
Abstract
Autologous tissue-engineered blood vessels (TEBVs) generated using adult stem cells have shown promising results, but many preclinical evaluations do not test the efficacy of stem cells from patient populations likely to need therapy (i.e., elderly and diabetic humans). Two critical functions of these cells will be (i) secreting factors that induce the migration of host cells into the graft and (ii) differentiating into functional vascular cells themselves. The purpose of this study was to analyze whether adipose-derived mesenchymal stem cells (AD-MSCs) sourced from diabetic and elderly patients have a reduced ability to promote human smooth muscle cell (SMC) migration and differentiation potential toward SMCs, two important processes in stem cell-based tissue engineering of vascular grafts. SMC monolayers were disrupted in vitro by a scratch wound and were induced to close the wound by exposure to media conditioned by AD-MSCs from healthy, elderly, and diabetic patients. Media conditioned by AD-MSCs from healthy patients promoted the migration of SMCs and did so in a dose-dependent manner; heating the media to 56°C eliminated the media's potency. AD-MSCs from diabetic and elderly patients had a decreased ability to differentiate into SMCs under angiotensin II stimulation; however, only AD-MSCs from elderly donors were unable to promote SMC migration. Gender and body–mass index of the patients showed no effect on either critical function of AD-MSCs. In conclusion, AD-MSCs from elderly patients may not be suitable for autologous TEBVs due to inadequate promotion of SMC migration and differentiation.
Introduction
U
Despite the success of autologous vascular tissue engineering in reducing intimal hyperplasia and thrombosis, 8 translatability to the clinic has been limited by improper testing of cell sources. Many preclinical investigations do not test the efficacy of cells from clinically realistic patient populations who would routinely see this type of therapy, opting for healthy human or animal cells instead.18–22,24,25,33–63 In addition, many of these healthy human cells are purchased from companies as opposed to being taken from an array of individuals, representative of a realistic patient population. While taken together, these studies have shown that cellularizing tissue-engineered grafts significantly improves the patency and regeneration over their acellular counterparts; these models hold limited relevance for clinical translation since in practice, cells would need to be harvested from patients at high cardiovascular risk such as diabetics64,65 and the elderly.66–68 In addition, these high-risk groups possess pathologic conditions such as a hyperglycemic environment69–72 and age-associated senescence,73–85 respectively, which can decrease the ability of their stem cells to proliferate, differentiate into specific mesenchymal cell types, and promote angiogenesis. Whereas bone marrow has been shown to be a source of stem cells, it is particularly important to consider the use of adipose-derived mesenchymal stem cells (AD-MSCs) due to their ease of isolation and abundance. Whereas there have been studies noting various changes in AD-MSC properties with aging74–76,78,79,82,83,85 and diabetes, 71 it is unclear how donor demographics affect functions related to vascular engineering such as the ability to secrete factors to induce SMC migration and their potential to differentiate into SMCs. Also, investigating if donor demographics alter the factors secreted by AD-MSCs will have a broad impact as the AD-MSC secretome has been used for a variety of applications.82,85–89
In this study, we use human AD-MSCs to test the hypothesis that cells sourced from high-risk populations (i.e., diabetic, elderly) will have a decreased efficacy to promote the migration of SMCs and ability to differentiate into SMCs themselves. We test this in vitro using a scratch wound assay and angiotensin II (AngII)-induced differentiation, respectively. Understanding the effect of donor demographics on the ability of human MSCs to produce SMC promigratory factors and differentiate into SMC is critical to the design of functional TEBVs and could have a wide impact of the stem cell therapy field.
Materials and Methods
Conditioned media from AD-MSCs
AD-MSCs were harvested from the adipose tissue of human patients using previously described methods.76,90 Only information on patient age, gender, body–mass index (BMI), and diabetic status was linked to the harvested cells to protect patient confidentiality, in accordance with an approved Institutional Review Board exempt protocol. Briefly, adipose tissue was isolated from patients, which was minced and digested in a collagenase solution (1 mg/mL, type II) for 30 min. The solution was then filtered through a gauze, centrifuged, resuspended in an NH4Cl erythrocyte lysis buffer (154 mM), and centrifuged again to obtain a cell pellet. The cell pellet was plated and cultured to obtain AD-MSCs. AD-MSCs were then classified into groups that represented a healthy status (<45 years of age, nondiabetic), a diabetic status (<45 years of age, diabetic), or an elderly status (>60 years of age, nondiabetic). To avoid confounding variables when comparing between groups, the age or diabetic condition was held constant and only female donors were used. However, to compare on the basis of gender, cells from different healthy male donors were also analyzed.
