
Abstract
Interaction between chondrocytes and the cartilage extracellular matrix (ECM) is essential for maintaining the cartilage's role as a low-friction and load-bearing tissue. In this study, we examined the influence of cartilage zone-specific ECM on human articular chondrocytes (HAC) in two-dimensional and three-dimensional (3D) environments. Two culture systems were used. SYSTEM 1: HAC were cultured on cell-culture plates that had been precoated with the following ECM molecules for 7 days: decorin, biglycan, tenascin C (superficial zone), collagen type II, hyaluronan (HA) (middle and deep zones), and osteopontin (deep zone). Uncoated standard culture plates were used as controls. Expanded cells were examined for phenotypic changes using real-time polymerase chain reaction. In addition, expanded cells were placed into high-density pellet cultures for 14 days. Neocartilage formation was assessed via gene expression and histology evaluations. SYSTEM 2: HAC that were cultured on untreated plates and encapsulated in a 3D alginate scaffold were mixed with one of the zone-specific ECM molecules. Cell viability, gene expression, and histology assessments were conducted on 14-day-old tissues. In HAC monolayer culture, exposure to decorin, HA, and osteopontin increased COL2A1 and aggrecan messenger RNA (mRNA) levels compared with controls. Biglycan up-regulated aggrecan without a significant impact on COL2A1 expression; Tenascin C reduced COL2A1 expression. Neocartilage formed after preculture on tenascin C and collagen type II expressed higher COL2A1 mRNA compared with control pellets. Preculture of HAC on HA decreased both COL2A1 and aggrecan expression levels compared with controls, which was consistent with histology. Reduced proteoglycan 4 (PRG4) mRNA levels were observed in HAC pellets that had been precultured with biglycan and collagen type II. Exposing HAC to HA directly in 3D-alginate culture most effectively induced neocartilage formation, showing increased COL2A1 and aggrecan, and reduced COL1A1 compared with controls. Decorin treatments increased HAC COL2A1 mRNA levels. These data indicate that an appropriate exposure to cartilage-specific ECM proteins could be used to enhance cartilage formation and to even induce the formation of zone-specific phenotypes to improve cartilage regeneration.
Introduction
C
Cells within the superficial zone (SZ) synthesize and secrete a specialized protein called the superficial zone protein (SZP),3–5 which is also known as megakaryocyte-stimulating factor, lubricin, and proteoglycan 4 (PRG4). 6 PRG4 plays a critical role in creating the low-friction properties at the cartilage surface, and, correspondingly, its expression is largely confined to chondrocytes that are located in the first 500 μm from the surface of articular cartilage with very few positive cells in the middle zone (MZ) and the deep zone (DZ). 7 Clusterin, a glycoprotein that is found exclusively expressed in SZ chondrocytes, regulates complement activation and cell death. 8 SZ chondrocytes also produce less collagen type II8–10 and keratan sulfate and other proteoglycans compared with DZ chondrocytes.2,11–14 However, the SZ appears to harbor higher levels of small leucine-rich proteoglycans (SLRPs).15,16 The major SLRPs present in cartilage include decorin, biglycan, asporin, lumican, and fibromodulin that play a role in tissue integrity, modulate metabolism, and mediate growth factor signaling.17–19 The glycoprotein, Tenascin-C (TN-C), is a modular multifunctional ECM molecule that is involved in cell adhesion, migration, and proliferation 20 and is found to be predominately expressed in the “transitional” region spanning the lower SZ and upper MZ in normal cartilage. 21
The MZ starts∼100–200 μm from the surface and is characterized by a high aggrecan content 22 and specifically high in hyaluronan (HA), dermatan sulfate, and collagen type II.23–28 Aggrecan's primary role is to maintain tissue hydration, which is an important element to endow cartilage with its mechanical properties. 29 HA functions as a scaffold for building proteoglycans and modulates cell adhesion and motility. 30 Collagen type II forms a scaffold or framework that provides tensile strength to the tissue. 31 Cartilage intermediate layer protein (CILP) is uniquely located in the MZ 32 and may mediate interactions between ECM components. 33
The radial or DZ comprises the lowest cell density and collagen content 34 ; however, collagen fibrils are thicker in this region and the proteoglycan content, including aggrecan, keratan sulfate, and chondroitin sulfate, is much higher than the upper zones.23,35–37 Osteopontin is expressed in the DZ38,39 and interacts with integrin receptors to promote cell adhesion, chemotaxis, and signal transduction. 40
It is plausible that the ECM environment may be an important factor in controlling and maintaining zone-specific cellular phenotypes. Engineering cartilage with zone-specific cellular and ECM phenotypes will be required to obtain biomechanical function and long-term survival similar to native tissue. Potentially, ECM components provide a tool to direct the differentiation of cells to the desired cartilage and, more specifically, a zonal phenotype. The behavior of various cell populations has been investigated after an interaction with bioactive ECM molecules, such as collagens type I, II, fibronectin, laminin, and HA.41–45 These proteins facilitate cell attachment, proliferation, differentiation, and the differentiated functions of cells.45,46 In particular, expansion on collagen type II and perlecan domain 1 can enhance chondrogenic differentiation.47–50
However, the effect of cartilage zone-specific molecules on chondrocyte differentiation, with regard to both general and zone-specific cartilage markers, is largely unknown. In addition, responses to ECM are likely to be different in monolayer (ML), or in three-dimensional (3D) pellet or hydrogel culture. Finally, pre- or coculturing chondrocytes in the presence of zone-specific ECM may preferentially direct cells toward zone-specific phenotypes.
