
Abstract
The periodontium, consisting of gingiva, periodontal ligament (PDL), cementum, and alveolar bone, is necessary for the maintenance of tooth function. Specifically, the regenerative abilities of cementum with inserted PDL are important for the prevention of tooth loss. Periodontal ligament stem cells (PDLSCs), which are located in the connective tissue PDL between the cementum and alveolar bone, are an attractive candidate for hard tissue formation. We investigated the effects of recombinant human plasminogen activator inhibitor-1 (rhPAI-1) on cementogenic differentiation of human PDLSCs (hPDLSCs) in vitro and in vivo. Untreated and rhPAI-1-treated hPDLSCs mixed with hydroxyapatite/tricalcium phosphate (HA/TCP) and dentin matrix were transplanted subcutaneously into the dorsal surface of immunocompromised mice to assess their capacity for hard tissue formation at 8 and 10 weeks posttransplantation. rhPAI-1 accelerated mineral nodule formation and increased the mRNA expression of cementoblast-associated markers in hPDLSCs. We also observed that rhPAI-1 upregulated the levels of osterix (OSX) and cementum protein 1 (CEMP1) through Smad2/3 and p38 pathways, whereas specific inhibitors of Smad3 and p38 inhibited the enhancement of mineralization of hPDLSCs by rhPAI-1. Furthermore, transplantation of hPDLSCs with rhPAI-1 showed a great ability to promote cementogenic differentiation. Notably, rhPAI-1 induced hPDLSCs to regenerate cementum-like tissue with PDL fibers inserted into newly formed cementum-like tissue. These results suggest that rhPAI-1 may play a key role in cementogenic differentiation of hPDLSCs. rhPAI-1 with hPDLSCs may be a good candidate for future clinical applications in periodontal tissue regeneration and possibly in tooth root bioengineering.
Introduction
P
Human DMSCs originate from various types of dental tissues, including PDL, pulp, periapical follicle, and alveolar bone, all of which have multilineage differentiation abilities.2–4 PDL, located between the cementum and alveolar bone, is specialized connective tissue containing fibroblasts, epithelial cells, undifferentiated mesenchymal cells, bone cells, and cementum cells. It has been reported that some of these cells have the potential to generate bone and cementum. The functions of PDL are supportive, sensory, nutritive, and remodeling of the periodontium, thus PDL generation is as important as cementum formation. Our previous studies showed successful isolation and characterization of human periodontal ligament stem cells (hPDLSCs) from PDL, thus increasing their potential as candidates for therapeutic application. 2 PDLSCs can differentiate into cementoblasts, osteoblasts, adipocytes, and chondrocytes.5–8 Recent studies indicate that PDLSCs have many osteoblast- and cementoblast-like properties and can differentiate into mineral-forming cells that express bone- and cementum-associated markers, such as alkaline phosphatase (ALP), bone sialoprotein (BSP), type 1 collagen (Col1), osteopontin (OPN), and osteocalcin (OCN).7,9–13 Cementum protein 1 (CEMP1) was first isolated and characterized as a human cementoblastoma-derived protein that is expressed in cementoblasts, and studies suggested that CEMP1 might play a critical role as a local regulator of cementoblast differentiation.14,15 F-spondin was recently identified as a highly expressed cementoblast-specific gene and a promoting factor for cementoblast differentiation. 16 Moreover, PDLSCs showed the capacity to generate cementum with PDL in vivo.3,17 Therefore, hPDLSCs would be the most appropriate candidates in clinical trials and tissue engineering for cementum regeneration. A typical tissue engineering strategy involves cells, biomaterial scaffolds, and bioactive molecules. To enhance the therapeutic effects, growth factors and cytokines are usually introduced into the systems by direct protein delivery or viral gene delivery.
Plasminogen activator inhibitor type 1 (PAI-1) plays important roles in many physiological processes, such as fibrotic disorders, metabolic disorders, and cancer.18–21 The plasminogen/plasmin proteolytic cascade, which includes two plasminogen activators, tissue-type plasminogen activator (tPA) and urokinase-type plasminogen activator (uPA), and can be inhibited by PAI-1, also plays an important role in extracellular matrix remodeling. 22 PAI-1 is expressed by bone cells and their substrates and present in bone matrix, known to be a regulator of bone remodeling. 23 We have previously demonstrated that platelet-rich fibrin (PRF) promoted the differentiation of the human alveolar bone marrow stem cells and high-level expression of PAI-1 in PRF. 24 Moreover, the combination of PRF and autogenous bone increased the rate of osteogenesis and the quality of the new bone. 25 Taken together, these findings indicate that PAI-1 contributes to hard tissue formation, although previous studies have not clarified the role of PAI-1 in cementogenic differentiation.