All AD-MSCs were cultured in 75-cm2 tissue culture flasks (Corning) with defined culture media [1:1 Dulbecco's modified Eagle's medium (DMEM, #11965; Gibco) to DMEM/F12 (#113300; Gibco) with 10% fetal bovine serum (#S11550; Atlanta Biologics), antibiotics (1% Pen/Strep, 0.5% Fungizone, 0.1% Gentamycin), and 10 μL dexamethasone] mixed with 25% Preadipocyte Growth Medium (#C-27410, #C-39425; PromoCell). To obtain conditioned media, culture media were replenished when flasks were near confluence and termed conditioned after 2 days when collected. Upon collection, the conditioned media were centrifuged to remove any cells or fragments and stored at−80°C until use. For most experiments, conditioned media were diluted with culture media to 100,000 conditioning cells/mL; however, for dose dependence studies, media were diluted from 500,000 conditioning cells/mL (dose 1:0) at dilutions of 1:2, 1:4, 1:9, and 1:19. To heat inactivate, the conditioned media were raised to a temperature of 56°C for 30 min. For all experiments, AD-MSCs between passage number 2 and 6 were used.
Culture of SMCs
Human SMCs were purchased from ATCC (#PCS-100-012) and grown in 75-cm2 culture flasks with SMC Growth Media (#311-500, #311-GS; Cell Applications). SMCs were removed from culture flasks with 0.25% trypsin-EDTA (#25200; Gibco) and placed in 24-well plates (TPP) at 10,000 cells/cm2 to achieve confluent monolayers 1 day before experimentation.
Scratch wound assay
Confluent layers of SMCs within each well had their culture media removed and were scratched with a single stroke of a 200-μL pipette tip to make a wound [scratch wound length: 682±105 μm (avg±SD, n=48)]. SMCs were then washed in a 1× Hanks' balanced salt solution to remove any cellular debris. SMCs were then incubated with SMC media containing 1 μL/mL of Cell Tracker Red (#C34552; Invitrogen) for 30 min to load cells for fluorescent visualization. The media were then replaced with AD-MSC conditioned media and SMCs were then allowed to migrate over 24 h (experimental schematic: Fig. 1). Nonconditioned AD-MSC media were used as a control to show the effect of AD-MSC secreted factors in the conditioned media to promote migration.
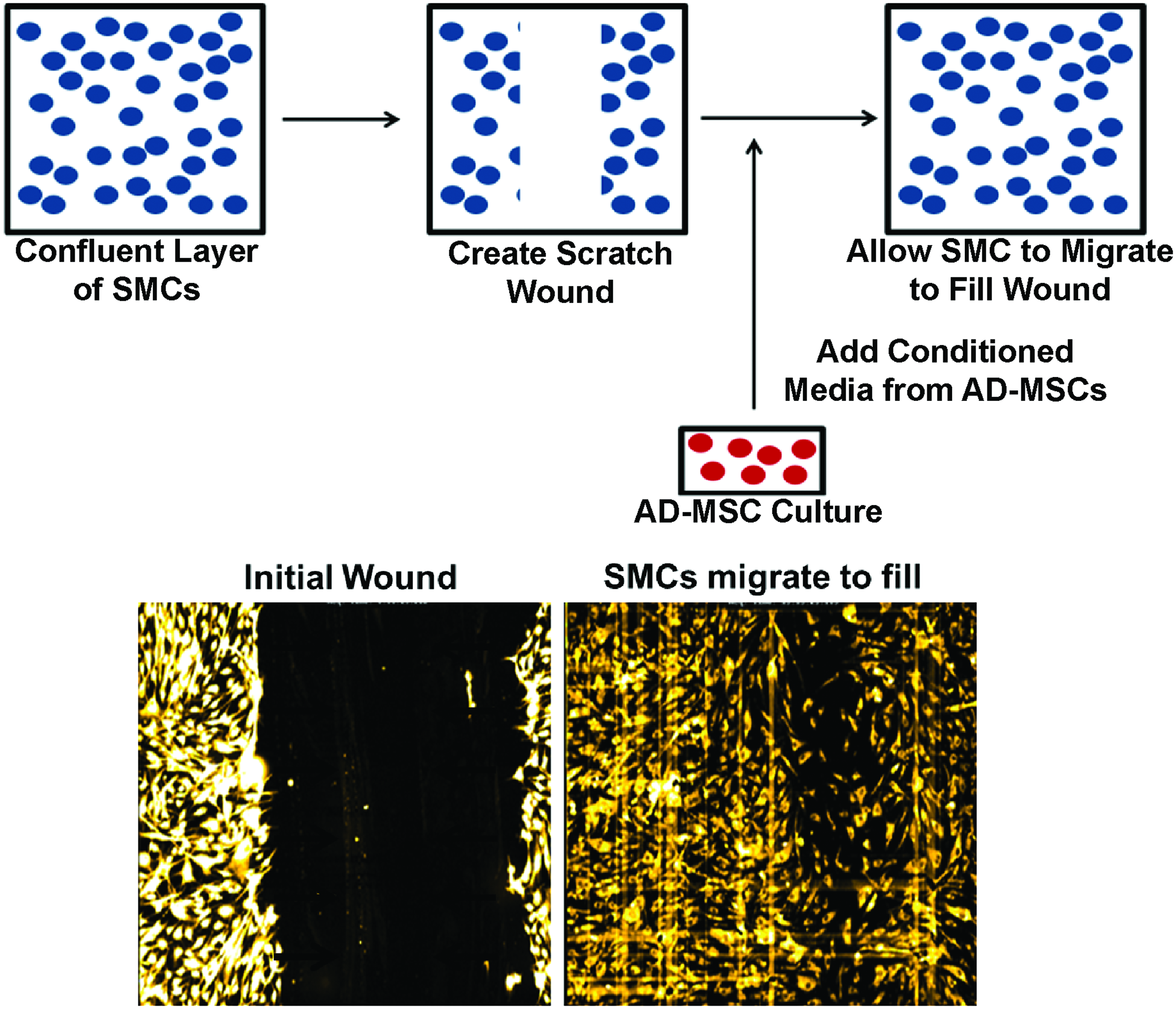
Scratch wound assay. Smooth muscle cells (SMCs) are induced to migrate through a scratch wound assay while being stimulated with adipose-derived mesenchymal stem cell (AD-MSC) conditioned media over the course of 24 h. Top: Schematic of migration assay. Bottom: Images of fluorescently labeled SMCs directly after scratch wound (left), and after migrating into wound area (right). Color images available online at www.liebertpub.com/tea
