
Abstract
Current approaches in bone tissue engineering have shown limited success, mostly owing to insufficient vascularization of the construct. A common approach consists of co-culture of endothelial cells and osteoblastic cells. This strategy uses cells from different sources and differentiation states, thus increasing the complexity upstream of a clinical application. The source of reparative cells is paramount for the success of bone tissue engineering applications. In this context, stem cells obtained from human bone marrow hold much promise. Here, we analyzed the potential of human whole bone marrow cells directly expanded in a three-dimensional (3D) polymer matrix and focused on the further characterization of this heterogeneous population and on their ability to promote angiogenesis and osteogenesis, both in vitro and in vivo, in a subcutaneous model. Cellular aggregates were formed within 24 h and over the 12-day culture period expressed endothelial and bone-specific markers and a specific junctional protein. Ectopic implantation of the tissue-engineered constructs revealed osteoid tissue and vessel formation both at the periphery and within the implant. This work sheds light on the potential clinical use of human whole bone marrow for bone regeneration strategies, focusing on a simplified approach to develop a direct 3D culture without two-dimensional isolation or expansion.
Introduction
I
A large number of co-culture studies using two different lineages from different sources and including differentiated cells mixed with undifferentiated cells (e.g., MSCs or progenitors) have been reported. Among these, human umbilical vein ECs, 25 human outgrowth ECs, 26 human microcapillary ECs, 27 and endothelial progenitor cells from cord blood have been studied for co-culture with human bone-derived cells, 28 osteoblast-like MG63 cells, 29 primary human osteoblast cells, 30 human osteoprogenitor cells from bone marrow, 31 and MSCs from adipose tissue 32 and bone marrow. 33 However, the co-culture of these two different lineages from different sources involves limitations with a view to clinical applications.
In this context, human bone marrow is promising, as it contains both MSCs and microvascular ECs that are able to support both osteogenesis and angiogenesis, respectively. Indeed, this approach has already been reported and although bone healing was improved, the hypothesized increase in vascularization and the impact on MSC differentiation toward the osteoblastic lineage was not determined. 34 In a recent clinical trial, it was also shown that the direct injection of concentrated bone marrow cells could successfully accomplish the regeneration of nonunion fractures, 35 supporting the potential of this cell source.
Several strategies have been used to achieve the local delivery of stem to in the injured site in order to promote tissue regeneration. To address this issue and to mimic a relevant 3D microenvironment, scaffolds are needed to create physicochemical macro- and microstructures that can promote cellular interplay and differentiation. A wide range of scaffolds have been used for bone tissue engineering, including inorganic, calcium-based phosphate matrices and polymeric structures. The latter can be considered suitable, as they can mimic the aqueous in vivo environment. 36 Moreover, the macroporous interconnectivity provided by the polymeric scaffold favors the migration of cells within the material and the invasion/integration of the host cells. 37 Owing to their composition and structure, they have inherent design flexibility and can, thus, be tailored to a specific need 38 such as the promotion of angiogenic events. The use of polysaccharides such as alginate, chitosan, pullulan, or cellulose39–44 has been extensively reported and numerous studies are in progress that focus on bone tissue engineering. These natural polymers hold promise as matrices for tissue regeneration strategies, as they are highly permeable, facilitating the transport of nutrients and metabolites. 45 Here, we focused on a macroporous matrix composed of the naturally occurring hydrophilic polysaccharides pullulan and dextran. These biodegradable matrices have already been shown suitable for cell therapy,46,47 as attested by their capability to sustain vascular cell growth 48 and the culture of MSCs derived from adipose tissue. 49 In addition, these matrices have recently been shown to favor the co-culture of human MSCs and human EPCs, promoting cell communication and improved bone formation. 50
Here, we investigated the potential of human whole bone marrow cells (hWBMCs) as a unique source of reparative cells for regenerating bone tissue and vasculature directly within polysaccharide-based macroporous matrices, without preliminary selection or two-dimensional (2D) expansion. We characterized (1) the population formed by the multicellular components of hWBMCs in 2D assays and thereafter in 3D matrices; (2) its contribution to promote bone and EC specific markers in 3D culture; and (3) the fate of the tissue-engineered constructs after ectopic implantation in a NOG mice model. Taken together, our findings provide new insights into the potential of this heterogeneous cell source within a 3D macroporous matrix for bone regeneration strategies that involve a limited ex-vivo culture and cell manipulation. This might help such strategies to move closer to the bedside.
Materials and Methods
Preparation of 3D macroporous scaffolds
Macroporous scaffolds (6 mm diameter, 2 mm thickness, 20 mm3) were produced using a blend of pullulan/dextran 75:25 (pullulan, MW 200,000, Hayashibara, Inc.; Dextran, MW 500,000, Sigma) prepared by dissolving 9 g of pullulan and 3 g of dextran into 40 mL of distilled water containing 14 g of NaCl, as previously described. 51 Chemical cross-linking was carried out using trisodium trimetaphosphate. Pores were created by a patented gas-foaming technique. Resulting scaffolds were cut into the desired shape, soaked in phosphate-buffered saline (PBS), and then extensively washed with a 0.025% NaCl solution. After freeze drying, scaffolds were stored at room temperature until further use. Regarding their characterization, pore size and pore area were determined using Environmental Scanning Electron Microscopy and confocal microscopy (FITC-Labeled dextran scaffolds). Water content and swelling ratio were determined as previously described. 52 Scaffolds with 6 mm diameter, 2 mm thickness, 20 mm3, porosity 68%±3%, and pore size 243±14 μm were used in this work.
Isolation and culture of whole bone marrow cell population
hWBMCs were isolated from human bone marrow samples as previously described. 53 Briefly, bone marrow was aspirated from the femoral diaphysis or iliac bone after obtaining consent from patients undergoing hip prosthesis surgery after trauma. Here, in the totality of this study, 10 different samples of hWBMCs from different donors were used, with an average donor age of 66±11 years. For each independent experiment, a different donor was used. The study was approved by the local institutional review board. Cells were separated by sequential passages through syringes fitted with 16-, 18-, or 21-gauge needles. After centrifugation for 15 min at 800 g, the pellet was resuspended in the following 50/50 culture medium containing 50% (v/v) of α-minimal essential medium (α-MEM; Invitrogen) supplemented with 10% (v/v) fetal calf serum (FCS; Gibco) and 50% (v/v) EC growth medium-2 (EGM-2; Lonza) and 5% (v/v) FCS before flow cytometry analysis or cell seeding within the scaffolds.
Flow cytometry
hWBMCs were isolated as previously described, and red blood cells were lysed using FACS lysing solution (BD Biosciences). Next, hWBMCs were washed with PBS 1× pH 7, and incubated with coupled antibodies against CD73 (cat#561254), CD34 (cat#555824), CD271 (cat#563451), CD105 (cat#561443) (markers for mesenchymal cells; BD Biosciences) and CD31 (cat#55445), CD34 (cat#555824), CD146 (cat#560846), CD29 (cat#563513) (markers for ECs; BD Biosciences) during 1 h at room temperature. 100,000 events were acquired and analyzed using a BD FACS Canto flow cytometer (BD Biosciences).
