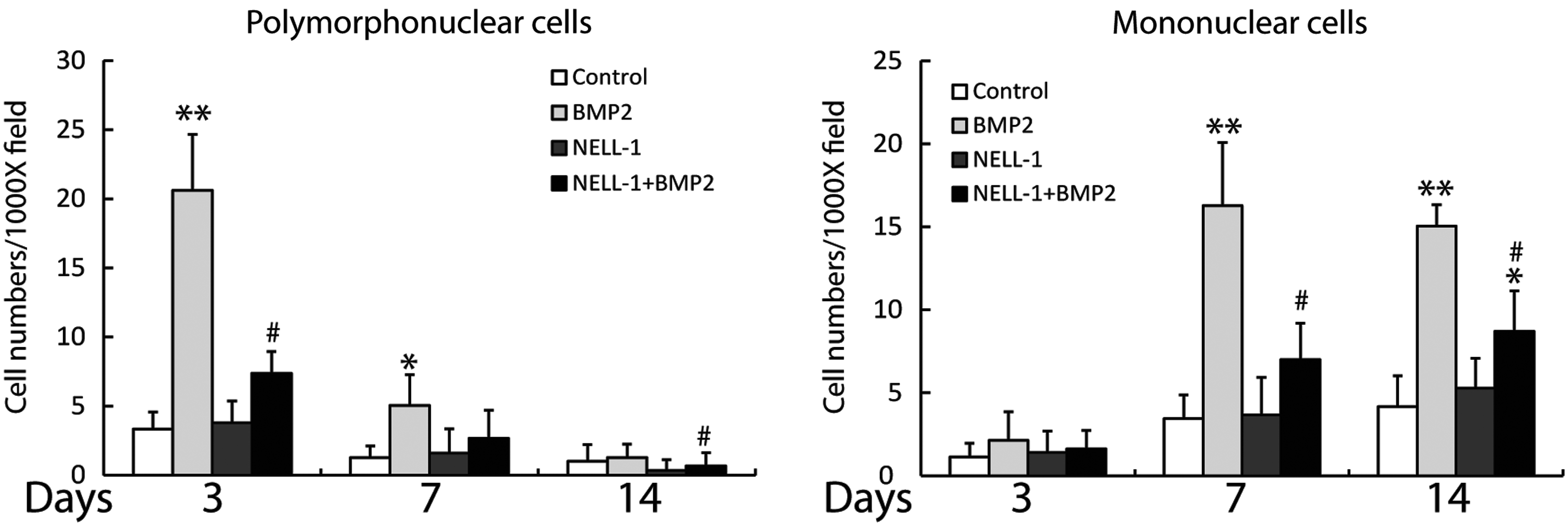
Quantification of inflammatory cell infiltrate in femoral onlay samples at 3, 7, and 14 days postoperative. Quantifications were based on eight random high power (1000×) fields obtained by four blinded observers. (Left) Polymorphonuclear cell number, representative of acute inflammation. (Right) Mononuclear cell number, representative of chronic inflammation. The doses of BMP2 and NELL-1, and the number of animals in each group are shown in Table 1. *p< 0.05, **p<0.01 compared to control. #p<0.05 compared to BMP2 alone.
NELL-1 inhibits BMP2-induced local inflammatory cytokine production
Next, we investigated the local expression of the proinflammatory cytokines TNFα and IL6 with BMP2 and/or NELL-1 treatment. IHC staining for TNFα confirmed our findings from H&E staining (Fig. 4). TNFα protein expression was dramatically induced at day 3 by BMP2 in comparison to control and staining was observed predominantly in areas of intense neutrophil infiltrate (Fig. 4A–F). In contrast, NELL-1 alone and NELL-1+BMP2 treatment groups both stained predominately negative for TNFα with focal areas of weak positivity (Fig. 4G–L). Semiquantification of TNFα immunostaining confirmed a significant induction by BMP2, which was reversed by NELL-1 protein addition (Fig. 4M). Local TNFα mRNA levels was also evaluated by microdissection and real-time PCR at 7 days (Fig. 6A). Similar to what has been seen from the IHC staining; BMP2 by itself significantly induced TNFα expression, which was inhibited by the addition of NELL-1.

