
Abstract
Human mesenchymal stromal cells (hMSCs) offer great potential for bone tissue engineering applications, but their in vivo performance remains limited. Preconditioning of these cells with small molecules to improve their differentiation before implantation, or incorporation of growth factors are possible solutions. Insulin-like growth factor-1 (IGF-1) is one of the most abundant growth factors in bone, involved in growth, development, and metabolism, but its effects on hMSCs are still subject of debate. Here we examined the effects of IGF-1 on proliferation and differentiation of hMSCs in vitro and we found that serum abolished the effects of IGF-1. Only in the absence of serum, IGF-1 increased proliferation, alkaline phosphatase expression, and osteogenic gene expression of hMSCs. Furthermore, we examined synergistic effects of bone morphogenetic protein-2 (BMP-2) and IGF-1 and, although IGF-1 enhanced BMP-2-induced mineralization, IGF-1 only slightly affected in vivo bone formation.
Introduction
Insulin-like growth-factor 1 (IGF-1 or somatomedin C) is one of the most abundant growth factors in the bone matrix, involved in growth, development, and metabolism. It is produced locally as well as systemically by the liver and its biological actions are regulated by the acid-labile subunit (ALS) and six binding proteins (IGF binding proteins [IGFBPs]), which can either enhance or inhibit its actions by preventing access to the IGF-1 receptor (IGF-1R). 7 Thus, systemic production of IGF-1 by the liver is regulated by growth hormone (GH) and this system is referred to as the GH/IGF-1 axis.8,9 Locally, IGF-1 is produced by a variety of cell types, which is regulated not only by GH but also by numerous other factors.
Several mouse models have demonstrated a key role of IGF-1 in bone formation.10,11 Complete disruption of IGF signaling, by deletion of the IGF-1R results in a reduction of cancellous bone volume, connectivity and trabecular number and increases trabecular spacing. 12 Interestingly, these mice display a dramatic decrease in mineralization, indicating an essential role for IGF-1 in the development of mature mineralized tissue. 12 IGF-1 null mice have reduced cortical bone size and femur length; although an increase in trabecular bone density and connectivity were also observed. 13 Impaired osteoclastogenesis may explain the latter findings, in line with in vitro studies showing that IGF-1 promotes osteoclastogenesis.13,14 Local overexpression of IGF-1 in osteoblasts resulted in increased bone mineral density and trabecular bone volume 15 and, since no increase in osteoblast number was observed, the effects of IGF-1 were suggested to be mediated via enhanced osteoblast function.
Due to its positive effect on osteoblasts, its presence at fracture sites and its involvement in the synthesis of bone matrix, 16 IGF-1 has gained interest as a therapeutic factor in fracture healing; either alone or in combination with other growth factors. In vitro IGF-1 was shown to enhance proliferation of C2C12 cells, osteoblasts, 17 osteoblast-like cells, 18 and periodontal ligament fibroblasts. 19 It also induces alkaline phosphatase (ALP) activity, osteocalcin, matrix calcium content, and nodule formation in osteoblasts 6 and has been proven as a chemotactic factor for these cells. 1 Furthermore, in fetal rat calvariae, IGF-1 was shown to enhance osteocalcin secretion and incorporation of proline into collagen type I 2 and to decrease collagen degradation. 3
In hMSCs, similar effects of IGF-1 on migration were shown,4,5 but the effects of IGF-1 on proliferation and differentiation are less clear. Some studies show that IGF-1 does not affect hMSCs at all,20,21 whereas on the other hand, MSCs transduced with an IGF-1 adenovirus displayed increased runx2 and collagen type I expression, but not ALP. 22 Also, incorporation of IGF-1 in scaffolds enhanced proliferation and differentiation of attached hMSCs and periodontal ligament fibroblasts.23,24 In contrast, it has been reported that IGF-1 stimulates chondrogenic differentiation of hMSCs in the presence of transforming growth factor-beta (TGF-β), 25 which was also demonstrated by adenoviral overexpression of IGF-1 and BMP-2 in adipose-derived MSCs. 26 In our own lab, we have shown that treatment of hMSCs with the small molecule db-cAMP enhances in vivo bone formation, which is accompanied by increased secretion of bone specific growth factors, such as BMP-2, but also IGF-1. 27 Conditioned medium from these treated cells increased proliferation of various cell types; thus, indicating a trophic effect, 28 but it is unknown if secreted IGF-1 can affect the implanted hMSCs.
In this study, we examined the effects of IGF-1 on proliferation and differentiation of hMSCs, both in the presence and absence of serum, which may contain proteins that neutralize IGF-1 function. In addition, with an adenoviral transduction system, we investigated if IGF-1 has effects on in vivo bone forming capacity of hMSCs, either alone or in combination with BMP-2.
Materials and Methods
Cell culture
Bone marrow aspirates (5–20 mL) were obtained from donors with written informed consent. hMSCs were isolated and proliferated as described previously. 29 Briefly, aspirates were resuspended using 20 G needles, plated at a density of 5×105 cells/cm2 and cultured in hMSC proliferation medium containing α-minimal essential medium (α-MEM; Life Technologies), 10% fetal bovine serum (FBS; Cambrex), 0.2 mM ascorbic acid (Asap; Life Technologies), 2 mM L-glutamine (Life Technologies), 100 U/mL penicillin (Life Technologies), 10 μg/mL streptomycin (Life Technologies), and 1 ng/mL basic fibroblast growth factor (bFGF; Instruchemie). Cells were grown at 37°C in a humid atmosphere with 5% CO2. Medium was refreshed twice a week and cells were used for further sub-culturing or cryopreservation upon reaching near confluence. For osteogenic experiments, hMSCs were cultured in basic medium, composed of hMSC proliferation medium without bFGF. The osteosarcoma cell line MG-63 was expanded and cultured in basic medium as described above. Recombinant human IGF-1 (rhIGF-1) was obtained from R&D Systems and used in concentrations between 10 and 150 ng/mL. Recombinant human bone morphogenetic protein-2 (rhBMP-2) was obtained from Biodoor Biotechnology Co., Ltd. and used at a concentration of 100 ng/mL, unless stated otherwise.