Cells were placed in a closed thermo-controlled (37°C) stage-top incubator (Tokai Hit Co.) atop the motorized stage of an inverted Nikon TiE fluorescent microscope (Nikon, Inc.) equipped with a 10×, 0.5NA plan apochromat lens (Nikon, Inc.). Cell Tracker Red was excited using a Lumencor diode-pumped light engine (SpectraX, Lumencor, Inc.) and detected using a DsRed longpass filter set (Chroma Technology Corp.) and ORCA-Flash4.0 sCMOS camera (Hamamatsu Corporation). NIS Elements software (version 4.0) was used to automatically image cell migration every 2 h while typical cell culture conditions were maintained (20% O2, 5% CO2, and 37°C). The resulting images were analyzed by measuring the area of the wound normalized to the area at the initial time point for each well. The migration rate was calculated by averaging the difference in normalized area between the first four time points (equivalent to 8 h) and was normalized to nonconditioned controls. In addition, images of healthy, diabetic, elderly, and control conditions were acquired every 15 min and joined sequentially as a time-lapse video to show migration of each side-by-side (Supplementary Video SV1; Supplementary Data are available online at www.liebertpub.com/tea).
ELISA on AD-MSC conditioned media
An enzyme-linked immunosorbent assay (ELISA) kit was utilized to detect the levels of vascular endothelial growth factor (VEGF) within the conditioned media of AD-MSC donors (#DVE00; R&D Systems). All protocols were followed according to the manufacturer's instructions. To get a greater sense of the population variability of VEGF present within conditioned media, a higher number of donors (n=12) were utilized for this assay. All donors investigated remained in the healthy AD-MSC group (i.e., <45 years of age, nondiabetic) and spanned a wide range of BMI (avg±SD: 27.3±4.3, range: 21.3–34.7).
AD-MSC differentiation into SMCs
To induce differentiation of AD-MSCs into SMCs, AD-MSCs were plated on PLL-coated coverslips (GG-22-pll; Neuvitro) at a density of 10,000 cells per coverslip. They were cultured in media (MEMα, #12561-056; Life Technologies) containing AngII (1 μM, #A9525; Sigma) with 10% serum. 91 After 4 days of culture, AD-MSCs were fixed in 4% paraformaldehyde and evaluated using a standard immunofluorescent chemistry protocol to detect calponin (1:250, #ab46794; Abcam), myosin heavy chain (1:250, #ab77967; Abcam), and smoothelin (1:250, #ab8969), to quantify percent expressing cells. F-actin was fluorescently labeled with FITC-phalloidin (1:250, #P5282; Sigma) to assess the cell shape (approaching a spindle-like SMC morphology). Calponin is expressed in aortic SMCs across their phenotype diversity, whereas myosin heavy chain and smoothelin are selective for mature contractile SMCs.92–94 All staining intensities were quantified using NIS Elements software. Primary human aortic SMCs (#PCS-100-102; ATCC) were used as positive controls for SMC staining.
