
Abstract
Skeletal muscle atrophy has been well characterized in various animal models, and while certain pathways that lead to disuse atrophy and its associated functional deficits have been well studied, available drugs to counteract these deficiencies are limited. An ex vivo tissue-engineered skeletal muscle offers a unique opportunity to study skeletal muscle physiology in a controlled in vitro setting. Primary mouse myoblasts isolated from adult muscle were tissue engineered into bioartificial muscles (BAMs) containing hundreds of aligned postmitotic muscle fibers expressing sarcomeric proteins. When electrically stimulated, BAMs generated measureable active forces within 2–3 days of formation. The maximum isometric tetanic force (Po) increased for ∼3 weeks to 2587±502 μN/BAM and was maintained at this level for greater than 80 days. When BAMs were reduced in length by 25% to 50%, muscle atrophy occurred in as little as 6 days. Length reduction resulted in significant decreases in Po (50.4%), mean myofiber cross-sectional area (21.7%), total protein synthesis rate (22.0%), and noncollagenous protein content (6.9%). No significant changes occurred in either the total metabolic activity or protein degradation rates. This study is the first in vitro demonstration that length reduction alone can induce skeletal muscle atrophy, and establishes a novel in vitro model for the study of skeletal muscle atrophy.
Introduction
Experimental models of disuse muscle atrophy employ physical means to initiate the atrophy process. In both animal (e.g., hindlimb suspension, limb immobilization, tenotomy, and denervation) and human (e.g., limb suspension, limb immobilization, and bed rest) models of muscle atrophy10,14,20–23 it is difficult, if not impossible, to isolate specific mechanical factors from the other influences that exist in the in vivo environment. Given the multifactorial nature of the skeletal muscle atrophy process, it would be desirable to have the ability to isolate individual factors that may lead to muscle atrophy. Since tension reduction appears to be a major factor leading to disuse muscle atrophy, a model that enables studying the effects of tension reduction alone on muscle atrophy would be highly beneficial.
Unfortunately, ex vivo-explanted muscle studies are normally limited by the short viability of explanted muscle tissue. 24 Advancements in tissue-engineering techniques have allowed the culturing of three-dimensional skeletal muscles (bioartificial muscle [BAM]) that have many in vivo characteristics and can be maintained in vitro for extended periods of time. 25 Tissue-engineered muscles are contractile and have functional characteristics similar to in vivo muscle.26–28 Mechanical stimulation of the tissue-engineered skeletal muscle has been shown to induce hypertrophy in BAMs 29 and spaceflight microgravity directly induces BAM myofiber atrophy in vitro. 13 While a two-dimensional in vitro muscle fiber starvation atrophy model has been reported, 30 no three-dimensional or tension-related in vitro model of skeletal muscle atrophy has been previously described. In the present study, tissue-engineered primary mouse BAMs were induced to atrophy by length shortening and tension reduction. This model system can be used to study specific mechanotransduction pathways that lead to skeletal muscle atrophy and possibly identify future drug targets to prevent or treat this condition.
Materials and Methods
Cell culture and tissue-engineering techniques
Primary mouse myoblasts (PMM) were isolated from 6-week-old male C3HeB/FeJ mice and individual myoblast clones isolated by standard dissection techniques as described previously. 31 The animal protocol was approved by the Miriam Hospital Institutional Animal Care and Use Committee. The cells were expanded in a supplemented growth medium (SGM) (1:1 DMEM [Gibco; #11995-040]: FGM [Cambrex; #3130] containing 20% fetal bovine serum [FBS; Gibco], 1% ITS+1 [Sigma], and 1% penicillin/streptomycin [p/s]) in collagen-coated 100-mm tissue culture dishes. The SGM contained 2 μg/mL insulin and 400 ng/mL of the human fibroblast growth factor (FGF). The cells were harvested when 80%–100% confluent with 0.25 mg/mL trypsin, pelleted by centrifugation at 1000 rpm for 5 min, resuspended in the SGM and either replated, used for BAM engineering, or cryopreserved.
BAM formation
Harvested myoblasts were tissue-engineered into BAMs containing 1×106, 2×106, or 3×106 cells each, as previously described. 31 Briefly, myoblasts were suspended in 0.4 mL of a 4°C solution of 1 part Matrigel™ (BD Biosciences):6 parts type I rat tail collagen (BD Biosciences) solution (1.6 or 0.8 mg/mL collagen in SGM) in 10×25-mm silicone molds with two stainless steel (ss) pins positioned 20 mm apart in the base of the molds. The molds were glued to the bottom of 35-mm-diameter Petri dishes. Solutions with double or half the concentration of Matrigel were also tested. Once the mixture was poured into the molds, they were placed in a humidified 37°C, 5% CO2 incubator for 8–10 h, and then flooded with the SGM (4 mL/well). Over the next 24–48 h, the cell/gel mixture contracted away from the mold and was suspended in place by the pins (Fig. 1A). Longitudinal tension in the cell/gel mixture resulted in parallel alignment of the myoblasts and their fusion into parallel arrays of postmitotic muscle fibers (Fig. 1C). BAMs were fed the SGM every 2 to 3 days. The BAMs were incubated for 3 days with 1 mg/mL cytosine arabinoside (araC) beginning days 3 and 7 postcasting, and weekly thereafter, to prevent the attenuation of force generation resulting from the continued proliferation of unfused myoblasts (data not shown). BAM tissue engineering was optimized based on maximizing the maximum isometric tetanic force (Po) by varying the number of cells plated during casting and the concentration of collagen and Matrigel. A combination of 2×106 cells/BAM in a suspension containing 1 part Matrigel:6 parts 1.6 mg/mL collagen solution was found to result in maximum Po generation (data not shown).