In this study, we examined (i) the response of human articular chondrocytes (HAC) expanded on a number of cartilage-specific ECM molecules in ML culture; (ii) the effect that preculturing cells on each ECM component had on neocartilage formation in pellet cultures; and (iii) neocartilage formation in the direct presence of ECM proteins in 3D alginate hydrogel cultures.
Methods
Tissues, cell isolation, and culture conditions
Human knee articular cartilage was obtained from tissue banks (approved by Scripps Institutional Review Board). Overall, 11 donors were used in this study (1 woman, 10 men; age=31±12 years). Chondrocytes were isolated from full-thickness cartilage shavings via enzymatic digestion and were cultured in ML for one passage on standard tissue culture flasks in 1×Dulbecco's modified Eagle's medium with 4.5 g/L glucose,
ECM substrates and ML cell culture conditions
ECM molecules were selected based on cartilage zone-specific expression patterns. Decorin, biglycan, and TN-C are predominately located in the SZ,16,21 and collagen type II and hyaluronan (HA) are present at higher proportions in the MZ and DZ.23–28 Osteopontin is almost exclusively located in the DZ.38,39 Precoating of culture plates (six-well) with either human decorin (R&D Systems, Minneapolis, MN), biglycan (R&D Systems), TN-C (R&D Systems), collagen type II (Chondrex, Redmond, WA) HA (Ferring Pharmaceuticals, Inc., Parsippany, NJ), or osteopontin (R&D Systems) was performed by suspending 1 μg/mL of each component in phosphate-buffered saline (PBS) and incubating it at 4°C overnight. The suspension was removed, and the plates were washed twice with 1×PBS. Plates not preincubated with any ECM solution were used as controls. Primary human chondrocytes, which were expanded on standard culture plates for one passage, were then seeded on the various precoated plates and maintained on the precoated-culture plates for 7 days in ML culture. No subsequent addition of ECM molecules was introduced during this 7 day culture phase. The cells were detached using accutase (Innovative Cell Technologies, Inc., San Diego, CA) and collected for either real-time polymerase chain reaction (RT-PCR) or high-density pellet cultures.
Pellet culture
The high-density pellet cultures were only used for chondrocytes that were cultured on each ECM molecule for 7 days. After culture on ECM molecules, the cells were detached, counted, and suspended at a density of 0.5×106 cells in insulin, transferrin, selenium (ITS) media (Sigma Chemical, St. Louis, MO) supplemented with 10 ng/mL transforming growth factor β1 (TGFβ1; PeproTech, Rocky Hill, NJ). The cells were centrifuged at 800 rpm for 5 min to form high-density-cell pellets, which were cultured in an incubator (37°C, 5% CO2) with loosened tube caps for 14 days as previously described. 47 Medium was changed every 3–4 days.
Chondrocyte embedding with alginate/ECM molecule hydrogels
For exposure of chondrocytes to ECM molecules in 3D alginate cultures, the chondrocytes were culture expanded to passage 1 in nontreated culture plates. A 2% alginate solution (PRONOVA UP LVG; NovaMatrix, Sandvika, Norway) in Hank's balanced salt solution was prepared as previously detailed. 51 The alginate was mixed with 1 μg/mL of either human decorin, biglycan, TN-C, collagen type II, or osteopontin. HA was added at a concentration of 1 mg/mL. The appropriate alginate/ECM combination was then mixed with chondrocytes at a density of 8×106/mL. The cell/alginate and ECM mixture were added as droplets (∼50 μL each using a 200 μL pipette) into alginate gelation buffer (120 mM CaCl2; 25 mM HEPES; and 150 mM NaCl) and incubated at room temperature for 20 min. The buffer was removed, and the alginate beads were washed thrice with 1×PBS. Afterward, at least five beads were transferred into each well of a six-well plate, cultured in ITS+ medium supplemented with 10 ng/mL of TGFβ1, and cultured for 14 days. 47 The medium was changed twice per week. Chondrocytes were released from the alginate beads for RNA extraction by the addition of 50 mM ethylenediaminetetraaceticacid (EDTA) in PBS. The released cells were centrifuged and washed in PBS before RNA was extracted.
RNA isolation and cDNA synthesis
Total RNA was isolated from chondrocytes by following the instructions provided in the RNeasy Mini Kit (Qiagen, Hilden, Germany). Total RNA was eluted into 30 μL of RNase-free water and was quantified using a NanoDrop ND-1000 V3.5.2 Spectrophotometer. Isolated messenger RNA (mRNA) was reverse transcribed using High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA).