TNFα immunohistochemical staining at 3, 7, and 14 days postoperative.
We next performed IHC staining for IL6, which revealed a similar pattern (Fig. 5). IL6 protein expression was dramatically induced by BMP2 in comparison to control, and remained persistently elevated throughout the 14 day study period (Fig. 5A–F). In contrast, NELL-1 alone treatment exhibited nearly entire negative staining for IL6 (Fig. 5G–I). Moreover, NELL-1 reversed BMP2 induction of IL6 expression, with only sporadic single cell staining throughout each sample (Fig. 5J–L). Again, semiquantification of immunostaining confirmed a significant induction by BMP2, which was reversed by NELL-1 protein addition (Fig. 5M). Similar results were seen by real-time PCR at 7 days postsurgery for local IL6 mRNA levels (Fig. 6B). In summary, BMP2 reliably induces an acute inflammatory response with an intensely TNFα+, IL6+ infiltrate in the femoral onlay model. In contrast, NELL-1 does not induce local inflammation in the same model and at similar dosages. Moreover, treatment with NELL-1 significantly attenuates the local TNFα, IL6 inflammatory infiltrate induced by BMP2.
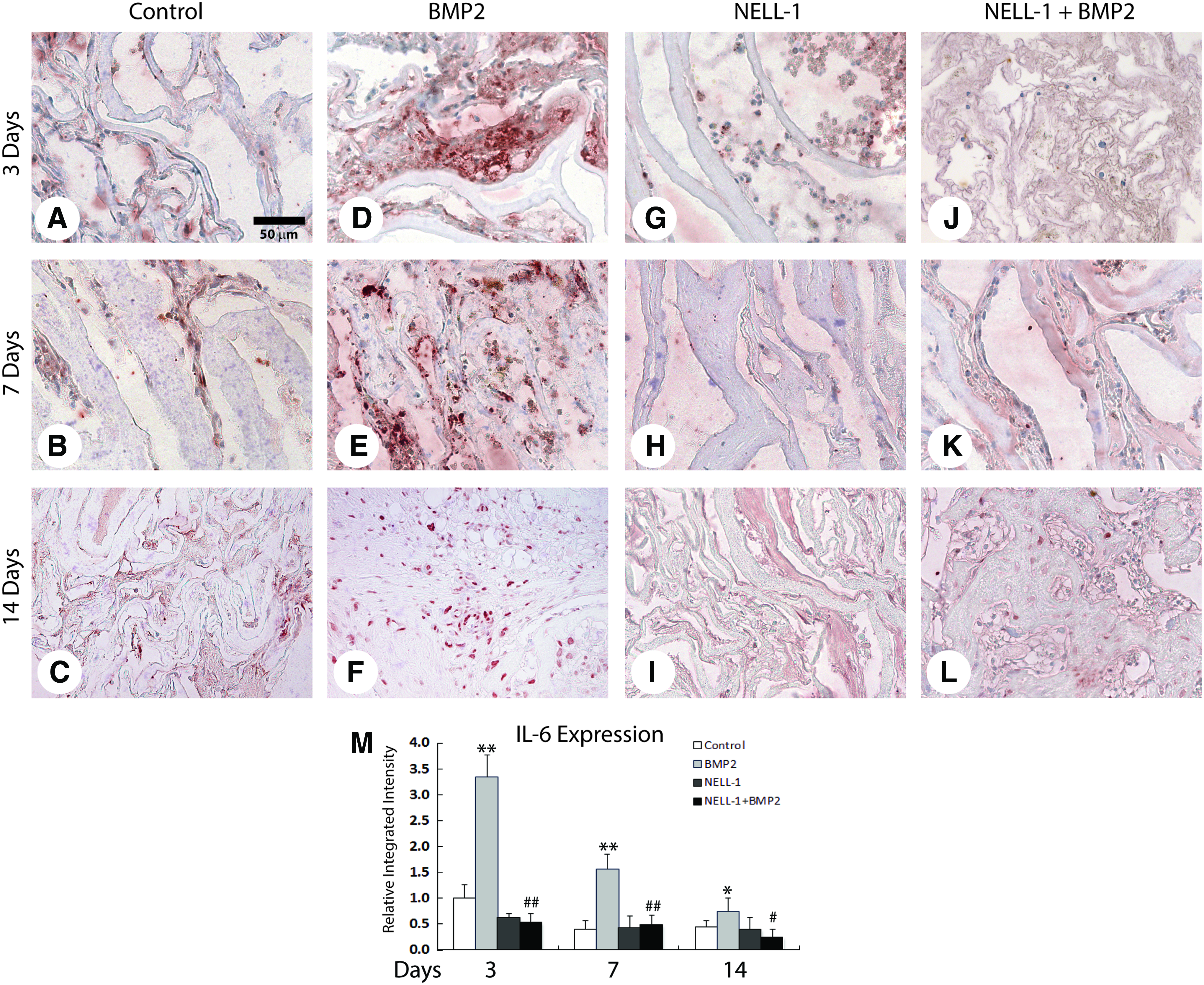
Interleukin 6 (IL6) immunohistochemical staining at 3, 7 and 14 days postoperative.
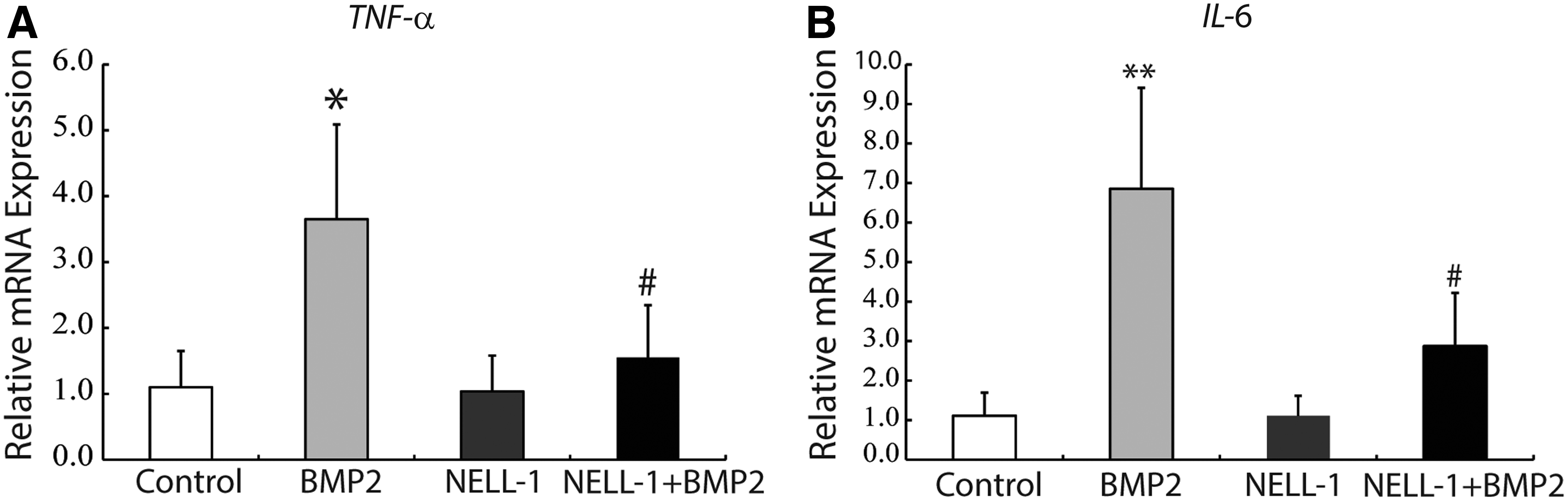
Local expression of TNFα and IL6 at 7 days postoperative by real-time PCR. 7 days after surgery, TNFα
NELL-1 inhibits BMP2-mediated increases in serum TNFα and IL6
To further investigate the proinflammatory effects of BMP2 and the extent to which this is reversed by NELL-1, we evaluated serum levels of TNFα and IL6 at 3,7, and 14 days postoperative by cytokine specific ELISA (Fig. 7). As before, BMP2 and/or NELL-1 were implanted overlying the femoral shaft and evaluation for systemic inflammation was performed at 3, 7, and 14 days postoperative. A significant, transient increase in TNFα and IL6 serum levels was observed with BMP2 treatment