Proliferation
hMSCs or MG-63 cells were seeded in triplicate in 24-well plates at 2000 cells/cm2 and allowed to attach for 10 to 15 h in proliferation medium or basic medium, respectively. Then, the medium was changed to basic medium with or without FBS with various concentrations of rhIGF-1. After 5 days, an Alamar Blue assay (BioSource) was performed. A 10% v/v solution of Alamar Blue in medium was prepared and 1 mL of solution was added to each well. After 4 h, the fluorescence intensity of the solution in each well was determined by measuring 200 μL of solution at an excitation wavelength of 545 nm and an emission wavelength of 590 nm (Victor; Perkin Elmer).
ALP expression
hMSCs were seeded in triplicate in six-well plates at 5000 cells/cm2 and allowed to attach for 10 to 15 h in proliferation medium. Each experiment was performed in triplicate with a negative control (cells grown in basic medium) and one or more experimental conditions (with or without FBS, various concentrations of IGF-1). After 4 days, cells were trypsinized and incubated for 30 min in block buffer (phosphate-buffered saline [PBS] with 5% bovine serum albumin [BSA; Sigma] and 0.01% NaN3), and then incubated with primary antibody (anti-ALP, B4-78 [Developmental Studies Hybridoma Bank, University of Iowa]) diluted in wash buffer (PBS with 1% BSA and 0.05% NaN3) for 1 h or with isotype control antibodies. Cells were then washed thrice with wash buffer and incubated with secondary antibody (rat anti mouse IgG PE; Dako) diluted in wash buffer for 30 min. Cells were washed thrice and resuspended in 250 μL wash buffer with 10 μL Viaprobe (Pharmingen) for live/dead cell staining. Only living cells were used for further analysis. With the isotypes set to zero, the amount of ALP-positive cells were analyzed on a FACS Caliber using cell-quest software (Becton Dickinson Immunocytometry Systems). ALP expression was then calculated relative to the respective negative control.
ALP activity
MG-63 cells were seeded in triplicate in 12-well plates at 5000 cells/cm2 and allowed to attach for 10–15 h. The next day, medium was changed to medium with or without FBS, with various concentrations of IGF-1. As a negative control, cells were cultured in basic medium. After 4 days of culture, an Alamar Blue assay was performed. Then, cells were lysed with CDP star lysis buffer and the lysate was used for a CDP star assay (Roche). Briefly, 30 μL of CDP star substrate was incubated with 10 μL of cell lysate for 30 min after which luminescence was measured (Victor; Perkin Elmer). ALP expression was normalized to metabolic activity, and data is depicted as percentage relative to control.
Calcium deposition
Cells were seeded in triplicate in six-well plates at 5000 cells/cm2 and allowed to attach for 10 to 15 h in proliferation medium. Then, either mineralization medium alone (composed of basic medium supplemented with 10−8 M dex and 0.01 M β-glycerophosphate [Sigma]) or mineralization medium supplemented with 75 ng/mL BMP-2, 100 ng/mL IGF-1 or a combination of both was added and cells were kept in culture for 14–21 days. Then, culture medium was aspirated, cells were washed twice with calcium- and magnesium-free PBS (Life Technologies) and incubated overnight with 0.5 M HCl on an orbital shaker at room temperature. Then the supernatant was collected and the amount of calcium was determined using a calcium assay kit (Quantichrom calcium assay kit; Gentaur), by measuring the absorbance at 620 nm (Tecan). Data are depicted as dg calcium per mL HCl.
Gene expression analysis
Cells were seeded in triplicate in six-well plates at 10,000 cells/cm2 in proliferation medium. The next day, medium was replaced with basic medium with or without serum, and 100 ng/mL IGF-1 was added to denoted conditions. RNA was isolated after 24 h, 48 h, 72 h, 5 days, and 10 days, using a Bioke RNA II nucleospin RNA isolation kit (Machery Nagel). For gene overexpression experiments, cells were kept in culture for 2 or 5 days after medium change and RNA was isolated. RNA concentrations were measured using an ND100 spectrophotometer (Nanodrop Technologies) and cDNA was synthesized from 100 ng of RNA, using iScript (BioRad) according to the manufacturer's protocol. For quantitative polymerase chain reaction (PCR), a master mix, containing distilled water, forward primer, reverse primer (Sigma Genosys), BSA, and SYBR green I mix (all from Invitrogen) was prepared. Real-time qPCR was performed in a Light-Cycler (Roche). Light-Cycler data were analyzed using the fit points method of Light-Cycler software. 30 The baseline was set at the lower log-linear part above baseline noise and the crossing temperature (Ct value) was determined. Ct values were normalized to the 18S housekeeping gene and ΔCt (Ct, control−Ct, sample) was used to calculate the relative increase in gene expression. Primer sequences are listed in Table 1.
ALP, alkaline phosphatase.
Confocal Raman microspectroscopy
UV grade CaF2 slides for Raman (Crystran Ltd.) were sterilized using 70% ethanol and subsequently coated with proteins by incubation with FBS for 30 min. Then, hMSCs were seeded in triplicate at 5000 cells/cm2 and allowed to attach overnight in proliferation medium. The next day, medium was replaced with mineralization medium, with and without IGF-1 (100 ng/mL) and BMP-2 (100 ng/mL). After 28 days of culture, cells were fixed using 10% formalin and Raman spectra were acquired. A krypton ion laser (Innova 90-K; Coherent, Inc.) emitting at 647.1 nm was used as an excitation source. A 63x/1.0NA water-dipping objective (Zeiss W-plan Apochromat; Carl Zeiss MicroImaging) was used to focus the laser light over the sample and the Raman scattered photons were collected and dispersed on an air-cooled electron-multiplying charge-coupled device (EMCCD: Newton DU-970N; Andor Technology). An excitation power of 35mW on the sample was used. Raman spectra were acquired from 15 randomly chosen cells in each culture condition.
Scanning electron microscopy
Cells were seeded in triplicate in six-well plates at 5000 cells/cm2 in proliferation medium. The next day, medium was replaced with basic medium with or without 100 ng/mL IGF-1 and/or 100 ng/mL BMP-2. After 10 and 17 days of culture, cells were fixed using 10% formalin. Samples were dehydrated through a graded series of ethanol (70%–100%), critical point dried using CO2 (CPD 030; Balzers), and sputtered with gold (Cressington). Scanning electron microscopy (SEM) images were obtained using a scanning electron microscope (XL30 ESEM; Philips) at an accelerating voltage of 5 or 10 kV.