Quantitative RT-PCR
Quantitative RT-PCR was performed using prevalidated TaqMan® gene expression reagents (Applied Biosystems) for the detection of COL1A1, COL2A1, COL10A1, PRG4, aggrecan, clusterin (CLU), vascular cell adhesion molecule 1 (VCAM-1), and CILP. Expression levels were normalized to GAPDH using the ΔCt method as previously reported. 52
Cell viability
Chondrocyte viability in alginate cultures was evaluated using the live/dead kit (Applied Biosystems) composed of Ethidium Homodimer-1 (detect dead cells; red) and Calcein-AM (live cells; green) as previously detailed. 53 After incubation with these dyes, the cell beads were viewed with a confocal fluorescence microscope (LSM-510; Zeiss, Jena, Germany) to assess the viability using a script written in MATLAB Software (MathWorks, Natick, MA), configured to enumerate green (live) and red (dead), cell signals were used. Viability is reported as the percentage of live cells.
Histology
Cell pellets and alginate beads were fixed in Z-fix (Anatech Ltd., Battle Creek, MI) for 1 h, washed with 1×PBS, and dehydrated in 70% ethanol before further dehydration and then paraffin embedded. Paraffin blocks were cut into 4 μm-thick sections, placed onto charged microscope slides, and kept at room temperature until staining with Safranin-O fast green.
Immunohistochemistry
Paraffin-embedded cell beads were sectioned at 4–6 μm and were placed on glass slides. The sections were then deparaffinized by immersion in pro-par clearant (Anatech Ltd.) thrice for 5 min, dehydrated in 70–100% ethanol for 3 min each, and then washed in distilled water. Sections for collagen type II staining were subjected to trypsin (Sigma-Aldrich, St. Louis, MO) digestion for 30 min at room temperature. Hyaluronidase (Sigma-Aldrich; for 30 min at room temperature) was used for aggrecan immunodetection.
After pretreatment, the slides were blocked with goat serum (2%) on the sample for 30 min. The appropriate primary antibody (each 10 μg/mL) for collagen type II (II–II6B3) or aggrecan (12/21/1-C-6) was incubated overnight at 4°C. All antibodies were obtained from the Hybridoma Bank (University of Iowa, Iowa City, IA). Mouse immunoglobulin G (IgG) was used as an isotype control to monitor nonspecific background staining. Human cartilage tissue was used as a positive control. After washing in PBS and blocking, the sections were incubated with the secondary biotinylated Goat anti-Mouse antibody for 30 min. Visualization of the signal was achieved using alkaline phosphatase (VECTOR Red Alkaline Phosphatase Substrate Kit; Vector Laboratories, Burlingame, CA). Sections were then dehydrated in ethanol, pro-par and were preserved in Refrax mounting media (Anatech Ltd.) under a glass cover slip.
In situ hybridization
PRG4 mRNA localization in pellets was assessed using an antisense probe under conditions previously described. 54 Multiple micrographs were taken of 3–5 sections per condition, and signal count intensity was assessed using a script written in MATLAB Software (MathWorks) that was configured to detect a probe signal above background signal. Average signal±standard error of the mean was calculated for each treatment.