at 7 days postoperative, which normalized by 14 days. In contrast, NELL-1 alone did not induce an increase in TNFα or IL6 serum levels at any time point. Excitingly, NELL-1 significantly reduced BMP2-mediated increases of both TNFα and IL6 serum peak levels at 7 days (#p<0.05). Thus and in summary, in our femoral onlay model, treatment with BMP2 protein induced expression of inflammatory markers both locally and systemically. In contrast, NELL-1 protein significantly attenuated BMP2 induced inflammatory markers, again at both local and systemic levels.
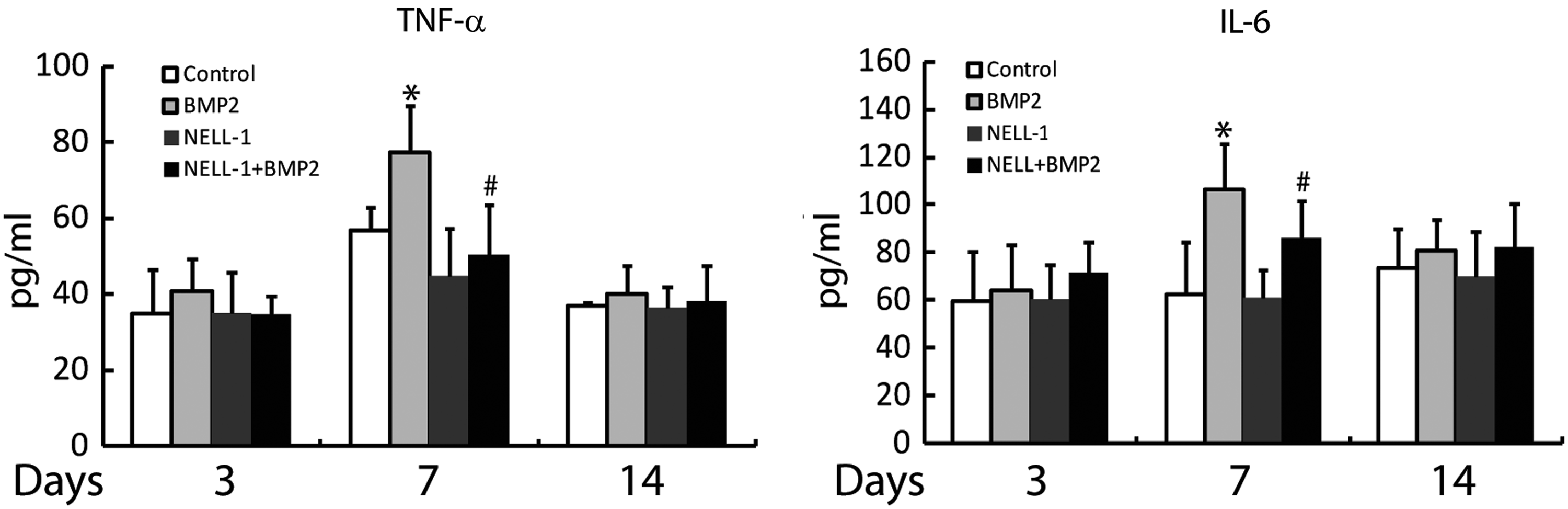
TNFα and IL6 serum levels at 3, 7, and 14 days postoperative. Serum level of TNFα and IL6 was measured by ELISA assays. BMP2 significantly increased serum levels of TNFα and IL6 at 7 days postoperation. This increase in cytokine levels was suppressed by combination treatment with NELL-1. N=5 animals per treatment group. *p<0.05 compared to control. #p<0.05 compared to BMP2 alone.