Adenoviral transductions
Adenoviruses containing an expression vector for IGF-1 (adIGF-1), BMP-2 (adBMP-2) or green fluorescent protein (adGFP, control) were purchased from Biofocus. For adenoviral transductions, hMSCs were seeded at 15,000 cells/cm2 and allowed to attach overnight. The next day, cells were transduced in basic medium using a previously optimized (data not shown) multiplicity of infection (MOI) of 1000 for adIGF-1 and adGFP and an MOI of 250 for adBMP-2 for 24 h, after which the medium was replaced with fresh medium. Transduced cells were then used for further in vitro experiments or, in case of the in vivo experiments, trypsinized, and seeded onto ceramic scaffolds for implantation.
Secretion of IGF-1 and BMP-2
hMSCs were seeded and transduced as described above. After the medium change, every 3 days medium was collected and replaced with fresh medium for a total period of 15 (control for in vitro experiments) or 30 (control for in vivo experiment) days. The concentrations of IGF-1 and/or BMP-2 in the medium were determined using ELISA (human IGF-1/BMP-2, Quantikine; R&D Systems), according to the manufacturer's protocol.
In vivo bone formation
About 1–2 mm biphasic calcium phosphate (BCP) particles were prepared and sintered at 1150°C as described previously. 31 hMSCs were seeded and transduced as described above. The next day, cells were trypsinized and seeded onto BCP scaffolds with 200,000 cells/four particles in basic medium and allowed to attach overnight. 20 min before surgery, nude female mice (Hsd-cpb:NMRI-nu; Harlan) were injected intraperitoneally with avertin. Particles were implanted in subcutaneous pockets, with 4 particles per pocket, 4 pockets per mouse, and 10 samples per condition. Incisions were closed using surgical staples and after 6 weeks the mice were sacrificed by cervical dislocation. Particles were explanted, fixed in 4% paraformaldehyde (Merck), dehydrated, and embedded in methyl methacrylate (L.T.I. Bilthoven) for sectioning. Undecalcified sections were processed on a histological diamond saw (Leica) and sections were stained with methylene blue and basic fuchsin to visualize bone formation. Quantitative histomorphometry was performed by pseudocoloring (bone in green, scaffolds in red) scanned images of high-resolution (7200 dpi) micrographs (Pathscan Enabler IV), after which the percentage of bone per scaffold area was determined using the pixel count option in Photoshop CS5 (Adobe).
Masson's Trichrome staining
hMSC/BCP constructs were firstly fixed in 4% formaldehyde at 4°C overnight before being decalcified in EDTA/PBS (pH 7.5) for 4 weeks. The decalcified implants were subsequently paraffin embedded and sectioned at 6μm. For Masson's Trichrome analysis, the sections were deparaffinised in Histoclear™ (Laborimpex), then methanol and rinsed with distilled water. The slides were then immersed in Weigerts iron hematoxylin solution for 10 min before rinsing in warm running tap water for 10 min. The sections were subsequently stained with ponceau-acid fuchsin solution (0.5% ponceau BS, 0.5% acid fuchsin in 10% acetic acid) for 15 min, washed in distilled water and differentiated in 5% phosphomolybdic acid 5% phosphotungstic acid solution for 15 min. The sections were then transferred directly to Masson's green solution (Klinipath) for 10 min before differentiation for 5 min in 1% acetic acid. The sections were then rinsed in distilled water, dehydrated through graded alcohol before being cleared in Histoclear for 2 min and mounted in Pertex.
Statistics
Data were analyzed in SPSS (PASW statistics) using one-way ANOVA followed by Tukey's multiple comparison test (p<0.05).
Results
IGF-1 increases proliferation of hMSCs and MG-63 cells in the absence of serum
To investigate the effect of IGF-1 on proliferation of hMSCs, cells were cultured in the presence of 10–150 ng/mL rhIGF-1 for 4 days, after which metabolic activity, as a measure for cell number, was determined. Since IGF-1 was previously shown to enhance proliferation of human osteoblasts in serum-free medium and the MG-63 cell line is frequently used as a model system for these cells, we used this cell line as a control. None of the used IGF-1 concentrations affected proliferation of MG-63 cells or hMSCs (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/tea) in culture medium containing FBS. In contrast, when FBS was removed from the medium, a clear dose-dependent increase in proliferation was observed for both cell types, although MG-63 cells showed a stronger response (50% increase in cell number) than hMSCs (20% increase in cell number) (Fig. 1a).