Immunolabeling of microvascular ECs in hWBMCs in 2D culture
hWBMCs cultured on plastic culture plates during 5 days without medium change were fixed with 4% (w/v) paraformaldehyde (PFA) at 4°C for 30 min and permeabilized with Triton X 100 at 0.1% (v/v) for 30 min at 4°C. Then, samples were blocked during 1 h in PBS 1× pH 7.4 (Gibco) containing 1% (w/v) bovine serum albumin (BSA) and then incubated with mouse anti-human VE-cadherin antibody (cat#sc-9989; Santa Cruz Biotechnology), or mouse anti-human CD31 antibody (cat#sc-13537; Santa Cruz Biotechnology), or rabbit anti-human Von Willebrand factor (vWF) antibody (cat#A0082; Dako) overnight at 4°C. Subsequently, cells were washed in PBS 1× pH 7.4 (Gibco) and incubated with Alexa fluor 568-conjugated rabbit anti-mouse IgG (cat#A11061; Molecular Probes) or Alexa fluor 488-conjugated goat anti-rabbit IgG (cat#A11008; Molecular Probes) for 2 h at 37°C. Cells were incubated with the nuclear probe 4′, 6′-diamidino-2-phenylindole (DAPI; Invitrogen) for 30 min at room temperature to label the nucleus and were thereafter observed with a fluorescence microscope (Nikon Eclipse 80i) equipped with a digital camera (Nikon Dxm 1200C).
Cell seeding within a 3D macroporous scaffold
Dried scaffolds (6 mm diameter, 2 mm thickness) were submitted to UV radiation for 30 min before cell seeding. For culture experiments, hWBMCs were cultured for 1, 6, and 12 days in the 50/50 culture medium (α-MEM and EGM-2 MV at a ratio 1:1 [v/v]). The cells were seeded at 4×105 cells/disk. Cells seeded into the 3D scaffold were detected after 24 h by calcein staining (cat#L3224; Molecular Probes) and examined with a modular stereo microscope (Leica MZ 10 F; Leica Microsystemes).
Alkaline phosphatase assay
Intracellular alkaline phosphatase (ALP) activity was assessed in cells cultured within scaffolds by using the conversion of a colorless p-nitrophenyl phosphate to a colored p-nitrophenol (Sigma diagnostic kit [85L-2]; Aldrich).
Cryo-sections of tissue-engineered constructs for microscopy
At designated time points, cellularized matrices were rinsed four times with PBS (1× pH 7.4) and then fixed for 30 min with 4% (w/v) PFA at 4°C. The matrices were then included in gel freeze (Labonord SAS). Sections of 10 μm thickness were obtained using a cryostat (Leica CM 1850 UV; Leica Microsystemes).
Immunolabeling of endothelial marker proteins and connexin43 into the 3D scaffold
After 1, 6, and 12 days of culture, the matrices containing hWBMCs were fixed with 4% (w/v) PFA at 4°C for 30 min and permeabilized with Triton X 100 at 0.1% (v/v) for 30 min at 4°C. Then, they were blocked for 1 h in PBS 1× pH 7.4 (Gibco) containing 1% (w/v) BSA before incubation with primary antibodies: mouse anti-human CD31 (cat#sc-13537; Santa Cruz Biotechnology), or rabbit anti-human vWF (cat#A0082; Dako) overnight at 4°C. Subsequently, cells were washed in PBS 1× pH 7.4 (Gibco) and incubated with Alexa fluor 488-conjugated rabbit anti-mouse IgG (cat#A11061; Molecular Probes) for 2 h at 37°C or Alexa fluor 568-conjugated goat anti-rabbit IgG (cat#A11008; Molecular Probes) for 2 h at 37°C. To label the nucleus, cells were incubated with the nuclear probe DAPI (Invitrogen) for 30 min at room temperature and were thereafter analyzed with a fluorescence microscope (Nikon Eclipse 80i) equipped with a digital camera (Nikon Dxm 1200C). For connexin43 (Cx43) labeling, cells were incubated with a primary antibody mouse anti-human against Cx43 (cat#MAB3068; Millipore). Subsequently, they were washed and incubated with Alexa 488-conjugated goat anti-mouse IgG (cat#A11001; Molecular Probes). Cultures were examined with a fluorescence microscope (Leica DMi 3000 B; Leica Microsystemes).
Immunolabeling of bone marker proteins
After 1, 6, and 12 days of culture, the matrices containing hWBMCs were fixed with 4% (w/v) PFA at 4°C for 30 min and permeabilized with Triton X 100 at 0.1% (v/v) for 30 min at 4°C. Then, they were blocked for 1 h in PBS 1× pH 7.4 (Gibco) containing 1% (w/v) BSA before incubation with primary antibodies: mouse anti-human Osteocalcin (OCN, cat#35-5400; Takara), or mouse anti-human COL1A1 (cat#M011; Takara) overnight at 4°C. Subsequently, cells were washed in PBS 1× pH 7.4 (Gibco) and incubated with Alexa fluor 488-conjugated rabbit anti-mouse IgG (cat#A11061; Molecular Probes) for 2 h at 37°C. For nuclear labeling, cells were incubated with the nuclear probe DAPI (Invitrogen) for 30 min at room temperature and were thereafter analyzed using a fluorescence microscope (Nikon Eclipse 80i) equipped with a digital camera (Nikon Dxm 1200C).
Alizarin Red staining
Alizarin Red staining was performed in order to assess the level of calcium deposition induced by the hWBMC culture. After 1, 6, and 12 days of culture, cells were fixed in 4% (w/v) PFA. Next, cells were stained with alizarin red S (2%, pH 4.2; Millipore) for 2 min. After aspiration of the overflow, cells were washed thrice with water. Extracellular matrix staining was observed using an optical microscope (Axiovert 25; Zeiss).
Quantitative real-time polymerase chain reaction
Total RNA was extracted using the RNeasy Total RNA kit (Qiagen), as indicated by the manufacturer, and 1 μg was used as the template for single-strand cDNA synthesis, using the Superscript system (cat#11904-018; Invitrogen). cDNA diluted at a 1:80 ratio was loaded onto a 96-well plate. Real-time polymerase chain reaction (PCR) amplification was performed using the SYBR-Green Supermix (Bio-Rad). Primers of the ubiquitary ribosomic protein P0, ALP, COL1A1, Cbfa1/runx2, OCN, Cx43 as previously described 50 and CD31 and vWF as previously described 54 were used at a final concentration of 200 nM. Data were analyzed using iCycler IQ software and compared by the ΔΔCT method. Quantitative real-time polymerase chain reaction (q-PCR) was performed in triplicate. Results were expressed relative to gene expression levels on day 1. Data were normalized to P0 (ribosomal protein) mRNA expression for each condition and was quantified relative to cbfa1/runx2, ALP, CD31, vWF, OCN, and COL1A1 gene expression in hWBMCs cultured alone in the matrix after 24 h, which was standardized to 1.