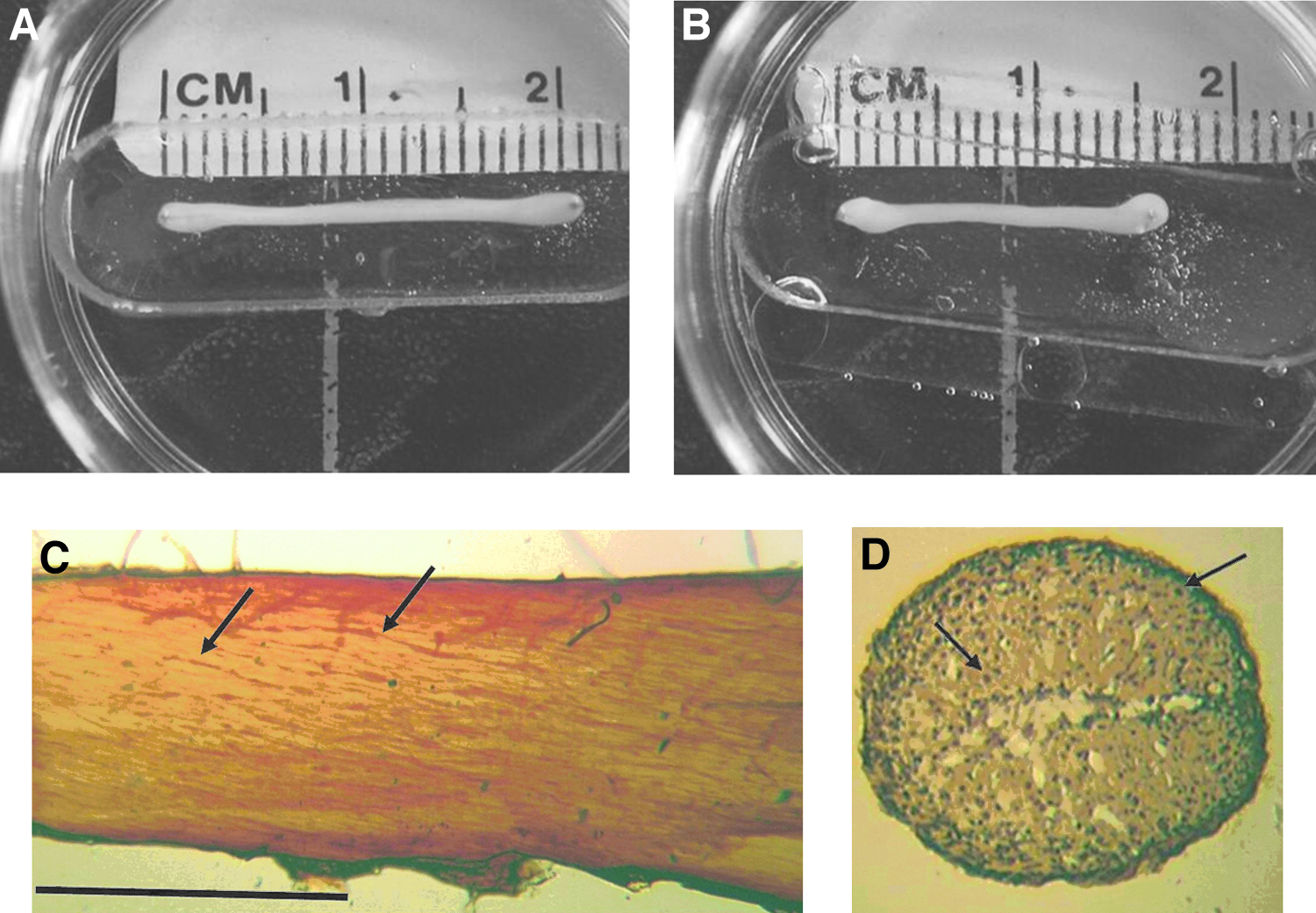
Bioartificial muscle (BAM) morphology. Representative 22-day-old control
BAM force measurements
BAM contractile properties were determined using a custom-designed force transducer system calibrated with a force generator as previously described. 26 Contractile properties were measured by attaching one of the two ss pins attached to the BAMs to the load arm of the force transducer, and then lifting the pin out of the silicone mold, while the other ss pin remained attached to the mold. Passive and active force measurements were recorded with an IBM-based computer-data acquisition system (LabVIEW software; National Instruments). BAMs were electrically stimulated via two platinum electrodes 4 mm apart on either side of the BAM. The voltage and stimulus were controlled using a custom-designed LabVIEW program. Preliminary studies were performed on 1- to 4-week-old BAMs to determine the requirements for peak twitch force generation by varying voltages and pulse durations. Peak twitch force (Pt) could be obtained by applying a 40 V, 4-ms-wide pulse, and Po was obtained by applying an electrical stimulus of 40 V at 40 Hz with 4-ms-wide pulses. These parameters were therefore used in subsequent studies.
