
Abstract
In Regenerative Medicine, platelet by-products containing factors physiologically involved in wound healing, have been successfully used in the form of platelet-rich plasma (PRP) for the topical therapy of various clinical conditions since it produces an improvement in tissue repair as well as analgesic effects. Measurement of endocannabinoids and related compounds in PRP revealed the presence of a significant amount of anandamide, 2-arachidonoylglycerol, palmitoylethanolamide, and oleoylethanolamide. Investigation of the activity of PRP on the keratinocyte cell line NCTC2544 in physiological and inflammatory conditions showed that, under inflammatory conditions, PRP induced in a statistically significant manner the production of these compounds by the cells suggesting that PRP might induce the production of these analgesic mediators particularly in the physiologically inflamed wounded tissue. Studies in a mouse model of acute inflammatory pain induced by formalin injection demonstrated a potent antinociceptive effect against both early and late nocifensive responses. This effect was observed following intrapaw injection of (1) total PRP; (2) lipids extracted from PRP; and (3) an endocannabinoid-enriched lipid fraction of PRP. In all conditions, antagonists of endocannabinoid CB1 and CB2 receptors, injected in the paw, abrogated the antinociceptive effects strongly suggesting for this preparation a peripheral mechanism of action. In conclusion, we showed that PRP and PRP lipid extract exert a potent antinociceptive activity linked, at least in part, to their endocannabinoids and related compound content, and to their capability of elevating the levels of these lipid mediators in cells.
Introduction
Based on this concept, platelet by-products in the form of platelet-rich plasma (PRP) have been extensively used for the topical therapy of various clinical conditions, such as musculoskeletal injuries, 1 wounds, and soft tissue injuries. 2 Major fields of application are sports medicine, orthopedics, dentistry, dermatology, ophthalmology, and plastic and maxillofacial surgery.3–7
Despite the extensive investigations performed on the growth and angiogenic factor content of PRP,1,8–10 little is known about the pathways involved in its activity during the repair/regeneration phase of wounded tissues. In a recent publication of ours, we investigated the mechanism of action of platelet releasate (PL) on the keratinocyte cell line NCTC2544, and showed that the PL enhanced the physiological early inflammatory response of the tissue to the wound by increasing the activation of P38 and nuclear factor kappa B (NF-κB) pathways, leading to more rapid tissue repair. 11
It has been described that PRP, in addition to its effect on tissue repair, also causes pain suppression.2,12–15 As these observations were made mostly in the course of clinical treatments, there are no reports about the possible factors involved in pain suppression by PRP. In the present study, the possible mechanisms of the analgesic effect of PRP were investigated. We considered the involvement of the endocannabinoid system because of its known role in pain control.16,17 First, we searched for the presence of endocannabinoid in the PRP preparations, then we evaluated a possible activity of PRP at inducing the production of these factors involved in pain control by cultured cells. For this study, we chose the NCTC2544 keratinocytic cell line because of our previous demonstration of the PL-induced repair in these cell monolayer scratch wound models indicating a good response of this cell line to platelet factors. 11 It is to note that keratinocytes are now considered to play an important role in pain perception and are involved in the analgesic actions of endocannabinoids. 18 In the present article, we report that, like PL, 11 also PRP, by far the platelet by-product most used for clinical applications, induced production of IL-8 and neutrophil gelatinase associated-lipocalin (NGAL) and activation of the NF-κB pathway in the NCTC2544 cell line.
We determined the content of endogenous cannabinoid receptor agonists and related mediators in PRP and we found a significant amount of the endocannabinoids anandamide (AEA) and 2-arachidonoylglycerol (2-AG), and of the AEA-related mediators, palmitoylethanolamide (PEA), and oleoylethanolamide (OEA). Furthermore, we showed that, in NCTC2544 keratinocytes, under inflammatory conditions, PRP was further elevating the levels of these mediators in a manner statistically significant, thus suggesting their possible involvement in PRP-induced pain control and wound healing. Indeed, studies in a model of acute inflammatory pain in mice showed a potent analgesic effect of PRP and allowed us to propose that such effect is exerted, at least in part, via endocannabinoid-related mechanism(s).
Materials and Methods
Platelet-rich plasma
PRP was prepared according to Zaky et al. 19 with slight modifications. In brief, PRP was obtained from healthy human blood donors from the Blood Transfusion Center of the San Martino Hospital, Genoa, Italy, in consensus with the guidelines of the institutional ethics committee. PRP was used at a final platelet concentration of 1×107 platelets/μL. The PRP was collected, frozen, and stored at −80°C until use. To minimize variations between donors, PRP preparations used for this study were derived from pooled PRP of at least 10 donors.
Cell culture
Keratinocytes from the cell line NCTC2544 were purchased from the Interlab Cell Line Collection (ICLC) of the Biological bank and cell factory (National Institute for Cancer Research—IST). NCTC keratinocytes were cultured as recommended by the cell line developer in the Eagle's minimum essential medium (MEM) with the Hank's balanced salt solution, supplemented with 2 mM glutamine, 100 U/mL penicillin, 100 μg/mL streptomycin (all from Euroclone), and 10% fetal bovine serum (Lonza) in a CO2 incubator at 37°C, 95% humidity, and 5% CO2.
Western blot analysis
Confluent keratinocytes were incubated for 24 h in a serum-free basal medium supplemented with one of the following conditions: (1) PRP 5%; (2) IL-1α (250 U/mL; Peprotech); (3) PRP 5%+IL-1α (250 U/mL); (4) serum-free medium with no supplements (control). Heparin (500 U/mL) was added to every cell culture dish before treatment. Cells were then extensively washed with phosphate-buffered saline (PBS) to remove residual stimulant factors, including PRP, and incubated in a serum-free medium for an additional 24 h after which, media were collected for Western blot analysis.
Electrophoresis was performed in reducing conditions using 25–50 μg of protein loaded on a 4%–12% NuPAGE Bis-Tris gel (Invitrogen) as described. 11 Images were then scanned using the Epson perfection 1260 scanner (Epson) and band densities were quantified using the ImageJ software.
NF-κB activity assay
For the NF-κB activity assay, binding of the NF-κB p65 subunit to the NF-κB binding consensus sequence 5′-GGG ACTTTCC-3′ was measured with the ELISA-based Trans Am NF-κB kit (Active Motif) using whole cell lysates as described. 11 Preparation of cell extracts was done as per the manufacturer's instructions. Each experiment was assayed in quadruplicate.
Cell treatment with PRP for endocannabinoid determination
Confluent keratinocytes were treated by using the following culture conditions: (1) serum-free medium with no supplements (control); (2) PRP 5%; (3) IL-1α (250 U/mL) (Peprotech); and (4) PRP 5%+IL-1α (250 U/mL). To all culture media, also, 500 U of heparin/mL was added. After incubation, cold methanol was added (over ice) at a 1:1 ratio with a cell culture medium. Cell layers and media were then collected by scraping. Dishes were washed with another 2 mL of methanol, which were combined with collected cells and media for endocannabinoid determination. For each culture condition, samples were collected at T0 and T24 h.
Endocannabinoid extraction, purification, and analysis
Cells+media in methanol were homogenized in 5 volumes of chloroform/methanol/Tris HCl 50 mM (2:1:1) containing 20 pmol each of d 8 -AEA, d 4 -PEA, d 4 -OEA, and d 5 -2-AG. Homogenates were centrifuged at 13,000 g for 16 min (4°C), the aqueous phase plus debris was collected and extracted again twice with 1 volume of chloroform. The organic phases from the three extractions were pooled and the organic solvents evaporated in a rotating evaporator. Lyophilized samples were then stored frozen at −80°C under nitrogen atmosphere to be analyzed later.