In this study, we aimed to investigate the effects of recombinant human plasminogen activator inhibitor-1 (rhPAI-1) on cementogenic differentiation of hPDLSCs in vitro and in vivo and to identify the molecular mechanisms by which cementogenic differentiation occurs. We found that rhPAI-1 significantly stimulated hPDLSCs to promote cementum formation with inserted PDL fibers in a transplantation model in vivo. These results suggest that rhPAI-1 is an important factor for regulating periodontal tissue regeneration in hPDLSCs.
Materials and Methods
Primary cell culture
Human third molars were collected from three healthy young males (18–25 years old) under the protocol approved by the Institutional Review Board of the Seoul National University Dental Hospital, Seoul, South Korea (IRB No. 05004). PDL was gently separated from extracted human third molars and the separated tissues were digested in a solution of 3 mg/mL collagenase type I (Worthington Biochem, Freehold, NJ) and 4 mg/mL dispase (Boehringer, Mannheim, Germany) for 1 h at 37°C. Single-cell suspensions were obtained by passing the cells through a 40-μm strainer (Falcon BD Labware, Franklin Lakes, NJ) and were cultured in the alpha-modification of Eagle's medium (α-MEM; Gibco BRL, Grand Island, NY) supplemented with 10% fetal bovine serum (Gibco BRL), 100 μM ascorbic acid 2-phosphate (Sigma-Aldrich, St. Louis, MO), 2 mM glutamine, 100 U/mL penicillin, and 100 μg/mL streptomycin (Biofluids, Rockville, MD) and incubated at 37°C in 5% CO2. The medium was changed after the first 24 h and then every 3 days. The colonies were picked and cultured separately. Three colonies of hPDLSCs were randomly selected and the cellular pool of those colonies was used for in vitro proliferation and differentiation studies. All primary cells used in this study were in passage 2 or 3.
Flow cytometric analysis
To characterize the immunophenotype of hPDLSCs, the expression of mesenchymal stem cell (MSC)-associated surface markers at passage 3 was analyzed by flow cytometry as described previously. 26 Briefly, cells in their third passage (1.0×106 cells) were fixed with 3.7% paraformaldehyde for 10 min and resuspended in phosphate-buffered saline (PBS) containing 1% bovine serum albumin (BSA) (ICN Biomedicals, Aurora, OH) for 30 min for blocking. Cells were then incubated with specific antibodies for CD34, CD13, CD90, or CD146 at 4°C for 1 h, followed by incubation with fluorescence secondary antibodies at room temperature for 1 h. All antibodies were purchased from BD Biosciences (San Jose, CA). The percentage of CD13-, CD90-, and CD146-positive and CD34-negative cells was measured with a FACSCalibur flow cytometer (Becton Dickinson Immunocytometry Systems, San Jose, CA) and the results were analyzed using CellQuest Pro software (Becton Dickinson Immunocytometry Systems).
Chondrogenic and adipogenic differentiation
For induction of chondrogenic and adipogenic differentiation, the cells were cultured in StemPro Chondrogenic and StemPro Adipogenic differentiation media (Gibco BRL), respectively, with the appropriate supplements. At week 3 postchondrogenic and postadipogenic induction, the cells were washed with PBS and fixed in 3.7% paraformaldehyde for 10 min. The cells were stained with 1% Alcian Blue (Sigma-Aldrich) and 0.3% Oil Red O dye (Sigma-Aldrich) for detection of proteoglycans and fat vacuoles as indicators of chondrogenic and adipogenic differentiation, respectively. Cells were visualized under an inverted light microscope (Olympus U-SPT; Olympus, Tokyo, Japan).
