
Abstract
Since there is currently no optimal treatment for chronic neck or back pain that restores full spine functionality and disc height, recent research has focused on developing a regenerative medicine approach. This requires a better understanding of the phenotype nature of the different cell types present in disc tissues. In particular, there is very little known about the differentiated states of cells that co-exist within the annulus fibrosus (AF), despite the critical importance of this tissue in maintaining disc functionality. Maintenance of the differentiated states of these AF cells is imperative to the production of appropriate extracellular matrix (ECM) macromolecules and to the engineering of functional AF tissues. The objective of this study was to determine whether inner annulus fibrosus (IAF) cells when grown on polycarbonate urethane (PU) scaffolds in vitro will produce ECM molecules characteristic of IAF cells and different from outer annulus fibrosus (OAF) cells. OAF and IAF cells isolated from bovine coccygeal intervertebral disc were grown on nanofibrous PU for approximately 14 days. The effect of culture time on ECM gene expression, DNA content, and the synthesis and retention of proteoglycans and collagens were evaluated for both OAF and IAF cells. The ECM accumulated was also characterized by immunostaining and Western blot. The tensile strengths of the tissue/scaffolds were evaluated at 14 days. Both OAF and IAF cells both attached to PU and had similar DNA contents over time. IAF cells maintained relatively higher levels of COL2A1, ACAN and VCAN gene expression, and relatively lower levels of COL1A1 gene expression when compared with OAF cells, by 10 days of culture. IAF cells synthesized and retained similar amounts of total collagen and proteoglycans when compared with OAF cells. While both OAF and IAF cells accumulated type I collagen, only IAF cells accumulated type II collagen. Both cell types accumulated similar amounts of aggrecan but IAF cells accumulated higher amounts of versican as determined by immunostaining. In conclusion, nanofibrous PU scaffolds enabled the maintenance of most of the characteristic features of the IAF cell phenotype, and were different from those of OAF cells.
Introduction
T
A number of different approaches have been developed to attempt to repair or replace the disc ranging from injection of cells to the replacement of the damaged tissue with in vitro formed tissue.7–9 Given that the disc is always weight bearing, attention has focused on replacing damaged tissue. However, identifying the conditions to recapitulate the AF has been particularly problematic. The AF has a complex organization composed of multiple lamellae of collagen fibrils angled at ∼60o to the vertical axis and although the angle is the same, the direction of the inclination alternates with each successive lamella, which is considered as helping confer resistance to tensile forces and permitting the twisting and bending of the spine. 6 The AF cells in the lamellae are aligned parallel to the collagen fibers. The composition of the AF varies depending on its location within the IVD. The inner aspect of the AF adjacent to the NP is rich in proteoglycans and contains lamellae composed predominately of type II collagen. These macromolecules, proteoglycans, and type II collagen decrease with increasing distance from the NP and the amount of type I collagen increases. Due to the differences in the extracellular matrix (ECM) composition, the AF is divided into an inner AF (IAF) and outer AF (OAF). Given the complexity of this organization, most of the attempts to bioengineer AF have consisted of growing cells on scaffolds in order to guide tissue architecture.10–14 A number of different scaffolds have been utilized, each with their unique strengths and limitations.10,14–16 More recently, scaffolds made of a degradable polycarbonate urethane (PU) and containing a surface modifier to bind fibronectin have been shown to be useful for soft tissue engineering, because they form a mechanically strong and elastic substrate for cell adhesion and ECM accumulation while undergoing resorption in vivo.17–19 This polymer can also be electrospun into aligned nanofibers, which enables one to simulate the orientation of the collagen fibers in a single lamella of the AF.18,19 Modulation of the PU scaffolds through the addition of an anionic dihydroxy oligomer (ADO) increases surface polar chemistry and improves AF cell attachment and ECM retention on the PU scaffold in a manner that mimics AF tissue development when coated with fibronectin.17,19 Furthermore, this biomaterial is reported to be biocompatible as it degrades into products that are not toxic to AF cells and ultimately generates CO2 and hydroxyl hydrocarbons which will not incite an inflammatory reaction.18,20 While the behavior of OAF cells on these PU nanofibrous scaffolds has been characterized, it is not known whether IAF cells will retain their different phenotype on these scaffolds. Given the differential nature of OAF and IAF cells, it is critical to assess this question in order to ultimately achieve successful application of these new scaffolds. Therefore, the aim of this study was to determine whether IAF cells when grown on PU scaffolds in vitro will produce ECM molecules characteristic of these cells and different from OAF cells.
