
Abstract
Background:
Current tissue-engineered cartilage constructs contain insufficient amounts of collagen, whose function is to resist tension. We postulate that dynamic tension is necessary to stimulate collagen formation. Another shortcoming is that tissue-engineered cartilage does not possess native zonal variations. We hypothesize that applying depth-varying mechanical cues would stimulate extracellular matrix (ECM) synthesis depth dependently. We developed a dedicated loading regime called sliding indentation, which enables us to apply dynamic tension as well as depth-varying strain fields to the chondrocyte-seeded agarose constructs.
Objective:
In 2 study designs, we explored whether sliding indentation would increase collagen content and induce depth-varying ECM distribution.
Methods:
In the first study, we developed an agarose-sandwich model that involves embedding of a thin chondrocyte-seeded 0.5% agarose layer between two cell-free 3% agarose layers. In the second study, 3-mm-thick chondrocyte-seeded agarose constructs were created. Sliding indentation at 10% depth and 1 Hz was applied to constructs in both studies for 4 h/day during 28 days, and unloaded constructs served as control.
Results:
Sliding indentation resulted in an increased amount of collagen in the produced cartilage layer. Further, sliding indentation for 7 days resulted in a depth-dependent response at gene expression levels, with the highest response in the regions that received highest strains. Analysis of protein expression after 28 days showed a similar depth-dependent distribution in all constructs, which further enhanced by sliding indentation.
Conclusions:
Sliding indentation can increase collagen content and enhances depth-dependent ECM distribution, and is therefore a promising strategy for culturing cartilage with improved properties.
Introduction
In healthy cartilage, the function of the collagen fiber network is to resist tension. We therefore hypothesize that to stimulate collagen formation, we need to apply tension to the tissue-engineered cartilage constructs. Recently, we have shown that collagen synthesis in periosteum-derived cartilage can be stimulated by application of dynamic tension induced by a novel sliding indentation loading regime, which involves an indenter compressed into the construct and slid over it to ensure dynamic straining.9,10 Based on these results, we postulate that this stimulatory effect of sliding indentation on collagen synthesis is generally applicable to the cartilage tissue engineering systems.
Therefore, the first aim of this study was to explore whether application of sliding indentation will enhance collagen synthesis by chondrocytes seeded in agarose. To enable this, the organ model system previously described 10 was adjusted such that we could apply sliding indentation to chondrocytes seeded in low-concentration agarose layers embedded between stiffer (3%) cell-free agarose layers. Low-concentration agarose (0.5%) was used, because we recently showed that reducing the agarose concentration results in a more uniform ECM distribution and an improved compressive equilibrium modulus (unpublished data).
Considering the importance of depth-dependent variations in normal articular cartilage, studies have aimed at engineering cartilage with zonal structure, function, or both. 11 Approaches to mimic the zonal structure and function include cell-based methods,11,12 scaffold-based methods,13,14 and a combination thereof.15,16 Here, we propose an alternative approach to induce depth-varying inhomogeneity within chondrocyte-seeded agarose constructs, based on mechanical loading. Since mechanical loading is a well-established trigger for matrix synthesis by chondrocytes in tissue-engineered cartilage, we hypothesized that by applying depth-dependent mechanical cues to the chondrocytes, these would be stimulated to produce a depth-varying tissue content. For this purpose, also the sliding indentation loading regime was used. Numerical evaluation of this back-and-forth sliding indentation has shown that it induced depth-varying strain fields in chondrocyte-seeded agarose constructs, with high-magnitude strains in the top half of the construct (Fig. 1; red) and lower strains in the bottom part of the construct (Fig. 1; green). 9
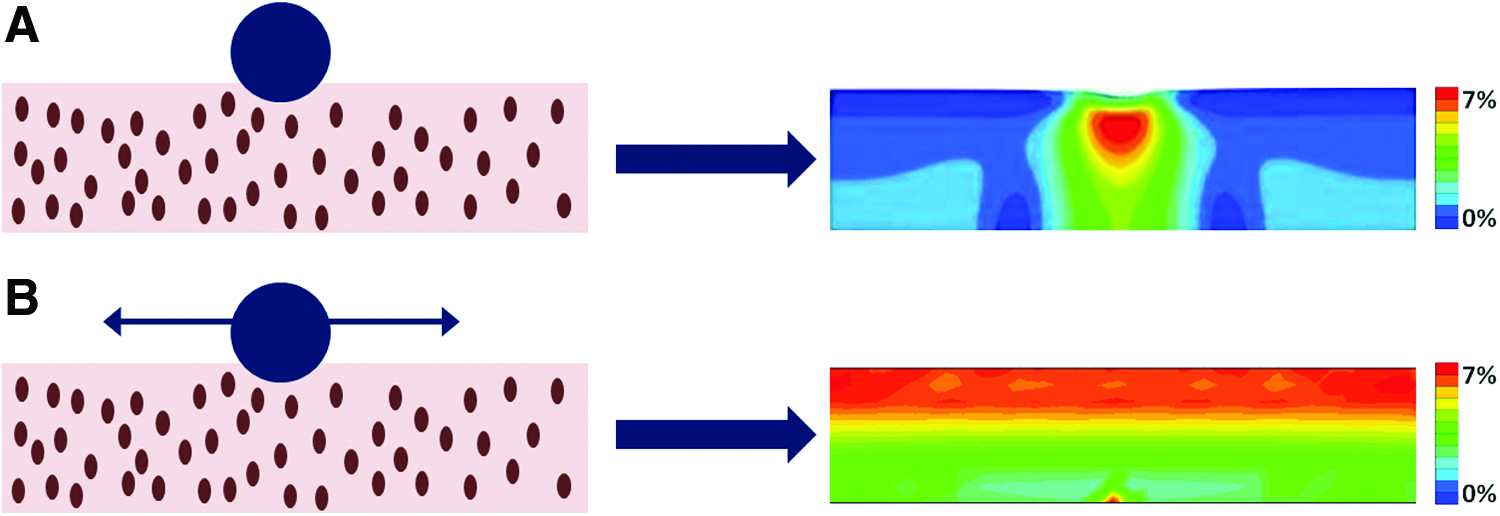
Numerical evaluation of local indentation
Based on the knowledge that sliding indentation induces a depth-dependent mechanical stimulus and that mechanical stimulation can enhance matrix synthesis, the second aim of the study was to evaluate whether sliding indentation to chondrocyte-seeded agarose constructs would result in a depth-dependent ECM deposition that corresponded with the applied strain field.