Statistical analyses
All statistical analyses were done utilizing Minitab software (version 16) to perform a t-test, ANOVA, or linear regression. Statistical significance was accepted at p<0.05.
Results
AD-MSC secreted factors can promote SMC migration
To investigate the ability of AD-MSCs to exert a paracrine promigratory effect on SMCs, we harvested human AD-MSCs from a variety of patients to obtain their conditioned media and employed a scratch wound assay with a confluent monolayer of human SMCs. Upon stimulation with AD-MSC conditioned media from healthy donors, SMCs showed an increased wound closure rate when compared with nonconditioned AD-MSC media (Fig. 2A and Supplementary Video SV1). These data were also expressed as the migration rate per hour over the first 8 h, and conditioned media displayed a significant increase in rate over nonconditioned controls (Fig. 2B). In addition, the factors promoting SMC migration were heat labile, as heating of conditioned media to 56°C removed the promigratory effect (Fig. 2B). Upon dilution of AD-MSC conditioned media to varying levels of conditioning cells/mL, a dose-dependent effect was observed (Fig. 2C). A dose of 1:19 (20-fold dilution) showed a similar effect to nonconditioned media.

AD-MSC secreted factors can promote SMC migration. SMCs are induced to migrate through a scratch wound assay while being stimulated with AD-MSC conditioned media over the course of 24 h. AD-MSCs promote the migration of SMCs compared to nonconditioned controls
To identify one factor that could be responsible for this paracrine effect on migration, we quantified VEGF levels in the conditioned media. VEGF was shown to be present at 4091±1903 pg/106 AD-MSCs.
AD-MSCs from diabetic donors can promote the migration of SMCs but have a decreased ability to differentiate into SMCs
Determining if AD-MSCs from clinically realistic patient groups can perform two main functions utilized in vascular tissue engineering—inducing SMC migration and differentiating directly into SMCs—is critical to the development of a stem cell-based vascular graft. To first assess if AD-MSCs from diabetic patients (i.e., a cohort of patients at high cardiovascular risk) would have a reduced ability to produce SMC promigratory secreted factors, we stimulated SMCs with conditioned media from the AD-MSCs of this patient population. Media conditioned by diabetic AD-MSCs induced faster wound closure than nonconditioned media (Fig. 3A, B and Supplementary Video SV1) in a similar manner to healthy AD-MSCs. This shows that a diabetic origin does not affect the functionality of these cells. Also, it is noteworthy that all AD-MSCs (healthy, diabetic, and elderly) were cultured in high glucose growth media (∼17 mM that is equivalent to 306 mg/dL, which is above the diabetic threshold), which suggests that the diabetic hyperglycemic environment does not limit the ability of AD-MSCs to produce SMC promigratory factors.

AD-MSCs from diabetic patients produce SMC promigratory secreted factors. Investigating diabetic patient AD-MSCs for the ability of their secreted factors to promote SMC migration, showed an increased migration compared to nonconditioned media [
To assess if AD-MSCs from diabetic patients had a reduced ability to differentiate into SMCs, they were stimulated with AngII. Diabetic cells displayed a less efficacious differentiation compared to controls indicated by a significantly lower expression of the SMC marker calponin (Fig. 4A, B) and a failure to adopt an SMC spindle-like morphology (Fig. 4C, D). Unstimulated AD-MSCs do not express calponin (data not shown) and maintain a flat morphology (Fig. 4C, D), whereas SMCs ubiquitously express calponin and a spindle-like morphology (Fig. 4E). In addition, no positive staining was seen with myosin heavy chain or smoothelin with healthy or diabetic donor AD-MSCs in either stimulated or unstimulated states, although SMCs stained positively in the same assay (data not shown).