Statistical analyses
Gene expression level, calculated percentage viability, and PRG4 in situ signal comparisons between control and treatments were conducted using Student t-tests using Excel (Microsoft Office Excel 2007). p-values less than 0.05 were considered significant.
Results
Cartilage zone-specific ECM differentially alters gene expression in chondrocyte ML culture
Culture on decorin, HA, and osteopontin for 7 days significantly (p<0.05) increased COL2A1 and aggrecan mRNA levels compared with controls (Fig. 1). Biglycan (SZ) significantly (p<0.05) up-regulated aggrecan without a significant impact on COL2A1 expression. TN-C (superficial/transition zone) significantly (p<0.05) reduced COL2A1 expression and generally down-regulated all genes analyzed (Fig. 1). Osteopontin, a deep-zone protein, was the only ECM to increase COL10A1 expression levels, Fig. 1E), although not significantly.
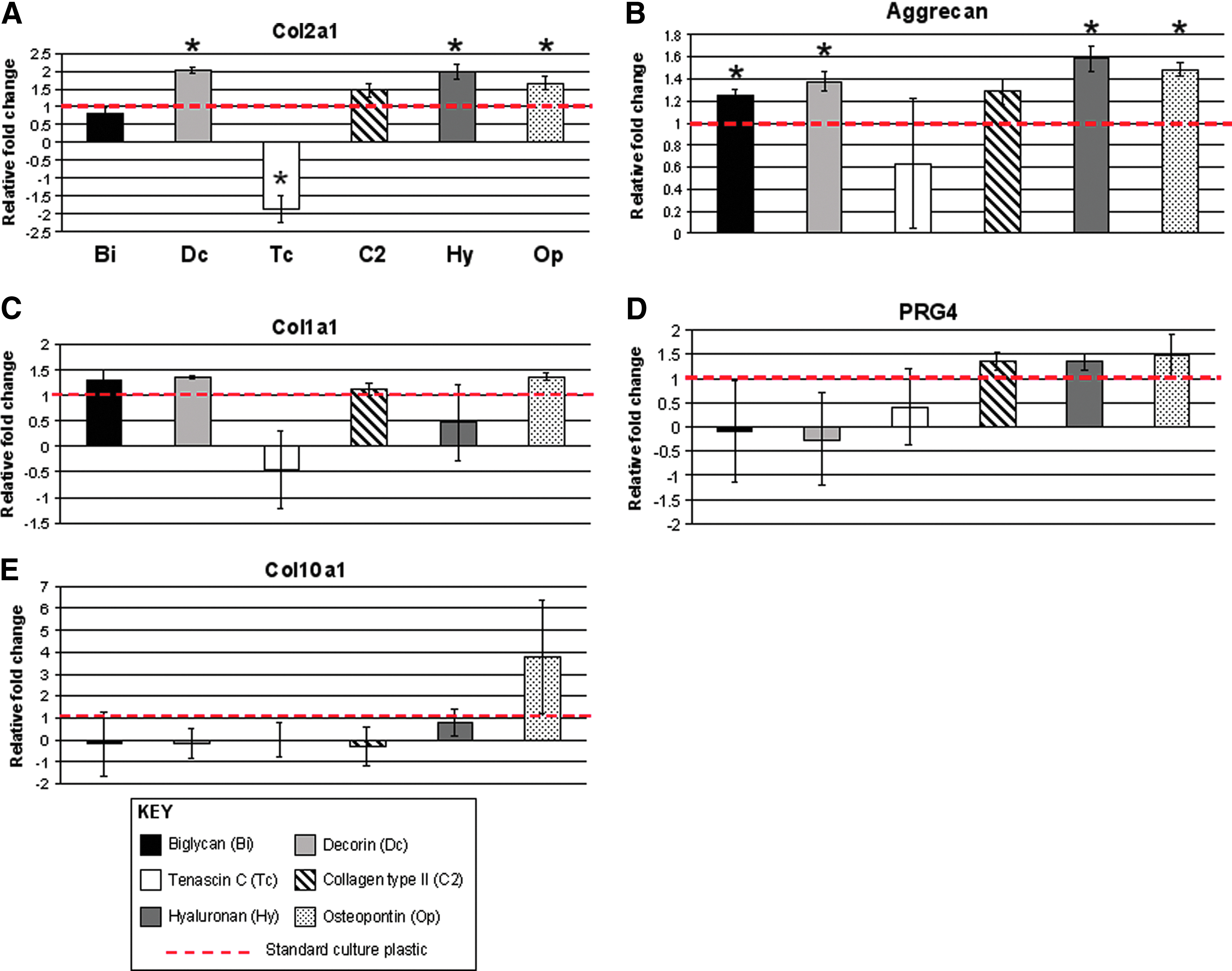
Gene expression profiles of human articular chondrocytes expanded in monolayer cultures on various extracellular matrix (ECM) molecules (7 days).
Chondrogenic cartilage matrix formation in chondrocyte pellet culture is altered by preculture conditions
Chondrocytes cultured on the SZ-specific proteins TN-C and collagen type II and subsequently cultured in high-density pellets for 14 days led to neo-tissues that expressed higher COL2A1 mRNA (1.6–2.5-fold) compared with control pellets (Fig. 2A). Decorin increased COL2A1 threefold but was not significantly different from controls (p<0.07). On the other hand, these ECM proteins did not change aggrecan expression. Preculture on HA decreased both COL2A1 (p<0.07) and aggrecan (p<0.05) expression levels compared with controls (Fig. 2A, B), which is consistent with histology (Fig. 2D). Cells cultured on the collagen type II substrate generated pellets with the most uniform Safranin-O stain and collagen type II ECM deposition (Fig. 2D). Culturing human chondrocytes with ECM proteins did not increase PRG4 expression (Fig. 2C); rather, it reduced its expression. A twofold reduction (p<0.05) in PRG4 mRNA was observed with biglycan and collagen type II (Fig. 2C). Osteopontin culture-treated cells produced a greater than threefold reduction in PRG4 at a significance level of p<0.06. HA trended to decrease PRG4 expression in pellets; however, due to variations in donor response, this was not significant compared with controls. Detection of PRG4 via in situ hybridization also revealed a decrease in PRG4 signal in pellets from chondrocytes precultured on HA and osteopontin (p<0.05); however, decorin significantly reduced PRG4 in situ signals (p<0.05) while biglycan reduced its signal at a nonsignificant level (Fig. 3). The hypertrophic marker COL10A1 was not detected via RT-PCR in any condition (data not shown).