Methods
Sliding indentation bioreactor system
A bioreactor was designed (LifeTec Group) to apply sliding indentation to chondrocyte-seeded agarose constructs (Fig. 2A). The bioreactor consists of a bottom plate with six separate containers, each containing a trough (length: 11.6 mm; width: 8 mm; height: 3 mm) for the constructs (Fig. 2C). The bioreactor system consists of an indenter unit, which positions three cylindrical glass indenters (Ø 4.5 mm) on top of three of the six constructs (Fig. 2B). The bioreactor is designed to move the indenters back and forth in an exact horizontal plane of motion of the constructs and the indentation depth of each indenter, as well as the medium conditions, can be adjusted separately. The indentation depth is chosen based on the outcome of numerical simulations in which the indentation depths of 5% and 10% were compared. 9 Both indentation depths resulted in a depth-dependent strain field, but an indentation depth of 10% resulted into more physiological tensile strains of 7% directly under the indenter, whereas strains were below 5% when an indentation-depth of 5% was used. A loading frequency of 1 Hz was chosen, since this has shown to be the most effective loading frequency in other loading regimes.17,18 During culture, the bioreactor is positioned in an incubator at 37°C, 5% CO2, and 21% O2. Gas exchange is ensured with an inlet on top of the system.
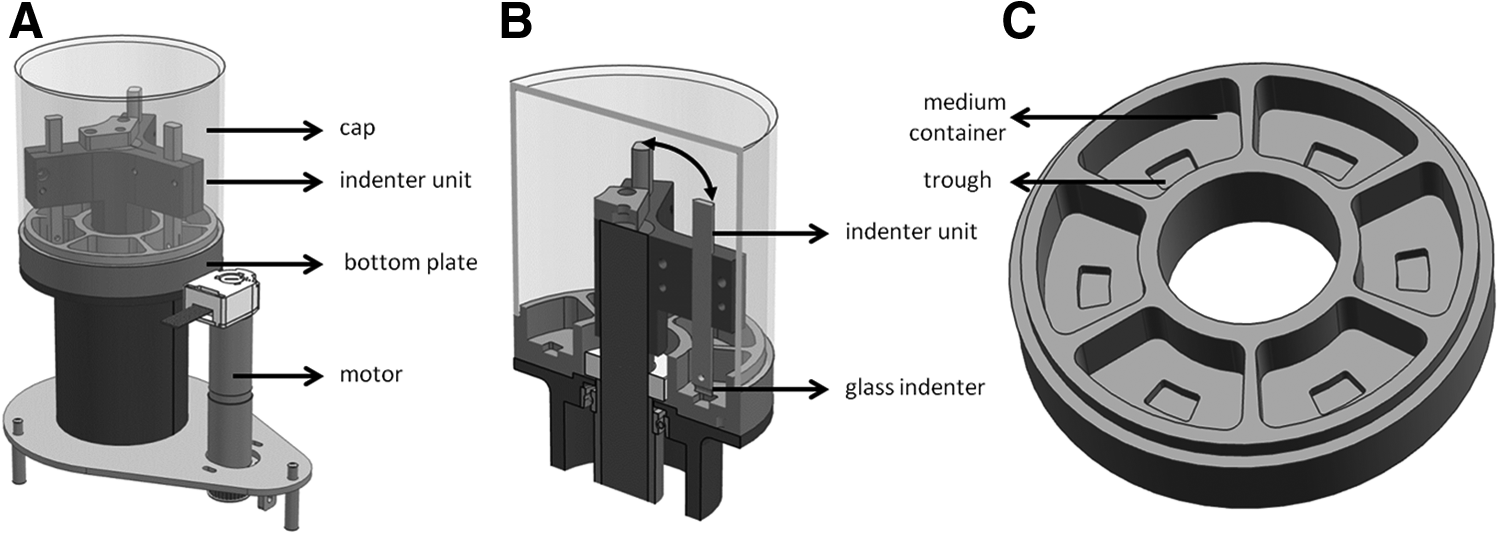
Overview of a complete sliding indentation bioreactor system, including motor
Cartilage and chondrocyte isolation
Full-depth slices of articular cartilage were removed from eight (n=3 for study 1; n=6 for study 2) carpometacarpal joints of the forelimbs of five young (8–12-month old) calves. The cartilage slices were minced and washed twice with phosphate-buffered saline (PBS) supplemented with 1% Pen-Strep (Lonza). Afterward, the slices were combined and digested with 0.5% collagenase type II (Worthington, Antonides–Interchema) at 37°C with stirring. After 16 h, the released chondrocytes were passed through a 70-μm pore-size cell strainer (Becton Dickinson), washed in DMEM (Gibco), and resuspended in DMEM supplemented 1% Pen-Strep and 0.1% amphotericin (Sigma).