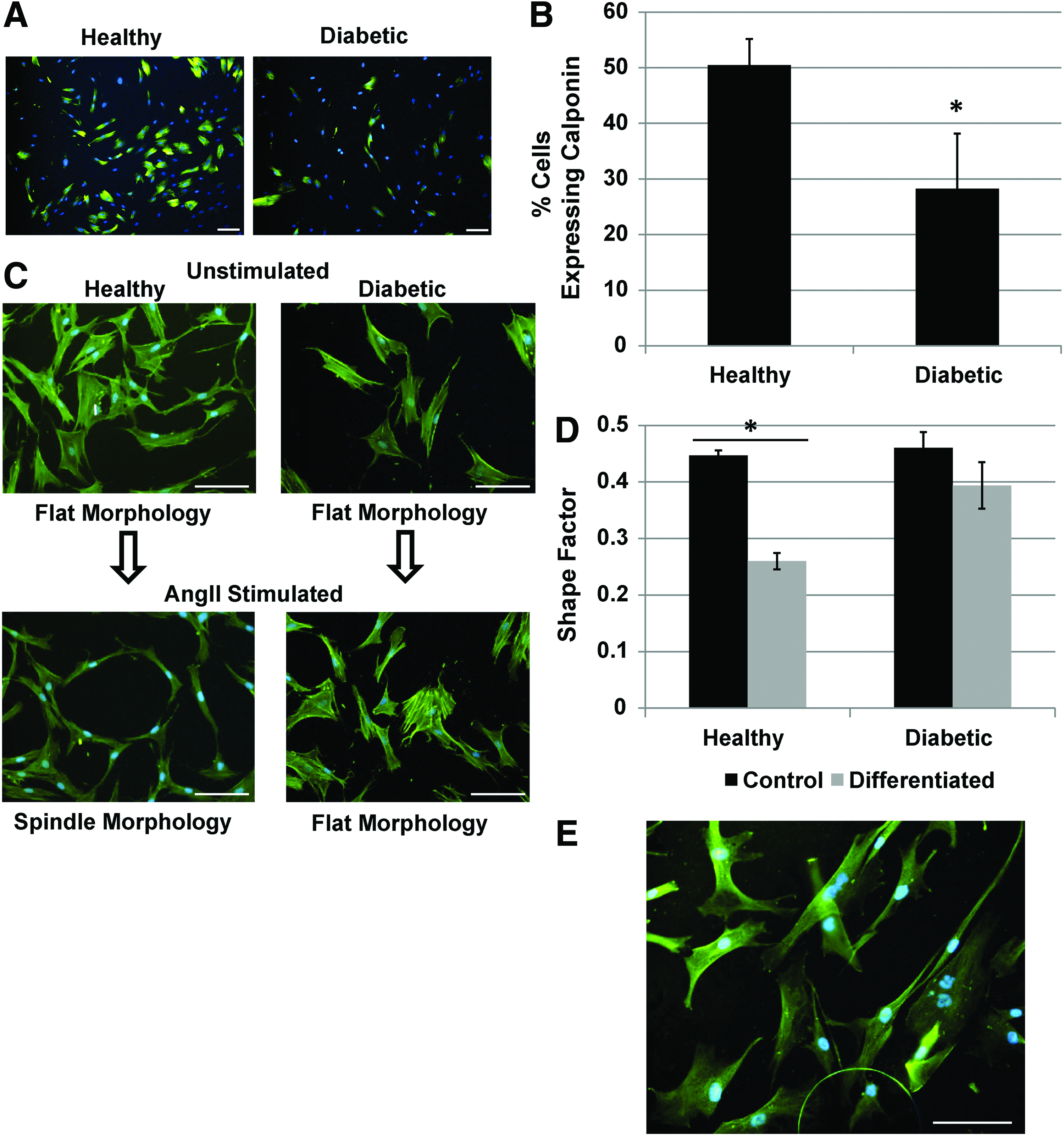
AD-MSC differentiation into SMC is decreased for diabetic patients. Diabetic AD-MSCs displayed a less efficacious SMC differentiation compared to healthy AD-MSCs in terms of the expression of calponin [
AD-MSCs from elderly donors have both a decreased ability to promote the migration of SMCs and differentiate into SMCs
Utilizing a scratch wound assay, AD-MSC secreted factors from elderly donors (i.e., another cohort of patients at high cardiovascular risk) were unable to promote the migration of SMCs (Fig. 5A, B and Supplementary Video SV1). The differentiation potential of elderly donor AD-MSCs toward SMCs was also found to be reduced based on the expression of calponin (Fig. 6A, B) and maintainance of a flat morphology (Fig. 6C, D) under AngII stimulation. AD-MSCs from elderly donors did not stain positively for myosin heavy chain or smoothelin in either stimulated or unstimulated states (data not shown).