NELL-1 inhibits BMP2 induced inflammatory response in vitro in a ROS-dependent mechanism
Finally, we sought to understand the mechanisms, by which NELL-1 suppresses BMP2-induced inflammation by performing a series of in vitro studies. NIH3T3 cells, an established cell type for the study of BMP2-induced inflammation, 29 were studied after being treated with NELL-1±BMP2. In striking similarity to our in vivo findings, treatment with BMP2 alone significantly increased mRNA expression of inflammatory cytokines TNFα and IL6, in a dose-dependent manner (Fig. 8A, B). The inflammation induced by BMP2, however, was significantly suppressed in the combined treatment with NELL-1 (observed across all BMP2 test doses) (Fig. 8A, B). Additionally, we evaluated TNFα expression in HUVEC (Fig. 8C), J774A.1 macrophages (Fig. 8D), and C2C12 mouse myoblast cells (Fig. 8E). Results confirmed both that BMP2 treatment increases transcription of the proinflammatory cytokine TNFα, and that NELL-1 can significantly suppress this expression.
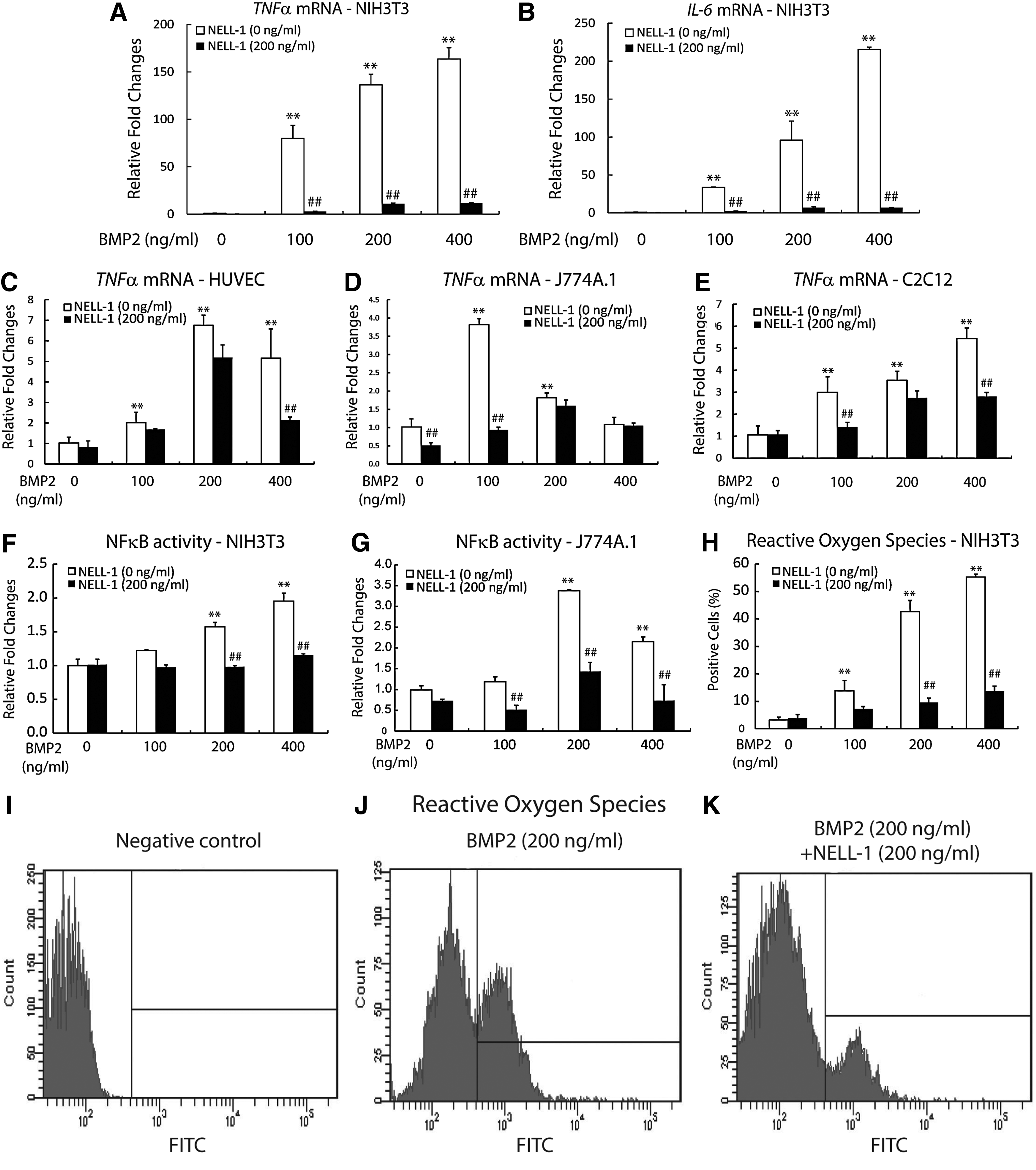
In vitro modulation of inflammation by NELL-1 and BMP2.
Considering NF-κB plays a key role in regulating immune and inflammatory responses, as well as being downstream of TNFα, 49 we next performed luciferase reporter assays using NF-κB promoter construct. Interestingly, NELL-1 suppressed BMP2-induced NF-κB transcriptional factor activity in NIH3T3 (Fig. 8F) and J774A.1 (Fig. 8G) cells. We then investigated ROS, which are crucial intracellular signaling molecules that trigger various transduction pathways, including inflammation. 50 We assessed ROS generation in NIH3T3 cells treated with NELL-1±BMP2. As documented in previous studies in other cell types,29,30 BMP2 increased ROS generation in a dose-dependent manner. Interestingly, NELL-1 significantly reduced ROS generation induced by BMP2 (Fig. 8H–K). In summary, NELL-1 attenuates BMP2 induced inflammation, associated with decreased transcription of TNFα and IL6, decreased NF-κB activity, and reduced generation of ROS.