Culture protocol study 1
Using 2 bottom plates, 12 agarose-sandwich constructs containing pooled cells from three different donors were created in the sliding indentation bioreactor system. First, for each sample (n=12) a 2-mm-thick layer of 3% low-melting agarose (type VII; Sigma) was added to the bottom of each trough and was allowed to gel for 20 min at room temperature. Afterward, a small volume of 60 μL of 0.5% agarose containing 50×106 cells/mL was added. This is in the exact plane in which, according to finite element simulations, high tensile strains would occur during sliding indentation (Fig. 1). After the agarose–cell suspension had solidified for 30 min, a 1-mm-thick 3% low-melting agarose layer was added. Then, 7 mL of serum-free culture medium was added to each construct consisting of hgDMEM supplemented with 1% Pen-Strep, 0.1 amphotericin, 0.1 mM dexamethasone, 50 mg/mL ascorbate 2-phosphate, 20 mg/mL
Culture protocol study 2
First, a short-term study (study 2a) was performed, in which constructs containing pooled cells from three donors were cultured for 7 days and were afterward analyzed for gene expression levels of aggrecan and collagen type I and II in depth. Second, a long-term study of 28 days (study 2b) was performed in which constructs containing pooled cells from three different donors were analyzed for the ECM content in depth.
Chondrocyte-seeded agarose hydrogel constructs consisting of 2% (w/v) agarose and a final cell concentration of 20×106 cells/mL were produced (n=12 per study) in bioreactor bottom plates (Fig. 2C). The cell suspensions were equally mixed with 4% (w/v) agarose (type VII). In addition, to determine the equilibrium-compressive moduli of cell-free agarose constructs, three constructs were produced by mixing equal amounts of the culture medium (without TGF-β3) and 4% (w/v) agarose. The constructs were allowed to solidify for 10 min, and subsequently redundant agarose was removed to level the constructs at exactly 3-mm height. Afterward, 7 mL of serum-free culture medium as described in study 1 was added to each construct. The constructs were allowed to equilibrate for 2 days, and two experimental groups were defined. In both studies, the control group contained constructs (n=6) cultured in the bioreactor system without receiving mechanical loading. The other group consisted of constructs (n=6) that were loaded by sliding indentation. Sliding indentation was applied with the same loading parameters as used in study 1. The medium with fresh TGF-β3 was changed every other day, and the removed medium was stored at −30°C for analysis of the released ECM components.
Gene expression analysis
Constructs of the short-term experiment in study 2 were divided into two parts after 7 days of culture. One part was used for full-depth analysis of gene expression levels. The other part was sliced in three parts with a vibratome, creating a top and middle layer with a thickness of each 750 μm and a bottom layer with a thickness of 1500 μm for analysis of the gene expression levels in depth. Total RNA was extracted from the samples using the RNeasy minikit (Qiagen) according to the manufacturer's instructions. Stability of the reference genes was investigated using a geNorm reference gene selection kit (PrimerDesign), and GAPDH showed to be most stable. Primer sets for collagen type I, II, and aggrecan (PrimerDesign) were developed and validated for qPCR. The qPCR (MyiQ; Bio-Rad) was performed using SYBR Green Supermix (Bio-Rad), primers, ddH2, and cDNA. Target gene expression was evaluated using the ΔCT method.
Mechanical testing
Samples (n=6 per group) from study 2 were subjected to mechanical testing in unconfined compression between impermeable platens using a standard material testing machine with a 2 N load cell (Zwick Roell Z010), after 28 days of culturing before further analyses, to determine the equilibrium Eeq with a testing protocol similar to that used by Mauck et al. 19 A cylindrical disc (Ø 6 mm) was punched out of each sample, and the thickness was measured with a microcaliper. Afterward, the samples were placed in a bath with PBS at room temperature and were subsequently equilibrated in creep to a tare load of 0.02 N. From this offset, stress relaxation tests were performed with a ramp speed of 1 μm/s until reaching 20% strain. After the equilibrium was reached, the equilibrium Young's modulus Eeq was calculated from the ratio of the equilibrium stress to the applied strain using testXpert II software.
Biochemical content
In study 1, the tissue-engineered cartilage layer could easily be separated from the cell-free 3% agarose layers with calipers. After 28 days of culture, half of each sample was used for biochemical analysis. In study 2, half of each sample was divided into two parts. One part was used for full-depth analysis of biochemical content, and the other part was sliced with a vibratome, creating top-middle and bottom layers, each with a thickness of each 1500 μm.
Samples from both studies were lyophilized, and subsequently papain digestion buffer (100 mM phosphate buffer, 5 mM
Histology and immunofluorescent staining
The other halves of samples from both groups in both studies (n=6 per group) were fixed for 24 h in 3.7% formalin, paraffin-embedded, and 5-μm slides were cut parallel with the loading direction. Some slides were stained with Alcian Blue (pH=2.5) and Picrisirius Red (both Sigma) to evaluate the distribution of sGAG and collagen, respectively, in the cultured constructs. Hematoxylin was used as nuclear counterstaining. Other slides were dewaxed, rehydrated, and incubated in a Tris–EDTA buffer at 100°C for 20 min. Nonspecific binding was avoided by incubation in 1% bovine serum albumin for 30 min. The samples were labeled with PBS containing monoclonal primary antibodies for type I collagen (1:100; C2456 Sigma) and type II collagen (1:50; II-II6B3 Developmental Studies Hybridoma bank). Alexa 488-conjugated goat anti-mouse secondary antibody labeling (Molecular Probes) for collagen type I and II and propidium iodide nuclear counterstaining were performed to visualize collagen and cells. Bovine tendon and native bovine cartilage were used as positive controls for collagen type I and II antibodies, respectively. Negative controls for both antibodies underwent the same procedure without primary antibody incubation.
Statistical analysis
Statistics were performed with Statgraphics (Statgraphics, Manugistics, Inc.). In both studies, the effect of sliding indentation on the different outcome parameters (gene expression levels of aggrecan, collagen I and II, equilibrium Young's modulus, and amounts of sGAG, DNA, and collagen) was examined for significant differences by a one-way ANOVA with Bonferroni post hoc testing. Statistical significance was assumed for p<0.05.