Lyophilized extracts were resuspended in chloroform/methanol 99:1 by volumes. The solutions were then purified by open bed chromatography on silica as described in.17,20 Fractions eluted with chloroform/methanol 9:1 by volume (containing AEA, 2-AG, PEA, and OEA) were collected, the excess solvent evaporated with a rotating evaporator, and aliquots analyzed by isotope dilution liquid chromatography/atmospheric pressure chemical ionization/mass spectrometry carried out under conditions described previously 21 and allowing the separation of 2-AG and AEA, PEA and OEA. Mass spectrometric detection was carried out in the selected ion monitoring mode using m/z values of 356 and 348 (molecular ions +1 for deuterated and undeuterated AEA), 304 and 300 (molecular ions +1 for deuterated and undeuterated PEA), 330 and 326 (molecular ions +1 for deuterated and undeuterated OEA), and 384 and 379 (molecular ions +1 for deuterated and undeuterated 2-AG). For 2-AG, the areas of the peaks corresponding to 1(3)-and 2-isomers were added together. The amounts of the compounds were expressed as picomoles/milligram of lipid extract.
For some experiments, the PRP was extracted using the same protocol described above for the cells, to quantify its content of AEA, 2-AG, OEA, and PEA, and the amounts of the compounds were expressed as picomoles/mL of PRP or picomoles/mg of lipid extract. One milliliter of PRP contained ∼7.3 mg of lipid extract.
Finally, for other experiments, the PRP extract, obtained without deuterated standards, was prepurified as described above, to obtain a fraction enriched in AEA, 2-AG, OEA, and PEA to be tested in the mouse model of acute inflammatory pain.
Mouse model of acute inflammatory pain
The experimental procedures applied in the test were approved by the Animal Ethics Committee of the Second University of Naples, Italy. Animal care was in compliance with the IASP and European Community guidelines on the use and protection of animals in experimental research (E.C. L358/1 18/12/86). All efforts were made to minimize animal suffering and to reduce the number of animals used. Mice received formalin (1.25% in saline, 30 μL) in the dorsal surface of one side of the hind paw. Formalin injection induces biphasic stereotypical nocifensive behavior. 22 Nociceptive responses are divided into an early, short lasting first phase (0–7 min) caused by a primary afferent discharge produced by the stimulus, followed by a quiescent period and then, a second, prolonged phase (15–60 min) of tonic pain. Each mouse was randomly assigned to one of the experimental groups (n=8–10), placed in a Plexiglas cage and allowed to move freely for 15–20 min. A mirror was placed at a 45° angle under the cage to allow full view of the hind paws. Lifting, favoring, licking, shaking, and flinching of the injected paw were recorded as nociceptive responses. The total time of the nociceptive response was measured every 5 min and expressed as the total time of the nociceptive responses in minutes (mean±SEM). Recording of nociceptive behavior commenced immediately after formalin injection and was continued for 60 min. The version of the formalin test we applied is based on the fact that a corelational analysis showed that no single behavioral measure can be a strong predictor of formalin or drug concentrations on spontaneous behaviors. We considered that a simple sum of time spent licking plus elevating the paw, or the weighted pain score, is in fact superior to any single (lifting, licking, shaking, and flinching) measure (r ranging from 0.75 to 0.86). Groups of 8–10 animals per treatment were used with each animal being used for one treatment only.
Before the formalin injection, experimental groups were treated with (1) total PRP; (2) a PRP lipid extract, and an endocannabinoid-enriched fraction of this extract (both reconstituted in dimethyl sulfoxide [DMSO] in the same volume of the total starting PRP [1 mL], and then injected after 1:10 or 1:100 dilution in PBS); (3) a mixture of synthetic AEA, 2-AG, OEA, and PEA in amounts identical to those found in the active dose of the PRP extract. All these preparations, that is, total PRP, PRP extract, and a mixture of synthetic compounds were in each case, administered into the paw in 10 μL of saline, 10 min before formalin. Data were compared to those obtained in animals treated only with formalin+vehicle (saline or DMSO in saline). In some experiments, cannabinoid receptors of type-1 (CB1, AM251; Tocris) and cannabinoid receptors of type-2 (CB2, AM630; Tocris) receptor antagonists were also administered intraperitoneally (i.p.) 10 min before the above preparations, or locally in the paw, 10 min before the above preparations, and 20 min before formalin. A transient receptor potential vanilloid type-1 (TRPV1) antagonist 5′ iodo-resiniferatoxin (I-RTX; Tocris) was also administered locally following the same procedure as the CB1 and CB2 antagonists. The assay is based on the immediate acute response to formalin; thus, the treatment with PRP was normally performed before formalin injection, to allow for the conditioning of the tissue. However, since after the first phase of the acute response there is a quiescent phase, and then a second phase of tonic pain, due, among other things, to sensitization, PRP was also injected just before the beginning of the quiescent phase, that is, 7 min after formalin injection. This allowed us to detect the possible activity of PRP in reducing the pain specifically in the second phase and after the first phase has finished, that is, before the occurrence of sensitization.
Statistical analysis
All data are presented as means and standard deviations. Statistical analysis was performed online using the unpaired Student's t-test by means of the GraphPad software (www.graphpad.com).
Results
PRP induces a proinflammatory response in resting NCTC2544 keratinocytes, which is intensified in an inflammatory environment
In a recent article, we reported that the lysate of purified platelets strongly increases the inflammatory response induced by IL-1α, by enhancing the activation of the p38 and NF-κB pathways, and the synthesis of IL-8 and of the antibacterial lipocalin NGAL. However, since the preparation normally used for clinical applications is PRP, that is, a concentrate of platelets in serum, we tested on NCTC2544 keratinocytes also the PRP activity. We confirmed the strong induction of IL-8 and NGAL, which again was further enhanced in an inflammatory environment mimicked by treatment of cells with IL-1α. In addition, as in the case of cells stimulated by the lysate, we observed that in the presence of PRP, the NF-κB pathway presented an additive effect with regard to the activation induced by IL-1α.
Figure 1A shows a Western blot of IL-8 and NGAL, released in the medium by the cells in the different culture conditions. The amount of secreted proteins were quantified (±SD) by the densitometric analysis of six determinations from three different experiments performed in duplicate dishes (Fig. 1B). The induction of IL-8 was statistically significant versus the control for all conditions p=0.0001. The treatment with IL-1α+PRP resulted in an induction of IL-8 significantly higher than the ones caused by PRP (p=0.031) and by IL-1α (p=0.002) alone. The induction of NGAL was statistically significant versus the control for all tested conditions, but the effect of IL-1α+PRP was not significantly different from that of IL-1 or PRP alone.