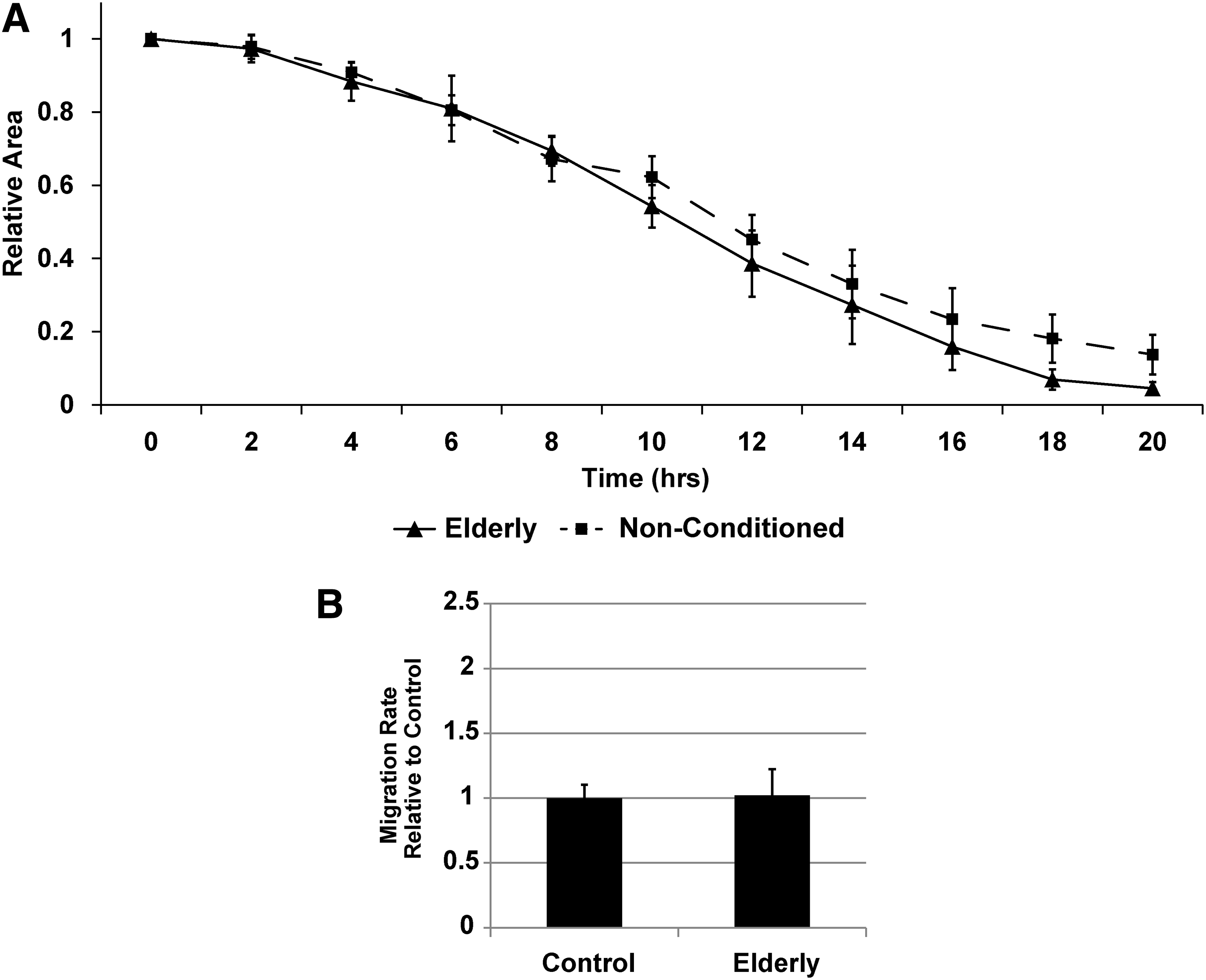
AD-MSCs from elderly patients do not produce SMC promigratory secreted factors. When comparing between healthy and elderly patient AD-MSCs, those from elderly patients displayed an inability of their secreted factors to produce SMC promigratory effects [

AD-MSC differentiation into SMC is decreased for elderly patients. Elderly AD-MSCs displayed a reduced ability to differentiate into SMCs under angiotensin II (AngII) stimulation for 4 days. AngII-stimulated AD-MSCs showed a significantly lower expression of calponin [
Gender and BMI do not affect the ability of AD-MSCs to promote the migration of SMCs or the ability to differentiate into SMCs
As all the donors utilized for the scratch wound assay were females, male donors from the healthy demographic were acquired and compared directly to the healthy female group to assess potential gender differences. The ability to produce SMC promigratory factors did not appear to be gender specific (Fig. 7A), as there was no difference in the migration rate observed between the groups. In addition, the gender did not affect the ability of AD-MSCs to differentiate into SMCs as both male and female donors displayed equivalent results in terms of expression of calponin postdifferentiation (Fig. 7B) and change to a SMC-like morphology in the presence of AngII (Fig. 7C). The BMI was also investigated in both assays by performing a regression analysis on the migration rate (Fig. 7D) and calponin expression (Fig. 7E) postdifferentiation, respectively. No significant trend was seen between either set of data (migration rate: R2=0.05, p=0.4; calponin expression: R2=0.001, p=0.96). Also, we correlated the concentration of VEGF (pg/106 cells) with BMI and showed no significant trend (data not shown, R2=0.01, p=0.74). From these data, altered abilities in AD-MSC function appear to be dependent on diabetes and age, but not gender or BMI.