Results
Study 1
Biochemical content and release
Sliding indentation did not have a significant effect on DNA and SGAG content after 28 days of culture (Fig. 3A, B). Collagen content was significantly higher in constructs that were subjected to sliding indentation compared to control constructs (p<0.05; Fig. 3C). sGAG was released with time in the culture medium in both groups (Fig. 3D). In the unloaded control group, sGAG was released at a constant rate. In contrast, in the loaded group, the sGAG release rate was higher during the first 18 days of culture and was diminished from day 18 till 28 (Fig. 3D). At 28 days, no significant differences in total sGAG release were observed (Fig. 3E).
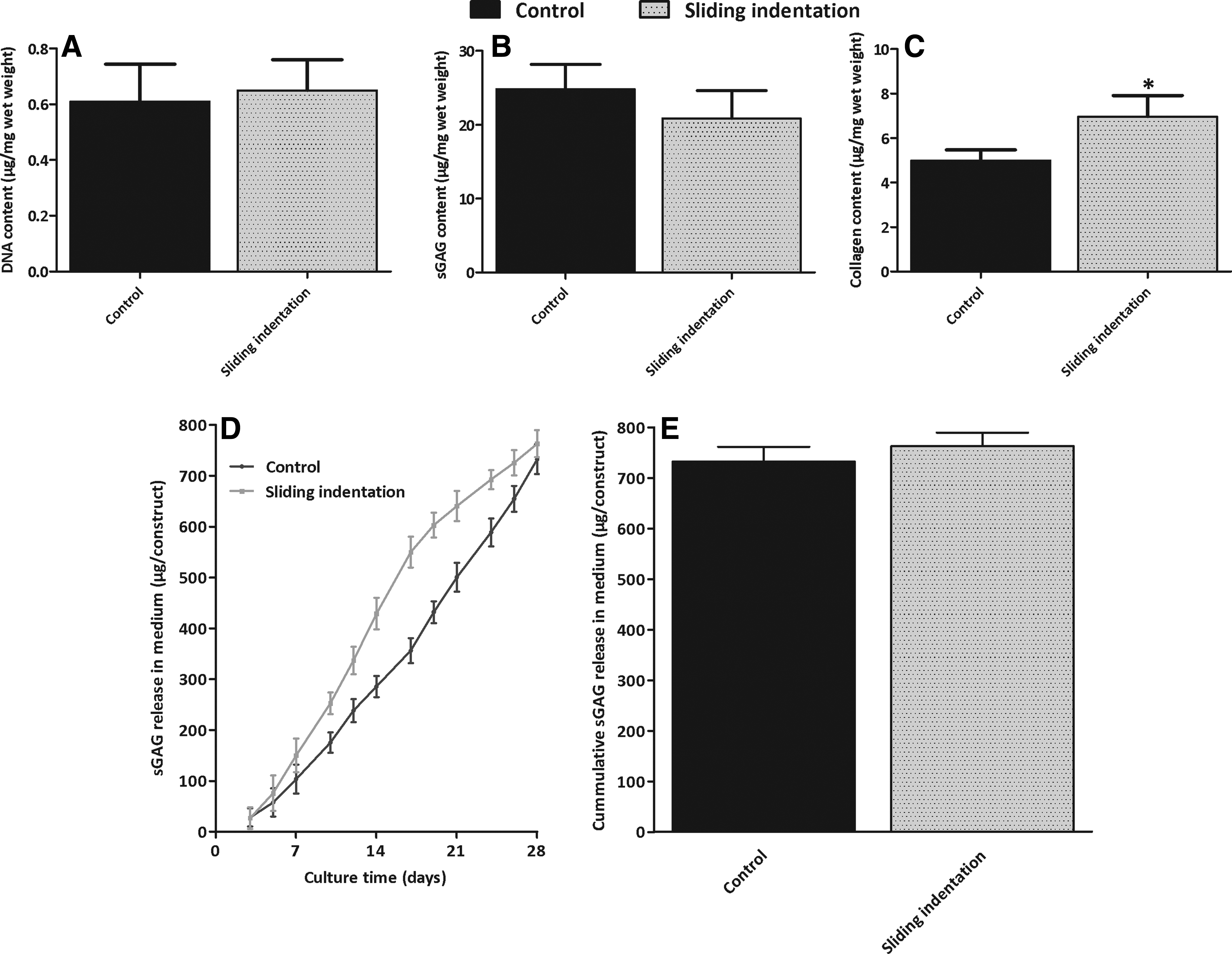
DNA, sGAG, and collagen content in engineered cartilage constructs
Histology and immunofluorescent staining
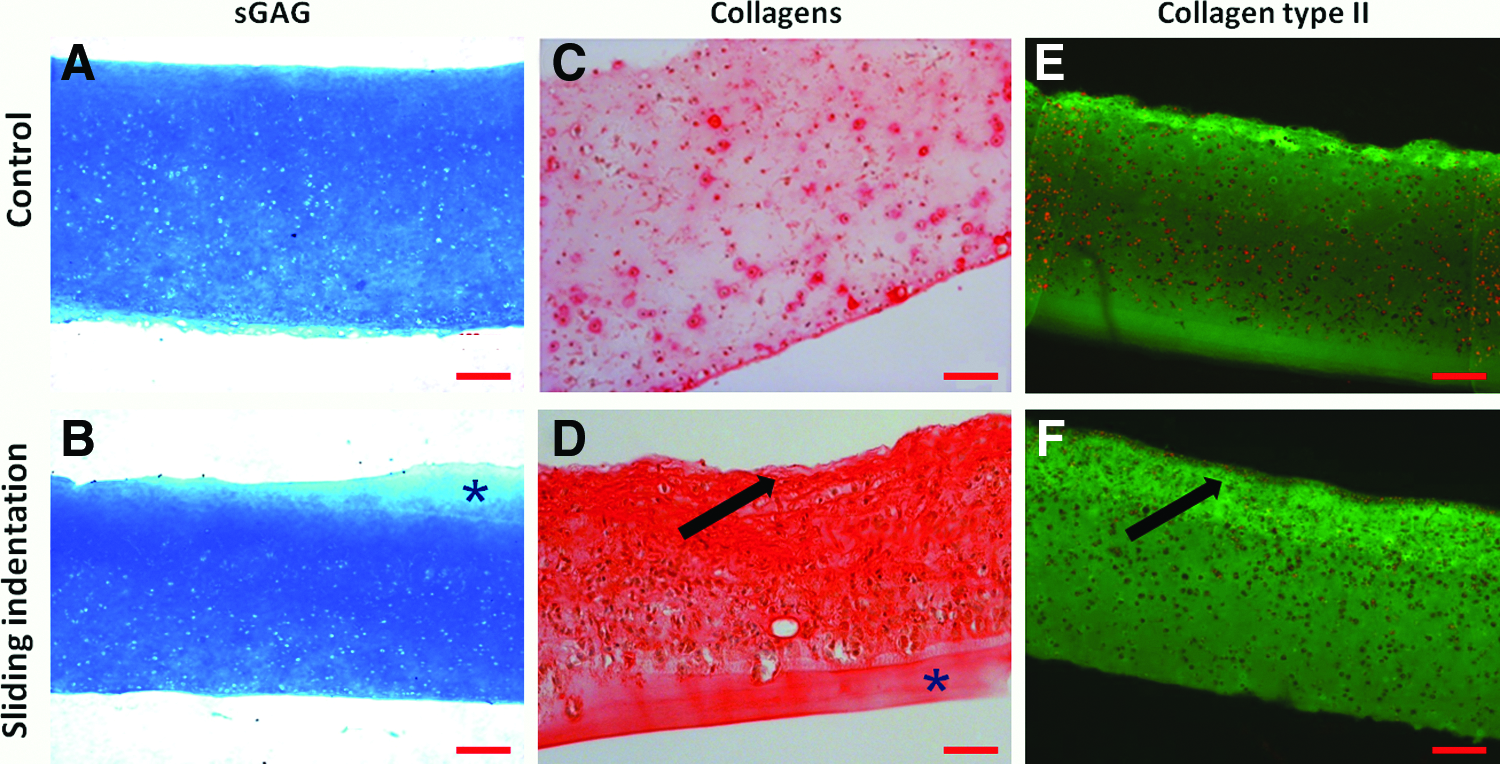
Histology followed the results obtained with the biochemical assays. No difference in the staining intensity for sGAG was found between the unloaded and loaded constructs (Fig. 4A, B). The distribution of the sGAG in the constructs was uniform in both groups. In loaded constructs, collagen staining (Fig. 4D, F) was more intense compared to that of the control constructs (Fig. 4C, E). Further, at the top of the loaded constructs, a denser layer with collagen (type II) was observed (see arrows in Fig. 4D, F), which appears to have a horizontal orientation at a higher magnification. This layer was never found in any of the control constructs. Immunofluorescent staining demonstrated that all the collagen deposited was type II (Fig. 4E, F), and staining for collagen type I was negative in all the samples (data not shown).
Alcian blue staining for sGAG
Study 2
Gene expression analysis
