
Abstract
Background:
The mechanical properties of articular cartilage are dominated by the interterritorial matrix, as the matrix in this region is stiffer, greater in volume, and more interconnected compared to that in the pericellular and territorial region. Hence, tissue-engineered constructs in which a newly synthesized matrix accumulates in the pericellular and territorial regions may be of a lower mechanical quality compared to constructs in which the interterritorial region contains abundant matrix.
Objective:
In this study, we explored the extent to which matrix distribution may be modulated by altering the agarose concentration and the presence of the transforming growth factor-β (TGF-β) and how this affects the mechanical properties of cultured cartilage constructs.
Methods:
Cartilage development in constructs with agarose concentrations varying from 1%, 2%, and 3% (study 1) and in constructs with no or very low agarose concentrations of 0.25%, 0.5%, and 1% (study 2) were compared. In both studies, the effect of TGF-β3 was compared to fetal bovine serum. After 21 and 42 days of culture, the matrix content and distribution were analyzed and mechanical properties were assessed at day 42.
Results:
Culture in lower agarose concentrations did not significantly influence the matrix content per wet weight, but did result in a more homogeneous distribution. Constructs cultured with less agarose also showed a higher equilibrium modulus. The presence of TGF-β3 resulted in an increased extracellular matrix (ECM) deposition, a more homogeneous matrix distribution, and an equilibrium modulus.
Conclusions:
Culturing with no or low agarose concentrations and TGF-β3 is favorable for cartilage tissue-engineering studies, because both stimulate the formation of a more homogeneous ECM and consequently result in improved mechanical properties.
Introduction
Agarose hydrogels are commonly used as a model scaffold for cartilage tissue-engineering studies in vitro,3,4 and has even shown potential for its use in vivo. 5 Chondrocytes cultured in agarose maintain their phenotype and synthesize cartilage-specific extracellular matrix (ECM) components, such as proteoglycans and collagen type II,6,7 which become entrapped in the hydrogel and coalesce into a functional ECM over time in culture. 4 Yet, the load-bearing properties of agarose-based tissue-engineered cartilage constructs are inferior to those of native cartilage. In chondrocyte-seeded agarose constructs, it is often observed that the matrix is not distributed homogeneously, with the newly synthesized matrix accumulated mostly in the pericellular region and mainly soft agarose in the interterritorial region.8–10 It can be postulated that constructs with such an inhomogeneous matrix distribution are of lower mechanical quality compared to constructs that also have their interterritorial region filled with stiff synthesized matrix. Unfortunately, experimental data that evaluate this theory does not exist, and it is not known how matrix distribution in cultured cartilage constructs could be modulated. In this study, we therefore explore the separate and combined effects of two approaches that may result in differences in matrix homogeneity at the microscopic level.
The first approach to change the matrix distribution is by modulating the transport rate of the newly synthesized matrix away from the cell before assembly, assuming that in more permeable scaffolds, the matrix would be more dispersed than in less permeable scaffolds. The concentration of the agarose gel determines its diffusivity. Higher agarose concentrations result in stiffer gels with decreased hydraulic permeability,11–13 due to decreased pore size 12 and reduced number of pores. 14 It was hypothesized by Ng et al. that, due to decreased pore size and number in stiffer agarose constructs (3%), newly synthesized matrix molecules are entrapped more efficiently, resulting in an increased amount of matrix within the constructs and a reduced loss into the culture medium.15,16 Indeed, using numerical modeling, the permeability of agarose has been shown to influence the distribution of matrix molecules within the agarose constructs.17,18 Reduced diffusion of matrix molecules was predicted in high-concentration agarose constructs, resulting in matrix islands around the cells. In low-concentration agarose hydrogels, a more homogeneous matrix distribution was predicted due to increased molecular diffusion. 17 Unfortunately, experimental data for evaluation of this prediction do not exist.
The second approach to change ECM distribution is by adding the transforming growth factor-β3 (TGF-β3) to the cultures. Although the exact mechanism remains unclear, it has been shown that the TGF-β3 not only stimulates ECM deposition,19,20 but also has an effect on matrix distribution within tissue-engineered cartilage constructs. 21
The aims of the present study are therefore to first explore the extent at which matrix distribution at the microscopic level may be modulated by each of the two approaches mentioned above and the combination thereof, and second, to evaluate the effect of ECM distributions on the mechanical properties of the engineered cartilage tissue.
Methods
Experimental design
Two different studies were carried out. In study 1, cartilage development in chondrocyte-seeded agarose hydrogel constructs with agarose concentrations 1%, 2%, and 3% was compared. In study 2, cartilage development in chondrocyte-seeded agarose constructs with no agarose (0%) or very low agarose concentrations (0.25%, 0.5%, and 1%) was compared. Because constructs with no or very low agarose concentrations are initially very fragile, constructs in study 2 were cultured in an agarose well setup to prevent disintegration. In both studies, the effect of serum-free culturing with TGF-β3 supplementation (the TGF medium, consisting of high glucose Dulbecco's modified Eagle's medium (hgDMEM) supplemented with 1% Pen-Strep, 0.1% amphotericin, 0.1 mM dexamethasone, 50 mg/mL ascorbate 2-phosphate, 20 mg/mL L-proline, 100 mg/mL sodium pyruvate, 1× ITS+, and 10 ng/mL TGF-β3) was compared to culturing with a standard culture medium supplemented with 10% fetal bovine serum (the FBS medium, consisting of hgDMEM supplemented with 1% Pen-Strep, 0.1% amphotericin, 50 mg/mL ascorbate 2-phosphate, and 10% FBS).