Ectopic implantation of tissue-engineered constructs in mice
The procedures and animal handling followed the principles of Laboratory Animal Care formulated by the National Society for Medical Research and were approved by the Animal Care and Experiment Committee of the University of Bordeaux. Experiments were carried out in accredited animal facilities according to European recommendations for laboratory animal care (directive 86/609 CEE of 24/11/86). Scaffolds were seeded with hWBMCs (4×105 cells/scaffold) 1 day before implantation. Then, matrices were implanted in a dorsal subcutaneous site of 12-week-old NOG female mice weighing 25–30 g (from the central animal facility of the University of Bordeaux, Bordeaux, France). For each condition and for each time of implantation, five samples were used for histological analysis. Five mice with two matrices were used for each condition. Animals were euthanized at 3 or 8 weeks. These time points were chosen as early and late time points of ectopic implantation in order to provide data regarding biointegration and osteoinduction. 55
Histological analysis
At 3 and 8 weeks postimplantation, samples were retrieved, fixed in 4% (w/v) PFA for 4 h at 4°C, dehydrated, and embedded in paraffin. Sections (4–5 μm in thickness) were de-paraffinized using toluene, rehydrated in decreasing concentrations of ethanol (100–50%), washed in distilled water, and, finally, stained with Mayer's Hemalum and Erythrosine Masson's trichrome (Invitrogen). The samples were observed with a Nikon Eclipse 80i microscope. The osteoid tissue area on paraffin sections was quantified using the ImageJ software. Two slides per sample with 15 average fields were used for quantification.
Immunohistochemistry
At 3 and 8 weeks postimplantation, samples were retrieved and fixed in 4% (w/v) PFA for 4 h at 4°C, dehydrated, and embedded in paraffin. Sections (4–5 μm in thickness) were de-paraffinized using toluene, rehydrated in decreasing concentrations of ethanol (100–50%), and, finally, washed in distilled water. Antigen recovery was performed in TBS buffer (10 mM Tris, 1 mM EDTA, 0.05% Tween 20, pH 9.0) at 100°C for 20 min. Then, endogenous peroxidase was quenched in 3% H2O2, in PBS, for 5 min. After washing with PBS, slides were blocked using 3% goat serum in PBS for 30 min. For CD31 immunolabeling, incubation was performed overnight with a rabbit polyclonal anti-human/mice CD31 antibody (cat#ab28364; Abcam) at 4°C. After two washes with PBS, the slides were incubated with the components of the anti-rabbit ABC kit (Vector Labs), as indicated by the manufacturer, and then revealed using DAB solution (Impact DAB; Vector Labs). Staining was stopped in distilled water, and samples were then counterstained in Mayer's hematoxylin and washed in running tap water for 10 min. Finally, samples were dehydrated and mounted using Entellan. Cell species identification was achieved by an overnight incubation at 4°C with a rat monoclonal anti-human HLA-1 antibody (cat#SM2012P; Acris). After two washes with PBS, the slides were incubated with the components of the anti-rat ImmPRESS kit (Vector Labs), as indicated by the manufacturer, and then revealed using DAB solution (Impact DAB; Vector Labs). Counterstaining and mounting were performed as previously described.
Statistical analysis
Data are represented as the mean±standard deviation resulting from independent experiments. The GraphPad Prism software (GraphPad, Inc.) was used for statistical analysis. For the q-PCR analysis, the two modes of analysis of variance were performed in order to compare the mean values between groups using the post-hoc Bonferroni test. For the histological analysis, the t test was used to compare the mean values between groups. Differences were considered significant when *p<0.05, **p<0.01, or ***p<0.001.
Results
Characterization of freshly isolated hWBMCs and 2D in vitro expansion: evidence for microvascular ECs
hWBMC characterization in terms of endothelial and mesenchymal cell-specific markers was performed to further establish the components of this heterogeneous population. Six independent samples of freshly isolated hWBMCs were processed for immunostaining and analyzed by flow cytometry. FACS analysis showed that 9.5%±1.0% of hWBMCs were positive for CD73 and negative for CD34, 10.0%±0.5% for CD105, and 12.0%±1.5% for CD271 (Fig. 1A). In addition, 6.3%±0.2% of the population was double positive for CD31/CD34 and 5.0%±1.5% positive for CD146 and 8.0%±1.5% for CD29 (Fig. 1B).
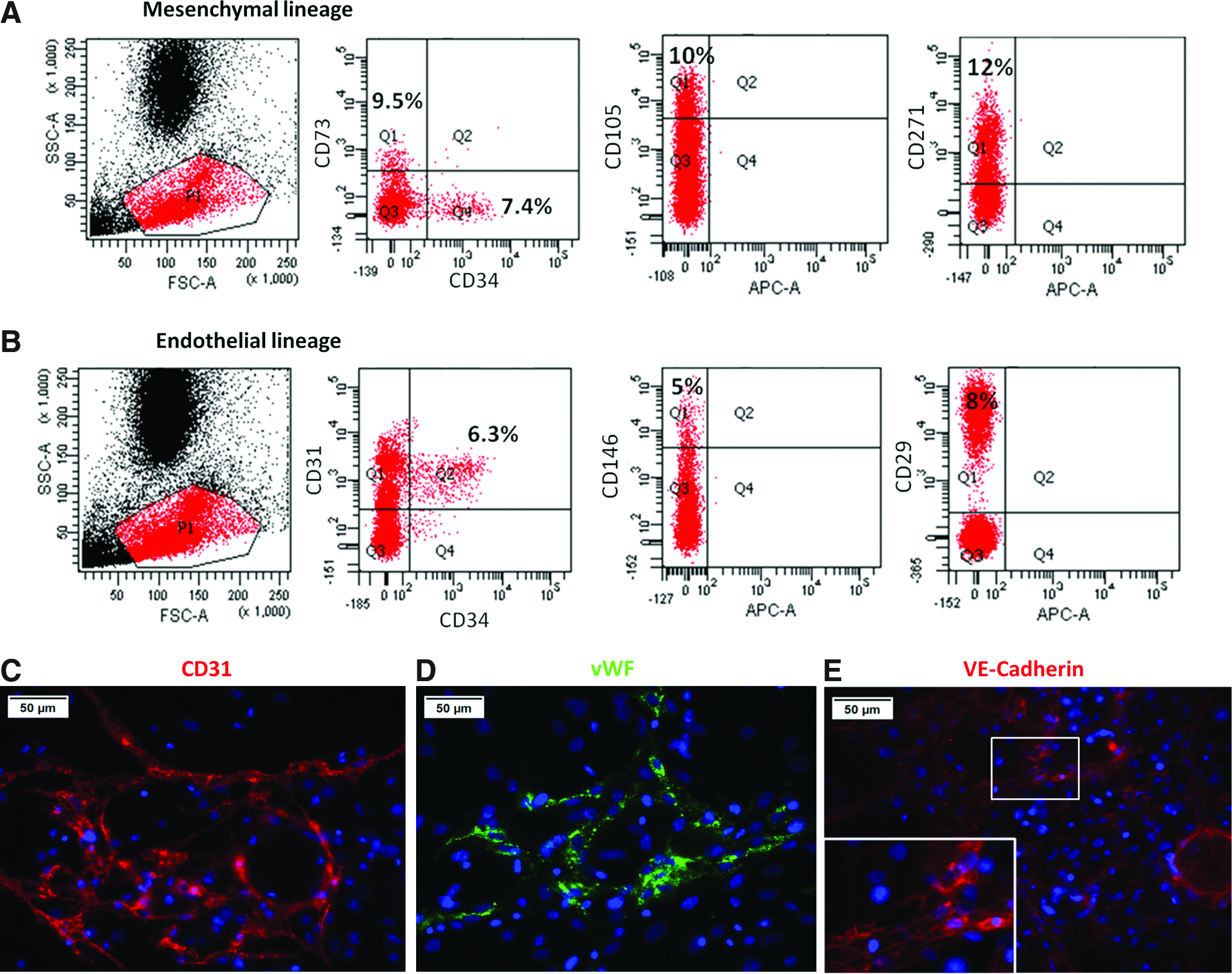
Flow cytometry analysis and immunolabeling of hWBMCs in 2D culture.