Materials and Methods
Synthesis of polycarbonate urethane (PU)
The base polyurethane PU was synthesized from the reaction of poly(1,6-hexyl 1,2-ethyl carbonate)diol, 1,6-hexane diisocyanate and 1,4-butanediol in N,N-dimethylacetamide (DMAC) at a temperature between 60°C and 70°C. 17 ADO was synthesized by reacting polytetramethylene oxide, hydroxyethylmethacrylate (HEMA), and lysine diisocyanate in DMAC overnight at a temperature between 50°C and 60°C. Lysine and HEMA ester groups were then hydrolyzed, forming hydroxyl and carboxylate groups. Synthesized ADO was added to PU at a concentration of 0.5% by weight, and PU nanofibrous scaffold was fabricated by electrospinning onto a rotating mandrel at 1200 rpm and an applied potential difference of 20,000V. The scaffold was dried in an Isotemp Vacuum Oven (Model 280A; Fisher Scientific, Pittsburgh, PA) overnight at 45°C. A circular sample of scaffold was cut using a biopsy punch (8 mm diameter) and was placed within a custom fabricated holder consisting of a 200 μL microfuge tube with the tip cut off, as previously described. 19 The construct was then sterilized by gamma irradiation (4 M Rad). Before cell culture, the scaffold was soaked in sterile phosphate-buffered saline solution without Ca2+ and Mg2+ (PBS−/−) containing fibronectin (20 μg/mL F0895; Sigma-Aldrich, St. Louis, MO) overnight at 4°C and then rinsed thrice with PBS−/−. 19
Cell isolation and culture
The IVDs were harvested aseptically from bovine caudal spines and placed in serum-free Dulbecco's modified Eagle's medium (DMEM) supplemented with 1% antibiotic-antimycotic (1000 U/mL penicillin G, 1000 μg/mL streptomycin sulfate, and 250 ng/mL amphotericin; Invitrogen, Carlsbad, CA). The OAF has a distinct lamellar organization that facilitates its identification on gross inspection of the disc. In contrast, the lamellar organization of IAF is not as prominent due to the intermixed proteoglycan-rich ECM, which makes the tissue softer and easy to identify as distinct from the OAF and NP. The efficacy of the dissection was confirmed by submission of random pieces of OAF and IAF for histological assessment. Each tissue type was cut into pieces and separately digested for 90 min with 0.3% protease. Subsequently, 0.2% collagenase for OAF and 0.1% collagenase for IAF were used in overnight incubation at 37°C. Cells were filtered using a sterilized mesh, washed thrice with DMEM supplemented with 5% fetal bovine serum (FBS). OAF and IAF cells were seeded separately onto Fn-coated PU scaffolds (2100 cells per mm2) in DMEM supplemented with 5% FBS in a 24-well plate. The next day, DMEM supplemented with 20% FBS was added to the cultures to a final volume of 2 mL. Ascorbic acid (Sigma-Aldrich) was added on the third day of culture (100 μg/mL final concentration). This media was changed every 2 or 3 days.
DNA content and cell viability assessment
Cultures harvested at 1, 3, 7, 10, and 14 days were digested with papain (40 μL/mL; 20 mM ammonium acetate, 1 mM-EDTA, and 2 mmol/DTT at pH 6.2; Sigma-Aldrich) for 48 h at 65°C. DNA content was determined using Hoechst 33258 dye (Polysciences, Warrington, PA) binding assay and fluorometry (excitation wavelength 365 nm, emission wavelength 458 nm) as previously described. 21 Cell viability was assessed at the end of the culture period using LIVE/DEAD® Viability/Cytotoxicity Kit (L-3224; Invitrogen) according to the manufacturer's instructions and visualized by confocal microscopy. At least 12 fields (ranging from 90 to 130 cells for each sample) were analyzed per sample.
Gene expression analysis
Total RNA was extracted from IAF and OAF cells using Trizol (Invitrogen) according to the manufacturer's directions. RNA concentration was measured at 585 nm using Nanodrop (Thermo Scientific, Waltham, MA). Two hundred nanogram of total RNA was reverse transcribed with Superscript III Reverse Transcriptase (Invitrogen) followed by relative quantitative PCR (qPCR) performed using SYBR Green mastermix (Qiagen, Germantown, MD) with 1 μM of each primer pair. After incubation at 95°C to activate the enzyme, the amplification parameters were 10 s denaturation at 95°C followed by 30 s annealing at 60°C. Mean relative quantification values were calculated using the ΔΔCT method with 18S rRNA serving as the endogenous control. The primers for 18S, ACAN, COL1A1, and COL2A1 were published previously elsewhere. 22 The VCAN primers were forward 5′-TCG GAC CAC AGG CAT TTT CT-3′ and reverse 5′-AGA TCG TGC TCT GGA TCT GT-3′. All primer pairs had an efficiency of >90%.
Quantification of collagen and proteoglycan synthesis
Synthesis of collagen and proteoglycan by OAF or IAF cells was quantified by incubating the cells with 2 μCi of both [3H]-proline and [35S]-SO4 (Perkin Elmer, Waltham, MA). After 24 h, the media was collected and the cultures were washed thrice (5 min per wash) with PBS calcium and magnesium-free (PBS−/−) to remove unincorporated isotopes. The cultures were then digested by papain as described earlier. To quantify the collagen and proteoglycan released, aliquots of the media was precipitated with either 70% ammonium sulphate or cold 100% ethanol, respectively. The precipitated collagen and proteoglycan were centrifuged at 14,000 rpm for 30 min at 4°C. The pellets were washed thrice with cold 70% ethanol and centrifuged for 10 min at 14,000 rpm after each wash. The collagen was resuspended in 10% sodium dodecyl sulfate (SDS), and the proteoglycans were solubilized in 4 M guanidine hydrochloride. The amount of radioisotope incorporated into the matrix or released into the medium was quantified by β-scintillation counter (Beckman Coulter, Mississauga, ON, Canada).