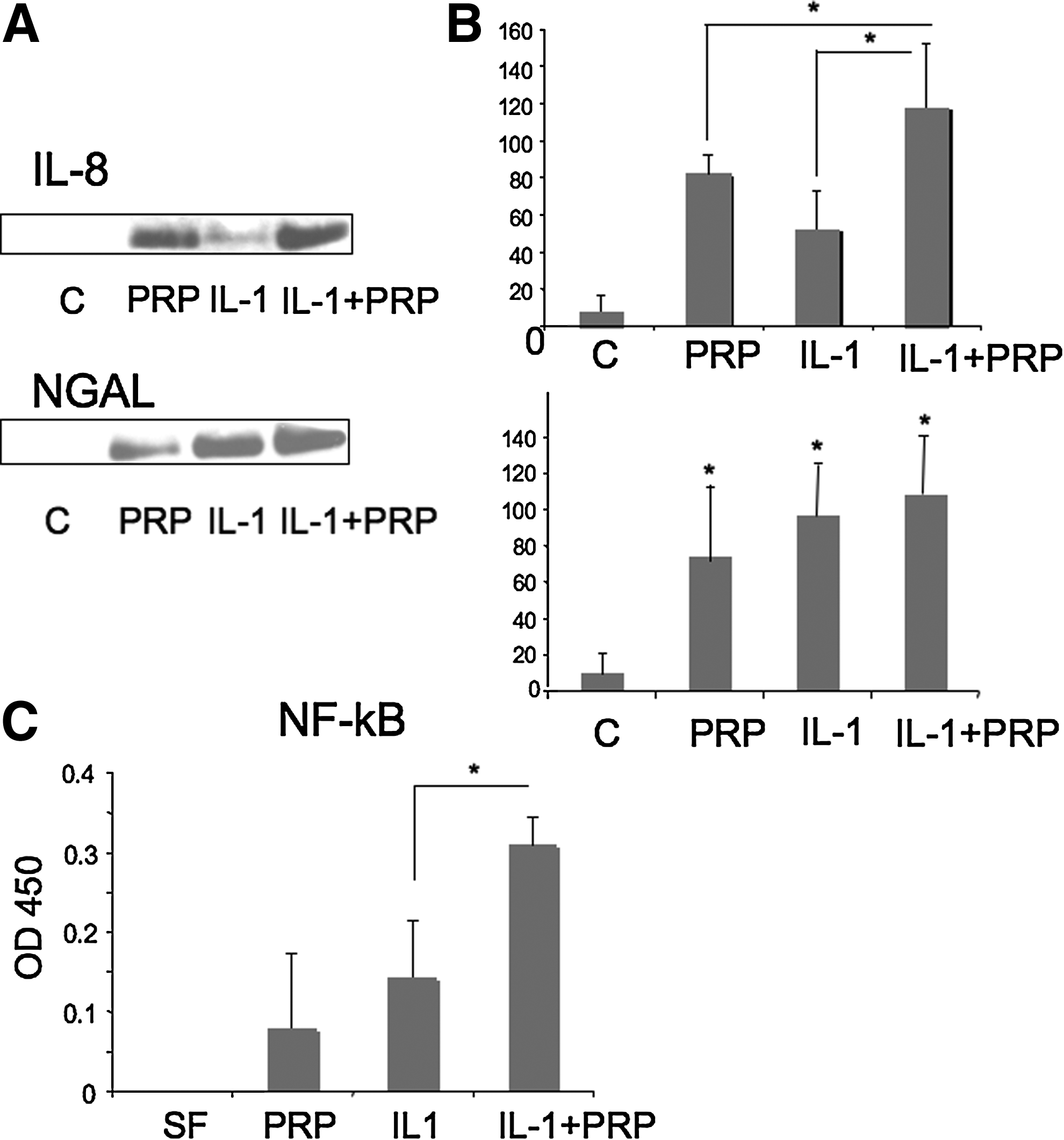
Platelet-rich plasma (PRP) effect on NCTC2544 keratinocytes in physiological and inflammatory conditions. Cells were incubated with either serum-free medium, 5% PRP, IL-1α (250 U/mL), or IL-1α+5% PRP.
Figure1C shows the activation of NF-κB by IL-1α and PRP and the additive effect observed with PRP+IL-1α. The average of three experiments analyzed in quadruplicate is shown with SD. The effect of IL-1α+PRP was significantly different from that of IL-1α alone (p=0.0001).
PRP contains a mixture of endocannabinoids and related mediators
Since an effect of PRP in pain suppression has been reported in different clinical applications,2,12–15 we measured the concentration of endocannabinoids and related analgesic mediators in two distinct PRP preparations and detected a significant amount of AEA, 2-AG, PEA, and OEA (Table 1).
AEA, amount of the endocannabinoids anandamide; 2-AG, 2-arachidonoylglycerol; PEA, palmitoylethanolamide; OEA, oleoylethanolamide.
PRP enhances the production of endocannabinoids and related mediators by cultured keratinocytes
We studied the effect of PRP on endocannabinoid production by NCTC2544 cells. Cells were treated with PRP in physiological and inflammatory-like conditions at T0 and T24 h and the amount of endocannabinoids was measured in the lipid extract. Figure 2 reports the variation in the levels of different endocannabinoids detected in the lipid extract after 24 h of cell treatment in the different culture conditions. For each condition, the amount of endocannabinoids present at T0 was subtracted from the 24 h value. The reported amount represents the mean (±SEM) of two independent experiments performed in quadruplicate (n=8). In PRP-treated NCTC2544 cells, the amount of AEA and 2-AG were increased in a statistically significant manner in inflammatory-like conditions (p=0.011 and 0.012, respectively) (Fig. 2A, B). Instead, in the same cells, PRP per se induced a strong elevation of the OEA level, which remained constant in inflammatory conditions (Fig. 2C). In both conditions, the increase was statistically significant with respect to the control (p=0.0002 for PRP and p=0.024 for IL-1+PRP). A different scenario was found for PEA, which was not increased after the cell treatment with PRP (Not shown).
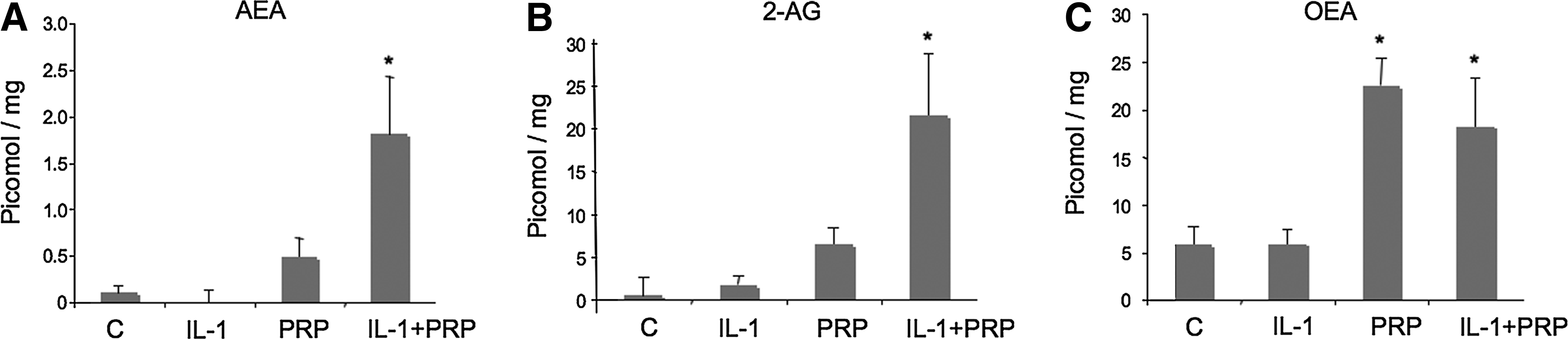
Induction of endocannabinoids and related compounds by treatment of NCTC2544 keratinocytes with PRP in physiological and inflammatory conditions. Cells were cultured in the presence of serum-free medium, 5% PRP, IL-1α (250 U/mL), or IL-1α+5%PRP. Cells+medium were extracted at T0 and T24 h for endocannabinoid and related compound content determination. T0 values were subtracted from T24 h values.