BAM excitability was determined by measuring rheobase50 (R50, the voltage required to cause a twitch force 50% of Pt) and chronaxie50 (C50, the pulse width that gives 50% of Pt at a voltage of twice the rheobase). BAM R50 was determined by giving twitch stimulations at 4 ms and varying the voltage from 0 to 50 V. Three measurements were made at each voltage. For BAM C50, twitch stimulations at 2×R50 were used at increasing pulse widths from 0 to 4 ms. The force–frequency curves were determined by plotting force against frequency during tetanic stimulations. The length–force curves were generated by determining both passive and peak twitch forces (40 V, 4 ms) for BAMs at 1-mm length increments ranging from −20% to +35% of the starting BAM length. The BAMs were allowed 1 min to recover between stimulations. One-minute rest times were determined to be adequate for the BAMs to fully recover their maximum Pt and Po for multiple stimulations (data not shown).
Length reduction
One day before shortening, BAMs were switched from the SGM to a standard maintenance medium (MM) containing 20% FBS and 1% p/s in DMEM. The shortening was carried out by manually bringing in one ss pin toward the other (Fig. 1B). The amount of shortening was 25% to 50% of the original length of 20 mm (i.e., final length of 15 and 10 mm, respectively). Media changes with either the SGM or MM±araC were made every 2 to 3 days. On the final day of the experiment, the BAMs were fed fresh media, lengthened to their initial length, and electrically stimulated to measure Po as described above. BAMs were then washed with ice-cold phosphate-buffered saline (PBS) and processed for histological and biochemical analyses.
Biochemical assays
BAMs were thawed and sonicated in a 300 μL sucrose buffer (0.25 M sucrose, 0.02 M KCl, pH 6.8) with seven 2-s bursts, while on ice. Fifty microliter aliquots were removed for noncollagenous protein determination. Noncollagenous proteins were extracted by the method of Lowry et al. 12 and the protein concentration determined using the BCA Protein Assay Kit (Pierce). 13
Total protein synthesis rates were determined at varying times after BAM shortening (6 h, 24 h, 3 days, and 7 days) by pulse labeling for 6 h in a tissue culture medium containing 1 μCi/mL 14-C phenylalanine (Amersham). The BAMs were rinsed and processed for radioactivity incorporated into cellular proteins as previously described. 13 Total protein degradation rates were determined by incubating a second group of BAMs for 40 h before shortening in a medium containing 1 μCi/mL 14-C phenylalanine. The BAMs were extensively rinsed, shortened, and media aliquots collected 1, 2, 6, 7, 9, and 10 days later. Radioactivity released into the medium was measured and radioactivity remaining in cellular proteins calculated as described previously. 13 Radioactivity was measured on a Packard Tricarb liquid scintillation analyzer (Packard Instruments).
The rate of lactate production by the BAMs was determined by measuring lactate concentrations in aliquots of tissue culture medium collected at varying times after shortening. The lactate measurements were carried out using a YSI Glucose/Lactate Model 2300 STAT Analyzer (Yellow Springs Instruments). The measurements were corrected for the duration of conditioning and volume of medium.
Histology and immunocytochemistry
BAMs were fixed in 3.7% formaldehyde for 45–60 min, paraffin embedded, and cut into 5-μm cross sections. The sections were deparaffinized in Histoclear (Sigma), rehydrated sequentially from 100% EtOH to 70% EtOH, and then dH2O. Localization of sarcomeric tropomyosin was visualized by first blocking the sections with 1% horse serum, 0.5% Tween in PBS (blocking buffer) followed by incubation for 1 h with a monoclonal anti-tropomyosin antibody (Sigma T-9283) diluted 1:100 in PBS containing 0.5% Tween. After rinsing with PBS, the sections were incubated for 30 min with biotinylated anti-mouse IgG (Vector Labs) diluted 1:200 in a blocking buffer, 30 min with the avidin/biotinylated enzyme complex, and then developed with 3,3′-diaminobenzidine (Sigma D5905) for 5–10 min. Sections not incubated with the biotinylated secondary antibody were used as negative controls.
Morphometric measurements
Cross-sectional analyses were made on 5-μm sections and imaged using an image analysis system (KS 300 v3; Carl Zeiss). Myofiber cross-sectional areas (CSA) were manually measured by outlining each myofiber and the CSA calculated by the software. Myofibers were measured from the entire cross-section of each BAM, totaling >100 myofibers/BAM.
Statistical analysis
Statistical analyses and data processing were carried out with SAS Enterprise Guide (version 5.1; SAS Institute, Inc.) and Microsoft Office Excel 2007 (Microsoft). Time-matched one-variable continuous data between 2 groups were compared using a nonparametric one-way analysis of variance (ANOVA) and Kruskal–Wallis test. For comparison of more than two groups of continuous data, a one-way ANOVA with a Bonferoni correction was performed. p<0.05 was accepted as being statistically significant.
Results
Characterization of PMM BAMs
BAMs made by casting an ice-cold collagen/Matrigel cell solution into silicone molds quickly gelled at 37°C and detached from the mold surface, remaining attached only to the pins at each end. The BAMs gradually contracted over several days to form nearly cylindrical structures by day 10 with a mean width and thickness of 1.20 and 0.98 mm, respectively (Fig. 1A). BAMs immunohistochemically whole-mount stained for sarcomeric tropomoysin showed parallel arrays of organized myofibers oriented longitudinally along its long axis (Fig. 1C, arrows). Similarly stained BAM cross sections revealed that myofibers were concentrated primarily along the periphery with fewer fibers located toward the center (Fig. 1D, arrows).