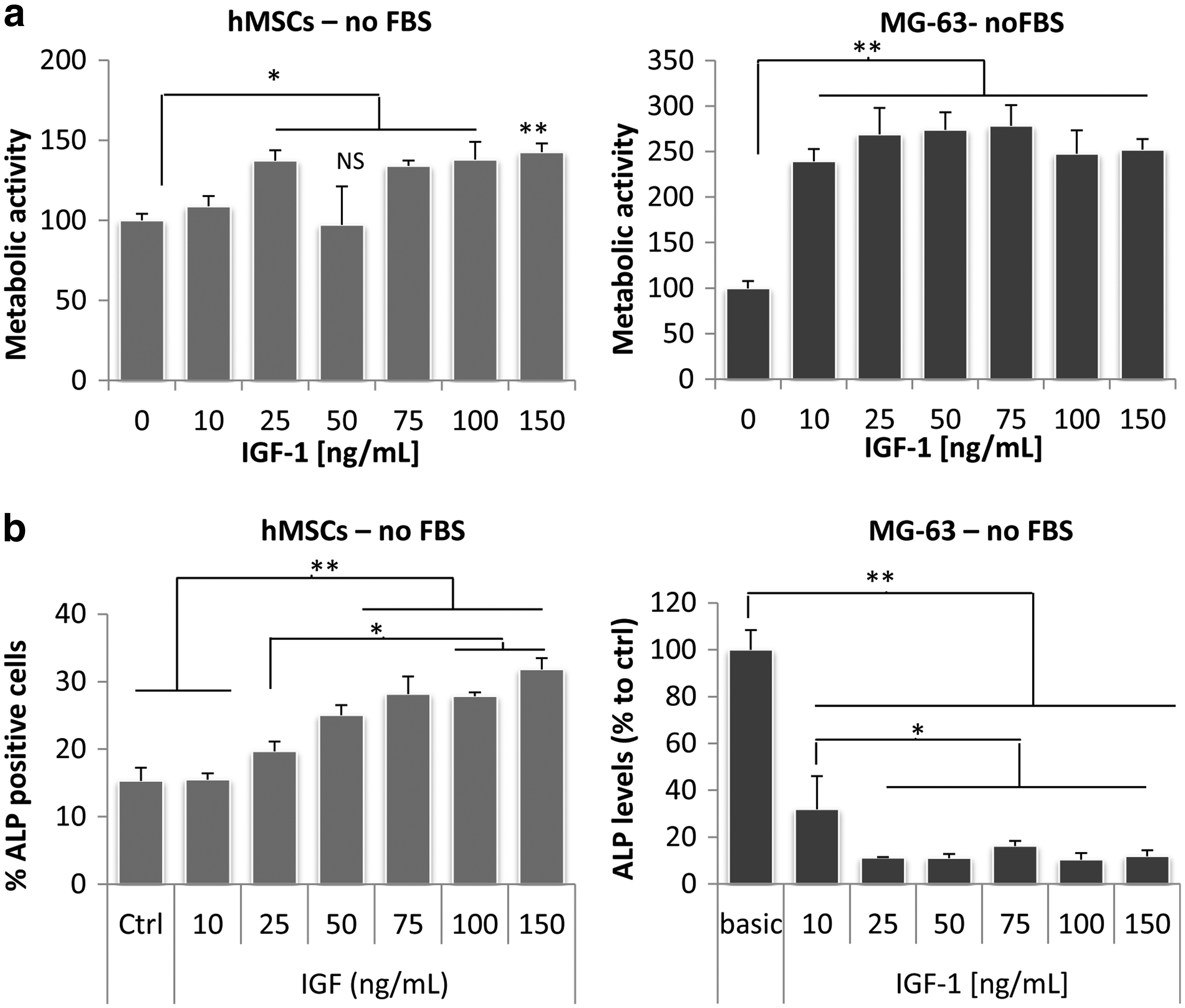
Insulin-like growth factor-1 (IGF-1) increases proliferation and differentiation of human mesenchymal stromal cells (hMSCs) and MG63 cells, but only in the absence of fetal bovine serum (FBS). hMSCs and MG63 cells were cultured in the presence of various concentrations recombinant human IGF-1 (rhIGF-1) and proliferation and alkaline phosphatase (ALP) activity were assessed after 4 days. When FBS was supplemented to the medium, IGF-1 did not affect proliferation (data not shown), but in the absence of FBS, IGF-1 significantly increased cell number
IGF-1 enhances differentiation of hMSCs in the absence of serum but decreases ALP expression of MG-63 cells
To investigate if IGF-1 also has a positive effect on osteogenic differentiation of these cells, hMSCs and MG-63 cells were cultured in the presence of rhIGF-1 with concentrations ranging from 10 to 150 ng/mL and ALP levels were determined. Again, in the presence of FBS there was no effect of IGF-1 (Supplementary Fig. S2), but in the absence of FBS, a dose-dependent increase in ALP expression was observed (Fig. 1b). In line with this, treatment of hMSCs with 100 ng/mL rhIGF-1 for a period of 1–10 days, resulted in an increase in the expression of bone marker genes, such as osteonectin, collagen type I, ALP. Runx2 expression was increased, but this was not significant and osteocalcin and S1004A were not affected, as demonstrated by qPCR (Fig. 2a). Again, IGF-1 did not induce expression of osteogenic genes when FBS was supplemented to the medium (data not shown). In contrast, ALP expression of MG-63 cells was significantly decreased with all concentrations of IGF-1, both in FBS-supplemented and FBS-free medium, although in the latter the effect was clearly stronger (Supplementary Fig. S2 and Fig. 1b). qPCR on MG-63 cells, using a concentration of 100 ng/mL rhIGF-1, confirmed the effect on ALP gene expression, although at day 10, ALP expression was slightly higher in IGF-1 treated cells. Expression of other osteogenic markers, such as runx2, collagen type I, and osteonectin, was also increased by IGF-1 at later timepoints (day 5–10), whereas there was no change in expression of osteocalcin (Fig. 2b), and S1004A expression was decreased. These data show that IGF-1 increases osteogenic differentiation of both MG-63 cells and hMSCs.
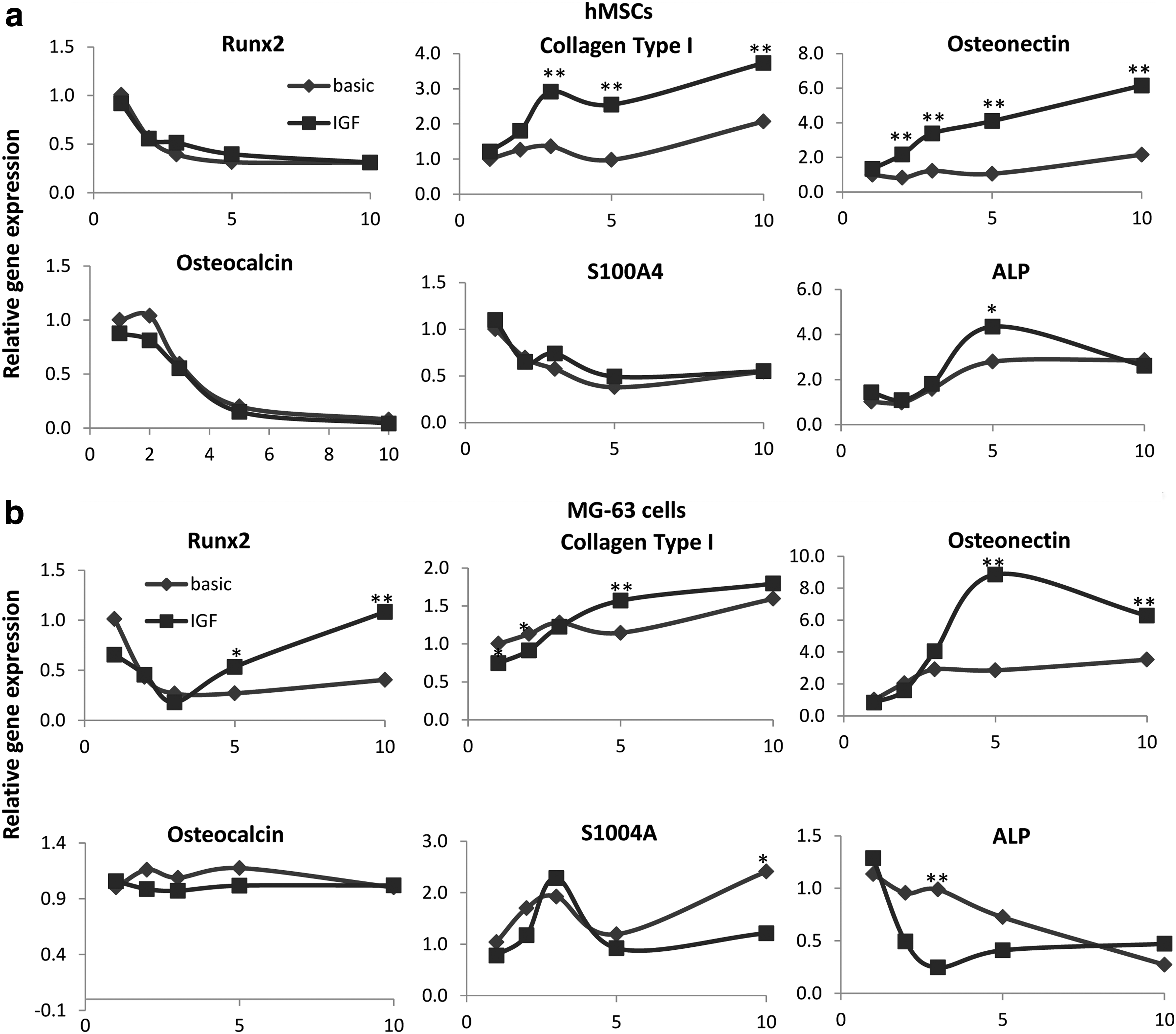
IGF-1 induces expression of osteogenic genes in hMSCs and MG-63. hMSCs and MG-63 cells were cultured in the presence of 100 ng/mL rhIGF, but in the absence of FBS for 1–10 days, after which gene expression levels were analyzed by quantitative polymerase chain reaction. IGF-1 significantly increased gene expression of Collagen type I, osteonectin, and ALP and non-significantly increased runx2 expression in hMSCs. No effect was found on osteocalcin and S100A4