Cell proliferation/cytotoxicity assay
Cell proliferation was measured using the colorimetric 3-(4,5-dimethylthazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay kit (Promega, Madison, WI). Briefly, hPDLSCs (3.0×103 cells/well) were seeded in 96-well plates and cultured for 48 h. Various amounts of rhPAI-1 (Prospec-Tany TechnoGene, Rehovot, Israel) were added in 100 μL of culture media per well for final concentrations of 0, 5, 10, 20, 50, and 100 ng/mL rhPAI-1. A premixed optimized dye solution (15 μL) was added at the end of the treatment. Cells were incubated in 5% CO2 at 37°C for 4 h and then 100 μL of a solubilization/stop solution was added to the cultures to solubilize the formazan product. Each condition was prepared in triplicate, and reactions were assessed using an ELISA reader at OD 595nm (reference, 655 nm).
Alkaline phosphatase activity assay and staining
Quantitative analysis of ALP activity was performed using the p-Nitrophenyl Phosphate (pNPP) Liquid Substrate System according to the manufacturer's instructions (Sigma-Aldrich). In brief, the ALP activity in cell lysates was determined with pNPP as the substrate in an assay buffer containing 5 mM MgCl2 and 50 mM Na2CO3. Absorbance at 405 nm was measured using an ELISA reader, and the ALP activity was calculated from a standard curve. For ALP staining, cells were fixed with 10% formalin, incubated with 0.1% Triton X-100 for 5 min, and stained using the Leukocyte Alkaline Phosphatase Kit (Sigma-Aldrich) according to the manufacturer's protocol.
Induction of mineralization and Alizarin red S staining
For mineralization, the hPDLSCs were cultured in the osteogenic differentiation medium with 50 μg/mL ascorbic acid, 10 mM β-glycerophosphate, and 100 nM dexamethasone (Sigma-Aldrich) for 3 weeks. On day 21, accumulation of mineral nodules was detected by staining with 2% Alizarin red S staining at pH 4.2 (Sigma-Aldrich). For the destaining procedure to measure the calcium content, 3 mL of 10 mM sodium phosphate–10% acetylpyrimidium (pH 7.0) solution was added to each stained well and incubated at room temperature for 15 min. The destained sample was transferred to a 96-well plate and the absorbance was measured at 562 nm.
Western blot analysis
The hPDLSCs (1.0×106 cells/dish) were seeded in a 60-mm culture dish and cultured for the indicated duration. Protein concentrations of cell lysates were determined using the DC Protein Assay Kit (Bio-Rad Laboratories, Hercules, CA). Equal amounts of protein (30 μg/lane) were resolved by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to a polyvinylidene difluoride membrane (GE Healthcare, Buckinghamshire, United Kingdom). Primary antibodies against phospho-ERK, ERK, phospho-p38, p38, phospho-JNK, JNK, Runx2, α-tubulin (Cell Signaling Technology, Danvers, MA), osterix (OSX; Abcam, Cambridge, United Kingdom), CEMP1, F-spondin, p-Smad2/3, and Smad4 (Santa Cruz Biotechnology, Santa Cruz, CA) were used for the mechanism study. Blots were developed using horseradish peroxidase-conjugated secondary antibodies (Cell Signaling Technology) and visualized using an enhanced chemiluminescence kit (GE Healthcare).
Reverse transcription–polymerase chain reaction
To evaluate gene expression levels in rhPAI-1-induced differentiated hPDLSCs, 1.0×106 cells were seeded in a 60-mm culture dish and cultured under osteogenic differentiation induction conditions. Total RNA was prepared using an RNeasy Mini Kit (Qiagen, Valencia, CA) according to the manufacturer's instructions, and cDNA was synthesized from 2 μg of total RNA using reverse transcriptase (Superscript II Preamplification System; Invitrogen, Gaithersburg, MD). Reverse transcription–polymerase chain reaction (RT-PCR) was conducted as described previously. 17 The relative expression of mRNA was compared in a histogram. The specific primer sets used for RT-PCR are listed in Table 1.
ALP, alkaline phosphatase; BSP, bone sialoprotein; CEMP1, cementum protein 1; Col1, type 1 collagen; OPN, osteopontin; OCN, osteocalcin.