The contractile characteristics of the BAMs were studied by measuring forces generated in response to transverse electrical stimulations from platinum wire electrodes. BAMs generated forces in response to electrical stimulation beginning 36 h postplating, and these forces continued to increase linearly for 3 weeks before plateauing at 2587±502 μN (mean±SD for all BAMs >25 days old). The BAMs remained viable in vitro for greater than 80 days with a minimal loss of force (Fig. 2). Based on the dimensions of BAM cross sections and the calculated CSA of 0.92 mm2, the mean specific Po (sPo) was 2.8±0.54 kN/m2.

Maximum isometric tetanic force (Po) as a function of BAM age. Po was measured for BAMs ranging in age from 1 to 82 days. The values were compiled from 11 separate experiments and each point represents the mean±SD of 3–31 BAMs.
The general appearance of electrically induced twitches, unfused tetani, and fused tetani are shown in Figure 3A–C, respectively. The excitability of the BAMs was determined by applying a series of pulses of variable pulse widths and voltages. The rheobase (R50) and chronaxie (C50) were used as measures of excitability (where lower values indicate greater excitability) and were calculated based on the effect of pulse amplitude (Fig. 4A) and pulse duration (Fig. 4B) on twitch force, respectively. For 30-day-old BAMs, R50 and C50 were 7.0±0.71 V and 0.37±0.12 ms, respectively (n =5 per group). The force-frequency diagram for the BAMs indicated that tetanic forces plateaued between 10 and 20 Hz (Fig. 4C).

Contractile characteristics of BAMs. Representative peak BAM twitch force

Excitability of BAMs.
Passive and peak twitch forces (4 ms, 40 V) were measured at increasing BAM lengths from −20% to +35% of the original length (20 mm, 0% stretch) to generate length–force curves for 2- to 3-week-old BAMs (Fig. 5). Passive forces increased linearly with increased stretch, whereas peak twitch forces reached a peak between +5% to +25% before decreasing with increased lengthening. Shortening of the BAMs beyond −15% of their original length eliminated both passive and active force generation.

Length–tension curve. Representative passive and peak twitch forces (4 ms 40 V) were measured on a 15-day-old BAM at various lengths ranging from −20% to +35% of the original length of 20 mm. Measurements were taken 1 min after each 1-mm incremental change in length. Each point represents three measurements made with a 1-min rest period between stimulations. Similar results were obtained from at least three other BAMs.
Inducing muscle atrophy through length reduction