Immunostaining
IAF and OAF cells cultured on PU were harvested at 2 weeks, fixed in 4% paraformaldehyde for 10 min at room temperature, washed thrice using PBS−/−, and then stored in PBS−/− at 4°C until they were stained. Cells were permeabilized with 0.2% Triton X-100 for 15 min at room temperature, blocked with 5% FBS in PBS−/− containing 0.1% Triton X-100 for 1 h in room temperature, and then incubated with anti-collagen type I antibody (dilution 1:250; rabbit polyclonal; Meridian Life Science, Memphis, TN), anti-collagen type II (clone 6B3, dilution 1:250; mouse monoclonal; Labvision, Fremont, CA), anti-aggrecan antibody (clone 6B4, dilution 1:50; mouse monoclonal; Abcam, Cambridge, MA), or anti-versican (clone 12C5, 1:50; mouse monoclonal; Developmental Studies Hybridoma Bank, University of Iowa, Iowa City, IA) overnight at 4°C. The samples were then washed thrice in PBS−/− (10 min each). The samples were incubated with secondary antibody, either goat anti-mouse IgG or goat anti-rabbit IgG (conjugated with AlexaFluor 488 or 596; Invitrogen) depending on the source of the primary antibody for 1 h at room temperature. After washing and counterstaining with DAPI, sections were mounted with Permafluor Mountant Solution (Thermo Scientific) and coverslipped. Images (60×) were collected at a single representative focal plane at a resolution of 1024×1024 pixels using Nikon Eclipse C1si (Nikon Corporation, Melville, NY). Aggrecan staining was quantified using Image J Software by normalizing the area stained by the number of cell nuclei per image. Twelve fields were counted per culture, and a total of 10 to 35 cells were counted per field.
Representative tissue from the bovine caudal disc were fixed in 10% formalin overnight, then in 30% sucrose in PBS−/− overnight and frozen in Optimal Cutting Temperature compound (Sakura Finetek, Tokyo, Japan). Seven micrometer sections were cut, pretreatment was done as previously described, 23 and immunostaining was performed as described earlier.
Western blot
OAF and IAF cultures were incubated in RIPA buffer (50 mM Tris-HCl, pH 7.4, 150 mM NaCl, 1% Nonidet P-40, 0.25% sodium deoxycholate, and 1 mM EDTA) containing protease inhibitors (PI) on a rotator for 30 min in 4°C and then were treated with an equal volume of chondroitinase ABC (Sigma-Aldrich), containing 0.2 units of chondroitinase ABC and PI (0.2 M Tris, pH 8.0, 100 mM sodium acetate) overnight at 37°C. 24 Three to four scaffolds were pooled to collect sufficient protein. The samples were boiled for 10 min, and 20 μg of protein was separated by SDS-PAGE (top half 4% and bottom half 8% polyacrylamide gel). After transfer to nitrocellulose at 30 V overnight, the blots were blocked in 5% skimmed milk powder. The blots were probed with mouse monoclonal anti-versican antibody (1:600 diluted in PBS with 0.00005% Tween® 20) that was kindly provided by Professor Firoz Rahemtulla, University of Alabama, Birmingham, AL. To normalize loading, the blots were probed with mouse monoclonal anti-α-tubulin antibody (1:2000 diluted in PBS with 0.00005% Tween 20) (mouse monoclonal; Abcam). Immunoreactivity was detected using horseradish peroxidise-conjugated goat anti-mouse secondary antibody. Immunoreactivity was visualized using an ECL detection kit (GE Healthcare, Waukesha, WI) and quantified by densitometry using Image J.
Tensile testing
After 2 weeks in culture, the PU scaffolds cultured with either OAF cells or IAF cells, or which were cell free were cut into rectangular strips (average size: 8 mm×2 mm×170 μm) and subjected to tensile testing. Samples were secured at either end using metallic clamps, and the elastic modulus was evaluated using a mechanical testing device (Instron® model 8501; Instron, Norwood, MA) under a tensile strain of 10 mm per minute to breaking point with a 50 N load cell. 18
Statistical analysis
The IVDs from at least two separate animals were pooled for each experiment to ensure sufficient number of cells. Each independent experiment was repeated thrice. All conditions (except for Western blots) were done in triplicate for each experiment, and results are expressed as mean±95% confidence intervals. Unpaired t-test was used to compare two groups. One-way analysis of variance with the Tukey's post hoc test was used to evaluate significance if more than two groups were compared. Significance was assigned at p<0.05.