IGF-1 enhances BMP-2-induced mineralization of hMSCs
In mouse knock-out models, IGF-1 was suggested to correlate with matrix formation, and therefore, we examined the effect of IGF-1 on mineralization of hMSCs. Previously, BMP-2 has been demonstrated to increase mineralization of hMSCs, which was confirmed here for concentrations of 75 and 100 ng/mL (Fig. 3a), and therefore, we also examined synergistic effects between these two growth factors. hMSCs were cultured in mineralization medium supplemented with 75 ng/mL BMP-2, 100 ng/mL IGF-1 or both. After 14 days, neither BMP-2 nor IGF-1 increased calcium deposition, but, compared to control, combined treatment resulted in significant higher amounts of calcium (Fig. 3b). After 21 days, a slight increase in calcium deposition was observed using BMP-2 or IGF-1 alone, but this was not significant. In contrast, addition of both factors resulted in a synergistic increase in calcium deposition, suggesting an interaction between these two factors. To investigate the mechanism behind the effects of IGF-1, mineralized samples were examined using Raman spectroscopy and SEM. Figure 3c shows Raman spectra after 28 days of culture. The calcium phosphate peak (961 cm−1) observed in cells cultured with BMP-2 and BMP-2/IGF-1 was higher compared to control, but no differences were observed between BMP-2 and BMP-2/IGF-1. Similarly, SEM images showed large amounts of mineralized nodules in both BMP-2 and BMP-2/IGF-1 groups, but we did not find differences in size or frequency of the nodules (Fig. 3d).
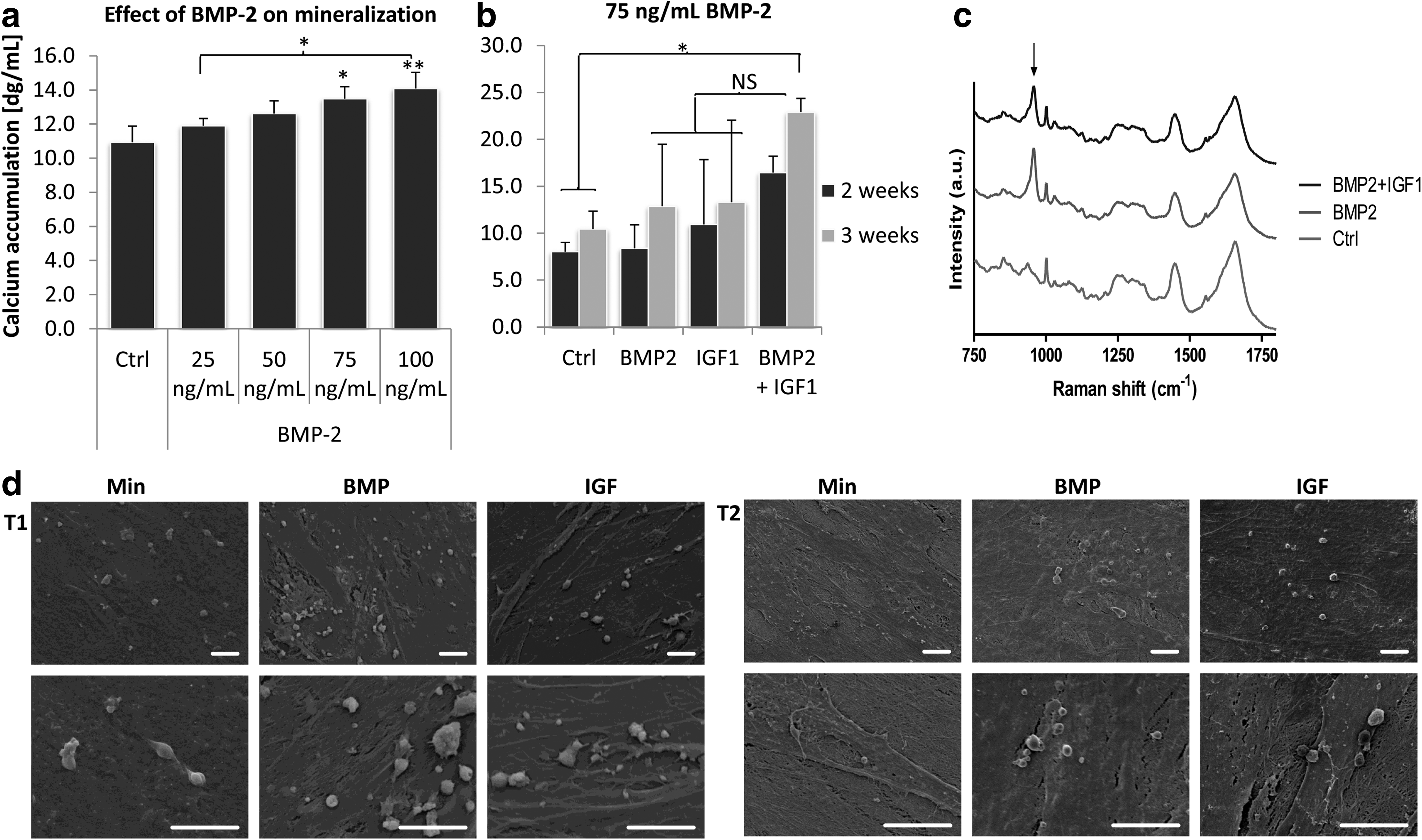
IGF-1 synergistically acts with bone morphogenetic protein-2 (BMP)-2 to induce mineralization in hMSCs. hMSCs were cultured in mineralization medium in the presence of various concentrations of BMP-2. 75 and 100 ng/mL BMP-2 significantly increased calcium deposition, whereas concentrations below had no effect
AdIGF-1 increases runx2 and osteonectin expression in vitro
To investigate the effects of secreted IGF-1 on in vitro differentiation of hMSCs, cells were adenovirally transduced with IGF-1 and after 5 days of culture, gene expression levels of runx2, collagen type I, ALP, and osteonectin were examined. As depicted in Figure 4a, transduction with IGF-1 resulted in high expression levels of IGF-1 already after 2 days. Compared to control, GFP transduced cells, IGF-1 also increased osteonectin and runx2 expression, but collagen type I levels were not affected and ALP levels were only slightly but not significantly increased after 5 days. The amount of secreted IGF-1 after adenoviral transduction was determined every 3 days for a total period of 15 days, and at the time of qPCR (2 and 5 days), cells secreted ∼6 or 20 ng/mL IGF-1, respectively (Fig. 4b).