To determine whether tension release by length reduction could induce myofiber atrophy in vitro, 3- to 5-week-old BAMs were shortened by 25% to 50% for 6 to 13 days (Fig. 1B). Changes in the maximal isometric force contraction, myofiber size, noncollagenous protein content, and total protein synthesis and degradations rates were measured.
Fifty percent BAM length reduction in MM for 6 days significantly decreased Po by 50.4% (n=4–5, p<0.05) (Fig. 6). The R50 of the shortened BAMs in MM increased significantly by 29% to 9.0±1.6 V (n=4–5, p<0.05), indicating reduced fiber excitability. No significant change occurred in C50 with shortening. Similar results were obtained with BAMs of varying ages when the duration of shortening was 6 days or longer, but 30-day-old BAMs shortened for 6 days gave the most reproducible results and, therefore, these parameters were used for subsequent studies.
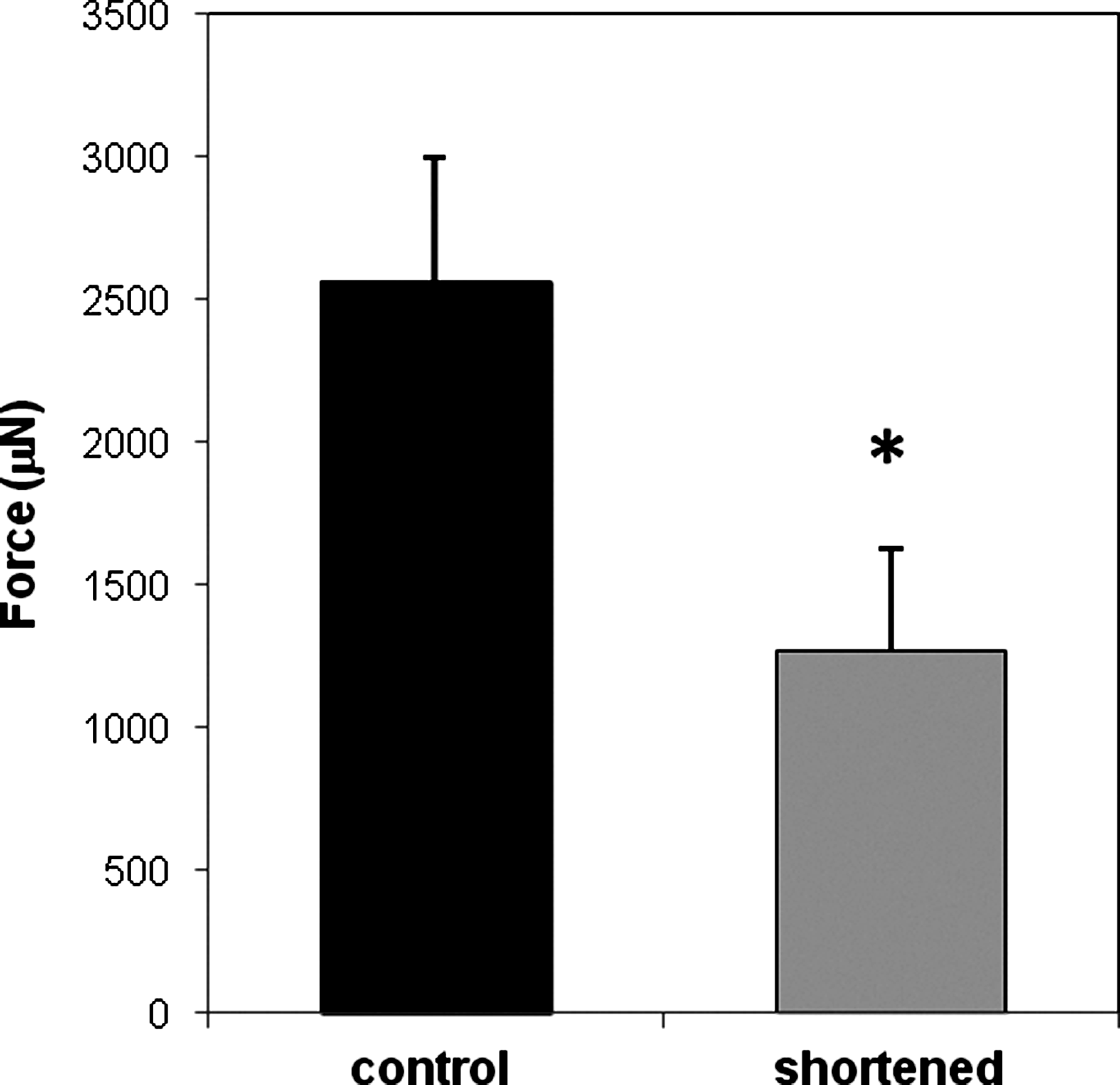
Maximum isometric tetanic force was reduced after BAM shortening and partially attenuated with medium growth factors. Po was measured on 30-day-old BAMs after 6 days of 50% shortening. BAMs were incubated in the maintenance medium. Bars represent the mean±SD of force measurements (n=4–5 per group; *p<0.05).
The rate of BAM metabolism after shortening was determined by collecting tissue culture medium fractions after 6 h of incubation with a fresh medium at 37°C and measuring lactate concentrations. There was no significant difference in the rate of lactate production between the control (57.9±2.6 nmol/day; n=6) and shortened BAMs (58.4±0.9 nmol/day; n=6). BAM shortening resulted in a small, but significant 6.9% (p<0.05) reduction in the total noncollagenous protein content in the BAMs (n=5–6 per group).
Protein synthesis rates were determined based on14-C phenylalanine incorporation into cellular proteins in BAMs during a 6-h pulse labeling period beginning 6 h to 7 days after 50% BAM shortening. Protein synthesis rates were significantly lower (19.0% to 22.0%, p<0.05) for BAMs shortened for 3 days or more compared to control BAMs (Fig. 7A). For protein degradation rates, aliquots of the tissue culture medium were collected over a 10-day postshortening period from BAMs whose proteins had been prelabeled with 14-C phenylalanine. The medium was assayed for released 14-C radioactivity and protein degradations rates calculated as outlined in the Material and Methods section. There were no significant differences at any time point (Fig. 7B), indicating that protein degradation rates were not significantly affected by shortening.
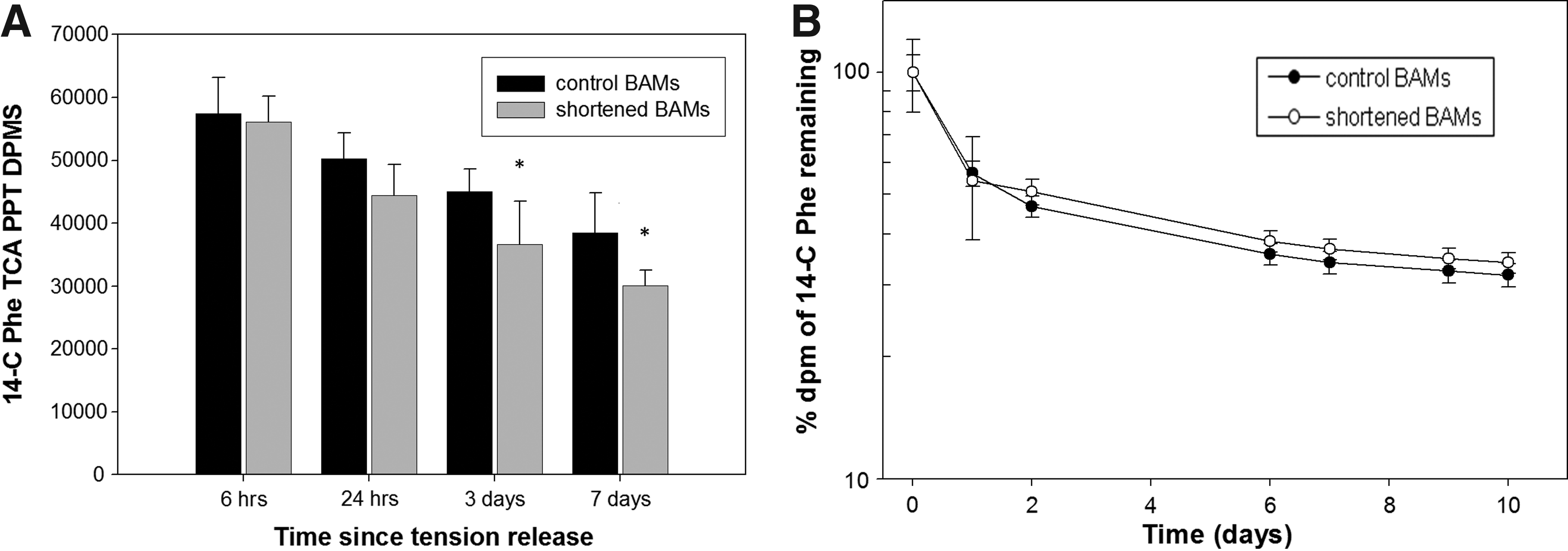
Protein synthesis, but not protein degradation, was reduced after BAM shortening.