The expression of these EC-specific markers was further confirmed by the immunolabeling of CD31, vWF, and VE-Cadherin in 2D hWBMC cultures, after 5 days of culture (Fig. 1C–E). In addition, CD31 staining revealed that labeled cells formed a specific tubular-like network within the monolayer of hWBMC culture (Fig. 1C).
Three-dimensional hWBMC cultures: evidence for spheroid formation
To establish the behavior and distribution of the hWBMCs once seeded within the 3D porous polysaccharide-based scaffold (Fig. 2A), we analyzed their fate after 24 h of culture. Optical microscopy after calcein staining revealed that hWBMCs, which had been initially seeded within the 3D structure (Fig. 2B), migrated during the first hours of culture to form well-organized spheroids (Fig. 2C–F) distributed within the pores of the material. The aggregates measured 150±50 μm and appeared homogeneously distributed within the pores of the matrix.
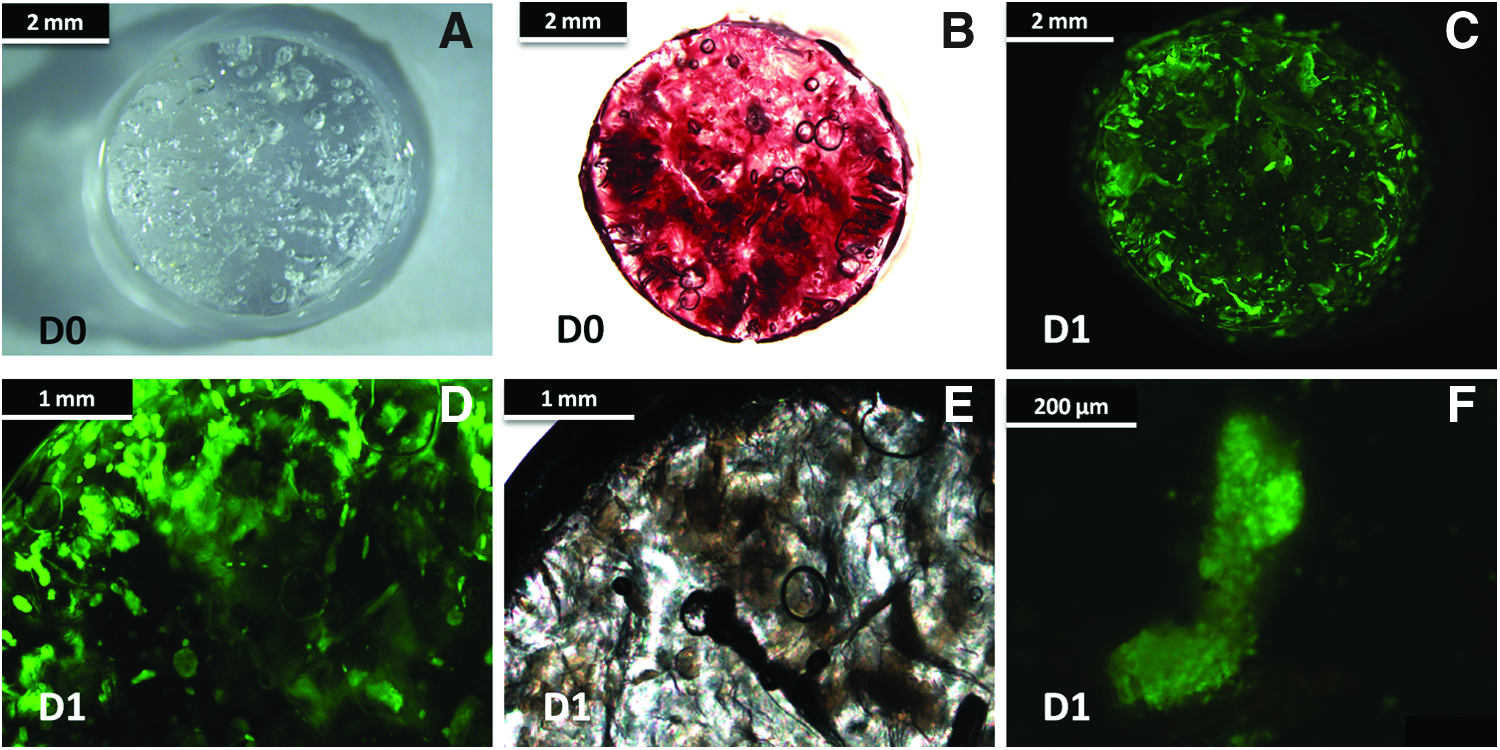
Spheroid formation within 3D matrix.
hWBMCs develop a network positive for endothelial markers within the spheroid formed in the 3D macroporous matrix
To evaluate the expression and distribution of endothelial-specific markers within the 3D hWBMC cultures, immunostaining for CD31 and vWF was performed after 1 day (D1), 6 days (D6), and 12 days (D12) of culture. A tube-like network was observed within the spheroids, as shown by the labeling for CD31 and vWF (Fig. 3A, B), well evidenced at days 6 and 12 in the cell aggregates for vWF labeling (Fig. 3B, C). The presence of vascular ECs was further confirmed by the quantification of CD31 and vWF gene expression from day 1 to 12 in the 3D cultures (Fig. 3D, E). Gene expression evaluation revealed a peak of expression of these two endothelial-specific markers after 6 days of culture within the matrices (Fig. 3D, E).