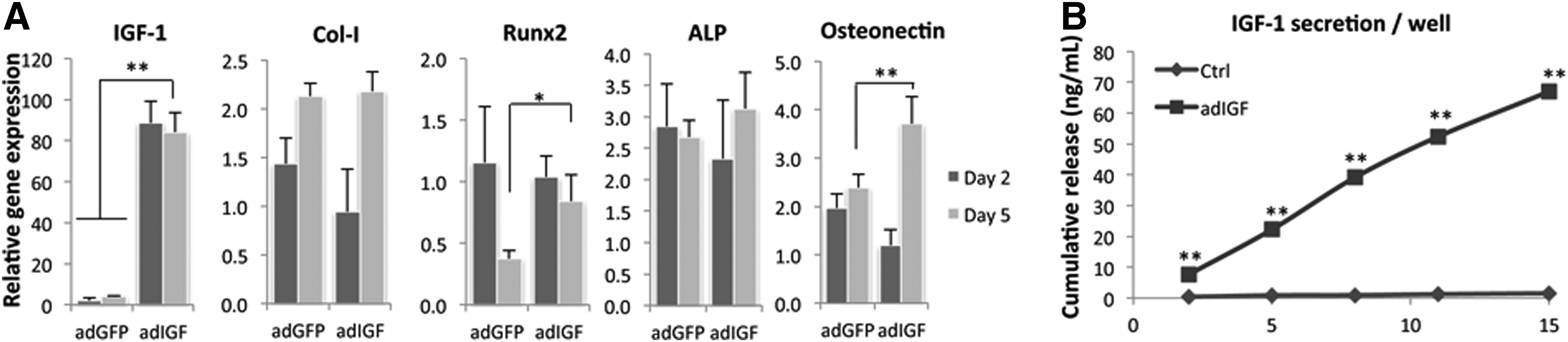
Adenoviral transduction with IGF-1 increases expression of runx2 and osteonectin. hMSCs were transduced with adIGF-1 and after 5 days of culture, Runx2, collagen type I, ALP, and osteonectin gene expression levels were determined. Overexpression of IGF-1 resulted in high expression of IGF-1, and, compared to the control (adGFP), significantly increased osteonectin levels, and runx2 levels after 5 days, but collagen type I expression was not affected. ALP levels were slightly increased after 5 days, but this was not significant
Cotransduction with adIGF-1 and adBMP-2 enhances BMP-2 secretion and in vivo bone formation
Then, to investigate the effects of IGF-1 and synergistic effects of IGF-1 and BMP-2 on in vivo bone formation, hMSCs were adenovirally transduced with IGF-1 or BMP-2, or both (IGF-1/BMP-2). As a control, cells were transduced with GFP. As a control, cells from the same transduction batch were kept in culture in vitro and the cumulative release profile was determined by collecting medium every 3 days for a total period of 30 days. As depicted in Figure 5A, the release of IGF-1 was significantly lower in cotransduced cells but, surprisingly, the secretion of BMP-2 was significantly increased in cells transduced with both BMP-2 and IGF-1, compared to cells transduced with BMP-2 alone. Therefore, to investigate the interplay between adIGF-1 and adBMP-2 transductions in more detail, hMSCs were cotransduced with IGF-1 (MOI 1000) and a range of BMP-2 (MOI 31.25–250). Figure 5B shows the cumulative release of IGF-1 and BMP-2 from single-transduced and cotransduced cells. As expected, transduction with IGF-1 alone, resulted in higher secretion of IGF-1 compared to cotransduced cells, and this decreases with increasing concentrations of BMP-2. Secretion of BMP-2 also increased with increasing MOI, but surprisingly, in samples with a low MOI of adBMP-2, BMP-2 secretion was higher after cotransduction with both IGF-1 and BMP-2, whereas secretion levels were similar upon cotransduction of adIGF-1 and high concentrations of adBMP-2. To determine if overexpression of IGF-1 induces secretion of BMP-2 via a feedback mechanism, BMP-2 secretion by adIGF-1 transduced cells was examined, but these levels were comparable to control (GFP).