PRP presents in vivo a potent antinociceptive activity related to endocannabinoid receptor activation
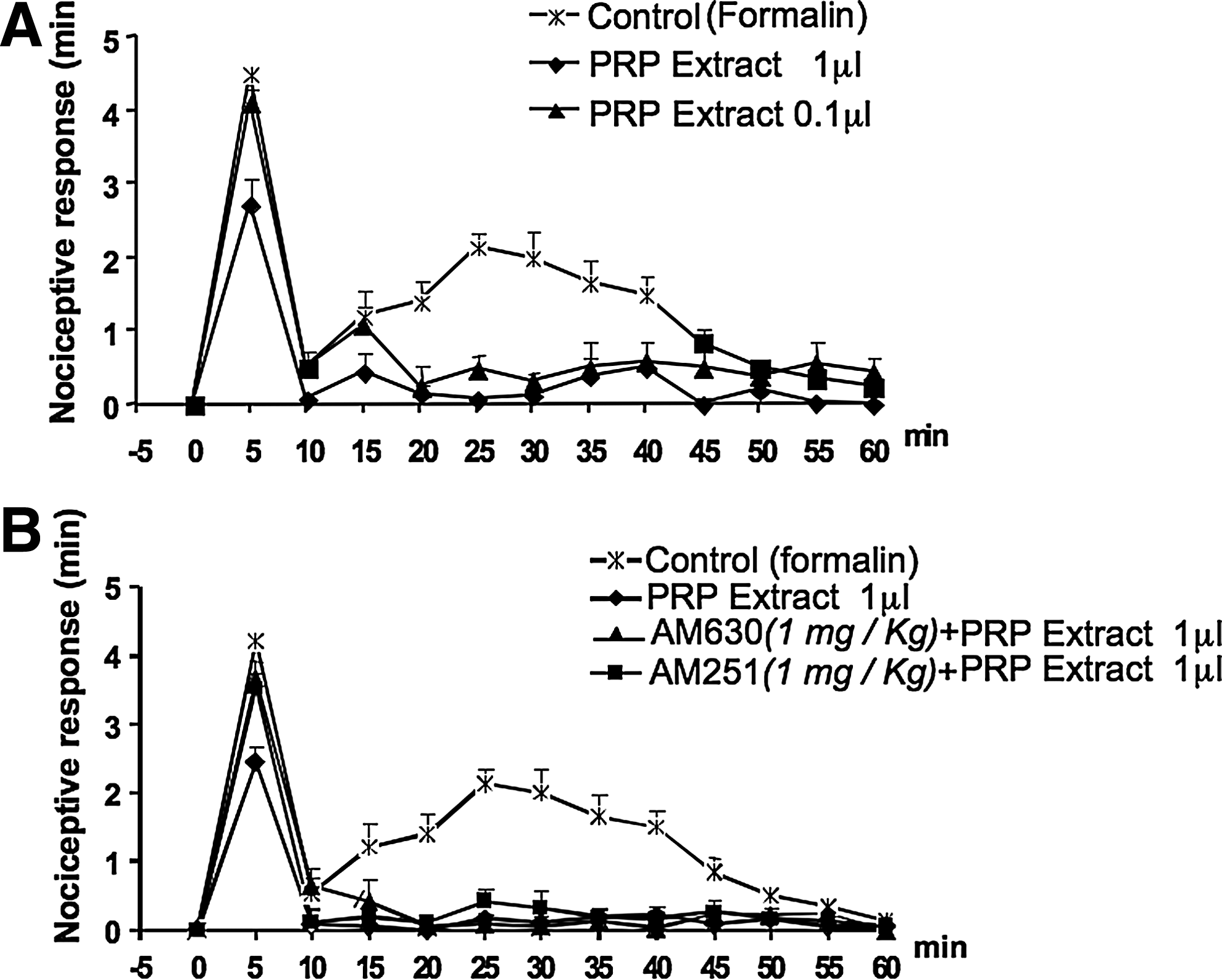
Next, it was of interest to investigate whether or not PRP displayed any analgesic activity in an in vivo model, and we chose the formalin-induced acute inflammatory pain test in mice. The effect of PRP on the nociceptive behavior of mice after subcutaneous formalin (1.25%, 30 μL) injections into the hind paws was investigated. We found that local administration of PRP, administered 10 min before, reduced the nociceptive behavior induced by formalin in a dose-related manner (Fig. 3A); 1 μL of PRP injected in 10 μL of saline (approximately containing 4.2, 20.7, 71.8, 35.0 fmol of AEA, 2-AG, PEA, OEA, respectively, see above), reduced by 42% the early response and completely abolished the late response. To establish whether the endocannabinoids found in PRP were involved in the antinociceptive response, the selective CB1 and CB2 receptor antagonists (AM251and AM630, respectively), were administered either systemically (i.p.) or locally (into the paw) 10 min before PRP injection. Both CB1 and CB2 receptor antagonists, when administered systemically (1 mg/kg, i.p.), only blocked the effect of PRP on the first phase (Fig. 3B), whereas when administered locally (30 μg/10 μL), abolished the antinociceptive activity of PRP on both the early and late nocifensive phases of the formalin test (Fig. 4B). I-RTX, an antagonist of TRPV1 channels, which are activated by PEA and OEA, also partly reduced the effect of PRP on both phases when injected locally (0.15 μg/10 μL) (Fig. 4B). Injection of the antagonists per se did not cause any effect (data not shown and Fig. 4A).

Antinociceptive effect of PRP on the effect of subcutaneous formalin injection into the hind paws of mice. Formalin (1.25%, 30 μL) was injected in the absence (control) or in the presence of PRP (administered 10 min before formalin).
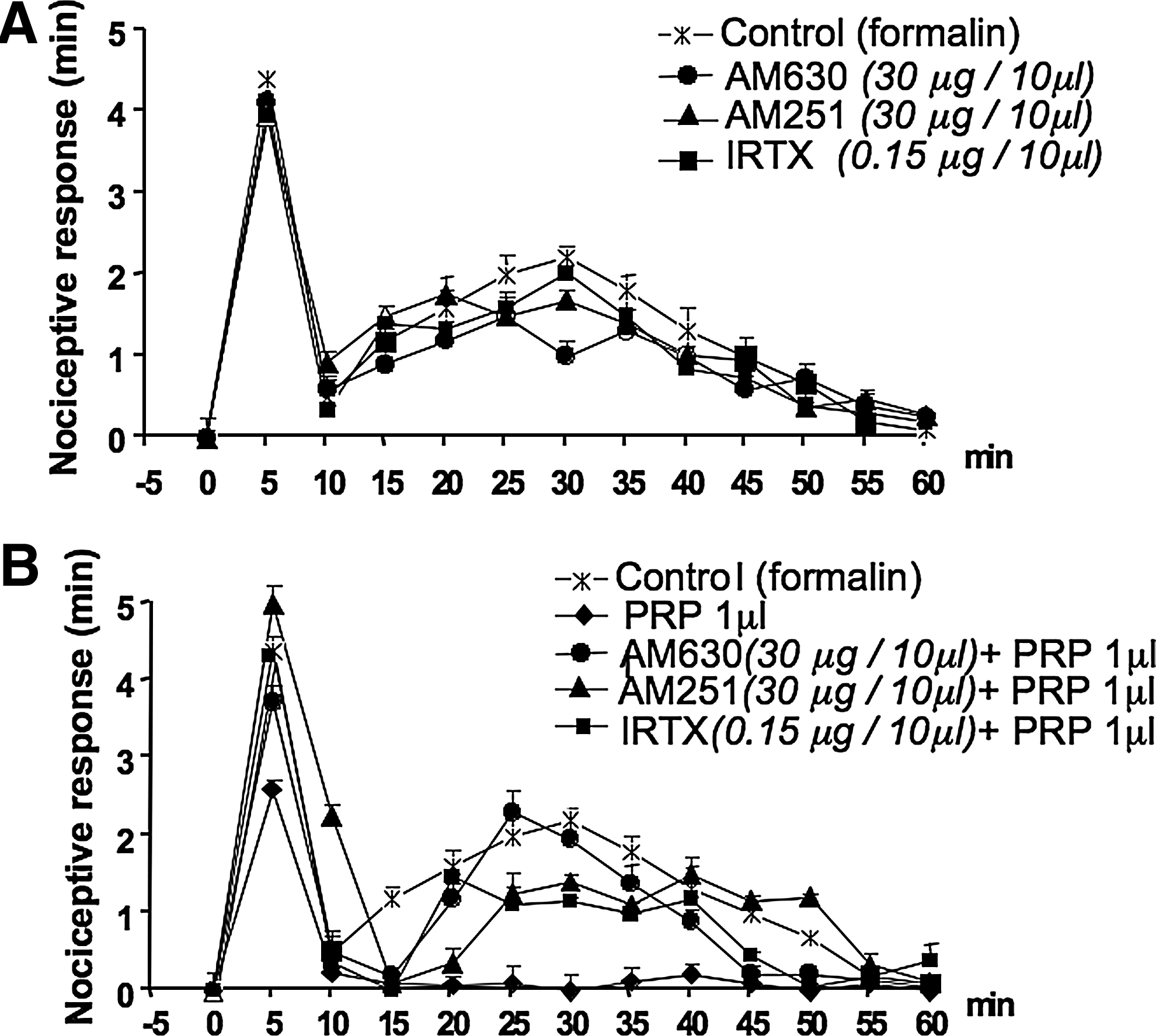
Effects of locally injected cannabinoid CB2, CB1 receptor antagonists and transient receptor potential vanilloid type-1 (TRPV1) receptor antagonist on the antinociceptive response of PRP on subcutaneous formalin injection. Panel
The same experiments were performed using a PRP lipid extract.