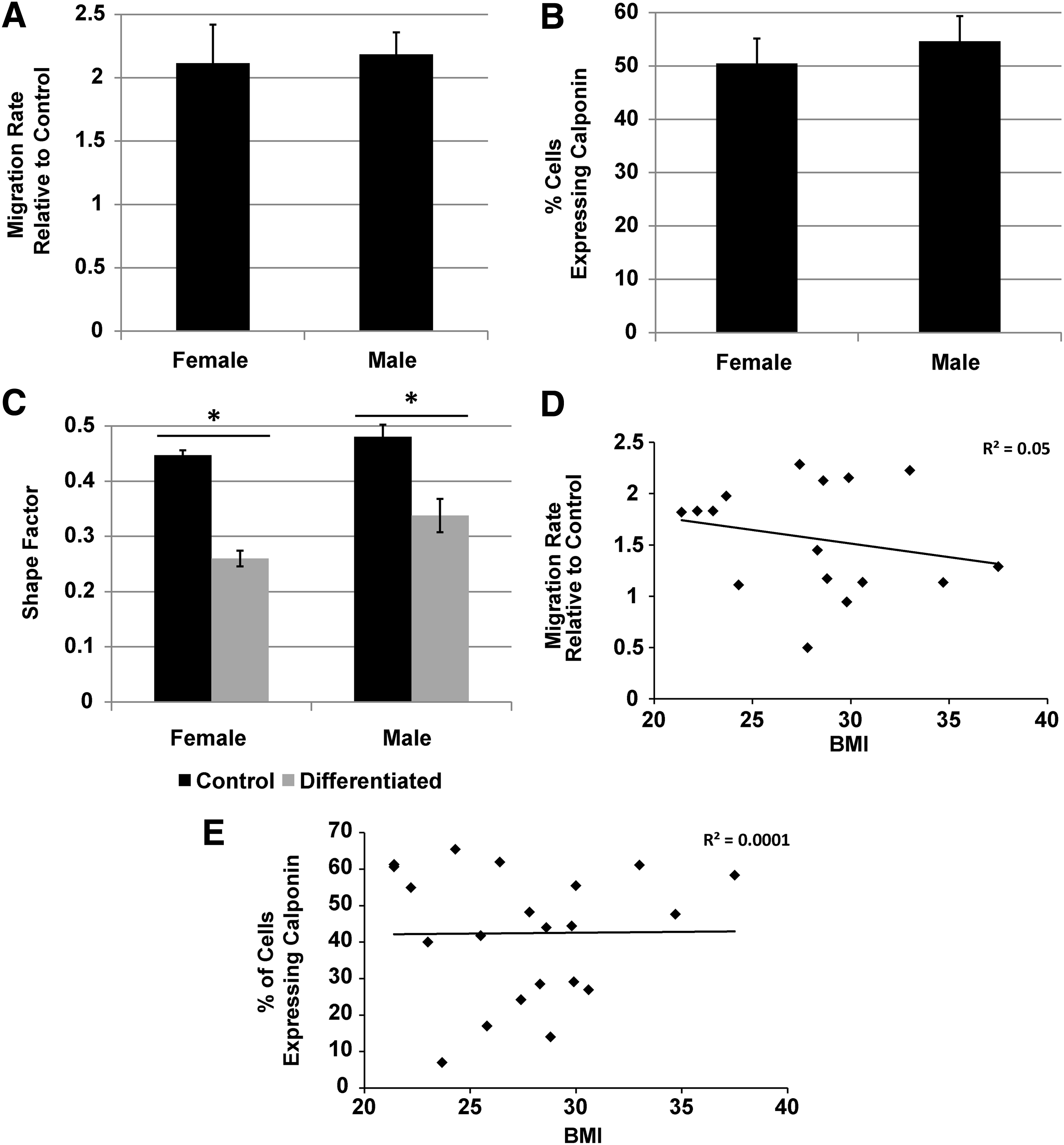
Gender and body–mass index (BMI) do not affect AD-MSC secreted SMC promigratory factors or AD-MSC differentiation into SMCs. Comparing gender of AD-MSCs from both healthy male and female donors produced equivalent SMC promigratory responses
Discussion
The ability of an implanted scaffold to become populated with functional cellular constituents is a critical factor for success in tissue engineering applications. For example, not only does a TEBV with more SMCs indicate a higher degree of vascular maturity, 50 including medial cellularity, myosin heavy chain expression, and contractility, but SMC content also correlates with desirable mechanical properties such as tensile strength and stiffness.39,41 These mechanical properties are critical for the long-term success of a TEBV as they can be predictive of failure by aneurysmal dilation/rupture and intimal hyperplasia. 95 As one of the main design strategies in tissue engineering is to utilize a biodegradable scaffold that is replaced with host tissue, if sufficient vascular cells and matrices are not recruited and produced to compensate the mechanical loss due to scaffold degradation, the TEBV could fail in the long term. Including MSCs that can guide this process is critical to avoiding failure.