Quantitative morphormetric measurements of BAM mean myofiber CSA were performed as described in the Materials and Methods section. In 30-day-old BAMs, 6 days of length reduction resulted in a significant 21.7% (n=4, p<0.05) decrease in mean myofiber CSA (Fig. 8).
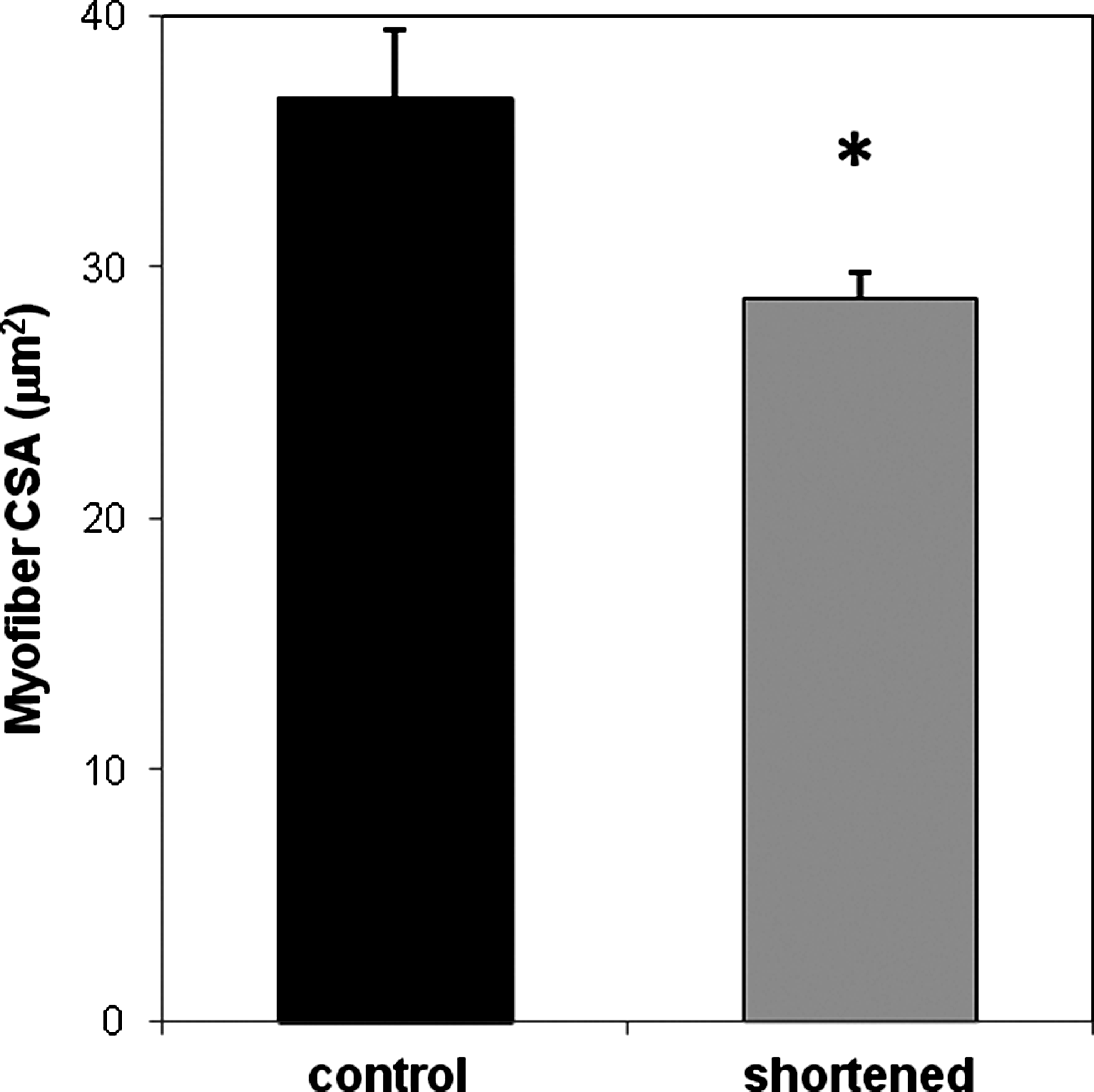
Myofiber cross-sectional areas (CSA) were decreased after BAM shortening. Mean myofiber CSA were determined for 30-day-old BAMs after shortening for 6 days. BAMs were fixed in 3.7% formaldehyde, paraffin embedded, sectioned (5 μm), immunocytochemically stained for sarcomeric tropomyosin, and myofiber CSA measured as described in Materials and Methods section. Bars represent mean±SD of mean CSA (>100 random myofibers/BAM) of four control and four 50% shortened BAMs (*p<0.05).