Gene expression levels of aggrecan (ACAN) and collagen type II (COL2A1) were quantified after 7 days of culture. In the unloaded control samples, gene expression levels of both aggrecan and collagen type II were similar in each layer in depth (Fig. 5). Sliding indentation induced a significant increase of aggrecan and collagen type II gene expression levels in total constructs (full depth) compared to the control. Further, it induced a depth-dependent response by chondrocytes. In loaded constructs, aggrecan and collagen type II expression levels were significantly higher in the top and middle layer compared to the bottom and unloaded control (all p<0.05). In the bottom layer, no effect of sliding indentation was observed. Collagen I (COL1A1) was not detectable in any of the samples (except in positive controls). No differences in the gene expression levels between the top and middle layer were observed in any of the constructs. Therefore, in the long-term study, the top and middle layers were taken together, and the constructs were divided in two layers of equal height (1500 μm) before quantification of the biochemical content.

Gene expression levels (2-ΔCT) of
Biochemical content and release
Sliding indentation had no significant effect on DNA content in full-depth constructs after 28 days of culture (Fig. 6A). In the control, significant less DNA was measured in the bottom layer compared to the top-middle layer and full-depth constructs (p<0.05). In the sliding indentation group, the top-middle layer contained significantly more DNA than the bottom layer and full-depth constructs (p<0.05). Sliding indentation significantly decreased sGAG and collagen content in the full-depth constructs and in the bottom layer compared to the control (p<0.05), but had no significant effect on sGAG and collagen in the top-middle layer (Fig. 6B, C). In both groups, significantly less sGAG and collagen were measured in the bottom layer compared to the top-middle layer (p<0.05). sGAG is released with time in the culture medium in both groups (Fig. 6D), but after 28 days of culture, 83%±7% more sGAG was released in the sliding indentation group (Fig. 6E; p<0.05).