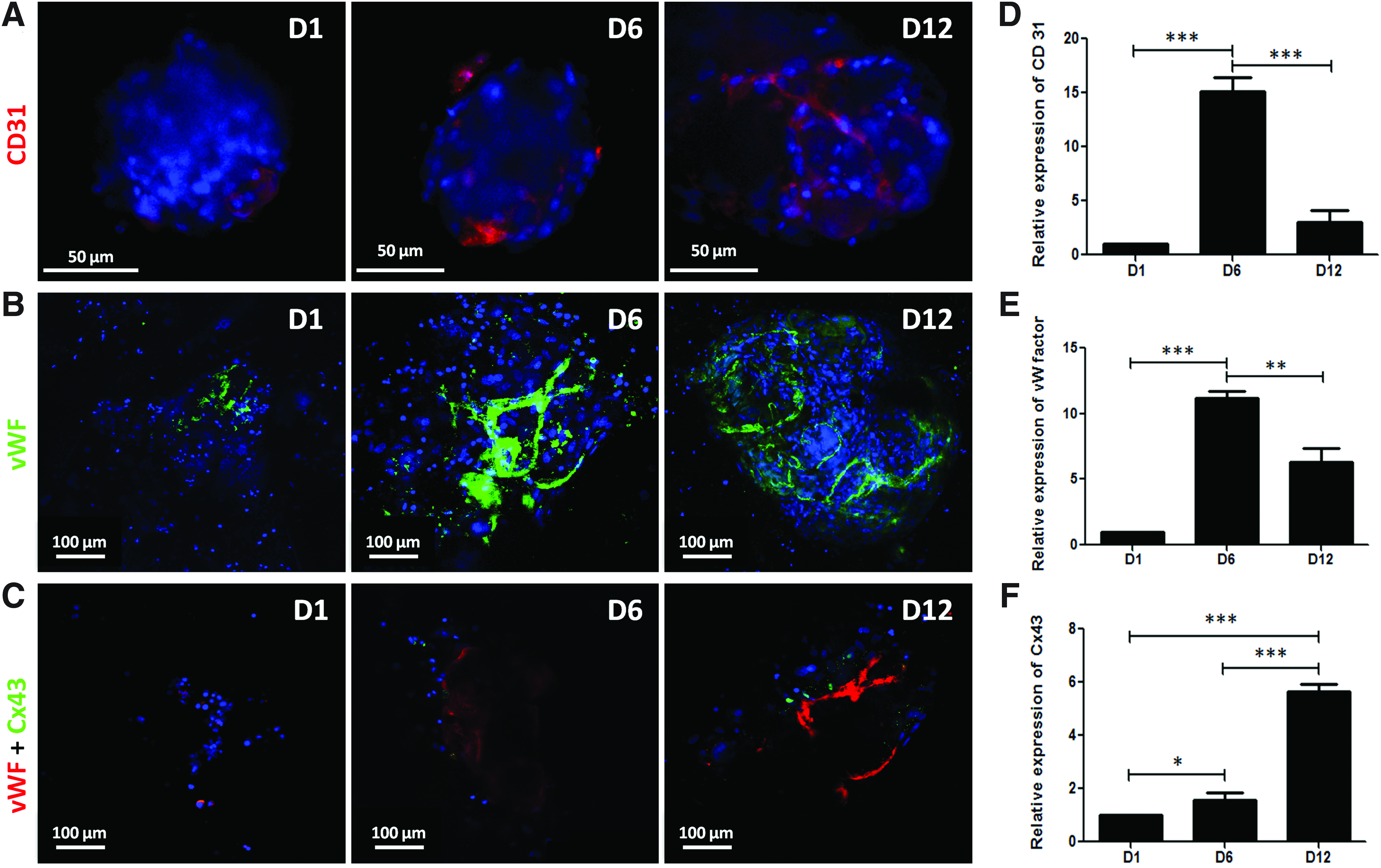
Immunolabeling and quantitative gene expression for endothelial specific markers of hWBMCs. Connexin43 (Cx43) immunolabeling and quantitative gene expression in hWBMCs.
hWBMC spheroid formation contributes to increased Cx43 expression
Since the 3D culture of hWBMCs within the matrix revealed a specific organization in spheroids, we assessed the expression of Cx43, a major actor of cell–cell communication in GAP junction protein family, after 1, 6, and 12 days of the 3D culture. Specific punctuated labeling of Cx43 appeared after 6 days of culture and was further evidenced after 12 days of culture. Cx43-positive cells were found to be in close proximity to vWF-positive cells (Fig. 3C, day 12). The quantification of Cx43 gene expression within the 3D matrices over time indicated a significant increase in the expression of this GAP junction protein from days 1 to 12. Indeed, a five-fold over-expression of Cx43 mRNA was obtained after 12 days of culture as compared with day 1 (Fig. 3F).
hWBMCs within spheroid conformation express bone-specific markers
We then assessed the ability of hWBMCs to express osteoblastic markers and to undergo osteoblastic phenotype differentiation within the 3D scaffold. The analysis of bone-specific markers in hWBMCs cultured within the 3D matrices was first qualitatively evaluated by cytochemistry for ALP activity, immunostaining for type 1 collagen and osteocalcin and alizarin red staining, on days 1 to 12 (D1 to D12) (Fig. 4A–D, respectively). The spheroids formed within the 3D matrix exhibited ALP activity and were positive for type I collagen during the time of culture (i.e., from day 1 to 12; Fig. 4A, B). OCN expression was shown at 1, 6, and 12 days of culture (Fig. 4C). Alizarin red staining showed an increased calcium deposition over the time of culture that was associated with the cellular aggregates (Fig. 4D). In addition, the gene expression of bone-specific markers (i.e., cbfa1/runx2, ALP, Col1A1, and OCN) was quantified in hWBMCs cultured within the 3D scaffold for 1, 6, and 12 days. Cbfa1 (a master gene for bone differentiation) expression was shown to increase during time of culture, till a three-fold increase at day 12, compared with day 1 (Fig. 5A). Both ALP and Col1A1 gene expression showed a peak at day 6 of culture. A significant decrease at day 12 was observed for ALP and Col1A1, but still with levels remaining significantly higher compared with day 1 (Fig. 5B, C). There was a significant steady increase in the gene expression of the late osteoblastic-specific marker OCN from day 1 until day 12 (Fig. 5D).

Immunolabeling of bone-specific markers in hWBMC 3D cultures.
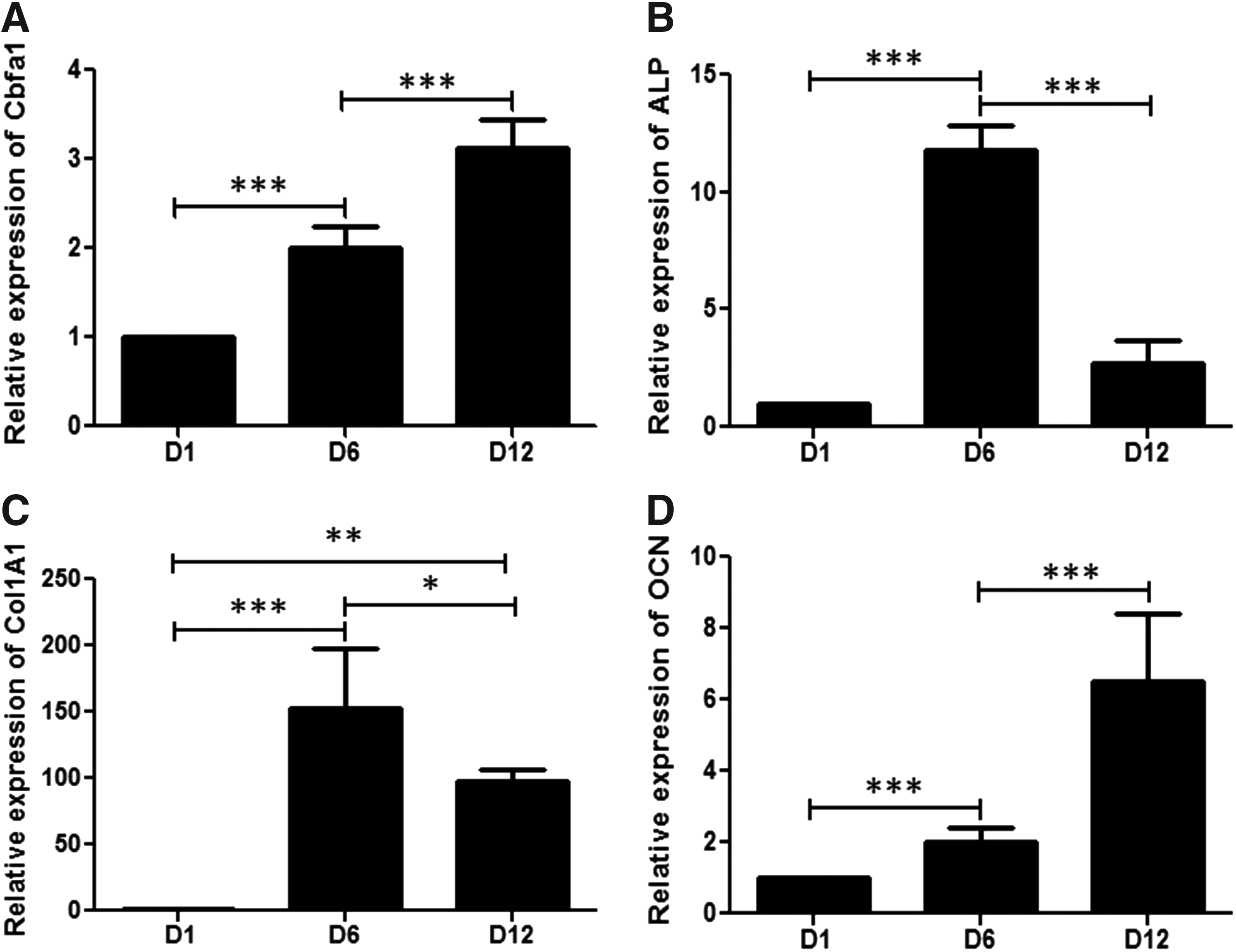
Quantitative analysis of bone-specific gene expression.