Results
OAF and IAF cells attach and proliferate on PU
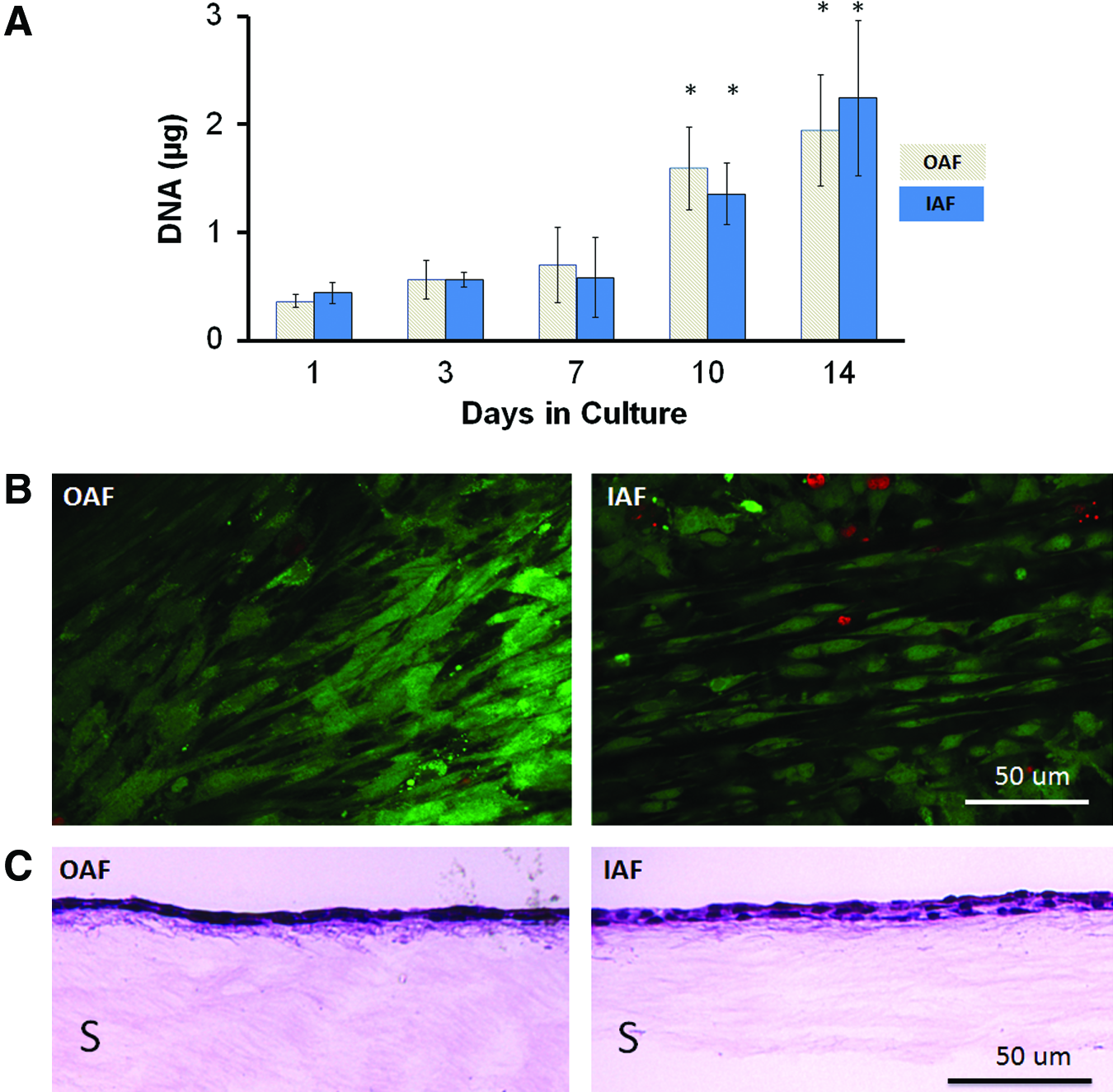
DNA quantification of cells on PU scaffolds harvested at 1, 3, 7, 10, and 14 days showed that OAF and IAF cellularity increased over time and was significantly increased by 10 days (p<0.01) when compared with day 1, 3, and 7 (Fig. 1A). There was no statistical difference in the amount of DNA content accumulated by OAF or IAF cells at any time point. The OAF and IAF cells grown on PU remained viable over the 2 weeks of culture (viability at 2 weeks: OAF cells: 94.5%±3.1% and IAF cells: 94.8%±1.9%) (Fig. 1B). Histological examination of the cultures at 2 weeks showed the presence of cells surrounded by a small amount of ECM (Fig. 1C).