Discussion
Although clinical trials have reported successful use of BMP2 for bone formation in open fractures of long bones, nonunions, and spinal fusion, there is a growing body of literature regarding clinical complications associated with the use of BMP2.7,27,51,52 The most worrisome (and potentially life threatening) complication is BMP2-induced local inflammation. Indeed, increased inflammation caused by BMP2 has a lengthy complication list, of which symptoms include neck-swelling resulting in dysphagia, and possible respiratory failure due to cervical inflammation.51,53,54 Additionally, radiculopathies and seroma formations have been reported in lumbar spinal fusions, due to inflammation induced by BMP2 to surrounding muscle and nervous tissue.18,20 Thus far, investigated methods to reduce BMP2 induced side effects, notably inflammation, have been limited to the administration of lower doses of BMP2,55,56 the prevention of BMP2 diffusion into surrounding tissue,57,58 and the administering of anti-inflammatory molecules like corticosteroids or BMP2 binding peptides.20,51,56,59–61 In this study, we successfully demonstrated that NELL-1 significantly suppressed BMP2-induce inflammation, including acute and chronic inflammatory cell infiltration, inflammatory cytokine production, soft tissue exudate, and even systemic levels of inflammatory markers. Notably, the osteoinductive potency of BMP2 was not diminished by addition of NELL-1 evidenced with osteoid and woven bone formation at site of implantation in comparison to BMP2 alone group (Fig. 2L, X). These findings were recapitulated in vitro using NIH3T3 fibroblasts, HUVEC, and C2C12 myoblasts, again finding that NELL-1 inhibited BMP2-induced inflammation. This was associated with a reduction in ROS generation, indicating one possible mechanism. In summary, our findings demonstrate the osteogenic potential of NELL-1+ BMP2 combination treatment for bone regeneration, whilst preventing the adverse effect of BMP2-induced inflammation.