Three-dimensional cultures of hWBMCs promote osteoid formation ectopically
The potential of these tissue-engineered constructs for promoting bone formation was evaluated after 3 and 8 weeks on subcutaneous implantation in NOG mice by histological analysis using Masson's Trichrome staining. Empty scaffolds did not reveal any osteoid formation or vascularization inside the implants (Fig. 6A-1, A-2) at both 3 and 8 weeks postimplantation. On the other hand, scaffolds seeded with hWBMCs (Fig. 6B-1) presented a vascular network mainly around the implant (Fig. 6B-2) and an osteoid tissue formation (Fig. 6B-3) at 3 weeks postimplantation. After 8 weeks of implantation, the scaffolds seeded with hWBMCs (Fig. 6C-1) exhibited vessels that invaded the scaffold (Fig. 6C-2). In addition, the amount of osteoid tissue formed increased in comparison with the 3-week time point (Fig. 6C-3). Semi-quantitative image analysis showed a significant increase in the osteoid surfaces, the average size of the osteoid surfaces, and the average fraction of the osteoid surfaces at 8 weeks as compared with 3 weeks (Fig. 6–F, respectively).
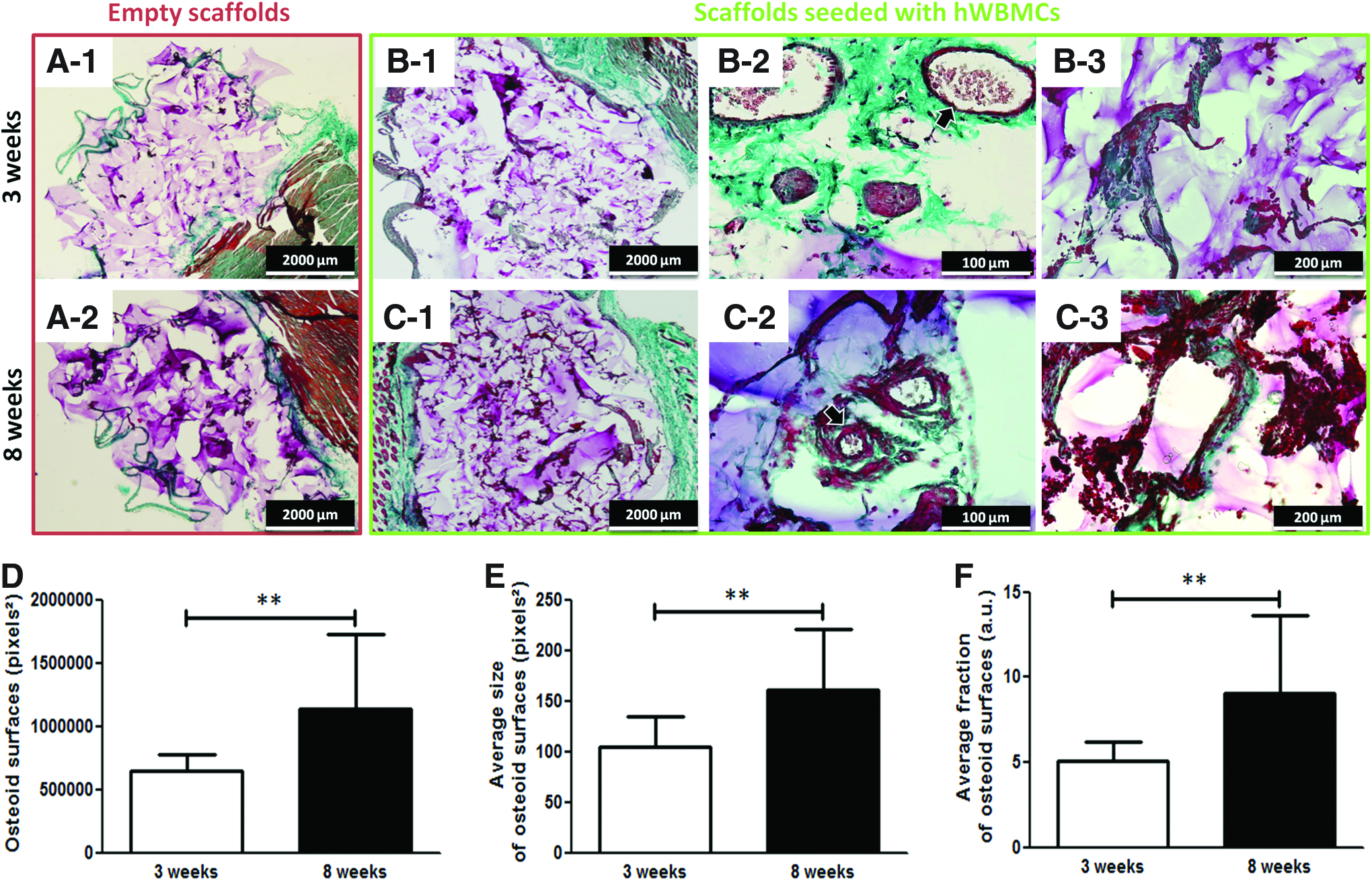
Subcutaneous implantation in mice of matrices seeded with hWBMCs: histological and quantitative analysis. Masson's trichrome staining of histological sections at 3 and 8 weeks of implantation of cylinders without cells
Three-dimensional cultures of hWBMCs within the matrix promote formation of blood vessels and are partially incorporated in autologous blood vessels
To assess the angiogenic potential of this tissue-engineered construct, the number of blood vessels was assessed both outside and inside the region of the implanted materials (Fig. 7A-1–A-3) by CD31 immunostaining. The outside region corresponds to the tissue in the vicinity of the implanted scaffold. CD31 immunolabeling showed that blood vessels and capillary structures were formed in the vicinity and inside the matrix (Fig. 7B-1–C-2) at both time points. When quantified, more vessels were observed outside the materials than inside the scaffold (Fig. 7D). Interestingly, the number of vessels significantly increased with time of implantation both outside and inside the matrix (Fig. 7D).