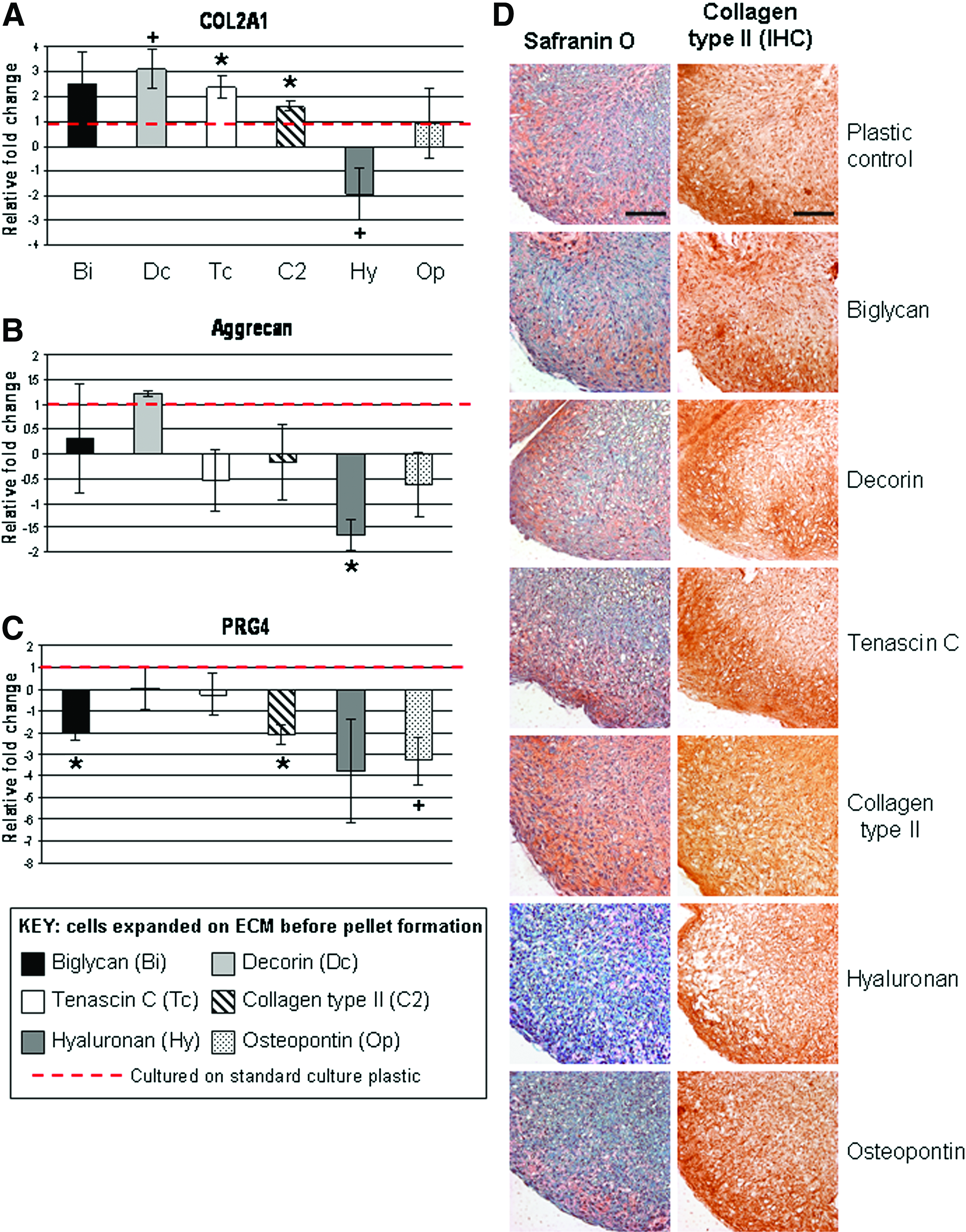
Neocartilage formation by pellet culture after expansion on extracellular matrix (ECM) molecules in monolayer culture. Gene expression profiles of human articular chondrocytes cultured in high-density pellets for

In situ hybridization for PRG4 messenger RNA.
Presentation of ECM molecules in alginate 3D culture alters neocartilage formation
The viability of chondrocytes cultured in alginate alone or with each ECM component did not significantly differ (Supplementary Fig. S1A, B; Supplementary Data are available online at www.liebertpub.com/tea). Exposing cells to HA directly in 3D-alginate culture (no HA culture expansion) had the most potent effect on neocartilage formation by enhancing COL2A1, Aggrecan gene (p<0.05), and protein expression (Figs. 4A–C and 5). Decorin treatments increased (p<0.06) COL2A1 mRNA levels compared with the control (Fig. 4). Chondrocytes cultured in alginate mixed with biglycan, collagen type II, TN-C, and osteopontin did not significantly change COL2A1, aggrecan, and COL1A1 mRNA levels (Fig. 4A–C), although collagen type II trended to enhance COL2A1 expression with a larger donor-to-donor variation (Fig. 4A). No significant change in COL10A1 mRNA levels was seen in all chondrocyte samples and based on cycle numbers, COL10A1 mRNA levels in all chondrocyte samples were very low.