Figure 5A and B show a dose response of the PRP lipid extract per se or after enrichment in endocannabinoids and related compounds by purification by silica gel chromatography. Both the total extract and the purified extract were active on the late, but not on the early, phase of the nocifensive response and, unexpectedly, were more active at low than at high concentrations, that is, at the dose of 2.5 μL (injected in 10 μL saline) of a solution obtained by dissolving in 1 mL saline raw or purified extracts from 1 mL of PRP. Indeed, the activity decreased at higher doses.
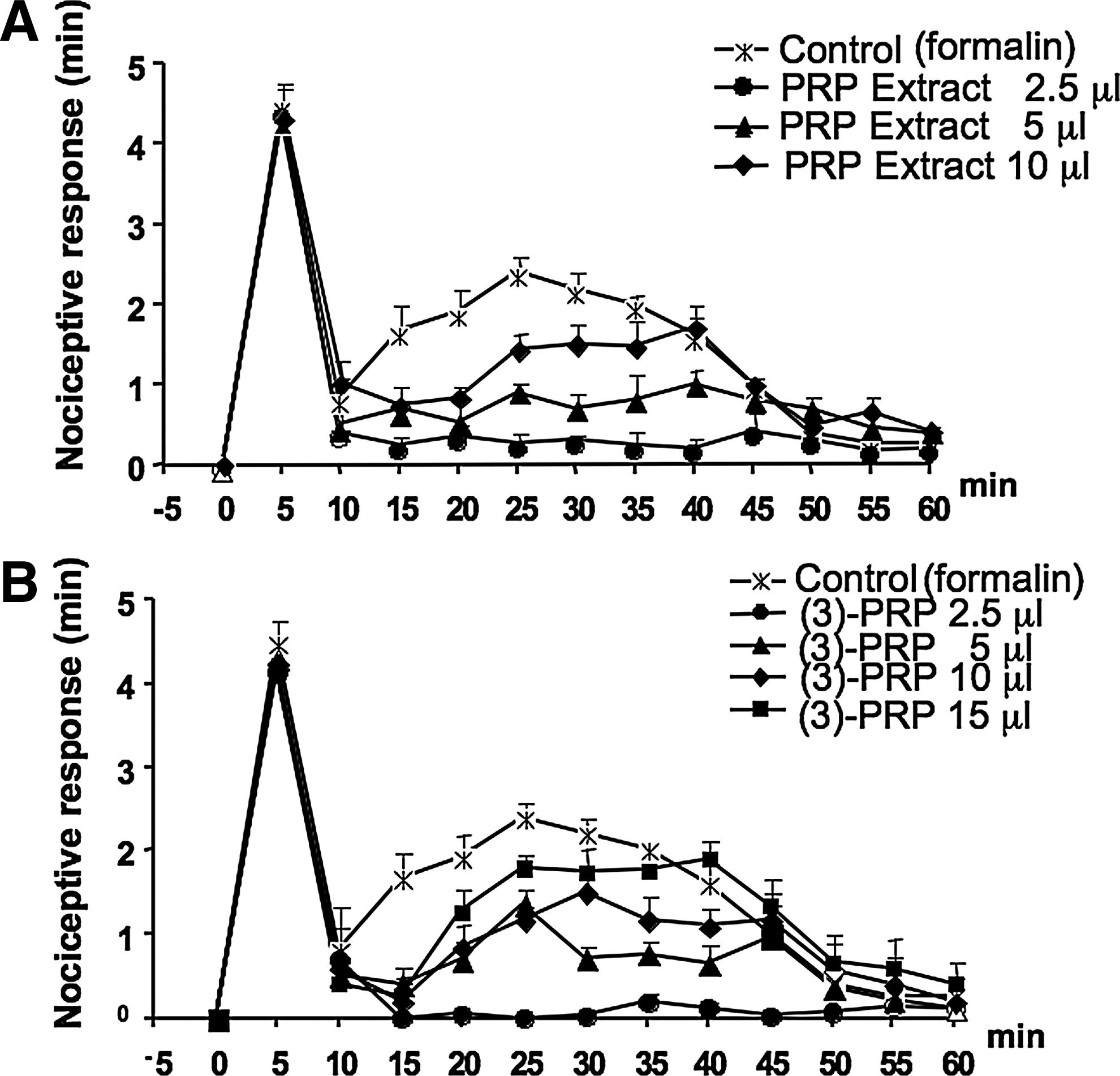
Antinociceptive response of PRP lipid extract and endocannabinoid-enriched fraction on the effect of subcutaneous formalin injections into the hind paws of mice. Formalin (1.25%, 30 μL) was injected in the absence (control) or in the presence of PRP lipid extract (2.5, 5, 10/10 μL saline)
Because of this finding, we repeated the experiments with lower concentrations of the unpurified lipid extract. In this case, we dissolved the lipid extract from 1 mL of PRP in 1 mL DMSO, and then diluted this solution 10-fold with PBS. Figure 6A shows the dose response for the injected doses of 1 and 0.1 μL (referred to the initial solution, that is, 10 and 1 μL of the 10-fold diluted solution) in 10 μL PBS. The 1 μL dose (containing in principle the same amounts of AEA, 2-AG, PEA, and OEA as in 1 μL of PRP, that is, 4.2, 20.7, 71.8, and 35.0 fmol) reduced by ∼40% the early response and completely abolished the late response. Another experiment performed with the extract initially dissolved in PBS instead of DMSO gave the same results (not shown).
Antinociceptive response of PRP lipid extract and the effect of systemically administered cannabinoid CB1 and CB2 receptor antagonists. Panel
To establish whether the endocannabinoids and related mediators in PRP lipid extract were involved in this antinociceptive response, the selective CB1 and CB2 receptor antagonists, AM251 and AM630, and the TRPV1 antagonist, I-RTX, were again administered either systemically (1 mg/kg, i.p.) or locally (30 μg/10 μL or 15 μg/10 μL) in the paw 10 min before PRP injection. As in the case of the animals treated with PRP, when given systemically, the CB1 and CB2 antagonists reversed only the effect of the lipid extract (1 μL/10 μL saline) on the first phase (Fig. 6B). However, again, when administered locally, all the antagonists, that were inactive per se (Fig. 7A), blocked the effect of the extract on the first phase, and only the CB2 and TRPV1 antagonists attenuated the effect of PRP extract on the second phase (Fig. 7B).