Cartilage and chondrocyte isolation
Full-depth slices of articular cartilage were removed from eight metacarpal-phalangeal joints of the forelimbs of young (8–12 month old) calves. Cartilage slices were pooled and digested overnight in a digestion medium (hgDMEM+10% FBS+1% Pen-Strep, all purchased from Gibco and 0.1% amphotericin B+0.5% collagenase type II; Antonides–Interchema) for 16 h in an incubator at 37°C and 5% CO2, while stirring. The resulting cell suspensions were purified by filtration through a 70-μm-pore-size cell strainer (Becton Dickinson), washed twice with hgDMEM, and resuspended in hgDMEM supplemented with 10% FBS, 1% Pen-Strep, and 0.1% amphotericin.
Culture protocol
For study 1, chondrocyte-seeded agarose hydrogel constructs with 1%, 2%, and 3% (w/v) agarose and a final cell concentration of 20 million cells/mL were produced. Cell suspensions were mixed with agarose (type VII; Sigma) as shown in Table 1 and transferred into a sterile stainless steel mold, containing holes measuring 6.5 mm in diameter and 2 mm in height (volume 57 μL). The chondrocyte/agarose suspensions were allowed to solidify at room temperature for 10 min to yield cylindrical constructs.
Final constructs contained 20 million cells/mL.
For study 2, chondrocyte-seeded agarose hydrogel constructs with 0%, 0.25%, 0.5%, and 1% (w/v) agarose and a final cell concentration of 20 million cells/mL were created in custom-designed agarose wells, adapted from Elder et al. 22 To create these wells, a stainless steel pin with a diameter of 6.5 mm was put in a well of a six-well plate, after which 7 mL of 3% agarose (type VII; Sigma) was added to the well and allowed to solidify at room temperature for 10 min (Fig. 1A). Afterward, the pin was removed, and subsequently, the agarose well was punched out with a 12-mm diameter punch (Fig. 1B) and transferred to the center of a culture well in a 12-well plate. The agarose well was fixed with a thin layer (0.5 mm) of 3% agarose (Fig. 1C, D). The agarose well was flushed for 2 days with hgDMEM supplemented with 1% Pen-Strep and 1% amphotericin. Afterward, 57 μL of agarose–cell suspension mixture (Table 1) was added to each agarose well, which was allowed to solidify at room temperature for 20 min. As a control, 1% agarose discs (as described for study 1; 1% disc) were additionally produced.

Overview culture setup study 2.
The constructs in both studies were subsequently cultured in either the TGF medium or the FBS medium for 21 and 42 days (n=12–18 per agarose concentration and medium composition). The medium was changed every other day and 300 μL of the removed medium per sample was stored at −30°C for analysis of the sulfated glycosaminoglycan (sGAG) content. In both studies, 1 construct per group was stained after 24 h and 42 days after fabrication with a standard calcein AM/propidium iodide live/dead assay (Invitrogen) to assess cell viability and distribution within the constructs.
Mechanical testing
After 42 days of culture, the equilibrium Young's modulus (Eeq) of samples of both studies (n=4 per agarose concentration and medium composition) was determined with a testing protocol similar to that used by Mauck et al. 23 The thickness of each sample was measured and afterward the samples were placed in a bath with phosphate-buffered saline at room temperature and were subsequently equilibrated in creep to a tare load of 0.02 N with an impermeable plate connected to a 2 N load cell using a standard materials testing machine (Zwick Roell Z010). From this offset, stress relaxation tests were performed with a ramp speed of 1 μm/s until reaching 20% strain. After equilibrium was reached, Eeq was calculated from the ratio of the equilibrium stress to the applied strain using testXpert II software (Zwick Roell).
Biochemical content
After 21 and 42 days of culture, halves of six constructs from each treatment group were lyophilized, and subsequently, 0.5 mL of the papain digestion buffer (100 mM phosphate buffer, 5 mM L-cysteine, 5 mM EDTA, and 140 μg papain per mL) was added to the samples, followed by melting for 1 h at 70°C. Afterward, the solution was incubated for 16 h at 37°C with an additional 3.5 μL of the agarase solution (1 kU/mL). After digestion with papain and agarase, the samples were centrifuged and the supernatant was used for quantification of DNA, sGAG, and collagen content.
The amount of DNA was determined using the Hoechst dye method 24 with a reference curve of calf thymus DNA (D4764; Sigma). The sGAG content of samples was determined using a modification of the assay described by Farndale et al., 25 and a shark cartilage chondroitin sulfate reference (C4348; Sigma). For analysis of the sGAG content in the collected culture medium, a buffered 1,9-dimethylmethylene blue (DMMB) solution with a pH of 3 was used. For determination of the collagen content, digested tissue samples were hydrolyzed in 6 M hydrochloric acid (Merck) and the orthohydroxyproline (OHP) quantity was assessed as described previously 26 using an assay modified from Huszar et al. 27 and a trans-4-hydroxyproline (H5534; Sigma) reference. A collagen:OHP ratio of 10:1 was used as a conversion factor. 28
Histology
Three half constructs from each treatment group were fixed for 24 h in 3.7% formaldehyde, paraffin embedded, and 5-μm-thick sections were stained with the Alcian blue dye to evaluate the distribution of sGAG or Picrosirius Red to evaluate distribution of collagen. Hematoxylin was used as nuclear counterstaining.
Statistical analysis
Statistics were performed with Statgraphics (Statgraphics; Manugistics, Inc.). The effects of treatment and time as well as their interaction were examined for significant differences by two-way ANOVA, with the equilibrium Young's modulus and amounts of sGAG, DNA, and collagen as the dependent variables. If no significant interaction was found, a one-way ANOVA with Bonferroni post hoc testing was used to search for significant differences in either time or treatment within the other factor level. In case of a significant interaction, an independent t-test with Bonferroni correction for a number of applied comparisons was used to test for significant differences between each specific time and treatment subgroup. Statistical significance was assumed for p<0.01.