Luciferase assay
The hPDLSCs were seeded on a 24-well plate at a density of 5.0×104 cells per well. After 24 h, the cells were transfected with Lipofectamine LTX and PLUS reagent (Invitrogen) according to the manufacturer's instructions. Luciferase reporter plasmid of pGL-basic 6× OSE was kindly provided by Dr. Z.H. Lee (Seoul National University). For each transfection, 0.4 μg of luciferase reporter plasmid pGL-basic vector (control) or expression vector containing six repeats of the consensus Runx2 binding site (6× OSE) promoter was used. Transfected cells were treated with or without rhPAI-1 for 48 h and then lysed for assessment of luciferase activity using the luciferase reporter gene assay system (Promega) according to the manufacturer's instructions. The measurements were performed with a luminometer (FLUOstar OPTIMA; BMC Laboratory, Offenburg, Germany), and the experiments were conducted in triplicate.
Preparation of human-treated dentin matrix and beagle dog periodontium
Periodontal tissue, including PDL and cementum, was completely removed using a surgical bur by grinding along the root surface. The roots were cut into two pieces following the midsagittal plane and the dental pulp, odontoblast, and predentin were completely removed. Finally, the dentin was trimmed to a cylinder 5.0 mm in height and 2.0 mm in diameter. Fabrication of a human-treated dentin matrix was performed as reported previously. 27
A beagle dog aged 10 months with 10 kg body weight was obtained and examined for good systemic and oral health. This study was reviewed and approved by the Institutional Animal Care and Use Committee (No. 070504-5, ilar.snu.ac.kr, IACUC) at Seoul National University (Seoul, Korea). The harvest procedure of periodontium was performed as reported previously. 17
Transplantation and histological analysis
To study the effects of rhPAI-1 on cementum formation in vivo, hPDLSCs (1.0×107 cells) were mixed with 100 mg hydroxyapatite/tricalcium phosphate (HA/TCP) ceramic powder (Zimmer, Inc., Warsaw, IN) and human tooth root dentin matrix, with or without rhPAI-1 (100 μg), on a 0.5% fibrin gel. Cells were then transplanted subcutaneously into immunocompromised mice (n=6) (NIH-bg-nu/nu-xid; Harlan Sprague Dawley, Indianapolis, IN). In this transplantation model, rhPAI-1 was used to induce cementogenic differentiation in hPDLSCs with HA/TCP and human tooth root dentin matrix. For histological analysis, samples were obtained 8 and 10 weeks after transplantation and fixed in 3.7% paraformaldehyde solution for 48 h at 4°C before decalcification with 12% EDTA (pH 7.4) at 4°C. The samples were embedded in paraffin and cut serially, starting from the outermost region, in the dentin midsagittal plane. Semiserial 5-μm sections were prepared for hematoxylin and eosin (H&E) and immunohistochemical staining and examined under a light microscope (Olympus U-SPT; Olympus).
For immunohistochemistry, the deparaffinized sections were immersed in 0.6% H2O2/methanol for 20 min to quench the endogenous peroxidase activity. Sections were then preincubated with 1% BSA in PBS for 30 min and incubated overnight at 4°C with the rabbit polyclonal antibody against CEMP1 (1:200), OSX (1:100; Abcam), Col1, and OPN (1:100; Santa Cruz Biotechnology). Sections were incubated for 1 h at room temperature with the secondary antibody and reacted with the avidin–biotin–peroxidase complex (Vectastain ABC Systems; Vector Laboratories, Burlingame, CA) in PBS for 30 min. After color development with 0.05% 3,3′-diaminobenzidine tetrahydrochloride (DAB peroxidase substrate; Vector Laboratories), the sections were counterstained with hematoxylin.
PDL was identified by staining with Picrosirius red staining to reveal collagenous fibers. The Picrosirius red solution (ScyTek Laboratories, Logan, UT) was added to the deparaffinized sections to completely cover the tissue and incubated for 1 h. Sections were rinsed quickly in two changes of acetic acid solution and absolute alcohol and then counterstained with hematoxylin.
Scanning electron microscopy
Samples were fixed in 0.1 M cacodylate buffer (pH 7.3) containing 2.5% glutaraldehyde for 30 min and in 0.1 M cacodylate buffer (pH 7.4) containing 1% osmium tetroxide for 1 h. After rapid dehydration through an ethanol gradient, critical point drying, and sputter coating with gold, the PDL fibers were observed under scanning electron microscopy (SEM, S-4700; Hitachi, Tokyo, Japan).