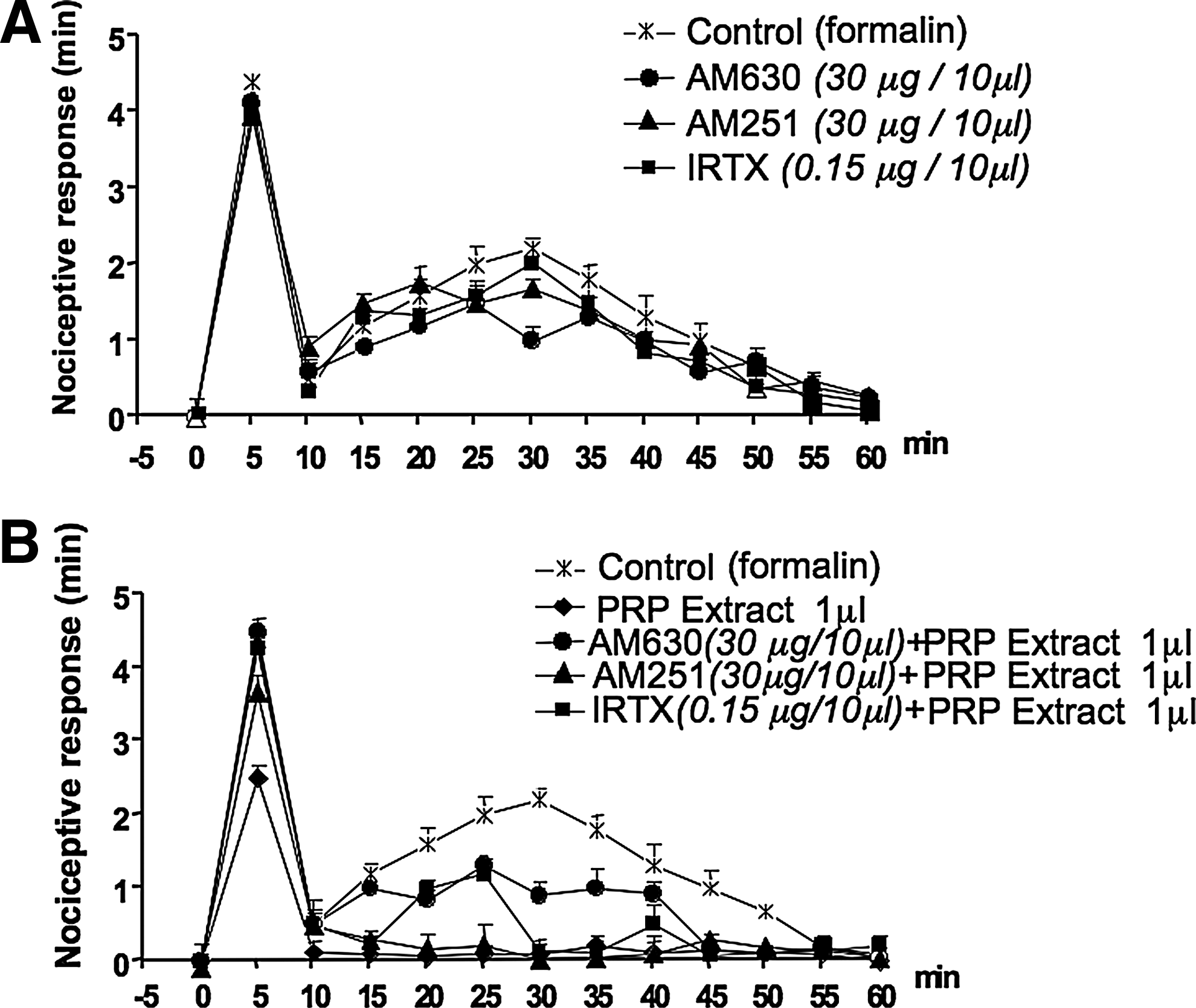
Effects of locally injected cannabinoid CB2, CB1 receptor antagonists and TRPV1 receptor antagonist (AM630, AM251, and I-RTX, respectively), on the antinociceptive response of PRP lipid extract on subcutaneous formalin injection. Panel
A mixture of endocannabinoids and related mediators presents an antinociceptive activity similar, but not identical, to that of PRP in vivo
A mixture of the endocannabinoids and related compounds was performed according to the content of the same compounds in 1 mL of PRP and, hence, containing 4.2, 20.7, 71.8, and 35.0 pmol di AEA, 2-AG, PEA, and OEA, respectively. This was dissolved in 1 mL PBS, and its activity was measured in the formalin test. The dose of 1 μL (containing the same fmol of AEA, 2-AG, PEA, and OEA), injected in 10 μL of saline, reduced the late phase of the nociceptive behavior induced by formalin, but did not show any activity on the early phase (Fig. 8A). This effect was attenuated by locally administered CB1 and CB2 receptor antagonists (30 μg/10 μL, paw), but not by a TRPV1 antagonist (0.15 μg/10 μL, paw) (Fig. 8B).
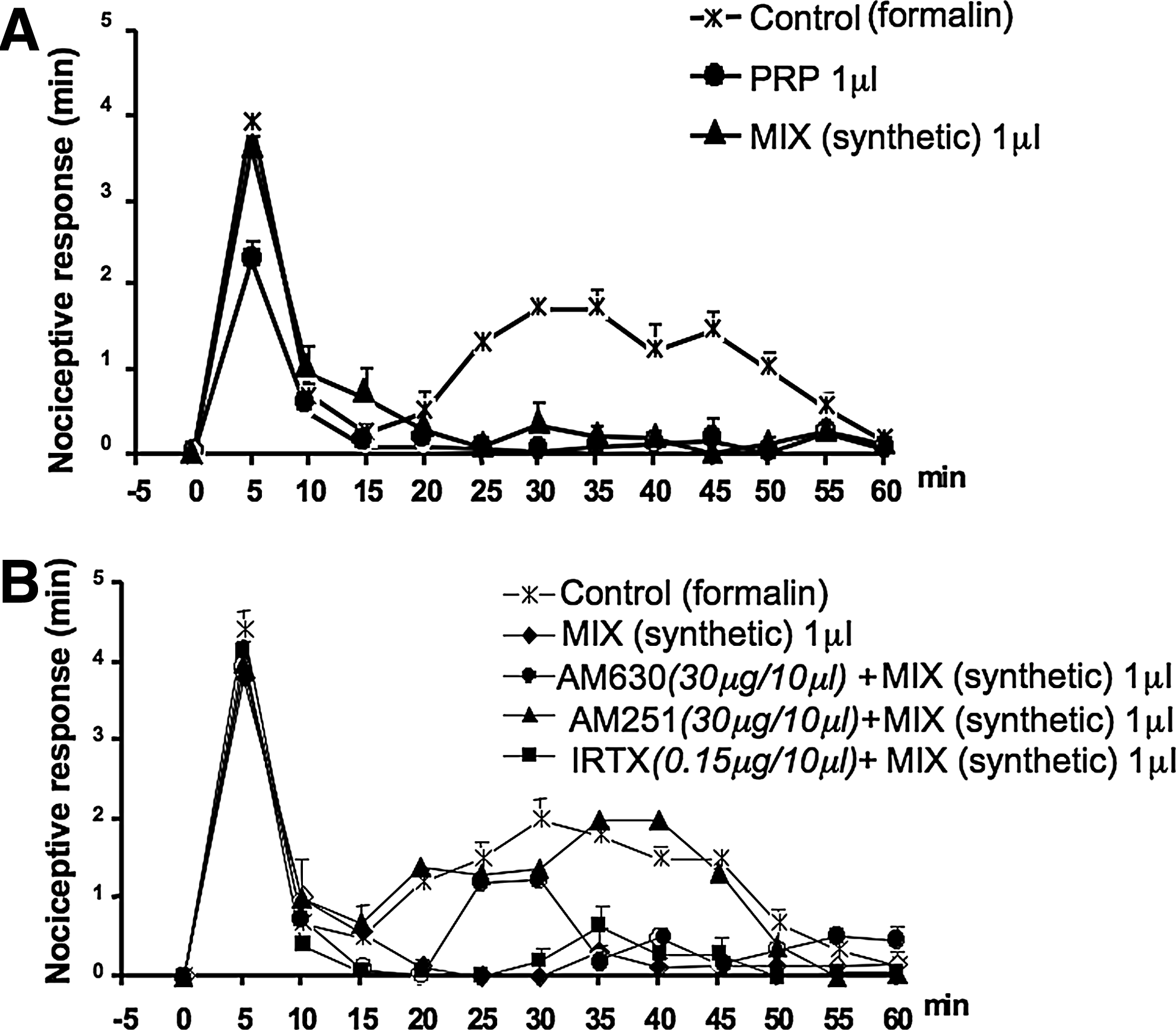
Antinociceptive effect of a mixture of synthetic amount of the endocannabinoids anandamide (AEA), 2-arachidonoylglycerol (2-AG), palmitoylethanolamide (PEA), and oleoylethanolamide (OEA) on the effect of subcutaneous formalin injection into the hind paws of mice. A mixture of the endocannabinoids and related compounds was prepared in the same amount and ratio of the same compounds in 1 mL of PRP and, hence, containing 4.2, 20.7, 71.8, and 35.0 pmol di AEA, 2-AG, PEA, and OEA, respectively. This was dissolved in 1 mL PBS and its activity was measured in the formalin test. The dose of 1 μL diluted to 10 μL with saline, and containing 4.2, 20.7, 71.8, and 35.0 fmol of AEA, 2-AG, PEA, and OEA, respectively, was injected in the paw. Panel
PRP and PRP lipid extract activity on the second phase of the response to formalin
In the experiments described above, the treatment with PRP was performed before formalin injection, to allow a conditioning of the tissue, and also because the response to formalin is immediate and transient. However, since after the first phase of acute response there is a quiescent phase, and then a second phase of tonic pain, which is due, among other things, to sensitization, we also injected PRP just before the start of the quiescent phase, to detect a possible antinociceptive activity of this preparation by selectively interfering with sensitization processes. As shown in Figure 9A, under these conditions, PRP at the concentration of both 0.1 and 1 μL/10 μL PBS did not reduce significantly the second phase of the nociceptive response except for the 15-min time point, whereas the PRP extract at the concentration of 1 μL/10 μL of PBS, was effective at reducing the response only in the first 25 min of the second phase (Fig. 9B).
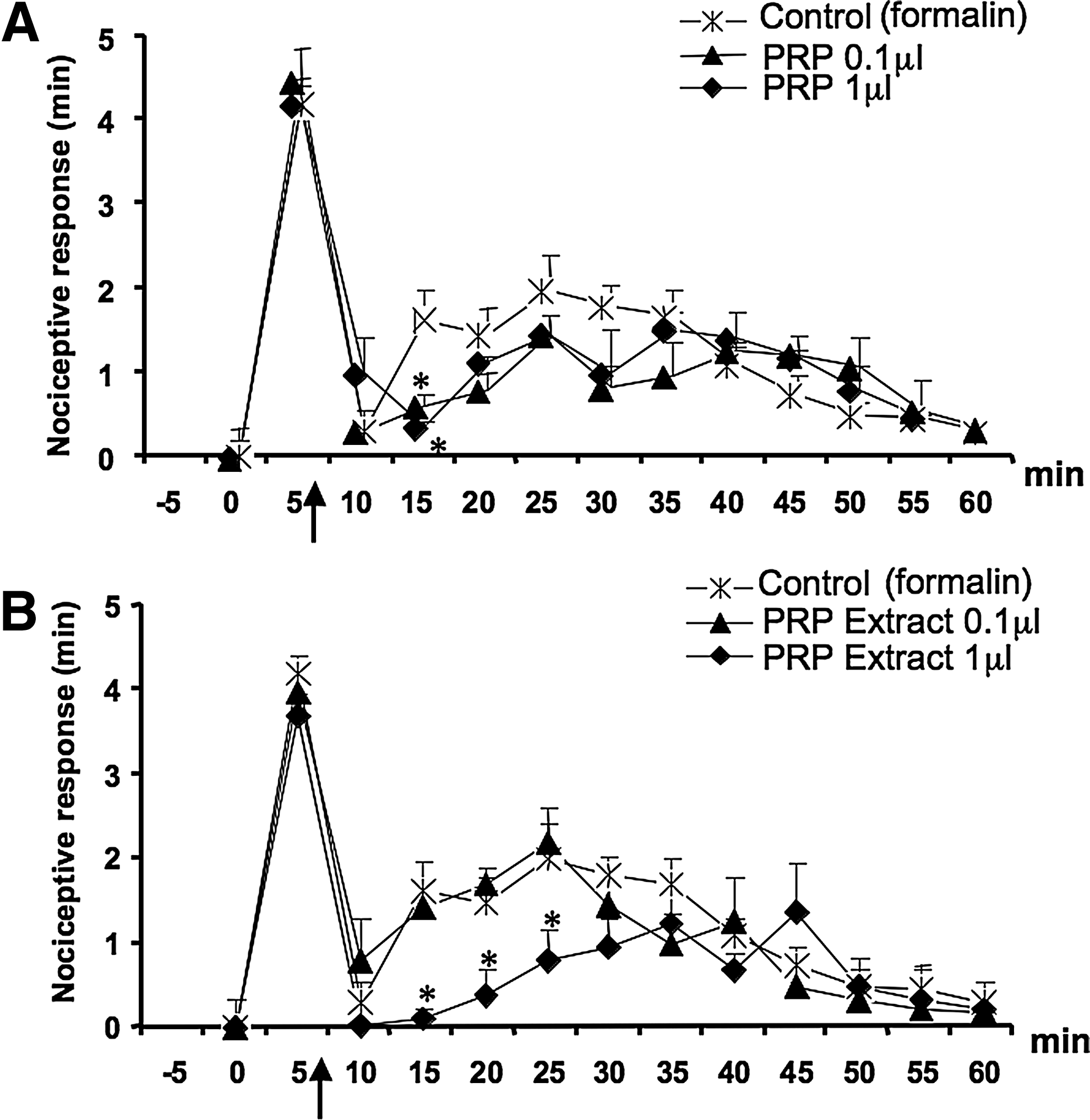