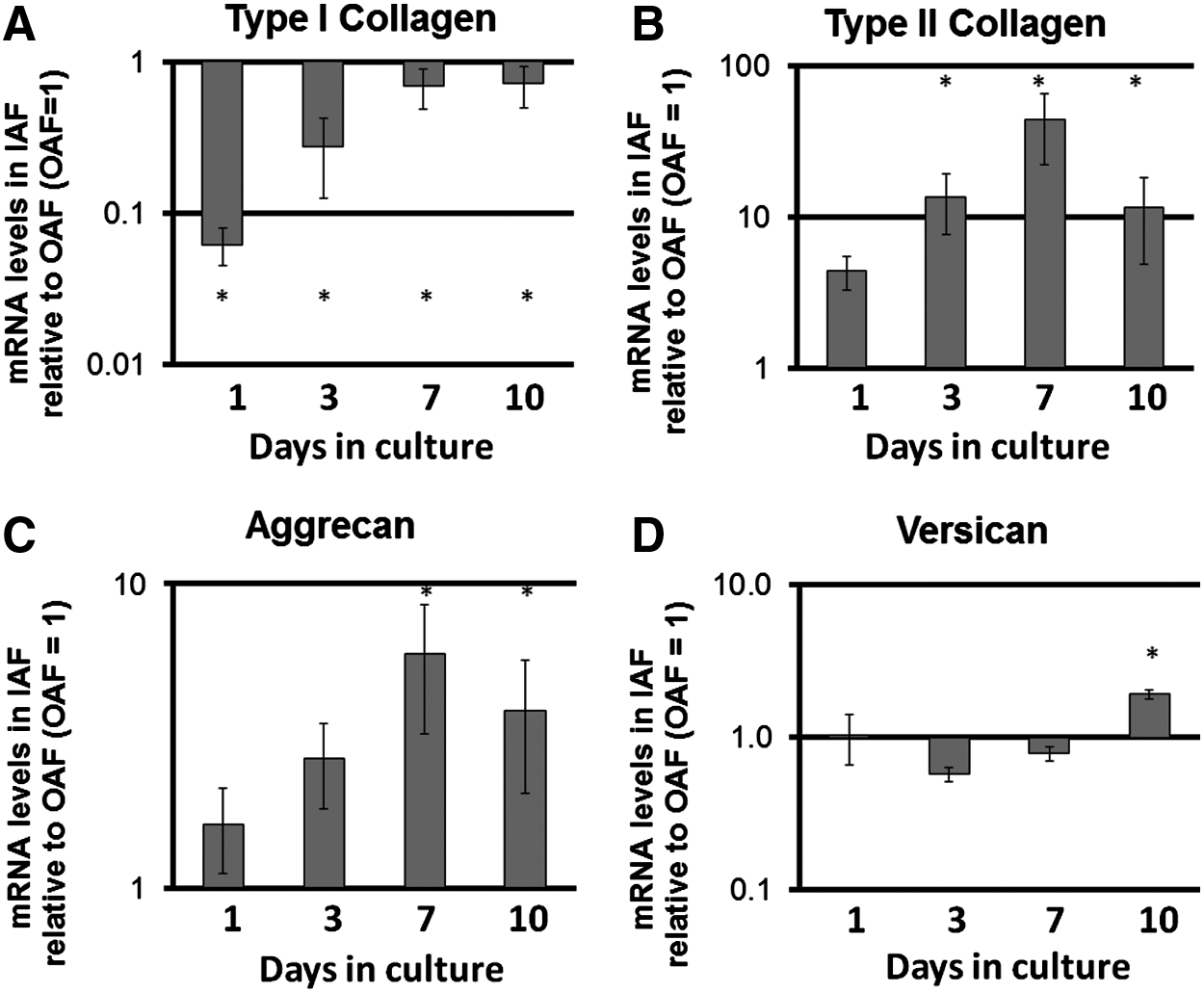
OAF and IAF cells differ in expression of ECM genes
IAF cells expressed significantly lower levels of COL1A1 and higher levels of COL2A1 when compared with OAF cells on days 3, 7, and 10 of culture (Fig. 2). IAF cells also showed a trend to relatively higher levels of ACAN expression, which was significantly greater by day 7 when compared with OAF cells (Fig. 2), and significantly higher levels of VCAN were expressed by day 10 (Fig. 2).
Relative expression levels of
OAF and IAF cells do not show significant differences in collagen and proteoglycan synthesis
OAF cells synthesized similar amounts of total collagen throughout the 14 days of culture, while collagen synthesis by IAF cells was variable (Fig. 3A). Collagen synthesis increased over time until day 10, after which it began to decrease to levels similar to those of day 1 of culture (Fig. 3A). Collagen retention was similar for OAF and IAF cells, except at day 7, at which time IAF cells retained significantly less collagen than OAF cells (Fig. 3B).

Collagen and proteoglycan synthesis and retention by OAF and IAF cells  were determined by quantifying radioisotope [3H]-proline (collagen)
were determined by quantifying radioisotope [3H]-proline (collagen)
Both OAF and IAF cells synthesized similar amounts of proteoglycans, which decreased significantly by day 10 (Fig. 3C). However, concomitantly retention increased significantly by 10 days of culture (Fig. 3D). There was no significant difference in the synthesis and retention of collagen and proteoglycans between OAF and IAF cells.
ECMs accumulated by OAF and IAF cells are distinct
The IAF and OAF cell cultures were stained for the major macromolecules that are known to be differentially expressed by the two cell types. Immunostaining for type I and type II collagen over time showed that OAF cells accumulated only type I collagen on PU, while IAF cells synthesize and accumulate both type I and type II collagen (Fig. 4A).