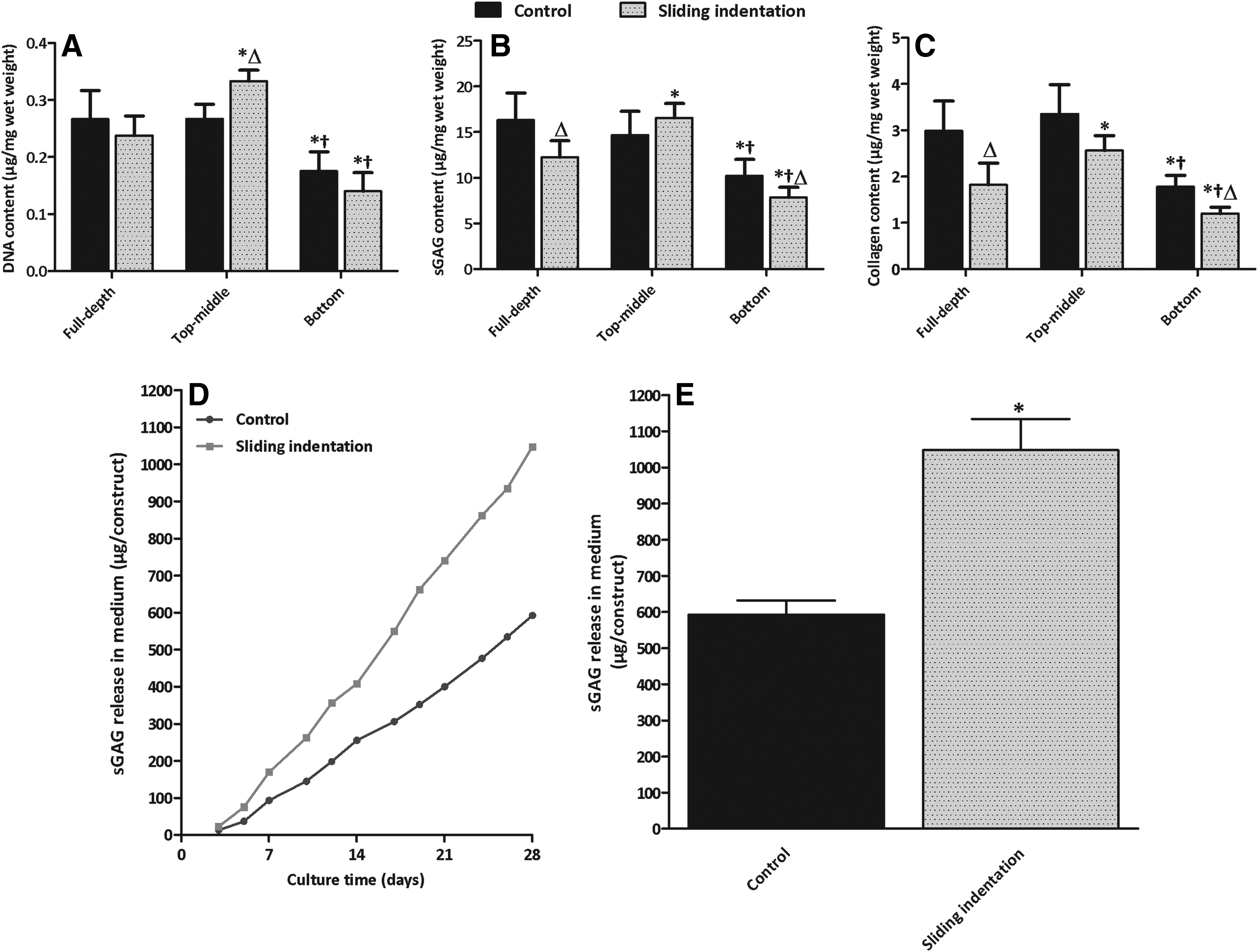
DNA, sGAG, and collagen content in engineered cartilage constructs
Histology and immunofluorescent staining
Histology followed the results obtained with the biochemical assays. In the unloaded constructs, no obvious gradient in Alcian blue staining in depth was observed, but staining for collagen (type II) was less intense in the bottom compared to the top-middle layer (Fig. 7A, C, E). Sliding indentation resulted in a depth-dependent staining intensity for both sGAG and collagen (Fig. 7B, D, F). The intensity of sGAG staining in the upper layer of loaded constructs (Fig. 7B) is comparable to that of the complete control constructs (Fig. 7A). In the bottom layer of loaded constructs, staining is less intense compared to the upper layer (Fig. 7B) and to the control constructs (Fig. 7A).

Alcian blue staining for sGAG
Mechanical testing
All constructs cultured in the bioreactor system for 28 days had a significantly higher Eeq than the empty 2% agarose constructs (both p<0.05). No significant difference was observed between the unloaded and loaded constructs (Fig. 8).
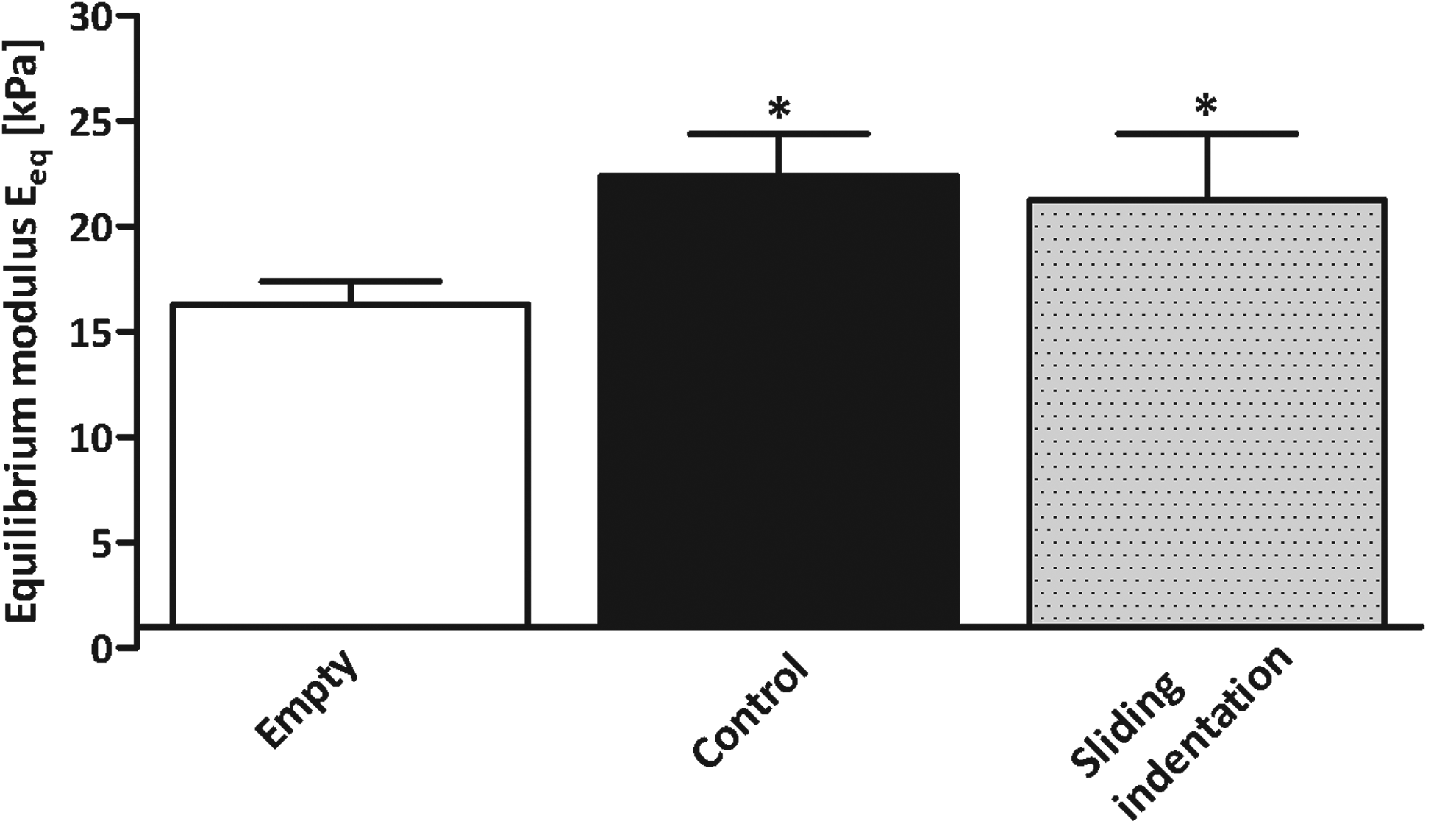
Equilibrium Young's modulus of empty constructs, unloaded controls, and loaded (sliding indentation) samples after 28 days of culture in study 2. Data represent the mean and standard deviation of six samples per group. *p<0.05 versus empty 2% agarose hydrogel.
Discussion
The results of our studies show that application of sliding indentation can increase collagen fractions (study 1) and enhances depth-dependent ECM distribution (study 2) in chondrocyte-seeded cartilage constructs.
Sliding indentation did increase collagen content, but did not influence sGAG content in the constructs, or total sGAG released from the constructs in study 1. However, the sGAG release rate in time did differ between groups. From the control samples, sGAG was released at a constant rate. In the loaded group, the sGAG release rate was highest during the first 18 days of culture and declined during the final 10 days. We speculate that sGAG release in the loaded group in the first phase of culture may be increased, because sGAG is being pushed out as a consequence of the sliding indentation loading regime. In the second phase, the sGAGs may be retained because they are entangled with the developed collagen network, which is known to be a major contributor to the structural integrity of the ECM network.24–26 Since only the collagen content in the loaded groups was increased, we speculate that collagen is a main contributor to better sGAG retention. However, also other factors, such as changes in the fibril diameter or amount of cross-links, or a combination of both, may have contributed to sGAG retention.
Interestingly, many of the loaded samples in study 1 exhibited a dense collagen layer in the top of the construct (Fig. 4D, F). Although this layer seemed to have some kind of fibrous appearance, we demonstrated that it contained collagen type II, while collagen type I was absent. Although we were unable to provide a proof using polarized light microscopy on sections stained with picrosirius red, it appeared as if the collagen fibers in this dense layer were oriented horizontally. This finding is interesting, since it is in agreement with the theories that predict that the fiber orientation may coincide with strain orientations.27,28 Using a numerical model, it was predicted that sliding indentation indeed induces horizontal strains in the region closest under the indenter. 