Our finding that BMP2 induces a pronounced local inflammation is in agreement with the available published literature.21,22,28 Although most studies focus solely on osteogenesis and bone formation, it has been thoroughly demonstrated that BMP2 possesses inflammatory effects in addition to its osteoinductive properties.18–21 Here we observed that BMP2 induces both acute and chronic inflammation, which is associated with increased expression of the proinflammatory cytokines TNFα and IL6. In vitro, BMP2 elicited substantial increases in generation of ROS and induction of NFκB activity. While literature suggests that BMP2 reliably induces inflammation, the manner by which NELL-1 prevents BMP2-induced inflammation is only partially elucidated. In the current study, we found that NELL-1 was able to inhibit BMP2-induced ROS generation. Since the cysteine redox status determines the level of inflammatory cytokines 62 and NELL-1 has a high ratio of cysteine composition (11.1%), the cysteine-rich composition of NELL-1 is currently being investigated as a potential reductive agent on the ROS, preventing the proinflammatory response.
In addition to preventing the inflammatory response induced by BMP2 treatment, NELL-1 possesses anti-adipogenic properties. 32 For example, in 3T3-L1 preadipocytes, NELL-1 directly reduces adipogenic gene expression (Pparg, Lpl, Ap2) and reduces lipid droplet accumulation. 32 This phenomenon has also been reported in primary human preadipocytes (adipose-derived stromal cells). 63 Interestingly and conversely, the proadipogenic effects of BMP2 have been well described both in vitro and in vivo.64,65 Indeed our research group previously found that high-dosages of BMP2 induced formation of cyst-like bony shells filled with adipose tissue in a rodent femoral segmental defect model. 26 It is interesting to speculate that the combination treatment of NELL-1 with BMP2 may mitigate this adipogenic phenotype.
Collectively, the findings of the present study suggest for the first time that NELL-1 has anti-inflammatory properties, and that NELL-1 may mitigate the adverse effect of high dose BMP2-induced inflammation, while potentially enhancing its osteoinductivity. These findings were observed in an in vivo femoral onlay model, with the reduction of proinflammatory markers TNFα and IL6, and recapitulated in vitro using NIH3T3 fibroblasts, HUVEC, and C2C12 myoblasts. This inhibition of BMP2-induced inflammation was associated with reduction of ROS production. Further studies are required to fully understand the mechanism behind the anti-inflammatory effects of NELL-1; however, current findings suggest promise for the combined use of NELL-1 and BMP2 protein for future efforts in bone tissue engineering.
Footnotes
Acknowledgments
This work was supported by the CIRM Early Translational II Research Award TR2-01821, NIH/NIDCR (grants R21 DE0177711, RO1 DE01607 and RO1 AR061399-01A1), UC Discovery Grant 07-10677, Eli & Edythe Broad Center of Regenerative Medicine and Stem Cell Research at UCLA Innovation Award. A.W.J. was supported by T32 training fellowships (5T32DE007296-14).
Disclosure Statement
Drs. K.T, and C.S. are inventors of NELL-1 related patents. Drs. K.T, and C.S are founders of Bone Biologics Inc., which sublicenses NELL-1 patents from the UC Regents.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.