Results
Gross appearance, cell distribution, and viability
The constructs cultured without agarose were notably smaller than the constructs containing agarose. Constructs without and with 0.25% agarose appeared weak and fragile at day 1 and where somewhat smaller than the higher agarose concentration constructs. With increasing agarose concentration and culture time, the constructs appeared to be firmer and stronger. A homogeneous cell distribution was observed in all the constructs. Cell aggregation was observed in constructs cultured without agarose and 0.25% agarose constructs appeared to have a somewhat increased cellular density. Cells in these constructs were closer together as the weak agarose mesh in these constructs appeared to have collapsed after casting. Cell viability was >90% in all constructs 24 h and 42 days after fabrication of the constructs (data not shown).
Biochemical content
DNA content
Culturing in the presence of FBS resulted in significantly more DNA in conventional agarose disc constructs compared to culturing with TGF-β3 (p<0.01) (Fig. 2A). This difference between FBS and TGF was absent in constructs cultured in the well setup (Fig. 2B). There are no significant differences in the DNA content in time or with increasing agarose concentration, except in study 2 between 0.5% and 1% agarose at day 21 in constructs cultured with FBS (p<0.01) (Fig. 2B).

The DNA content in cultured cartilage constructs study 1
GAG content
No significant differences in the GAG content were observed between the three agarose concentrations in study 1 (Fig. 3A). In study 2, the agarose concentration did not influence the GAG content in constructs cultured with FBS, except for the 0.25% group at day 21, which contains less GAG (significant for 0.5% and 1%; p<0.01) (Fig. 3B). In the presence of TGF-β3, constructs cultured without or with very low agarose (0.25%) contained significantly more GAG at day 42 compared to constructs cultured with more agarose (p<0.01) (Fig. 3B). In both studies, the GAG content increased significantly with time in all groups (all p<0.01) and constructs cultured in the presence of TGF-β3 contained more GAG compared to constructs cultured with FBS.

The glycosaminoglycan (GAG) content in cultured cartilage constructs study 1
GAG release