9 It may be useful to explore other methods to detect early changes in collagen alignment in future studies, such as Fourier-transform infrared imaging spectroscopy 29 or second-harmonic imaging microscopy. 30 Also, it is likely that our 28-day culture period was insufficient to enhance collagen synthesis and alignment, and therefore longer culture periods are required.
Sliding indentation increases collagen content in tissue-engineered constructs, and it is therefore likely to improve the tensile strength as well. Unfortunately, we did not measure the tensile strength of constructs in this study, but it is apparent that an increase in collagen fractions leads to increased tensile stiffness of engineered cartilage constructs,6,31 which is favorable for the mechanical quality of the cartilage.
Recently, we have shown that culturing chondrocytes in low agarose concentrations in the presence of TGF-β3 results in a more homogeneous ECM distribution and leads to improved mechanical properties (revisions submitted to this journal). Using the sandwich model, it is possible to apply sliding indentation to the cell-seeded gels with low agarose concentration, without damaging the initially mechanically weak constructs. Using this model, we have previously shown that collagen deposition in chick periosteum is enhanced in response to dynamic tension induced by sliding indentation. 10 Now, we know that sliding indentation can improve the collagen content in thin chondrocyte-seeded agarose layers, and the next step will be to extend the model to more feasible thicknesses of constructs, without limiting diffusion.
The gene expression data from study 2 confirm our hypothesis that a depth-dependent mechanical stimulus can induce a depth-dependent response of chondrocytes. Numerical evaluation showed that the loading regime induces largest strains in the upper part of the construct (Fig. 1). In this same region, we found significantly increased aggrecan and collagen type II gene expression levels. In the bottom layer, where strains are low, no differences in the gene expression levels between were found between loaded samples and controls. This is in agreement with Li et al., who also demonstrated increased mRNA levels for different chondrogenic genes in the top layer of chondrocyte-seeded 3D polyurethane constructs that were subjected to surface motion. 32 However, in native articular cartilage, the proteoglycan content is higher in the middle and the deep zone compared to the superficial zone. Our results and those from others31,32 suggest that a sliding loading regime has a general stimulating effect on chondrogenic gene expression in the region close to the indenter and shows that cells in these region are mechanosensitive. In addition, it should be noted that cells in the top and middle layer are experiencing a complex mechanical environment, with compressive and tensile strains, fluid flow, and fluid pressurization in different directions.