Many recent studies have begun to investigate the mechanisms by which TEBVs remodel to achieve a mature native-like vessel. Studies utilizing cell tracking have shown that seeded cells are eventually replaced with migrating host SMCs, with the seeded cells being necessary for providing initial antithrombogenicty and initiating the remodeling process.18–22 Providing further evidence for this phemonmenon, implanted scaffolds with low porosity28,30,32 and even TEBVs with already intact SMC layers 24 have been shown to be repopulated with host vessel wall cells. In addition, vascular engineering methods have begun to take advantage of host recellularization by creating acellular approaches, utilizing bound growth factors or drug delivery methods,19,23,96 or designing porous scaffolds to facilitate this cellular infiltration.31,46,97,98 In all of these cases, establishing a mature TEBV populated by SMCs requires recruitment of host cells.
In this study, we show that donor demographics in human AD-MSCs can play a role in their ability to produce secreted factors that are responsible for SMC migration and for them to differentiate into SMCs, both of which are important for remodeling and maturation of TEBVs. Age is critically important to both the abilities of AD-MSCs to produce secreted factors and differentiate into SMCs, whereas a diabetic condition only affected the ability of AD-MSCs to differentiate. However, gender and BMI did not seem to play a role in either stem cell function, which is surprising since female AD-MSCs proliferate at a higher rate than male ones99,100 and BMI is negatively correlated with the AD-MSC differentiation capacity. 101
One area of interest that was not yet investigated in the migration experiments is the effects of each AD-MSC demographic on their respective SMC demographic, such as the effect of diabetic AD-MSC conditioned media on diabetic SMCs. However, SMCs sourced from diabetic and elderly humans have been previously shown to have increased 102 and decreased 103 migration, respectively. This parallels the above conclusions that the combination of AD-MSC and SMC of diabetic patients is promigratory, whereas the combination of the elderly is not.
As we have utilized human cells in this study, particularly those from clinically realistic groups, and shown functional differences in a mechanism of action by which TEBVs remodel and the ability of AD-MSCs to form SMCs, this represents a critical step in the clinical translation of TEBVs. We have shown that elderly AD-MSCs may cause poor TEBV remodeling in terms of their ability to recruit SMCs. While these cells may have auxiliary functions such as providing resistance against thrombosis, 20 this clearly could reduce the effectiveness of an autologous TEBV therapy. This an important concern as more than 4 million people in the United States are over 65 years old 104 and these represent one of the highest cardiovascular risk groups. 67 In addition, as many TEBV approaches utilize AD-MSCs to differentiate into SMCs,14–17 those utilizing elderly and diabetic cells could be heavily affected due to the reduced differentiation potential for both groups. One point worth noting in this regard is that with 1 μM AngII stimulation for 4 days, AD-MSCs do not achieve complete differentation into SMCs, as evidenced by the absence of staining for myosin heavy chain or smoothelin (both late differentiation markers).
We have also shown that the ability of AD-MSCs to induce SMC migration is decreased upon heat inactivation indicating that these secreted factors likely operate on a protein level. One likely explanation could be that growth factors secreted by MSCs are responsible for this promigratory function. Indeed, we have shown that one known factor to promote SMC migration, VEGF, is present in our conditioned media. However, many growth factors produced by AD-MSCs are promigratory for SMCs 105 and can also induce other mitogenic, proteolytic, extracellular matrix producing, inflammatory, and angiogenic effects. 5 This is concerning for studies that target elderly patients, as the same growth factors that lead to the deficiency in promoting SMC migration may also affect other applications beyond TEBVs. Also, with AD-MSC secreted factors showing a dose-dependent response, the need to optimize the number of cells utilized in cell therapy approaches is clear.
In conclusion, by utilizing conditioned media from AD-MSCs to induce SMC migration, we have shown that age and not diabetes, gender, or BMI results in an inability. In addition, this effect is dose dependent and functions on a protein level. Also, we have shown that differentiation of AD-MSCs from diabetic and elderly patients is decreased but no effect was seen due to gender or BMI. With respect to TEBVs, AD-MSCs from elderly patients may be suboptimal in autologous TEBVs, as they lack the ability to induce host recellularization.
Footnotes
Acknowledgments
This work was supported by the American Heart Association (AHA #12PRE12050163 to J.T.K.) and the National Institutes of Health (R21 #EB016138 to D.A.V.).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.