Effect of PRP and PRP lipid extract on the second phase of the nociceptive response to formalin. PRP and PRP lipid extract were locally administered 7 min after formalin injection (arrow) to detect a possible specific antinociceptive response on the second phase. Panel
Discussion
Local applications of platelet gels have been proposed for the treatment of various clinical conditions, such as musculoskeletal injuries, 1 wounds, and soft tissue injuries. 2 Recently, PRP injections have been used for the treatment of osteoarticular diseases, such as osteoarthritis (hip, knee) and recalcitrant epicondylitis.12–15 These literature articles reported that PRP reduced pain, decreased the symptoms, and improved the functions of the organs. On this background, it is very important to investigate all the aspects of this complex preparation that appears to be so important for tissue repair following insults and wounding. In particular, it is important to investigate about the pathways activated by PRP trying to dissociate the plethora of activities, opening possibilities also in the field of pharmacology for developing compounds able to activate specific pathways physiologically activated in tissue repair. In this respect, it is crucial to investigate the pharmacological activities underlying the analgesic actions of PRP and dissect them from those that are instead involved in other pharmacological aspects of this preparation. This effort, if successful, would open new avenues also in the field of pharmacology, for the development of more standardized compounds able to activate those pain control pathways specifically activated during tissue repair. In a recent publication of ours, we described the effect of a lysate obtained from purified platelets (PL) on the keratinocyte cell line NCTC2544, and showed that PL-increased wound healing suggested that PL increases the physiological response of the tissue to the wound, leading to more rapid tissue repair and providing a defense to bacterial infection. In the present article, we studied the effects of PRP, the platelet preparation, including also plasma and membranes that is commonly used for clinical applications, on the same cells. We focused on the possible mechanism involved in the described analgesic property of this preparation12–14 postulating a role for the endocannabinoid system, because of its known function in pain control.16,17 Endocannabinoids, such as AEA and 2-AG, are fatty acid derivatives that act as endogenous agonists of the G-protein-coupled receptors for the major Cannabis psychoactive constituent, Δ 9 -tetrahydrocannabinol, and known as CB1 and CB2. 23 Both cannabinoid receptors and endocannabinods participate in several ways in the control of pain, and do so at the peripheral, spinal, and supraspinal level. 17 Furthermore, the endocannabinoid AEA is often produced in cells together with two congeners, PEA and OEA, which do not act directly via cannabinoid receptors, but can produce analgesic and anti-inflammatory effects via several other mechanisms, including the activation and subsequent desensitization of TRPV1 channels. 24
Our present results can be summarized as follows.