Statistical analysis
Statistical analysis was performed using Statistical Package for Social Science 13.0 software (SPSS, Inc., Chicago, IL). Normal data with equal variance were analyzed using one-way analysis of variance (ANOVA) with a Tukey's procedure. Significance was defined as p≤0.05. Values in each graph represent mean±standard deviation. All assays were performed at least thrice and representative data are presented.
Results
Characterization of hPDLSCs
The isolated cells formed single-cell-derived colonies, and three different colonies from each of the three donors were randomly selected as hPDLSCs for studies. Flow cytometric analysis showed that ∼95.7% of hPDLSCs expressed CD13, 96.84% expressed CD90, 90.17% expressed CD146, and 1.05% expressed CD34 (Fig. 1A). The percentage of positive cells was determined by the relative intensity of antibody-binding cells. CD13, CD90, and CD146 are putative positive markers of MSCs,8,28 whereas CD34 is an MSC-negative marker that marks primitive hematopoietic progenitors and endothelial cells. 29 To investigate the multilineage differentiation potential of hPDLSCs, the cells were incubated without or with osteogenic medium supplementation to induce mineralization in vitro. After 3 weeks of osteogenic induction, hPDLSCs formed extensive amounts of Alizarin red S-positive mineral deposits throughout the adherent layers. The cells were also capable of undergoing adipogenic and chondrogenic differentiation after incubation with adipogenic-inductive and chondrogenic-inductive supplements for 3 weeks. Oil Red O and Alcian Blue staining showed lipid droplet formation and Alcian Blue-positive nodules, respectively (Fig. 1B).

Characterization and multilineage differentiation of hPDLSCs.
Recombinant human PAI-1 induces cementogenic differentiation of hPDLSCs in vitro
To examine the effects of rhPAI-1 on cell viability in vitro, hPDLSCs were treated with 0, 5, 10, 20, 50, or 100 ng/mL of rhPAI-1. MTT assay showed that rhPAI-1 did not have any cytotoxicity after 48 h of culture, even at high doses. Furthermore, rhPAI-1 still had no cytotoxicity in hPDLSCs after long-term culture of 1, 2, and 3 weeks (data not shown). In addition, we observed that rhPAI-1 increased proliferation of hPDLSCs at a low dose of 10 ng/mL (Fig. 2A). Next, we investigated the effects of rhPAI-1 on the differentiation of hPDLSCs in vitro. After hPDLSCs were cultured in the osteogenic differentiation medium treated without or with rhPAI-1 for 2 weeks, we observed only 50 and 100 ng/mL of rhPAI-1-induced osteogenic/cementogenic differentiation. Calcium deposition and ALP activity of hPDLSCs were significantly increased in 100 ng/mL of rhPAI-1-treated group compared to 50 ng/mL of rhPAI-1-treated group and control group (Fig. 2B, C). Higher calcium contents were observed in the rhPAI-1-treated group after a destaining procedure of Alizarin red S staining (Fig. 2B, right panel). The mRNA expression of osteogenic/cementogenic differentiation markers, such as ALP, Col1, OPN, OCN, BSP, and Runx2, increased significantly after culture with rhPAI-1 compared to untreated cells (Fig. 2D). Importantly, expression of F-spondin and CEMP1, well known as cementoblast-specific markers, was also significantly increased by rhPAI-1. To assess cementogenic differentiation ability of rhPAI-1, we measured the transcriptional activity of 6× OSE promoter. As shown in Figure 2E, rhPAI-1 treatment stimulated the transcriptional activity of 6× OSE promoter. These results suggest that rhPAI-1 can enhance the Runx2 signaling pathway to stimulate osteoblast/cementoblast-associated gene expression through Runx2 activation. Therefore, the Runx2 signaling pathway may play a critical role in cementogenic differentiation as well as osteogenic differentiation. Thus, in the osteogenic differentiation medium, rhPAI-1 could induce the differentiation of hPDLSCs into mineral-forming cells, such as cementoblasts in vitro.

Effects of rhPAI-1 on cementogenic differentiation of hPDLSCs.