GAG is released with time in the culture medium in all groups in both studies (Fig. 4A, C). In the presence of FBS, the agarose concentration has no effect on GAG release in both studies. In study 1, release of GAG from constructs cultured in the presence of TGF-β3 significantly decreased with increasing agarose concentration (Fig. 4B; p<0.01), but this effect was absent in study 2. A culture medium of constructs cultured in the presence of TGF-β3 contained significantly more GAGs compared to constructs cultured with FBS for all agarose concentrations in study 1 (Fig. 4B; all p<0.01). In study 2, the same trend was observed, but was only significant for 1% agarose (both disc and well setup). Interestingly, significantly more GAGs were released in the culture medium of the 1% agarose discs, compared to the constructs that were cultured in the customized agarose wells (Fig. 4D; all p<0.01).
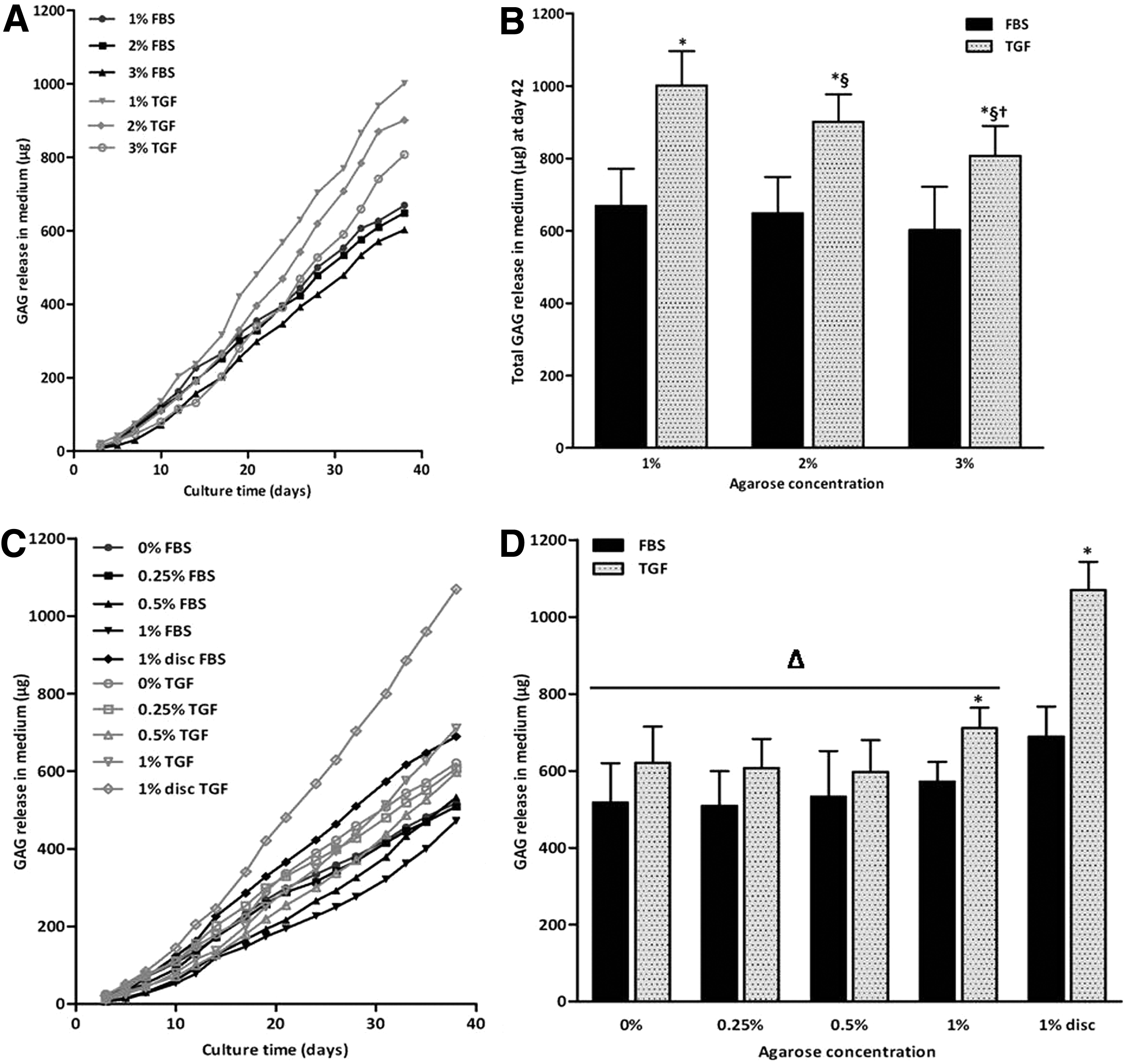
GAG release in the culture medium over 42 days of culture of study 1
Collagen content
The agarose concentration did not affect the collagen content of constructs cultured with both medium compositions in study 1 (Fig. 5A) and neither did in constructs cultured with FBS in study 2 (Fig. 5B). In contrast, the collagen content in constructs that were cultured in the presence of the TGF-β3 significantly varied with varying agarose concentrations, but no clear trend was visible (Fig. 5B). Constructs without agarose cultured in the presence of the TGF-β3 contained significantly more collagen than any of the other constructs (p<0.01). The collagen content increased significantly with time in all groups in study 1 (all p<0.01), whereas in study 2, no significant differences in the collagen content between day 21 and 42 were observed, except in constructs and discs cultured with 1% agarose. Further, culturing with TGF-β3 resulted in more collagen compared to FBS in both studies (p<0.01). No collagen was detectable in the culture medium.
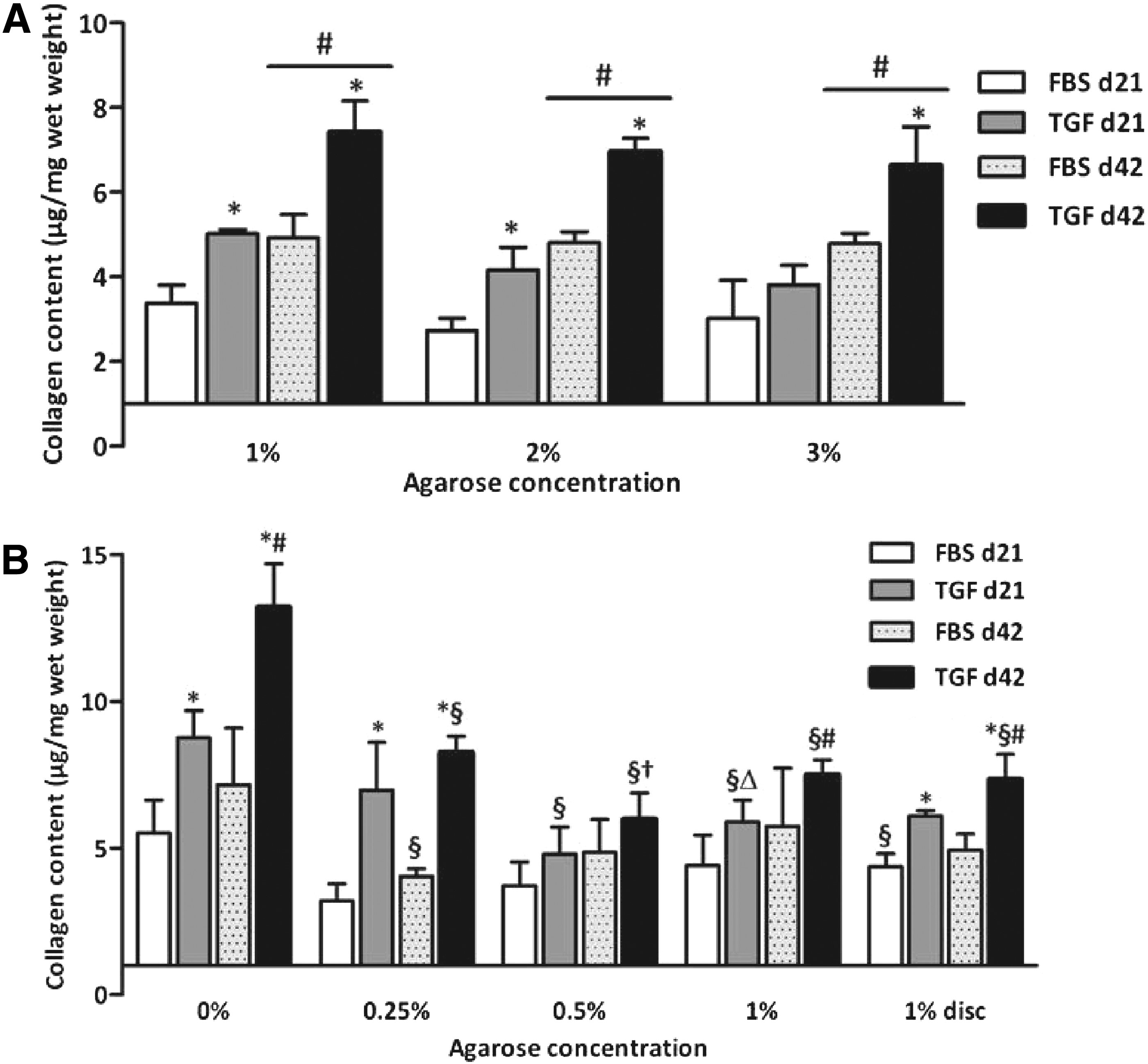
The collagen content in cultured cartilage constructs in study 1
Histology
Alcian Blue staining and Picrosirius Red staining of the cultured constructs showed that GAG and collagen deposition was evident in all groups (Figs. 6 and 7). Constructs cultured in the presence of FBS without or with a low agarose concentration (0%–0.5%) showed a more diffuse deposition of GAGs and collagen compared to the higher agarose concentrations. Between the higher agarose concentrations (1%–3%), no obvious differences in tissue distribution could be observed in FBS-containing cultures. In contrast, in TGF-β3-supplemented cultures, this effect of the agarose concentration on matrix distribution was absent and these constructs showed a more uniform GAG and collagen distribution compared to FBS cultures in both studies.