Discussion
In this study, we have demonstrate that length reduction alone induces atrophy in tissue-engineered skeletal muscle, as measured by decreases in Po, mean myofiber CSA, noncollagenous protein content, and protein synthesis rates. These BAMs generate a Po greater than 2500 μN and have an in vitro viability of more than 80 days.
To the best of our knowledge, Po of 2587±502 μN for our PMM BAMs is greater than any other reported for tissue-engineered skeletal muscle constructs made from mammalian cells. Previously, Dennis et al. 27 showed that myooids made from C2C12 cell lines+fibroblasts, and primary mouse and rat muscle cultures had Po ranging from 115 to 576 μN. In a method using primary rat cultures in a fibrin gel casting, Huang et al. reported tissue-engineered skeletal muscle constructs with a Po of 805.8±55 μN. 32 Avian organoids had an absolute isometric force (passive+active) of 2850 μN and a net Po of 1360 μN. 33 Bian and Bursac engineered porous skeletal muscle tissue constructs using a mesoscopic hydrogel-molding approach that generated twitch force amplitudes as high as 1390±210 μN when exposed to agrin. 34
Although this study was not designed to determine the maximum in vitro duration of BAM survival, it was found that they can remain viable and generate force for greater than 80 days, maintaining Po at a steady-state level long term. There is no reason to suspect that these BAMs could not remain viable for a significantly longer period of time. Neonatal rat, adult rat, and mouse myooids were reported to have lifespans of 52±2.5 days, 74±6 days, and 101±12 days, respectively.26,27 This extended lifespan of tissue-engineered muscle provides a distinct advantage to the use of these constructs for long-term studies over ex vivo-explanted skeletal muscle preparations, which have a limited viability on the order of hours to days. 24 Although BAMs in this study were kept in static cultures with regular medium changes, bioreactors allowing constant perfusion of a fresh medium, such as those successfully utilized to maintain BAMs in experiments designed for spaceflight atrophy studies,13,35 may permit even longer periods of viability.
When corrected for BAM CSA, the sPo of PMM BAMs was 2.80±0.54 kN/m2. Although this sPo is ∼1% of the ∼280 kN/m2 sPo reported for adult mouse muscle, 36 it is similar to those reported for rat myooids (2.9 kN/m2) 26 and avian organoids (0.9 to 3.6 kN/m2), 33 and less than those reported by Dennis and coworkers for mouse myooids (19.2 kN/m2) 27 and by Huang et al. (36.3 kN/m2). 32 The relatively small sPo reported for PMM BAMs is partly due to the fact that, whereas ∼90% of in vivo muscle CSA is occupied by muscle fibers, 37 only a small percentage of the BAM's CSA is composed of myofibers (2%–10%), a figure that is similar to that reported for human BAMs. 29 Myooids bioengineered with only cells and no extracellular matrix had a smaller CSA and a higher density of muscle fibers, thereby generating greater sPo values. 27 To be consistent with sPo reported for individual ex vivo or in situ myofiber preparations, Po should ideally be normalized to the total CSA of only the myofibers in the BAMs. The alternative, calculating sPo for individual PMM BAM myofibers, is currently not technically feasible. The relatively small Po in PMM BAMs and murine myoids compared to in vivo muscle is most likely due to several factors besides myofiber density or tissue size, including low sarcomeric protein content per fiber, embryonic and neonatal isoforms of contractile proteins rather than adult isoforms, and the fact that the type I collagen used to bioengineer the PMM BAMs is stiffer than normal extracellular matrix components formed in situ.
Mean myofiber CSA in the control PMM BAMs of 36.7±2.7 μm2 is consistent with those reported for human BAMs 29 (∼35 μm2) and C2C12 mouse BAMs 38 (∼20 μm2), both of which were bioengineered with collagen/Matrigel as the extracellular matrix material. These were somewhat less than myofiber CSA for avian BAMs/organoids 13 (∼85–140 μm2) and rat myooids 26 (∼80–400 μm2), both of which were made without the use of an extracellular matrix scaffold material.
Based on our evolving, but limited understanding of mechanotransduction mechanisms involved in disuse skeletal muscle atrophy, 11 we hypothesized that in vitro length reduction, hence tension reduction, alone can induce atrophy in tissue-engineered BAMs. With PMM BAM length reduction, we observed a decrease in myofiber CSA, a loss of total muscle proteins, a decrease in tetanic force, and a downregulation of protein synthesis, all of which are hallmarks of skeletal muscle atrophy.
However, some of the observed changes in response to length reduction are modest, particularly reductions in the noncollagenous protein content, protein synthesis rates, and CSA. The decrease in total protein in different muscles in hindlimb-unloaded rodents ranged between 14% and 58% 10 compared to 6.9% for the PMM BAMs. The 22% decrease in protein synthesis rates in the PMM BAMs is lower compared to 59% reduction seen in hindlimb-suspended rats 10 and 48.5% reduction seen in human vastus lateralis muscles after 21 days of bed rest. 39 Depending on the species and duration of unweighting, hindlimb-unloaded rodents typically demonstrated decreases in CSA of type I soleus muscle fibers between 40%–70% 10 compared to the 21.7% reduction seen in BAMs. Some of these discrepancies may be partially explained by the lack of certain external factors that can influence the skeletal muscle atrophy process, such as endocrine effects, motoneuron innervation, and vascularization. The likely expression of immature isoforms of contractile proteins in tissue-cultured myofibers, 40 the lack of a higher level myofiber organization, and a generally shorter duration of tension reduction in our experiments are all possible contributing factors as well.