(1) We first characterized the response of the cultured keratinocytes to PRP in physiological and inflammatory-like conditions and proved that the PRP, like the PL, induced in these cells the production of IL-8 and of NGAL and the activation of the NF-κB pathway. These effects appeared to be stronger in Il-1α-treated cells, suggesting an enhancement of wound healing by the inflammatory phase.
(2) We observed that PRP-treated keratinocytes exhibited a concomitant increase of AEA, 2-AG, and OEA production, which was again stronger in IL-1α-treated cells, suggesting that PRP might induce the production of these analgesic mediators particularly in the physiologically inflamed wounded tissue. This is an interesting finding that correlates with the activation of NF-κB by PRP. Indeed, it was reported that AEA was induced in macrophages by LPS by a mechanism involving NF-κB activation. 25 After the initial proinflammatory stimulation, PRP enhances tissue repair.11,26 Since endocannabinoids were described to have also an anti-inflammatory activity, 27 it is also possible to hypothesize the existence in keratinocytes of a pro-resolving loop involving endocannabinoids.
(3) We then determined the content of these mediators in the PRP and found that they were present together with the other OEA congener, PEA, in significant amounts (although lower than those found in keratinocytes after stimulation with PRP).
(4) Next, we investigated whether PRP displayed any activity in an in vivo model of nociception and we chose the formalin-induced acute pain test in mice. The effect of PRP on the nociceptive behavior of mice after subcutaneous formalin injection into the hind paw was investigated. First, we found that the local administration of PRP before formalin reduced the early response of the nociceptive behavior induced by formalin by 42%, and completely abolished the late response. Then, to establish whether the endocannabinoids found in PRP were involved in the antinociceptive response, we coadministered with PRP selective CB1 and CB2 receptor antagonists (AM251and AM630, respectively), or a TRPV1 antagonist (I-RTX). We observed that, especially when locally injected in the paw, these antagonists abolished the antinociceptive activity of PRP both on the early and late phase of the response to formalin. The stronger attenuation of PRP effects observed with local versus systemic injection of the antagonists strongly suggests for this preparation a peripheral mechanism of action. Finally, a PRP lipid extract and a fraction of the extract enriched in endocannabinoids and related compounds showed a dose-related antinociceptive activity, and locally injected CB1, CB2, or TRPV1 receptor antagonists abolished the antinociceptive activity of the former preparation both on the early and late phases of the response. It should be noted that the activity of the extract was maximal in the range of 0.1–2.5 μL in 10 μL of vehicle, and decreased at higher concentrations, somehow resembling typical bell-shaped dose–response curves often observed with endocannabinoids. 28
Interestingly, PRP and the PRP lipid extracts were active at doses containing very low concentrations of endocannabinoids and related compounds, that is, in the order of the 10–100 fmol/10 μL of vector, which is below the active concentrations reported in the literature for each of these compounds. However, one should not forget that, as shown here, PRP is capable of inducing elevation of endocannabinoid levels in keratinocytes. If this phenomenon, which might be due to nonendocannabinoid constituents of PRP, occurs also in other cell types in the mouse paw, it is possible that the local concentrations of these mediators after injection of PRP are much higher than those found in PRP per se. Furthermore, it has been previously reported that N-acylethanolamines like AEA and PEA, synergize at producing antinociceptive effects in the formalin test in mice. 29 Therefore, synergistic effects among AEA, 2-AG, OEA, and PEA might also explain why antinociceptive effects were observed here with amounts of PRP and PRP lipid extract containing very low concentrations of these compounds. Indeed, we found here that a mixture of synthetic AEA, 2-AG, OEA, and PEA, in the same amounts as those contained in the maximally effective dose of the PRP lipid extract, also produced antinociceptive effects in the second phase of the formalin test, and in a way attenuated by antagonists of CB1, CB2, or TRPV1 receptors. However, the effects observed with this preparation were qualitatively different from those seen with PRP in terms of efficacy on the two phases of the formalin response. Indeed, it is likely that both PRP and PRP lipid extract contain other constituents that might modulate nociception via cannabinoid receptor- and noncannabinoid receptor-mediated mechanisms. This possibility is supported by the observation that similar, although not identical, sensitivity to different antagonists was observed with PRP, PRP lipid extract, and the artificial mixture of AEA, 2-AG, OEA, and PEA. In addition, since the second phase of tonic pain is due among others, to sensitization mechanisms, we also injected the PRP just before the beginning of the quiescent phase to detect the possible activity of this preparation at specifically counteracting these mechanisms, independently from the first phase, which causes sensitization. The PRP extract was effective at reducing the response in the first 25 min of the second phase. These data suggest that PRP also directly affects sensitization mechanisms, and that the effects seen on the second phase in the experiments in which PRP was administered before formalin were not merely due to interference with nociceptive mechanisms occurring during the first phase. We would like to point out that the finding that PRP contains endocannabinoids and is able to induce further endocannabinoid level elevation in cell cultures opens the possibility of a specific involvement of these lipid mediators in tissue repair in addition to pain control. Future studies should be aimed at investigating the composition of other possible lipidic components active in the PRP and also at investigating whether endocannabinoids and related mediators can underlie also some of the wound healing11,25 therapeutic properties of PRP. This last possibility is supported by the previous finding that cannabinoids improved colonic epithelial 29 and periodontal30,31 wound healing, and that cannabinoid receptors were overexpressed at the site of skeletal muscle wounds. 32
Conclusion
We reported here that PRP, a platelet product widely used in the clinic, has a potent antinociceptive effect and owes at least a part of its analgesic properties to peripheral endocannabinoid-mediated mechanisms.
This is the first report of a specific analgesic effect of both total PRP and PRP lipid extract in an animal model. The finding that the lipid extract has a potent antinociceptive activity is especially relevant and makes this material suitable for analgesic preparations to be used in different possible clinical applications.
Footnotes
Acknowledgments
We acknowledge Dr. Paolo Strada for providing platelet blood fractions and Dr. Niels Borregaard for the gift of anti-NGAL monoclonal antibodies. We would also like to appreciate Dr. Maddalena Mastrogiacomo and Anita Muraglia for valuable suggestions and providing PRP preparations. This work was partially supported by funds from the European Commission FP7 (Angioscaff and GAMBA projects) and from Regione Liguria (P.O.R. 2007–2013).
Disclosure Statement
We wish to confirm that there are no known conflicts of interest regarding this publication and that there has been no financial support that could have influenced its outcome. F.D., V.U., R.C., F.P., L.L., F.P., L.G., S.M., V.D., state no conflict of interests. However, R.C. wants to acknowledge that he is a shareholder of the company Biorigen Srl, a university spin off whose mission is the R&D in the Regenerative Medicine field with particular focus on skeletal tissues.