The gene expression results did not translate into similar results on the protein expression level on the long term. sGAG and collagen contents were significantly lower in the loaded constructs compared to the control constructs. Even though the gene expression levels increased in the upper half of the loaded constructs, this did not translate into more sGAG or collagen in the same region. Based on our observations on sGAG and collagen quantities as well as their distributions, we may interpret these results as follows. Sliding indentation resulted in more aggrecan gene expression, which translated into more sGAG protein synthesis. However, as a consequence of the loading regime, sGAGs were expelled from the constructs into the culture medium. In unloaded constructs, in total 1570 μg of sGAGs was synthesized and in loaded constructs 1823 μg. However, in the loaded group, 57% of the totally synthesized sGAGs were released in the culture medium, whereas this was only 38% in the control. These results indicate that sliding indentation has a positive effect on ECM anabolic activity by chondrocytes, but a negative effect on matrix retention, especially in the region that is in close contact with the indenter. In the deeper zone, sliding indentation did not significantly enhance sGAG synthesis, but some sGAGs may be released from this area as well, resulting in a finally reduced content compared to the control.
sGAG and collagen contents were lower in the bottom layer of both unloaded and loaded constructs. This depth-varying effect in unloaded constructs may be caused by gradients in nutrient, oxygen, or growth factor transport toward chondrocytes in the deeper zones, due to the fact that the constructs were positioned on an impermeable bottom plate. However, this did not induce the differences in viability in depth in any of the constructs (data not shown), but it may have influenced cell metabolism. It has been reported that a nonhomogeneous oxygen, nutrient, and growth factor distribution can lead to reduced proliferation and ECM synthesis in the inner or deep regions of cultured cartilage constructs,33,34 and that this is dependent of the scaffold thickness, diameter, concentration,35,36 and ECM development within the constructs. 37 Possible diffusion limitations may be prevented by replacing the impermeable bottom plate by a permeable one. In future studies, it would be worthwhile to augment the numerical studies to include mass transport through the constructs and the effect of the sliding indentation loading regime thereon, since these may influence each other.
Interestingly, in study 1, we do see an increase in collagen content, but this effect is absent in the top-middle layer of loaded constructs in study 2b, in which the predicted strains were similar to study 1. In constructs cultured in study 1, the cell density was higher (2.5 times), and agarose concentration was lower (4 times) compared to the constructs in study 2. Both factors are known to influence the microenvironment of cells. For example, cells seeded in 0.5% agarose may be more mechanosensitive than cells in stiffer 2% agarose, since they can more easily deform and adapt their morphology in response to mechanical loading. Besides, we have recently shown that ECM deposition by chondrocytes is increased and more uniform in agarose percentages below 1%, probably due to increased pore size and number (unpublished data). On the other hand, the increased cell density in study 1 compared to study 2 may have led to differential cell–cell and cell–matrix interactions, which may in turn have affected the mechanosensitivity and mechanoresponsiveness of the chondrocytes.
The equilibrium Young's modulus of cartilage is determined to a great extent by sGAGs, which function to resist compressive forces. 38 Although the sGAG content in loaded constructs was significantly lower than in the unloaded constructs, this did not lead to differences in equilibrium Young's modulus of the constructs. Similarly, in a study by Bian et al. using a comparable loading regime, the equilibrium Young's modulus was significantly increased in loaded constructs compared to unloaded, but no differences in bulk biochemical content were observed. 39 Based on our results, we speculate that the depth-varying distribution affects the bulk equilibrium Young's modulus of the constructs, although the mechanism behind this connection is unclear. This speculation is in agreement with computational predictions. 40 Besides that, sliding indentation might lead to differences in the amount of other matrix molecules, such as COMP, link protein, fibronectin, other collagen types, and collagen cross-linking, which may also contribute to the mechanical properties of the engineered tissue.25,41,42 Thus, these data show that it is not the absolute amounts of sGAG and collagen determining the mechanical properties of the constructs, but that distribution of matrix components or presence of other matrix components may be responsible, and this requires further investigation. Also, it would be interesting to evaluate depth-dependent inhomogeneity in mechanical properties, by assessing compressive and tensile moduli for the layers in depth separately. Further, it should be noted that the mechanical stiffness of engineered cartilage achieved in this study is still well below that of the native cartilage tissue. Although for clinical application, it is still to be determined what level of mechanical maturity will be required for the engineered cartilage implants to survive in load-bearing joints, it is likely that enhanced mechanical properties will reduce the potential of mechanical failure.
Our loading regime leads to loss of matrix components into the culture medium, especially when using conventional agarose constructs. Such a loss of matrix may be prevented by starting the loading regime after a period of preculture, or possibly by modifying the agarose-sandwich model in such a way that a depth-varying mechanical stimulus prevails, while protein retention is impaired.
Conclusion
In conclusion, this study demonstrates that sliding indentation is a promising loading regime for enhancing collagen synthesis by chondrocytes seeded in an agarose-sandwich model, and to induce a depth-dependent response by chondrocytes seeded in agarose. This is a major conclusion that may help developing tissue with depth-dependent matrix content, structure, and consequently mechanical properties and mechanical properties closer to that of native cartilage than currently possible.
Footnotes
Acknowledgments
The authors thank Mehdi Khoshgoftar for performing the finite-element simulations. This study was supported with funding from the Dutch Technology Foundation STW (VIDI-EGT.07970).
Disclosure Statement
No competing financial interests exist.