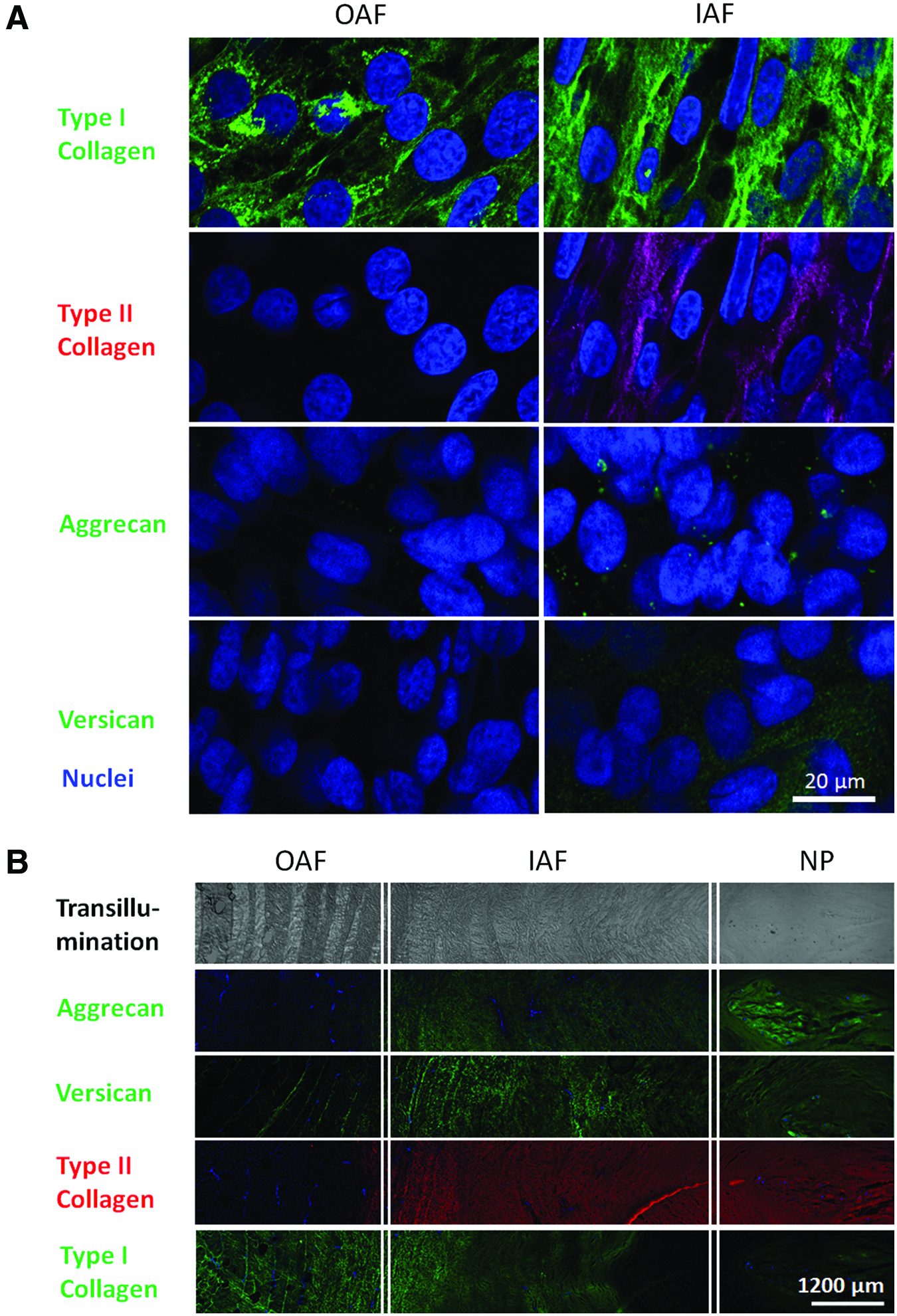
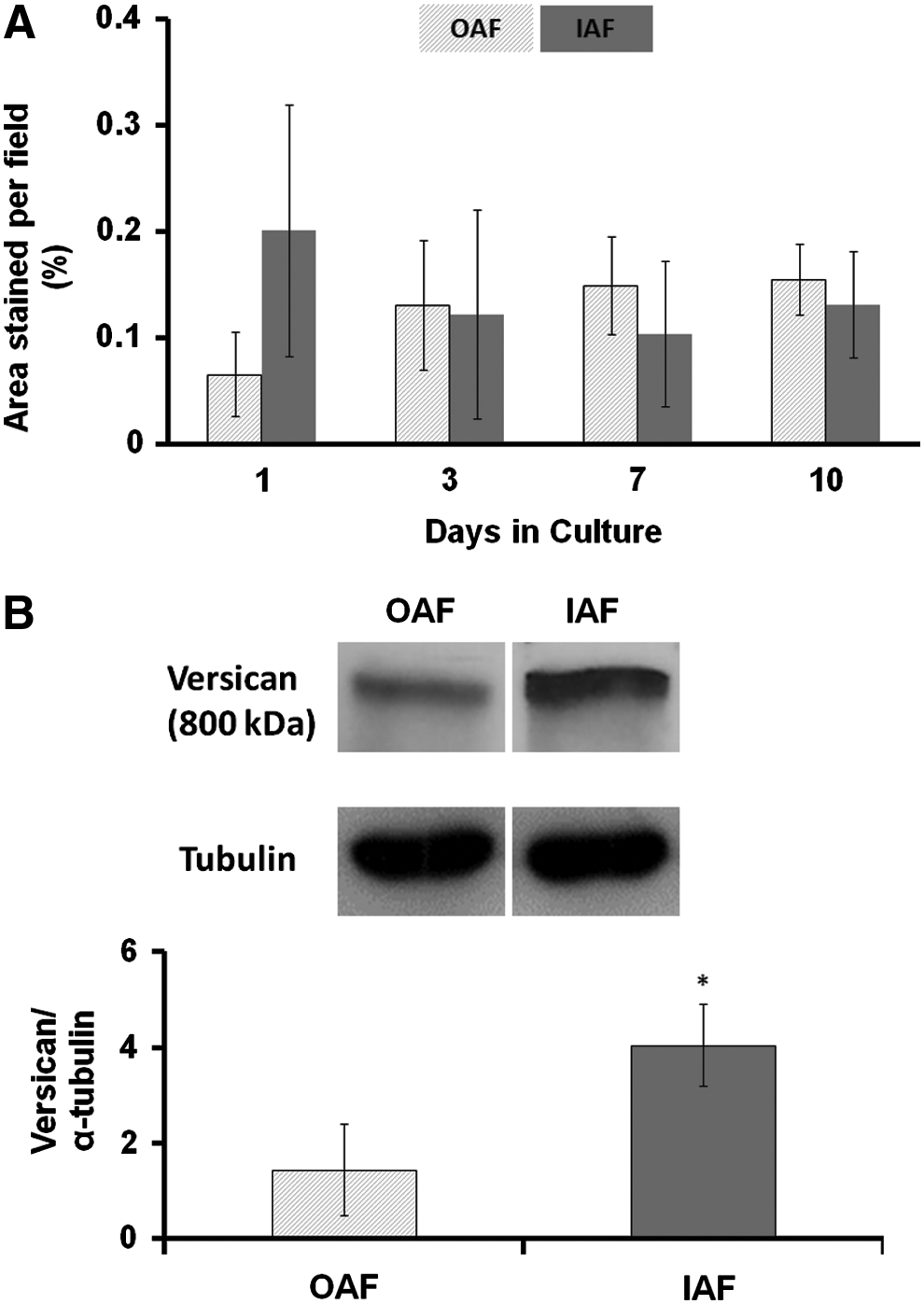
Both IAF and OAF cells accumulate similar amounts of aggrecan in their ECM as shown by immunostaining (Fig. 4A) and quantification by morphometric measurement (Fig. 5A). However, IAF cells accumulated more versican than OAF cells as seen by immunostaining (Fig. 4A). This difference was confirmed by quantification of versican by western blotting and densitometry (Fig. 5B). Native bovine disc is shown for a comparison (Fig. 4B).
OAF and IAF cells increased the tensile strength of PU
The cells accumulated enough ECM sufficient to increase the strength of the scaffold by 14 days of culture. The elastic modulus and ultimate tensile strength measured for PU scaffolds cultured with OAF or IAF cells for 2 weeks were significantly higher than PU scaffolds grown in culture without cells (p<0.05) (Fig. 6). There was no significant difference in the elastic modulus and the ultimate tensile strength measured between PU cultured with OAF cells or IAF cells.

Discussion
This study demonstrated that both OAF and IAF cells attach to a degradable PU scaffold and accumulate ECM which increases the strength of the scaffold as early as 2 weeks in culture, as shown by a significant increase in ultimate tensile strength. In addition, the cells maintained aspects of their individual phenotype in vitro. IAF cells showed differential collagen accumulation as OAF cell cultures contained only type I collagen while IAF cells accumulated both type I and type II collagen, characteristic of IAF cells in the native disc.1,25 IAF cells also accumulated more versican when compared with OAF cells as shown by immunostaining and confirmed by western blot and densitometry. This is also a characteristic feature of IAF cells in the native bovine disc. 26
This is believed to be the first study which demonstrates in an in vitro setting using a synthetic elastomeric degradable scaffold that type II collagen is accumulated by IAF cells but not OAF cells, as other reports in the field have only shown that OAF and IAF cells can have differential gene expression profiles of COL1A1 and COL2A1 when grown in vitro. In a study by Chou et al., OAF and IAF cells were cultured in a monolayer on rigid plastic tissue culture polystyrene (TCPS) and passaged twice, showing that the cells maintained their respective genotypes, as gene expression of COL1A1 was higher for OAF cells when compared with IAF cells; while IAF cells expressed higher levels of type II collagen when compared with OAF cells. 27 When these P2 cells were placed in 3D culture in alginate or polyester ester mesh, they lost these differences in COL1A1 and COL2A1 gene expression and analysis by ELISA showed no differences in type I or type II collagen accumulation between IAF and OAF cells by 14 days in culture. 27 This study underlines the importance of the material on which disc cells are grown, as it can influence the maintenance of cell phenotype. In keeping with this, porcine IAF and OAF cells have also been shown to undergo a shift in gene expression levels when grown as a monolayer on TCPS when compared with being grown in 3D within alginate. 28 IAF cells expressed lower levels of aggrecan when cultured as a monolayer on TCPS as compared with culture in alginate. 