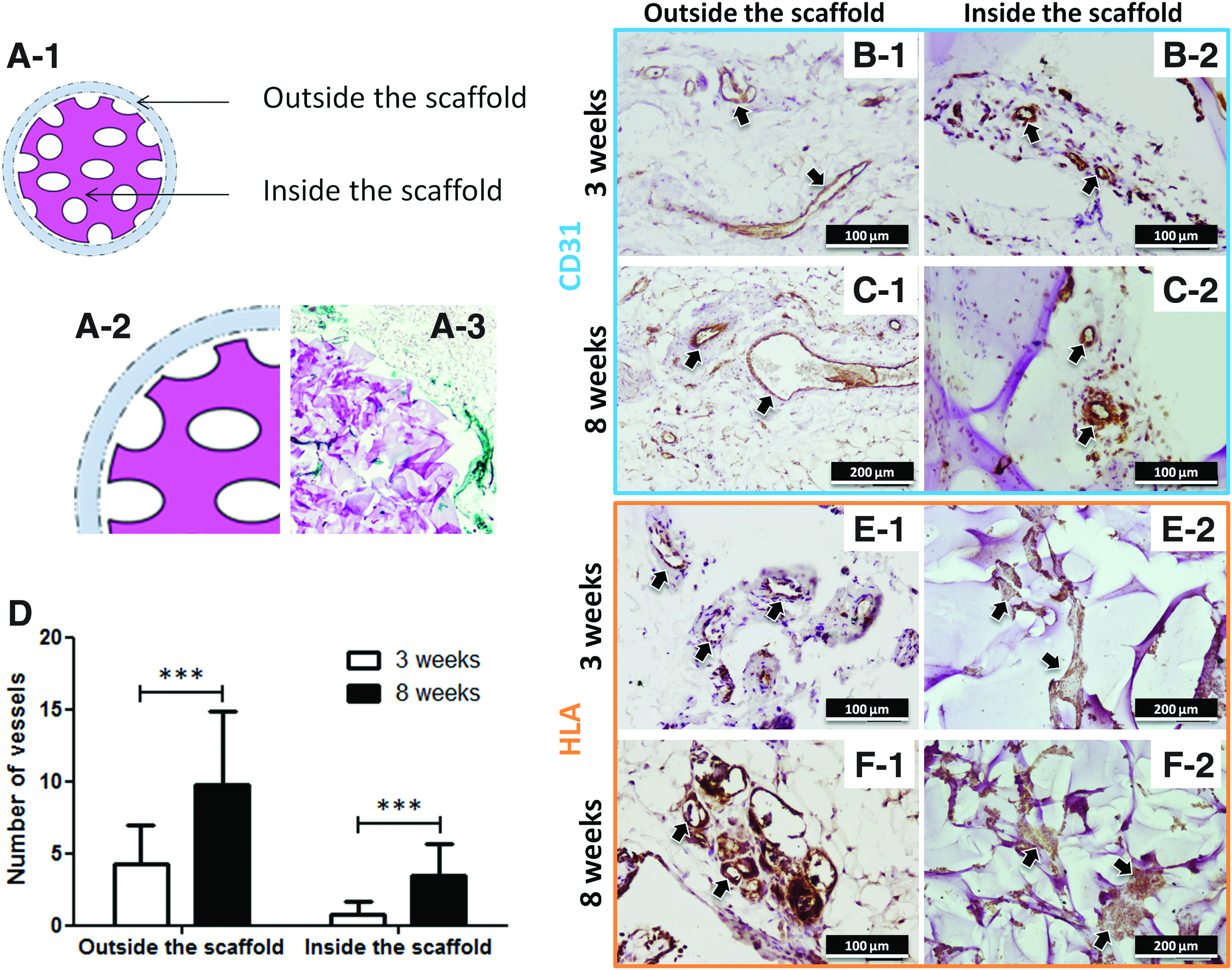
Subcutaneous implantation in mice of matrices seeded with hWBMCs: immunohistochemistry for CD31 and HLA-I.
The fate of the human cells after implantation was analyzed with human HLA-I specific immunolabeling. Cells positive for human HLA-I were observed in the vicinity of the material in structures resembling blood vessels (Fig. 7E-1, F-1, for 3 and 8 weeks, respectively). In addition, the cellular aggregates were maintained for approximately 8 weeks inside the scaffolds, as demonstrated by the HLA-I positive cells inside the materials (Fig. 7E-2, F-2, for 3 and 8 weeks, respectively).
Discussion
In this work, we focused on a further characterization of whole bone marrow cells and tested their potential to induce bone and vessel formation when seeded within a 3D matrix, without selection and 2D cell expansion. We found that the biodegradable natural scaffold composed of pullulan and dextran is an appropriate 3D microenvironment for promoting (1) the maintenance of these human bone marrow cells; (2) a specific cellular organization in spheroids promoting cell interactions and the establishment of junctional communications; (3) the activation of endothelial and osteoblastic markers; and (4) the formation of a microvascular network within the spheroids, observed both in vitro and in vivo, after ectopic implantation in mice.
For many years, bone marrow extracts have been used for treating nonunion gaps in orthopedic surgery. However, the understanding of the cellular interplay involved in the coupling between angiogenesis and osteogenesis remained elusive. Recently, the interplay between MSCs and ECs was reviewed. 56 This study shed light into the underlying mechanisms by which MSCs promote or inhibit neo-angiogenesis and into the interactions between MSCs and ECs in various physio-pathological conditions and in tissue regeneration. ECs derived from bone marrow have been shown to produce angiocrine factors that promote organ recovery and restore normal organ function, including bone tissue function. 57
Numerous studies have reported the use of multipotent MSCs as promising candidates to support bone formation and vascularization of tissue constructs. However, in tissue engineering applications, a common strategy consisted of the isolation of autologous MSCs from bone marrow samples before their seeding within a scaffold. 58 However, owing to the preselection of cells, these cultures were deprived of interactions with the other cell types present in the whole bone marrow suspension, such as cells from the hematopoietic compartment. In addition to MSCs, ECs or their progenitors from different sources (e.g., peripheral blood, cord blood) have been shown to successfully promote the formation of vessels and, thus, directly contribute to the formation of an intrinsic microvasculature within the implants. However, their application in clinical practice is hampered owing to the difficulties of co-culturing both autologous cell types from different sources, after their respective isolation and amplification, as shown by the limited number of clinical trials reported in the literature for bone tissue regeneration.
In this work, we confirmed the potential of whole human bone marrow as a candidate for bone tissue engineering for promoting both vasculature and bone formation. The characterization of cells from bone marrow by flow cytometry revealed that they express typical MSC markers such as CD73, CD105, and CD271 and are negative for CD45, the latter being a hematopoietic stem cell marker. In addition, we also detected a population expressing CD31, CD34, CD146, and CD29, which are typical EC markers. These markers are commonly used for bone marrow characterization by either positive or negative selection. 59 Our flow cytometry results reflect the variable proportion of MSC cells detected among different patients and are consistent with previous reports.2,60–62 The existence of an endothelial microvascular population from human bone marrow and their subsequent purification has already been described, 63 but the corresponding quantification and characterization were not undertaken.