Recombinant human PAI-1-induced OSX and CEMP1 expression is mediated by p38 and Smad2/3 pathways in hPDLSCs
To inspect the key molecular mechanisms by which rhPAI-1 induces differentiation of hPDLSCs, the first targets to be examined should be among the members of transcription factors involved in osteogenesis/cementogenesis. Recent studies indicated that Runx2 and its downstream target OSX play critical roles in cementogenic differentiation, which is important for osteogenesis.30,31 Furthermore, OSX particularly controls the formation of cellular cementum. 32 During in vitro cementogenic differentiation, rhPAI-1 increased Runx2 expression at 7 days and subsequently increased OSX expression at 14 days. CEMP1 and F-spondin expression also increased at 14 days of culture (Fig. 3A). The expression levels of mitogen-activated protein kinases (MAPKs) and Smads were examined as these proteins are thought to be involved in the early signaling pathways. 33 Among these proteins and their phosphorylated forms, rhPAI-1 increased p-p38 activation without affecting p-JNK and p-ERK. As expected, rhPAI-1 upregulated the levels of p-Smad2/3, whereas Smad4 showed no change (Fig. 3B). To confirm whether p38 and Smad2/3 play an essential role in rhPAI-1-induced cementogenic differentiation of hPDLSCs, we investigated the effects of their specific inhibitors, SB203580 and SIS3, respectively, on differentiation induction.34,35 Compared to Alizarin red S staining positivity of rhPAI-1 group after 14 days of differentiation induction, decreased calcium nodule formation was observed in the presence of rhPAI-1 with the SB203580 and SIS3 group (Fig. 3C). The calcium contents induced by rhPAI-1 with SIS3 and SB203580 treatment were verified after destaining procedures (Fig. 3C, bottom panel). Therefore, SB203580 and SIS3 inhibited rhPAI-1-induced mineralization of hPDLSCs in vitro. Taken together, these results suggest that OSX and CEMP1 seem to play an essential role in rhPAI-1-induced cementogenic differentiation of hPDLSCs through p38 and Smad2/3.
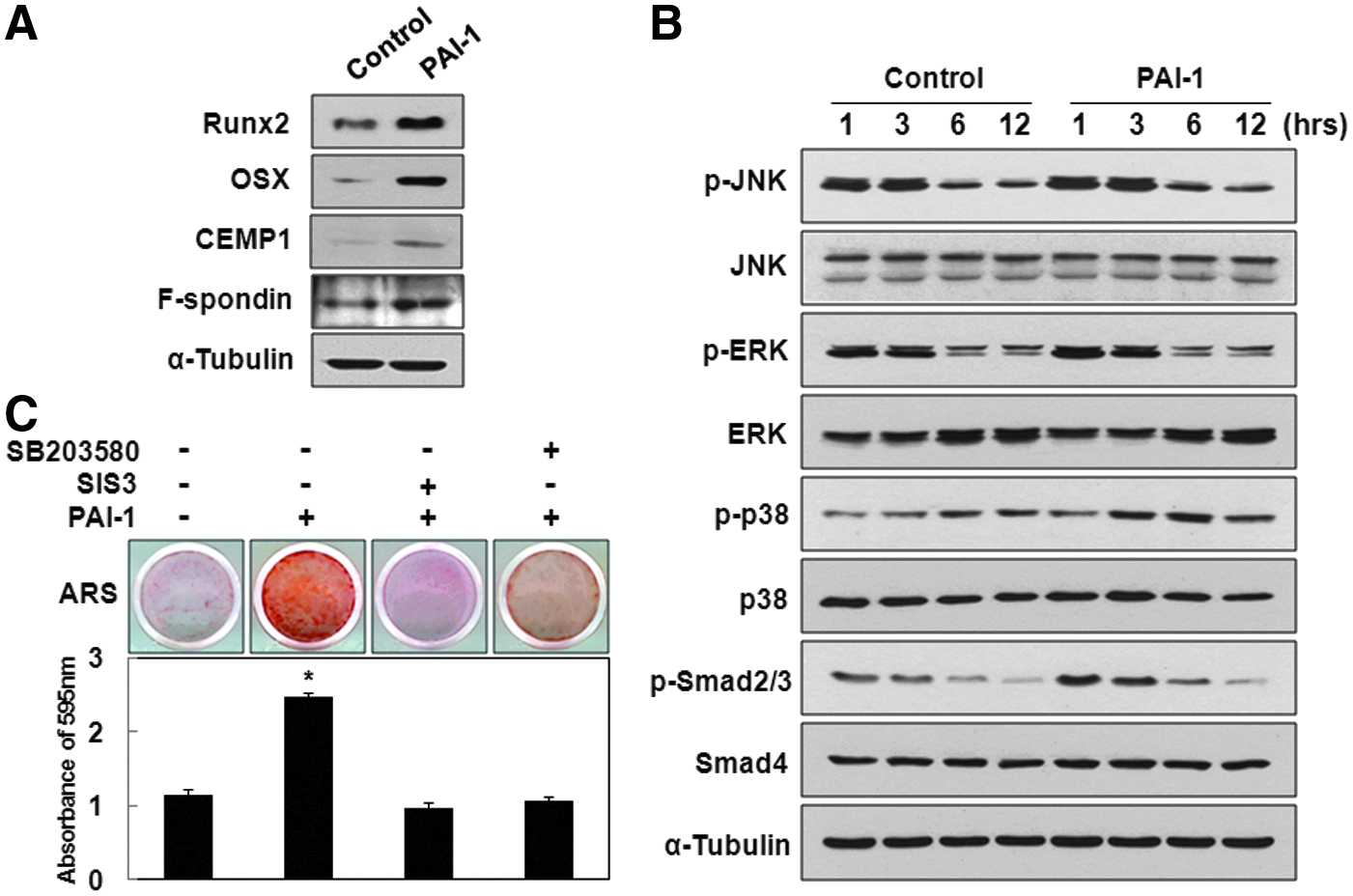
Recombinant human PAI-1-induced OSX and CEMP1 expression was mediated by p38 and Smad2/3.
Recombinant human PAI-1 promotes newly formed cementum on the surface of HA/TCP