28 When cultured as a monolayer on TCPS, IAF cells also expressed similar levels of COL2A1 as OAF cells. 28 Bovine IAF and OAF cells exhibited greater proteoglycan synthesis in alginate or collagen gels versus cells grown in a monolayer culture on TCPS. 29 While bovine IAF cells exhibited greater proteoglycan synthesis when cultured in alginate and as a monolayer on TCPS, as compared with OAF cells cultured in the same environment, the difference in proteoglycan synthesis was lost when these cells were cultured in collagen gel. 29 Passaged human AF cells, when cultured in collagen sponges, demonstrated the most ECM production in comparison to cells grown on collagen gel, fibrin, agarose, and alginate gels. 30 This further supports the concept that biomaterials influence the types of molecules which are synthesized and accumulated and that this microenvironment can profoundly influence cell metabolism and phenotype. This has been shown for other cell types as well such as osteoblasts and chondrocytes.31,32
It is not known why PU scaffolds promote the maintenance of AF cell phenotype, although it was shown in a previous work that these materials enable proteins (such as fibronectin and collagen) to adsorb in a manner that permits differentiation in their ability to direct cell shape and protein synthesis. 19 Polyurethanes have polar and nonpolar domains that, if well distributed on the surface, may minimize denaturation of adsorbed proteins.17,33,34 The studies by Attia et al. further showed that the manner by which matrix was accumulated by AF cells when grown on PU, in some ways, mimicked fetal AF development in vivo. 19 During IVD formation, the sclerotome surrounds the notochord and establishes cell alignment before the deposition of collagen, either type II collagen in the IAF primordium or type I collagen in the OAF primordium.35,36 AF cells grown on PU also show alignment before matrix accumulation occurs. Perhaps alignment is the defining factor, as studies using keratinocytes have shown that scaffold alignment can influence collagen synthesis and the type of matrix produced. 37 Hence, it may be very possible that both the molecular distribution of chemistry on the fibers in combination with the nanostructural arrangement of the fibers induce the cells to synthesize and accumulate the appropriate collagen. This hypothesis will require further studies.
With time in culture, proteoglycan synthesis decreased for both IAF and OAF cells while proteoglycan retention increased. The reduction in proteoglycan synthesis, which was accompanied with an increase in proteoglycan retention, was previously seen in bovine chondrocytes cultured on Millicell CM® filter inserts. 38 Although the rationale for why this occurred is unclear, one could speculate that proteoglycan synthesis is regulated by the amount of proteoglycans accumulated in the pericellular matrix. 