It is believed that the initial stimulus for disuse muscle atrophy is the shortening of muscle fibers and the subsequent reduction in tension. In hindlimb-suspended rats, the working length of the soleus muscle contracts an estimated 20% after 4 days. 23 Other animal models of atrophy such as limb immobilization, myomectomy of opposing muscles, and denervation, also shorten the muscle and prevent its use (i.e., reduce contraction and tension generation). With current in vivo models, it is not possible to isolate the effects that length reduction alone may have on skeletal muscle atrophy due to the compounding systemic effects of the in vivo environment. It is for this reason that the availability of a reliable and reproducible in vitro model of skeletal muscle atrophy may be beneficial. Such a model would enable the study of specific mechanisms linking length and tension reduction to muscle atrophy, including the study of different signaling pathways and gene expression patterns. With this knowledge, various pharmacologic and mechanical countermeasures can then be targeted and tested. Research arising from the use of such an in vitro model has potential clinical implications as well. It may allow for a better understanding of how tension reduction contributes to the development of skeletal muscle atrophy as seen not only in debilitated, immobilized, or paralyzed patients on earth, but also to humans exposed to extended periods of microgravity in spaceflight.
In addition to using this model for studying atrophy resulting from length or tension reduction, it can also be used to investigate the effects of other known promoters of skeletal muscle atrophy, including starvation, glucocorticoids, and certain cytokines. Although BAMs are not vascularized, a starvation-related model can potentially be used to replicate the effects of ischemia-induced skeletal muscle atrophy as well.
However, the use of an in vitro BAM model to study skeletal muscle atrophy does have its limitations. First, although there are many similar physiological characteristics, BAMs do have significant differences when compared to in vivo muscle. The relatively large amount of extracellular matrix used as scaffolding makes myofibers a significantly smaller proportion of the entire muscle unit. This may have implications on the mechanical and functional properties of BAMs. In an attempt to overcome this, attempts were made to reduce the amount of collagen and Matrigel. However, Matrigel is a solubilized basement membrane preparation that is rich in extracellular matrix proteins and appears to be an essential component of the BAM matrix. Its components, which include laminin and collagen IV, as well as growth factors such as TGF-beta and FGF, likely play an important role in enhancing the structural matrix required for BAM formation. Although BAMs made with both 1.6 and 0.8 mg/mL collagen have been found to be comparable with respect to their metabolic, morphologic, and functional properties, those made with lower collagen concentrations tended to be more fragile and more difficult to handle when processing for physiological and morphometric analyses. The stiffness of the matrix can also have a significant effect on the formation of sarcomeres in the myofibers. 41 Also of note, the BAMs were not under tension when fixed before the immunocytochemical analysis of CSA. Although there is no evidence to suggest this was the case in our experiments, it is possible that differences seen in myofiber CSA of shortened versus control BAM were confounded by differential formaldehyde-induced contraction in the two groups.
Another limitation is the fact that BAMs are not vascularized or innervated, likely contributing to some of the observed physiological differences with in vivo muscle. The lack of vascularization or an intramuscular capillary network significantly limits the potential size of the BAMs since myofibers are dependent on distance-limited diffusion for nutrients. This impacts not only the organization of the myofibers, but also the BAM's overall functional capacity and profile. With regard to innervation, two-dimensional in vitro motoneuron muscle cocultures have been readily studied and utilized,42,43 and the establishment of an in vitro innervated BAM model may be soon possible. A recent study of miniaturized PMM BAMs demonstrated effective clustering and morphological maturation of acetylcholine receptors induced by agrin and laminin, indicating the functional activity and plasticity of BAMs toward innervation. 40 The lack of innervation highlights one of the other limitations of the current BAM model, which is that BAMs express neonatal isoforms of contractile proteins. 40 This is not surprising given that myofiber or myotube innervation appears to be necessary for the development of adult isoforms of myosin heavy chain. 42
Despite current limitations of this in vitro model of skeletal muscle atrophy, the model nevertheless offers many potential advantages both to existing two-dimensional tissue culture models and to available animal and human models of atrophy. Further development, refinement, and characterization of the model, including work toward in vitro vascularization and innervation of BAMs may make this model an even more powerful tool for the study of skeletal muscle atrophy.
In summary, we have characterized tissue-engineered BAMs made from PMM and have demonstrated that length reduction alone can induce atrophy in BAMs, hence, establishing a potential in vitro model of skeletal muscle atrophy that may be used to complement other in vivo and in vitro atrophy models. The use of such in vitro models may lead to a better understanding of the mechanisms linking mechanical stimuli to a variety of signaling pathways, and ultimately, physiological changes seen in skeletal muscle after tension reduction. These data may ultimately aid in the development of new pharmaceuticals to prevent, attenuate, or treat muscle wasting resulting from a variety of disorders or conditions.
Footnotes
Acknowledgments
The authors thank Paul Kosnik, PhD, Bernard Creswick, and Janet Shansky for their technical assistance. We also thank Tina Hernadez-Boussard, PhD, MPH, for her statistical expertise and guidance. The work was supported by NASA NCC2-1062.
Disclosure Statement
No competing financial interests exist for either author.