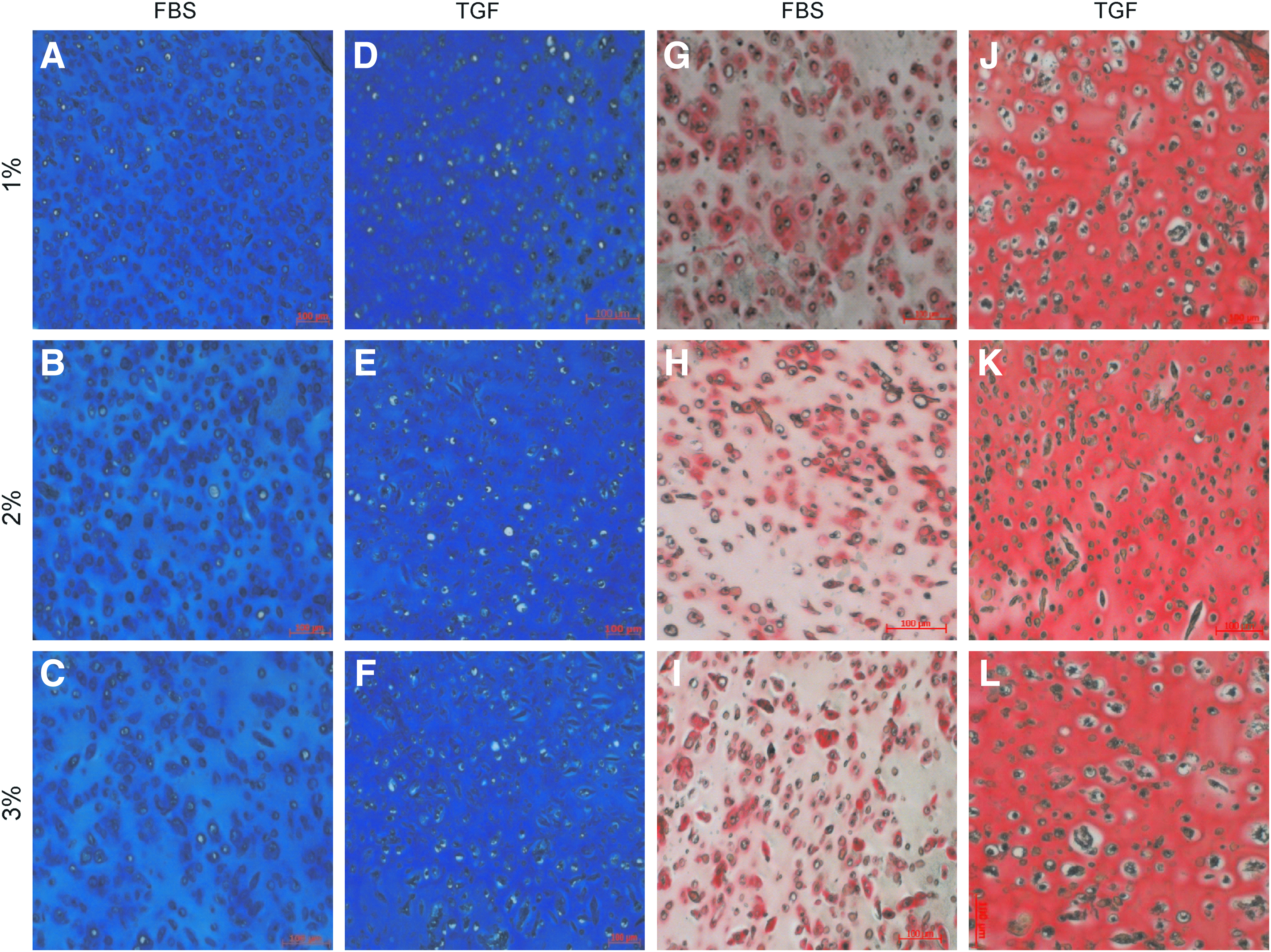
Alcian blue staining for sulfated glycosaminoglycan (sGAG)
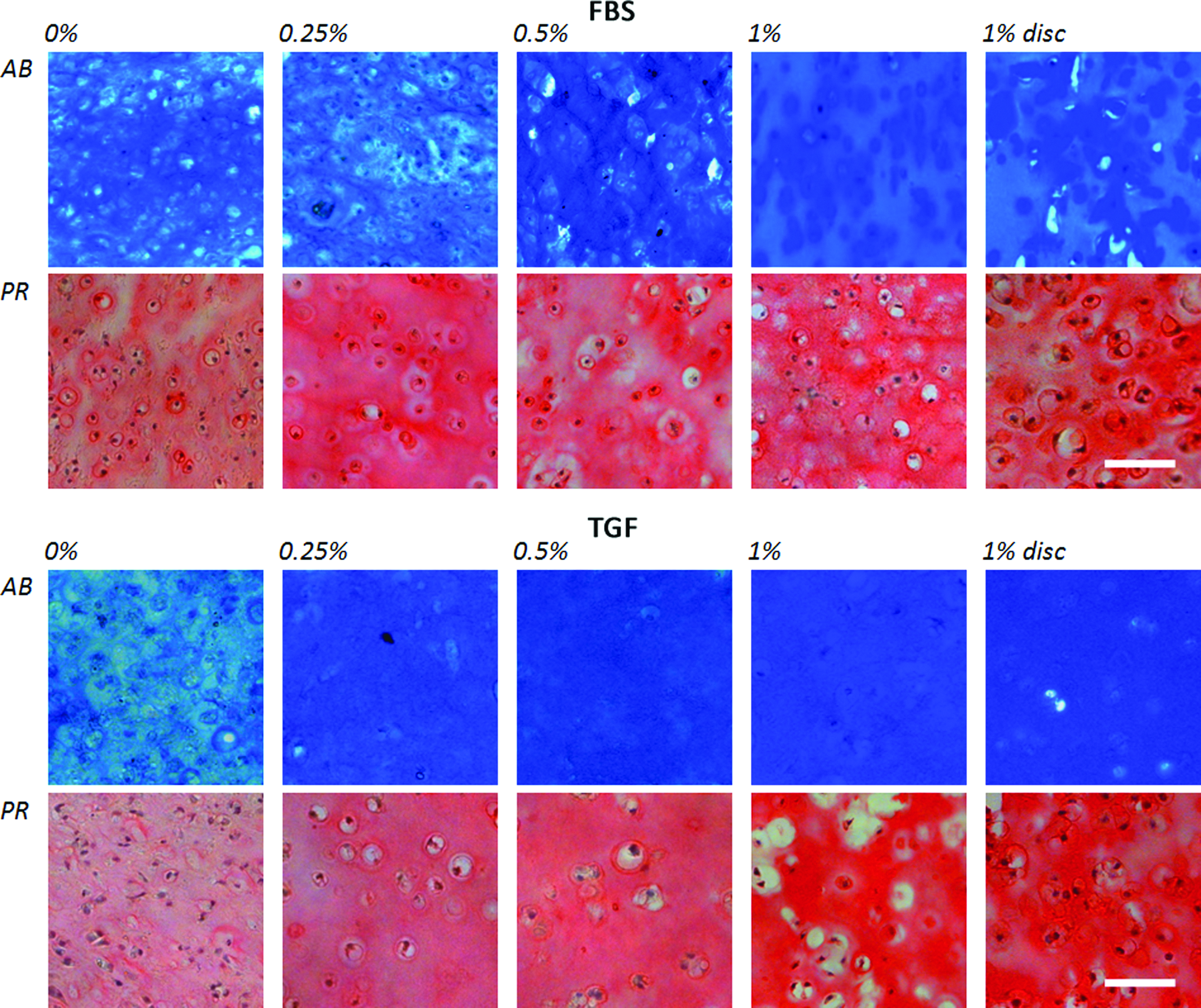
Alcian blue staining for sGAG and Pircrosirius Red staining for collagen of cross-sectional slides of 5 μm thickness of the cultured chondrocyte-seeded agarose constructs after 42 days of culture with hematoxylin as nuclear counterstaining. Color images available online at www.liebertpub.com/tea
Mechanical properties
Agarose concentration, medium composition, and their interaction significantly affected the equilibrium compressive Young's modulus (Eeq) of cultured constructs at day 42 (all p<0.01). In study 1, Eeq increased significantly with increasing agarose concentration in all groups (p<0.01), except between 1% and 2% agarose in constructs cultured in the presence of TGF-β3 (Fig. 8A). In addition, it was seen that with increasing agarose concentration, the equilibrium modulus of the constructs was to a lesser extent determined by the modulus of the agarose and more by the deposited tissue. In study 2, no significant differences in Eeq were observed between agarose concentrations of constructs cultured with FBS. In the presence of TGF-β3, Eeq of constructs cultured without agarose was significantly higher compared to any of the constructs with agarose (p<0.01). No differences between agarose concentrations were observed, except for 0.5% agarose constructs, which exhibited a significantly decreased Eeq compared to 0.25% agarose (p<0.01) (Fig. 8B). Constructs cultured without agarose demonstrated a significantly higher Eeq compared to constructs that were cultured with agarose (all p<0.01). Eeq of empty constructs could not be determined for agarose concentrations below 1% in study 2, since they were too weak to be accurately measured.