38 As the amount of proteoglycans in the pericellular matrix increased, the retention decreased. However, further studies would be required to confirm this. Both IAF and OAF cells accumulated similar amounts of proteoglycans, which was unexpected given that IAF tissue is rich in proteoglycans when compared with the OAF in the mature disc, and gene expression for versican and aggrecan was greater for IAF than for that of OAF cells. There are several possible explanations for this. First, the assay measures all sulfated proteoglycans and cannot differentiate between different types of proteoglycans. Although the IAF is rich in aggrecan, the OAF tissue has other proteoglycans, such as fibromodulin, decorin, biglycan, and versican, particularly in the interlamellar space, in addition to aggrecan.26,39 Second, it may take time for the tissues to show differential accumulation of proteoglycans and 2 weeks of culture may not have been sufficient time for this to occur, as work by Alini et al. has shown that AF cells cultured in collagen-hyaluronan scaffolds increased their chondroitin sulfate content slowly. 40 Third, it is possible that mechanical loading, as present in vivo, may be necessary for differential proteoglycan accumulation as suggested by Korecki et al. 41 Finally, this accumulation may reflect the early stages of AF tissue formation. In the human fetal developing disc, since type I and II collagen are accumulated in the IAF and OAF, respectively, there is little aggrecan in the IVD. 42 As the disc matures, aggrecan is accumulated in the IAF but not in the OAF. Hence, the findings in these cultures may just reflect a defined stage in tissue development.
The maintenance of the differential expression and accumulation of versican by IAF and OAF cells is particularly intriguing. The functional role of versican in the AF has been speculated but is still relatively unknown. Versican, similar to aggrecan, is a large proteoglycan with chondroitin sulfate side chains and has the capability of aggregation and stabilization by link protein. 43 This may contribute to binding water and, thus, confer weight-bearing capability. Alternatively, versican has also been shown to regulate cell adhesion, cell proliferation, and the binding of chemokines and growth factors.44,45 It may play a role in early matrix organization, sequestering growth factors, such as TGF-β, similar to that observed for articular cartilage.46–48 Future work with this culture system will enable investigation of the role of versican in AF development and its role in disc function.
In conclusion, a PU scaffold containing an ADO promotes the maintenance of AF disc cell phenotype, as examined to date, with the IAF and OAF cells accumulating different ECMs. IAF cells accumulate more versican and type II collagen than OAF cells, similar to the native disc. This study suggests that there are fundamental differences between IAF and OAF cells and, thus, raises the possibility that maintaining these features, perhaps through appropriate scaffold selection, may be critical for bioengineering a functional and phenotypically correct IVD. Furthermore, this experimental system can serve as an in vitro model of OAF and IAF tissues that can be used to investigate tissue development and the effect of mechanical loading on these cells.
Footnotes
Acknowledgments
This research was supported by CIHR (M0P114991). The authors would like to thank Harry Bojarski and Ryding-Regency Meat Packer for providing bovine tissues, Dr. Meilin Yang for synthesizing PU scaffolds, and Dr. Firoz Rahemtulla for providing the anti-versican antibody.
Disclosure Statement
No competing financial interests exist.