Cotransduction of IGF-1 and BMP-2 facilitates BMP-2 secretion and results in increased amounts of bone formation in vivo. hMSCs were transduced in vitro with IGF-1, BMP-2 or both and implanted in vivo. The concentration of secreted IGF-1, as tested in vitro, was reduced upon cotransduction of IGF-1 and BMP-2, whereas the secretion of BMP-2 was increased upon cotransduction
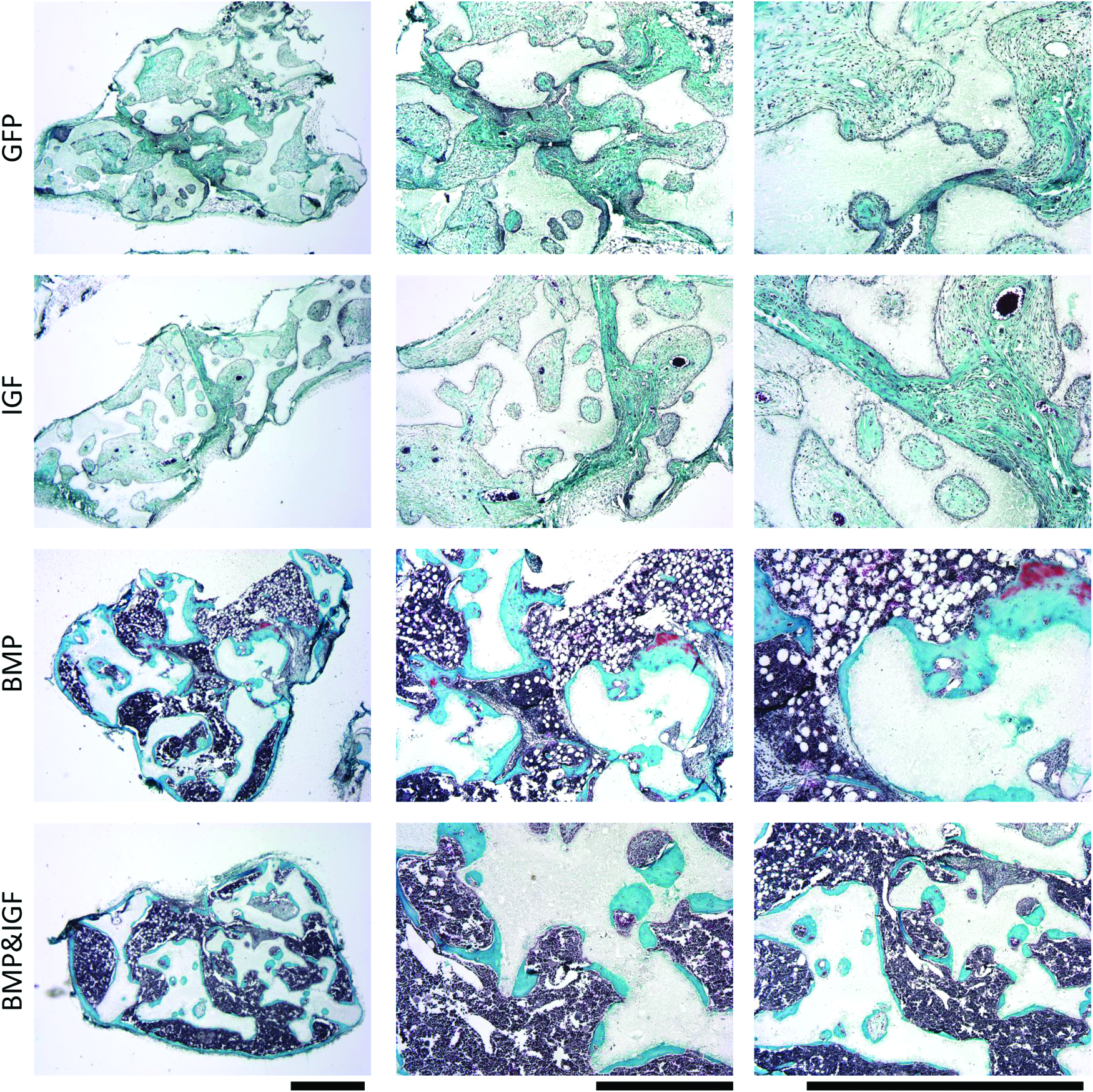
In line with these findings, after 6 weeks of implantation, bone formation in the IGF-1/BMP-2 group was higher than in the BMP-2 group (39% and 48%, respectively). In contrast, no bone was formed in the GFP group, whereas some scaffolds in the IGF group displayed a low percentage of bone (0.6%) (Fig. 5C). The in vitro release profile suggests that the increased amounts of bone may be related to enhance BMP-2 secretion. On the other hand, histological observations showed that bone in the BMP-2/IGF-1 group displayed a lower level of mineralization, indicating a less mature state, compared to BMP-2, or IGF-1 groups. In both BMP-2 and BMP-2/IGF-1 groups, bone marrow was formed inside the scaffolds pores, but the marrow was more densely packed and more granular in the BMP-2/IGF-1 group (arrows, Fig. 6). The less mature state of the bone in the BMP-2/IGF-1 group was confirmed by Masson's Trichrome staining (Fig. 7), which showed that only the BMP-2 group contained areas of mature bone (red).