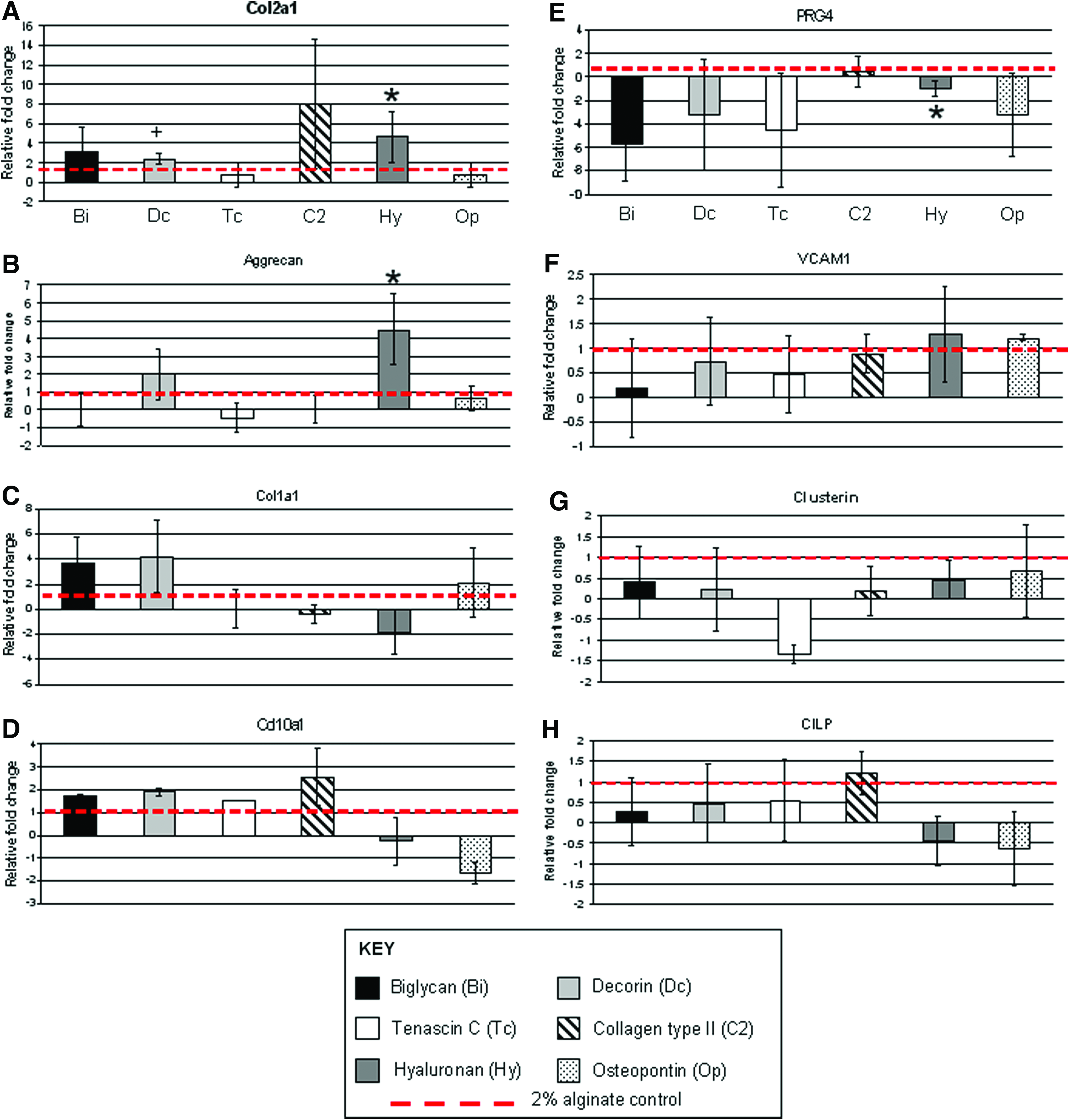
Gene expression profiles of human articular chondrocytes in 2% alginate in the presence of different ECM proteins.
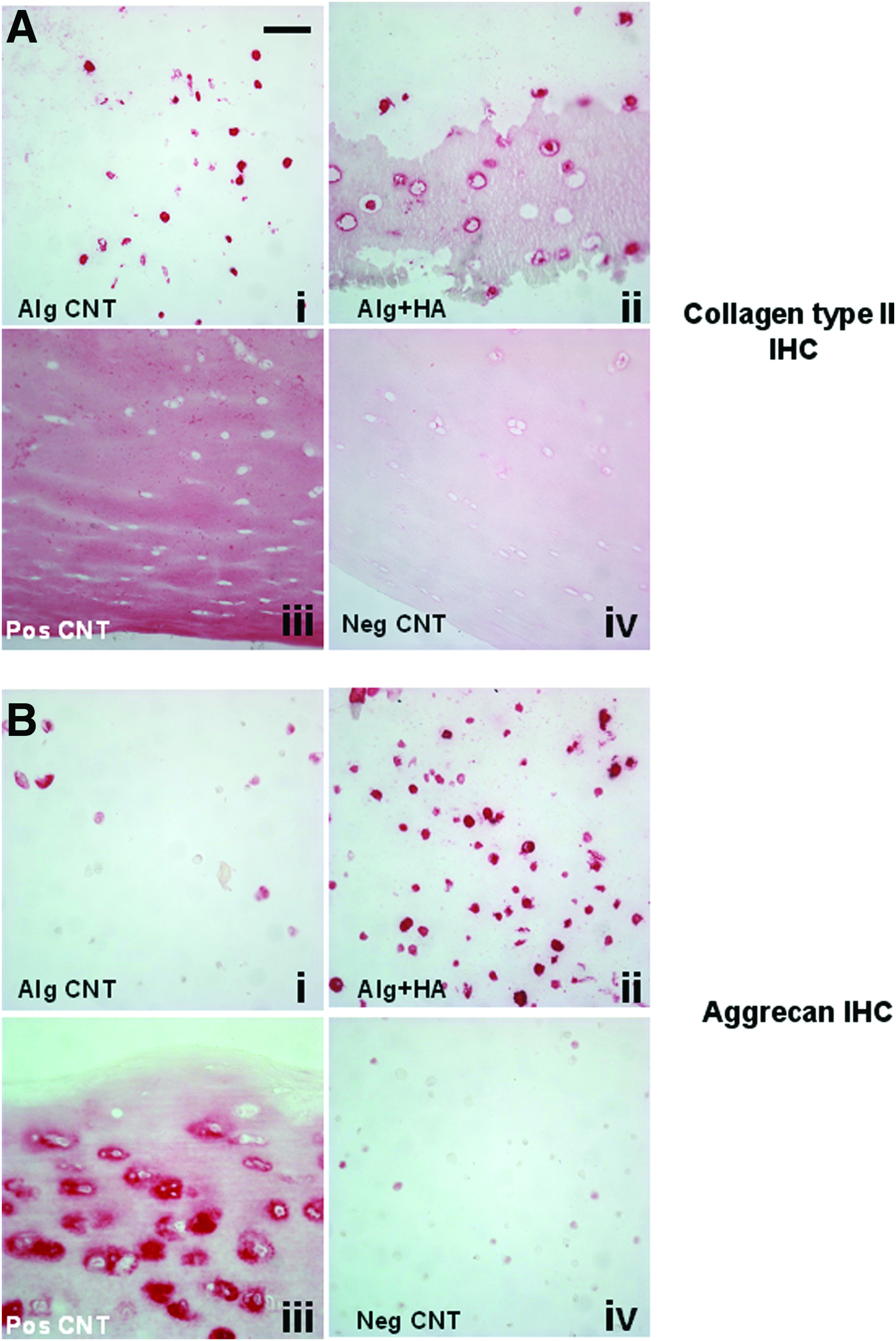
Immunohistochemistry (IHC) for
The chondrocyte response to cartilage zone-specific ECM molecule exposure resulted in a few changes in mRNA levels of genes deemed to be zone specific. PRG4 mRNA levels trended to be reduced with biglycan, decorin, TN-C, and osteopontin treatment (Fig. 4E); however, high donor variation indicates a nonsignificant response. VCAM-1 and CLU are almost exclusively expressed in the cartilage SZ (VCAM-11, CLU 8 ); however, ECM molecule treatments did not significantly alter the expression of these SZ expression markers (Fig. 4F, G). No treatment effect was seen (compared with controls) with the expression of CILP, a cartilage MZ marker (Fig. 4H).
Discussion
To engineer cartilaginous tissue with structure, organization, and function close to that of native adult articular cartilage, knowledge of the composition and intimate interaction between cells and their ECM environment is essential. In this study, we have demonstrated that cartilage-specific ECM molecules alter cell phenotype in ML culture and alter cell responses to 3D culture systems in a manner that may be used to better guide cartilage tissue formation and to better emulate the natural state.
The paradigm of isolating and culturing cells to increase cell numbers is still required for many current tissue engineering approaches. Including a step of presenting appropriate ECM molecules in the ML stage, particularly decorin, TN-C, and collagen type II, may promote better neocartilage formation. Exposure to some ECM molecules in ML leads to direct cartilage zone-specific expression profiles.
Decorin, HA, and osteopontin increased COL2A1 and aggrecan expression after 7 days in ML compared with standard plastic culture; however, this shift did not correspond to the eventual phenotype observed in the pellets. Only cells that expanded on decorin resulted in a shift in COL2A1 RNA expression relative to control pellets. Conversely, TN-C reduced COL2A1 in ML; however, it significantly enhanced its expression in pellets. While collagen type II did not alter COL2A1 expression in ML, it significantly enhanced COL2A1 expression in the pellets as previously demonstrated.55,56 HA had the opposite effect in ML; it significantly increased COL2A1 and aggrecan in ML and markedly reduced neocartilage formation in the high-density pellets. The contrasting responses that were observed in these two systems are not easily resolved with the data available in the present study. However, some mechanisms may be postulated for all molecules.