While hWBMCs formed a multilayer with time of 2D culture, they were organized in spheroids for approximately 24 h of 3D culture that were distributed within the pores of the matrix when seeded in a polysaccharide-based scaffold. Here, we used a macroporous hydrophilic matrix composed of natural polysaccharides. Owing to their biochemical similarity with the extracellular matrix and inherent biocompatibility, pullulan and dextran are relevant candidates for the formation of tissue engineering constructs that are able to support cell viability and differentiation.47,64 Both have recently been used as a bioactive scaffold for human bone marrow MSCs co-cultured or not with human endothelial progenitor cells. 50 This 3D macroporous structure was prepared in the absence of organic solvents by the cross-linking of the biopolymers. This matrix exhibited interconnected pores and different porosities, key factors for improving cell migration and enabling the formation of blood vessels inside the scaffold. One of the main features of these matrices is their ability to support cellular interactions and to promote the formation of cell clusters. Our previous work showed that human bone marrow stromal cells co-cultured with endothelial progenitor cells from cord blood also generated spheroids in the same scaffold. 50
Three-dimensional spheroids have been extensively used in the field of angiogenesis.65–67 In particular, when spheroid-based engineering of a human vasculature was performed in mice, ECs could be grafted as spheroids into a matrigel-fibrin matrix and gave rise to a complex 3D network of human neovessels connected to the mouse circulation. 66 The authors developed a quantitative spheroid-based EC transplantation technique for the formation of durable vascular networks in vivo. Several approaches for generating cell spheroids have been described in the literature, including the hanging drop technique, 68 the carboxymethylcellulose technique, 66 and the liquid overlay method. 69 In this study, we used a simple biocompatible macroporous matrix that promotes the spontaneous formation of spheroids measuring 50 to 200 μm homogeneously distributed through the pores of the matrix. Moreover, they led to the formation of CD31-positive vascular structures that were well evidenced after 12 days of culture. vWF staining confirmed the presence of differentiated ECs, mainly in the periphery of the spheroids after 12 days of 3D culture.
Other scaffolds used for bone tissue engineering, such as polyurethane, 25 polycaprolactone starch scaffold, 70 polylactic acid 71 seeded with co-cultures of ECs, and osteoblastic cells from different sources and at different stages of cell differentiation, have been shown to support the formation of tube-like structures and vessel-like formation. Here, 3D culture with hWBMCs induced the conformation of aggregates, unlike the conformation observed with the 2D monolayer cultures of hWBMCs, which led to a tubular-like network formed by CD31 or vWF-positive cells. Indeed, within the 3D matrix, tubular structures were restricted to the spheroids, and, in some cases, networks between two or more spheroids could be observed.
In our study, spheroid aggregation allowed cell stabilization during the first day of culture and improved physiological conditions due to augmented cell-to-cell contact formation. Moreover, the spheroids formed exhibited an increasing expression of Cx43, a major protein involved in gap junctional communication that plays a key role in bone cell differentiation. 72 Cx43-positive labeling was evidenced at day 12 and was found close to the CD31-positive vascular structures, strongly suggesting its effect in cell differentiation toward osteoblastic lineage. However, the identity of these labeled cells remains elusive. Cell-to-cell contacts observed with hWBMC cultures were also shown for co-cultures of MSCs and PDECs in the same scaffold. The key role of this protein in bone cell differentiation was previously demonstrated with the use of inhibitory peptides for Cx43. 50
Cell-to-cell contact drives fundamental events for cell phenotype establishment. It is widely thought that the spheroid 3D microenvironment markedly influences the behavior and function of the incorporated stem cells. Our findings demonstrate that hWBMC cultures in 3D spheroids and within the matrix possess the ability to differentiate into microvascular ECs. The fact that CD31 and vWF in vitro expression decreased between days 6 and 12 could suggest a senescence of these human microvascular ECs, which are present in the whole bone marrow fraction, over the long-term culture period in vitro. Although spheroidal aggregation may protect cells from apoptosis, they could not survive in static culture conditions, as the diffusion of oxygen and nutrients in static conditions is probably limited within the scaffolds and the spheroids73,74 in static cultures.
Regarding the fate of MSCs and their orientation to the osteoblastic lineage, quantitative analysis performed by q-PCR confirmed the increased expression of all bone-specific markers for hWBMCs cultured within the matrix, with expression profiles in accordance with the corresponding bone markers. Cbfa1, an early bone-specific marker, was over-expressed from day 1 to 12 in the hWBMC culture. Maximal stimulation of ALP mRNA, an early bone marker, occurred after 6 days of 3D culture and then decreased. The same expression pattern was observed for Col1A1. Both genes are early markers for bone differentiation, which might explain the gene expression peak at 6 days of culture and the subsequent decrease at day 12. The late osteoblastic marker OCN involved in matrix mineralization showed an increased expression from day 1 to 12 in hWBMC cultured cells. The calcium deposition in the matrix confirmed the mineralization of the extracellular matrix. Cytochemical analysis, immunostaining of bone markers, and their quantitative analysis confirmed that this tridimensional cellular conformation was able to sustain the expansion and differentiation of osteogenic cells arising from hWBMCs without the need for preliminary 2D selection or expansion cultures.
Ectopic implantation of the matrix loaded with hWBMCs validated the suitability of the spheroidal structures to support bone and vessel formation in vivo. Osteoid formation increased with time of implantation from 3 to 8 weeks. Importantly, we also demonstrated that hWBMC spheroid-seeded scaffolds supported an increased vascularization in the periphery of the scaffold and inside the matrix with time of implantation. The role of the hWBMCs loaded within the matrix and their impact in the neovascularization process of the implant was assessed by immunolabeling a specific human marker (i.e., HLA-I). Human cells were detected in the aggregates after 8 weeks of implantation inside the scaffold, suggesting their contribution to bone formation and/or the vascularization of the tissue. In addition, positive cells associated with vessels were observed in the periphery of the implants. However, the exact role of these human cells loaded in this scaffold for tissue regeneration and their potential interactions with the murine cells remain unknown. Scaffolds seeded with hWBMCs may contribute to the vascularization of the implant by the release of growth factors, including chemoattractive factors and angiogenic factors, thereby facilitating the recruitment of cells inducing in vivo the formation of new vessels arising from the host cells. In addition, further studies considering the use of this approach in a bone defect are underway, aiming at confirming the potential of this cell source, in combination with the selected biomaterial, to improve bone regeneration.
Conclusion
This study demonstrates that the hWBMC population, without preliminary isolation or 2D expansion, can be used directly after extraction for the development of a bone regeneration and vascularization strategy, once seeded in an appropriate bioactive matrix. The ability of the 3D matrix to promote spheroids spontaneously contributes to the formation of a vascularized tissue. This approach is, therefore, suitable for improving the efficacy of MSC-based vascularization strategies in tissue-engineering applications without the need for preliminary 2D or 3D cultures, enabling a simplified approach with higher potential to reach the bedside.
Footnotes
Acknowledgments
This work was supported by grants from INSERM (National Institute for Health and Medical Research), from University of Bordeaux, Universities of Paris 7 and Paris 13, and by grants from the French Research National Agency (ANR-10-EMMA-009-01 MATRI+; ANR-12-TecSan-0011 INEOV). The authors thank Vincent Pitard and Santiago Gonzalez from the cytometry facility (University of Bordeaux, France). They are also grateful to R. Cooke for editing this article.
Authors' Contributions
Julien Guerrero: Conception and design, article writing. Hugo Oliveira: Conception and design, article writing. Sylvain Catros: Conception and design of in vivo experiments. Robin Siadous: Conception and design. Mohammed Derkaoui: Conception and design, provision of study materials. Reine Bareille: Conception and design, provision of study materials. Didier Letourneur: Financial support, data analysis and interpretation, and article writing. Joëlle Amédée: Financial support, data analysis and interpretation, and article writing.
Disclosure Statement
S.M.D. and D.L. are shareholders of the company IMMATIS for the development and commercialization of the reported matrices. For all other authors, no competing financial interests exist.