Histological overview of in vivo bone formation. No bone was formed in the GFP group, whereas in the IGF group small amounts of mature bone were observed in some implants. Large amounts of mature bone were formed in the BMP-2 group, whereas in the BMP-2/IGF-1 group, large amounts of osteoid were formed. In addition, in both the BMP-2 and the BMP-2/IGF-1 group, large amounts of bone marrow (arrows) occupied spaces between the scaffolds, but this was denser and more granular in the BMP-2/IGF-1 group. Magnifications; top 40×, middle 100×, bottom 200×. Color images available online at www.liebertpub.com/tea
Mature bone was observed in the BMP-2 group, but not in the BMP-2/IGF-1 group. Mason's Trichrome staining of decalcified in vivo samples showed parts of mature bone formation (red) in the BMP-2 group. This was surrounded by immature bone (green), which was also formed in the BMP-2/IGF-1 group. Scale bar=1 mm. Color images available online at www.liebertpub.com/tea
Discussion
Numerous studies have investigated the effects of IGF-1 on MSCs from different sources and, whereas some studies show increased proliferation and differentiation of hMSCs in response to IGF-1, others fail to show an effect. Culture protocols vary greatly between these studies, which makes it difficult to compare them. Walsh reported that a concentration of 10 ng/mL IGF-1 had no effect on proliferation or ALP expression of CFU-Fs 20 , whereas a study by Koch et al. showed increased expression of runx2 and collagen type I after adenoviral transductions, with the levels of secreted IGF-1 around 120 ng/mL 10 days post-transduction. 22 These data suggest that low concentrations of IGF-1 do not affect proliferation or differentiation of hMSCs, which was also demonstrated by our ALP experiments, which show a clear dose-dependent effect. In addition, we show here that with a concentration of 100 ng/mL rhIGF-1, expression of osteonectin and runx2 increase 3–5-fold, whereas adenoviral overexpression (resulting in 20 ng/mL secreted IGF-1 5 days post-transduction) only slightly increased runx2 and osteonectin expression and did not affect collagen type I. Similarly, treatment with rhIGF-1 resulted in an increase in ALP levels after 5 days of culture, which could not be repeated using adenoviral overexpression of IGF. Other studies using scaffold-release systems that result in a high concentration of IGF-1 (80–100 ng/mL) indeed also show positive effects on proliferation and osteogenic differentiation.23,24
The presence of serum in the medium also plays a key role when investigating the effects of IGF-1. In the absence of serum, we found an increase in both proliferation and differentiation of hMSCs, whereas in the presence of serum IGF-1 had no effect. Most likely, serum contains proteins that neutralize IGF-1 function,
IGFBPs and the ALS, which were shown to inhibit IGF-1 function in mouse models. Overexpression of ALS, 32 IGFBP-1, 33 −2, 34 −3 35 or −5 36 resulted in deficiencies in the skeleton, such as reduced body weight, growth retardation, diminished mineralization, decreased cortical thickness, and reduced cortical bone volume. Although total IGF-1 serum levels were not affected in these models, the amounts of extracellular fluid IGF-1 available to bind to tissue receptors were decreased; thus suggesting that these proteins inhibit IGF-1 activities.
The potential of IGF-1 to improve fracture healing has been demonstrated in in vivo models, but studies are not consistent. Both local and systemic delivery of rhIGF-1 were reported to enhance bone formation in a distraction osteogenesis mouse model, 37 but systemic administration in a rat femoral defect had no effect. 38 Also, systemic delivery increased IGF-1 serum levels, which could potentially result in negative side effects. Therefore, local application using controlled release systems offers a better alternative, especially for tissue engineering applications, but also in this case reports are not consistent. Meinel showed enhanced bone formation by IGF-1-releasing scaffolds, 39 whereas a study by Laffargue et al. shows a slight increase at best. 40 So far, better results are obtained using dual release systems. Implants releasing both IGF-1 and TGF-β resulted in more advanced bone remodeling in fracture models in rats and sheep41,42 and the combination of IGF-1 and platelet-derived growth factor also improved bone regeneration.43,44 BMP-2 is the main player in stimulating fracture repair, and therefore, we investigated the combined treatment/secretion of IGF-1 and BMP-2. 45 We demonstrate that IGF-1 potentiates BMP-2-induced mineralization in vitro, whereas IGF-1 alone does not affect mineralization; thus, showing a synergistic effect. This is in line with a mouse model, which demonstrates that IGF-1 is involved in mineralization. 6 Unexpectedly, using a range of BMP-2 concentrations, mineralization of hMSCs (donor 134A; donor numbers correspond to our internal databank) was increased by 75–100 ng/mL BMP-2, but in a second study (donor 102) using both BMP-2 and IGF-1, a concentration of 75 ng/mL BMP-2 did not significantly affect calcium deposition, although synergistic effects between BMP-2 and IGF-1 were still observed. We speculate that this is a donor-dependent effect.
In addition to these in vitro data, we found higher amounts of newly formed bone by cells cotransduced with both IGF-1 and BMP-2, compared to BMP-2 alone. Surprisingly, the amount of BMP-2 secreted by cotransduced cells was higher than by cells transduced with BMP-2 alone. Additional experiments using a range of adBMP-2 MOI's also demonstrated that, when using low MOI, the secretion of BMP-2 increases upon cotransduction, suggesting that adIGF-1 facilitates adBMP-2 transduction, BMP-2 gene expression or BMP-2 protein secretion. This is interesting, since secreted IGF-1 is reduced upon cotransduction with adBMP-2, and also negatively correlates with increasing MOI's of adBMP-2. Further experiments are required to investigate this in more detail. Overexpression of IGF-1 alone resulted in small amounts of new bone in vivo, but did not lead to clinically relevant amounts of bone. Due to the increase in BMP-2 secretion after cotransduction it is difficult to draw conclusions regarding the effect of IGF-1 on BMP-2-induced bone formation. We have previously demonstrated that scaffold incorporation of 1–40 μg rhBMP-2 does not result in morphological differences of newly formed bone, although more bone marrow was found with increasing concentrations of BMP-2. 46 Therefore, the increased BMP-2 secretion probably accounts for the increase in bone and bone marrow; however, it is unlikely that the osteoid appearance of the bone is due to higher concentrations of BMP-2, which suggests that IGF-1 has a negative effect on the maturation of bone.
Conclusions
We demonstrated that IGF-1 enhances proliferation and osteogenic differentiation of both hMSCs and MG-63 in vitro, but its effects are highly dependent on the used concentration and the presence of serum. In vivo, IGF-1 had small effects on bone formation of hMSCs, although it seemed to inhibit the maturation of BMP-2-induced bone formation.
Footnotes
Acknowledgments
The authors gratefully acknowledge the support of the TeRM Smart Mix Program of the Netherlands Ministry of Economic Affairs and the Netherlands Ministry of Education, Culture, and Science.
Disclosure Statement
All animal experiments were approved by the local animal ethics committee. The authors state that they have no conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.