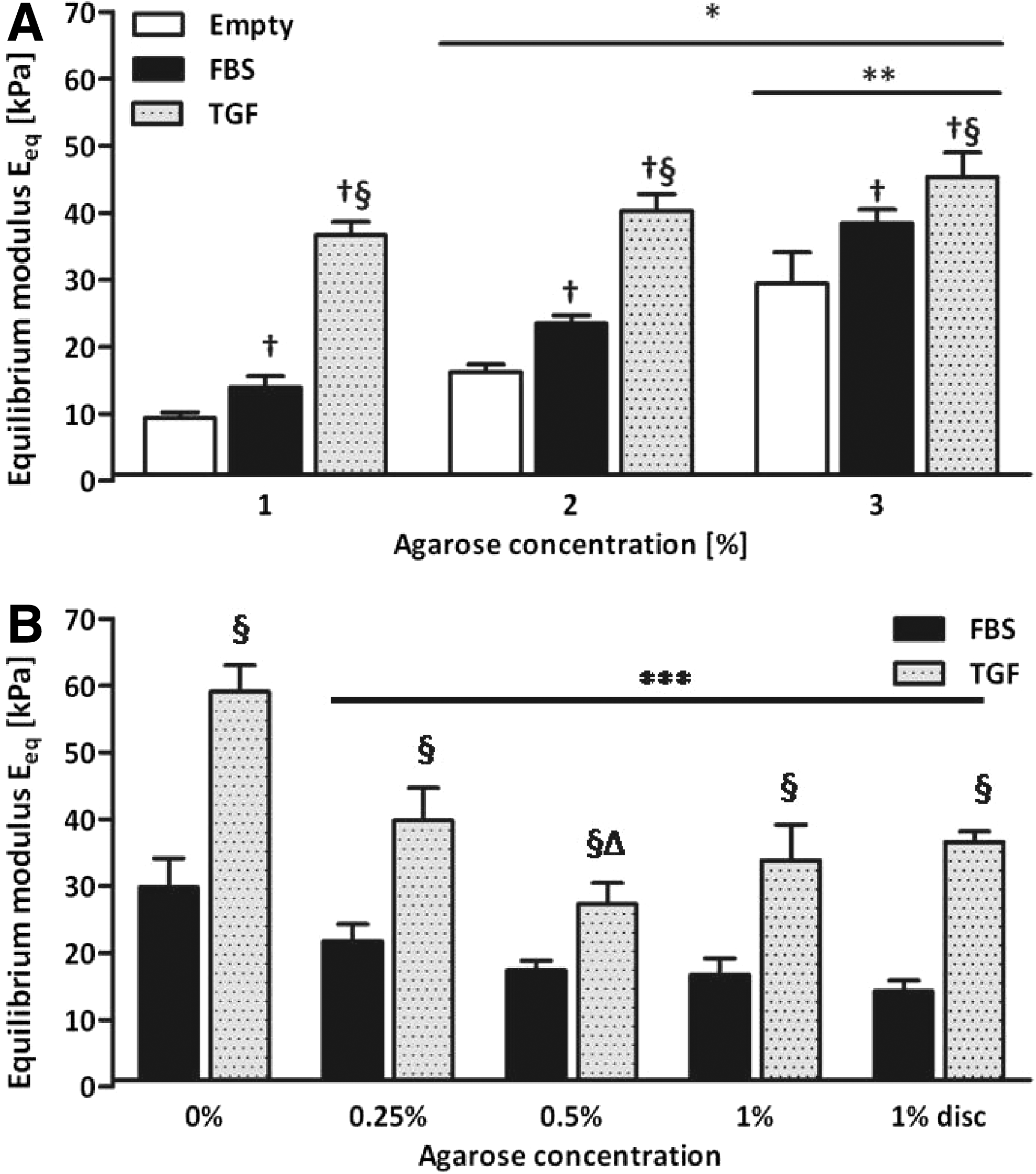
The equilibrium modulus of cultured cartilage constructs from study 1
In both studies, constructs cultured in the presence of TGF-β3 exhibited a significantly higher Eeq than constructs made of the same agarose concentration, but cultured in the presence of FBS (p<0.01).
Discussion
We have demonstrated that lowering the agarose concentration does result in a more homogeneous distribution of GAGs and collagen and improved mechanical properties. Further, we have demonstrated that culturing in the presence of TGF-β3 results in increased matrix content and a more homogeneous matrix distribution, compared to culturing with FBS. Culturing without agarose in the presence of TGF-β3 leads to highest mechanical properties and biochemical content, compared to any of the constructs containing agarose.
Lowering the agarose concentration resulted in a more homogeneous distribution of matrix components. Constructs cultured with a lower percentage agarose and agarose-free constructs (0%, 0.25%, and 0.5%) were smaller and cells appeared more densely packed, which was probably because cells were drawn closer together as the weak agarose mesh in these scaffolds collapsed after casting. Because cells were closer together, it is suggested that the ECM in the pericellular and territorial regions could easily unite, which gives it a more homogeneous appearance of the deposited matrix. An increased deposition in the interterritorial region was, however, visible between 0.5% and 1% agarose constructs, in which both had the same cell density, indicating that the agarose concentration indeed modulated ECM distribution. The reduced size of the constructs could theoretically also have influenced the release of the ECM in the culture medium, since the distance for synthesized molecules to diffuse out of the constructs decreases in smaller constructs. However, we did not observe this in our study.
To our knowledge, there are no other studies, which have examined the influence of agarose concentration on ECM distribution and release of matrix components in the culture medium. Studies that varied crosslinking density of poly(ethylene glycol) (PEG) hydrogels, which impacts molecular diffusion, 29 have reported a similar trend of increasing interterritorial ECM deposition in PEG hydrogels with reduced crosslinking.30,31 The results of their and our study were in agreement with numerical predictions by Sengers et al. and van Donkelaar et al., who have predicted that in less permeable hydrogels, matrix deposition would be mostly pericellular, whereas in gels with an increased diffusion rate, a more diffuse distribution pattern would emerge.17,18
There is no direct correlation between the agarose concentration and quantity of matrix within constructs. Furthermore, lowering the agarose concentration did not result in increased release of matrix components into the culture medium. We therefore assume that production and capturing of matrix products within constructs is not directly dependent on the pore size or number, but that other factors, such as matrix–matrix interactions may be involved. These interactions have previously been shown to affect the assembly of matrix molecules. The increased diffusion rate of synthesized matrix molecules led to improved collagen fibrillogenesis. 32 Also, a smaller agarose pore size could have been a limiting factor for large proteoglycan and hyaluronan aggregate formation. For example, 2% agarose hydrogels have a pore diameter of ±0.12 μm, 15 which is small compared to the length of an aggrecan molecule (±0.4 μm). 33 Hyaluronan aggregates can even contain more than 100 proteoglycan units and have lengths up to 4 μm. 34 In lower agarose concentration constructs that have larger pore sizes, proteoglycan aggregate formation was possibly less hindered, resulting in larger, but not more, aggregates and this may have improved mechanical properties.