First, collagen type II protein has been demonstrated to enhance chondrogenesis of adipose tissue-derived mesenchymal stem cell (MSC) through the integrin-mediated activation of RhoA/Rock pathway. 57 TN-C is known to be involved in promoting early chondrogenesis,58,59 possibly by suppressing RhoA, focal adhesion, and actin stress fibers,60,61 which is critical for chondrogenesis. 62 Salinas and Anseth 63 demonstrated that the KLER sequence, a binding site from decorin protein, was incorporated into polyethylene glycol (PEG) gels to enhance MSC chondrogenesis. Decorin is known to bind strongly to collagen type II and is involved in matrix organization. 63
The contrasting action of HA is more difficult to resolve. While enhancing chondrogenic gene expression in ML, it significantly reduced neocartilage formation in high-density pellet culture after exposure to HA in ML. Studies including HA indicate a positive influence to enhance chondrogenesis,64,65 which is what we also found in the alginate 3D system in this present study. To our knowledge, no other peer-reviewed published study has examined the cartilage-forming response of chondrocytes after expansion on HA in ML culture; however, a similar observation was made by Ryan et al., 66 who showed reduced chondrogenic potential of MSC cultured on an HA-prepared surface. The basis for the negative effect of expanding cells on HA and the subsequent poor neocartilage formation is not clear; however, it may be related to a number of factors. One factor could be due to the interaction of HA with its receptor, CD44 while in ML culture. Sugahara et al. 67 showed that HA can induce cleavage of CD44 from the surface of tumor cells. CD44 and HA interaction is known to be important in early chondrogenesis 68 and in 3D systems to drive chondrogenesis of progenitor cells. 69 HA binding of CD44 in bovine chondrocytes enhanced neocartilage formation through interactions between CD44 and TGFβ1 receptor signaling. 65 Since chondrocytes express high levels of CD44 in ML culture 70 and reduced CD44 surface marker levels are associated with reduced chondrogenic capacity, 71 it is plausible that HA exposure in ML may reduce CD44 surface levels and hinder subsequent CD44 and HA interactions needed for robust neocartilage formation when placed into the high-density pellet system.