Release of matrix components into the culture medium was higher from 1% agarose discs, than from 1% agarose constructs cultured in the confined agarose well setup. In the well setup, there is only 1 way out to the culture medium, whereas discs can release matrix at all sides and which may explain the observed difference. We also investigated whether GAGs could have diffused into the surrounding agarose wells in the well setup by analyzing the agarose wells for GAGs, but amounts were below the detection limit. Culturing in the confined setup is likely to be beneficial for matrix retention within the chondrocyte-seeded constructs.
Culturing with TGF-β3 instead of FBS resulted in improved mechanical properties and increased GAG and collagen content, which is in agreement with other studies.20,26,35,36 In addition, in the presence of TGF-β3, most of the matrix components were laid down in the interterritorial region, further away from the chondrocytes, a finding that is in accordance with a previous study by van Osch et al. 21 In FBS-supplemented cultures, most of the matrix components were deposited close to the chondrocytes in the pericellular and territorial regions. Since mainly the interterritorial matrix contributes to the mechanical properties of the construct, 2 this difference in ECM distribution may explain at least part of the increased Eeq of constructs cultured with TGF-β3. It is unclear why matrix components are laid down further away from the chondrocyte when cultured in the presence of TGF-β3 compared to FBS. It has previously been demonstrated that TGF-β downregulates the formation of collagen pyridinoline crosslink formation by articular chondrocytes in alginate culture. 37 The reduced amount of crosslinks in the cultures with TGF-β3 could have had a twofold effect. First, we speculate that less crosslinking resulted in smaller collagen fibrils, which made them less prone to entrapment and because the diffusion coefficient of molecules inversely related to the molecular size, these smaller fibrils were able to diffuse over larger distances and therefore distribute more evenly. Second, new matrix components are entrapped less efficiently in the surrounding matrix, which enables them to diffuse further away from the chondrocytes. This idea is supported by the increased release of GAGs into the culture medium in TGF-β3 cultures.
Constructs that were cultured without agarose showed improved mechanical and biochemical properties over constructs that were cultured in an agarose hydrogel scaffold. Chondrocytes maintained their phenotype and synthesized cartilage-specific ECM components, which is in agreement with literature.22,38,39 Previous studies have indicated that a scaffold-less approach can lead to relatively high aggregate moduli compared to cultures with scaffold material,40,41 which is in agreement with our results.
Decreasing agarose concentration and culturing in the presence of TGF-β gives more ECM in the interterritorial region compared to constructs with a higher agarose concentration, where the ECM deposition is limited to the pericellular region. Increased matrix deposition in the interterritorial region is more favorable, since this is the main load-bearing part of the matrix. Often there is only the pericellular matrix observed directly around the cells and hardly any ECM in the interterritorial region. This region consists only of the scaffold material, which is usually less stiff than the natural ECM and will therefore result in inferior bulk mechanical properties. This situation is not ideal, and therefore, we aimed at engineering cartilage with the ECM not only in the pericellular region, but also in the interterritorial region. We call this homogeneous, since the matrix is not only around the cell, but also more diffuse in the tissue construct. This situation is one important step closer to the ultimate goal, which is a situation in which there is abundant stiff interterritorial ECM and less stiff pericellular ECM, but which we cannot create as yet. Engineering constructs with inhomogeneity mimicking the in vivo situation is a topic of further investigation in which we possibly need to culture our constructs longer and add stimuli such as mechanical loading to stimulate tissue synthesis, deposition, and remodeling in the interterritorial region.
A disadvantage of constructs without or with low amounts of supporting scaffold material is that they are initially mechanically very weak and will break down as a result of mechanical loading, which is an essential trigger for stimulation of cartilage growth in tissue-engineering approaches.1,8,26,42 A possible solution to circumvent this problem is to not apply loading before the ECM network within the construct has sufficiently developed. 43 This way the beneficial effects of using no or low amounts of agarose and application of mechanical loading could be combined, which will likely improve the quality of tissue-engineered cartilage and make it favorable for in vivo cartilage repair studies.
It is worthwhile to translate the results of our study to possible in vivo outcomes after implantation. Constructs cultured without or with low agarose concentrations in the presence of TGF-β show superior mechanical properties over the other conditions. In order for implanted constructs to withstand the demanding load-bearing environment of the joint, these improved mechanical properties of the implant will be beneficial. However, it should be taken in account that a considerable in vitro preculture period is necessary for the initial weak constructs to reach mechanical stability. The ultimate goal is a balance between sufficient mechanical properties by improved tissue distribution and adequate porosity to stimulate diffusion and tissue integration, which should be further established. 44
In conclusion, the results of this study highlight first the significance of culturing chondrocytes in constructs with no or low-concentration agarose, because it stimulates formation of a more homogeneous ECM distribution. Second, this study shows that besides stimulation of matrix synthesis, the presence of TGF-β3 is also beneficial because it stimulates distribution of matrix components. Both stimuli result in constructs with improved mechanical properties.
Footnotes
Acknowledgments
This study was supported with funding from the Dutch Technology Foundation STW (VIDI-EGT.07970).
Disclosure Statement
No competing financial interests exist.