Encapsulating chondrocytes in alginate leads to expression of COL2A1 and Aggrecan without the presence of ECM molecules. However, similar to the pellet culture results, exposure to biglycan, decorin, and collagen type II tended to further enhance COL2A1 mRNA levels, although with a high variation in donor response. While TN-C enhanced COL2A1 mRNA levels in the pellet cultures, no change was seen in the 3D alginate environment. Osteopontin did not affect chondrogenesis in any of the systems tested. As discussed earlier, HA supported the most robust profile of neocartilage formation with decreased mRNA levels of COL1A1 and increased expression levels of COL2A1 and Aggrecan; in contrast to the pellet cultures of cells expanded on HA. The enhanced neotissue formation in the presence of HA in alginate is likely due to a combination of the 3D-alginate environment per se, the high CD44 levels, usually found in dedifferentiated chondrocytes that are able to bind to the available HA in the gel to thus rapidly initiate the associated prochondrogenic signaling pathways.
It was postulated that ECM proteins may be involved in modulating the cartilage-zonal phenotype. As a read-out, we primarily focused on the expression of PRG4 as a representative SZ marker, although changes in VCAM-1 and clusterin mRNA levels were also examined. These latter two markers were not significantly altered by any ECM molecule treatment; however, an overall tendency of reduced PRG4 mRNA levels was observed for most ECM molecule treatments (relative to controls). Biglycan and decorin are reported to be localized at higher concentrations in the SZ, 16 and TN-C is localized in the transitional zone between the SZ and upper MZ. 21 Both biglycan and decorin repressed PRG4 expression compared with controls, yet TN-C did not significantly alter its expression. It is possible that differences in the local concentration of these specialized ECM proteins may modulate the production of SZP levels by certain cell populations in the SZ. In addition, the presence of osteopontin and HA, found in higher concentrations in the middle and DZs, also appear to exert a significant inhibiting effect on PRG4 levels to mimic its expression profile seen in native cartilage tissue.
In this present study, we did not test the effect of various combinations of ECM molecules, nor did we examine the effect of using different concentrations; however, such an approach may be useful for fine-tuning zonal phenotype. To promote the SZ phenotype, presentation of the appropriate concentrations of TN-C, biglycan, and decorin will likely permit the expression of PRG4 and COL1A1, while reducing COL2A1. Both biglycan and decorin trended to increase COL1A1 mRNA levels, which is one feature of the SZ. 72 Using HA alone or in combination with biglycan and collagen type II, 3D cultures may be sufficient for promotion of the MZ and DZ cartilage phenotypes. Various combinations of these ECM proteins may also be more important, especially in mediating growth factor presentation or utilizing enhanced ECM organization.
Since no single ECM protein was fully successful in inducing a zone-specific phenotype, perhaps the use of naturally derived ECM from cartilage itself or from isolated cartilage zones may be more desirable than the purified ECM molecules used in this study. Examples of this approach have been reported for heart and meniscus regeneration strategies.73,74 We examined a mixed population of cells; however, studies focused on the responses of zone-specific cells and the corresponding zone-specific ECM would be interesting and may provide greater insights in re-establishing zone-specific phenotypes.75,76
With regard to the effects observed as a consequence of ML exposure to the various molecules, the process of coating the culture plastic may have altered the biological activity of the molecules used in this study. While we detected HA and collagen type II on the plates via Safranin O staining and immunostains, respectively (data not shown), we cannot be certain that all molecules tested in this study equally adhered to the culture surface. The dynamics of mixing the various ECM molecules within the alginate hydrogels is not clearly defined in terms of whether the molecules remained within the gels, whether the concentration was uniform throughout, and whether the cells were able to interact with the molecules while encapsulated within the cross-linked hydrogel. Thus, variations in response by cells may be due to these factors.
Overall, we have shown that exposing chondrocytes to ECM molecules in different contexts can alter the subsequent neocartilage formation and can even modulate cartilage zone-specific gene expression profiles. While no single ECM protein may be ideal to promote zone-specific phenotypes, specific combinations and concentrations may be needed. Appropriate exposure of some ECM proteins during chondrocyte or progenitor cell culture may be a practical way to improve neotissue formation and to improve cartilage repair.
Footnotes
Acknowledgments
Funding was provided by NIH/NIAMS P01 AG007996, NIH UL1 RR025774, and CIRM TR1-01216. Technical assistance by Nick Steklov, Peter Chen, Shantanu Patil, Stuart Duffy, and Lilo Creighton is appreciated. The authors thank Judy Blake for manuscript formatting and copyediting. Antibodies used in this study were obtained from the Developmental Studies Hybridoma Bank developed under the auspices of the NICHD and maintained by The University of Iowa, Department of Biology, Iowa City, IA 52242.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.