To assess the capacity of hPDLSCs for cementum formation in response to rhPAI-1 in vivo, we aimed to reproduce the microenvironment of human tooth root as shown in Figure 4A. The hPDLSCs were mixed with HA/TCP and dentin matrix without or with rhPAI-1 and then subcutaneously transplanted into the dorsal surface of immunocompromised mice. First, we examined the area of the red box for cementum formation. At 8 weeks after transplantation, the rhPAI-1-treated group showed significant generation of new cementum-like tissue, whereas the control group generated a limited amount of cementum-like tissue on the surface of HA/TCP (Fig. 4B, C). Furthermore, we could observe a larger number of differentiated cementoblast-like cells and cementocyte-like cells in the rhPAI-1-treated group compared to the control group (Fig. 4D, E). Histomorphometric analysis showed that cementum-like tissue area in the rhPAI-1-treated group was 2.13-fold larger than that in the control group (Fig. 4F).

Histological analysis showed the effects of rhPAI-1 on cementum formation in vivo.
To further prove the effects of rhPAI-1 on cementoblast differentiation and cementum formation in hPDLSCs in vivo, we examined the expression of cementum matrix-associated proteins using immunohistochemical staining. Col1 is the most abundant collagenous protein of cementum matrix. 36 As expected, the expression of Col1 in newly formed cementum-like tissue was much stronger in the rhPAI-1-treated group than in the control group (Fig. 5A–D). During cementum formation, cementoblasts and cementocytes have been reported to produce OPN, which plays an important role in cementogenesis. 13 OPN was hardly expressed in the control group, but cementoblast-like cells and cementum-like tissue in the rhPAI-1-treated group strongly expressed OPN (Fig. 5E–H). Expression of OSX was also much stronger in the rhPAI-1-treated group than in the control group (Fig. 5I–L). Cemp1, which is regarded a specific marker of cementoblasts, was expressed more strongly in cementoblast-like cells of rhPAI-1-treated group than control group (Fig. 5M, N).

Immunohistochemical analysis showed the effects of rhPAI-1 on cementogenic differentiation of hPDLSCs in vivo.
Recombinant human PAI-1 regenerates new cementum on the dentin matrix surface
To investigate the effects of rhPAI-1 on cementogenic differentiation with human tooth root dentin matrix, we next evaluated cementogenesis in the area of the red dotted box in histological analysis. At 8 weeks after transplantation, rhPAI-1 induced hPDLSCs to differentiate into cementoblast-like cells, which arranged in a line on the surface of dentin matrix, but there was little change in the control group (Fig. 6A–D, triangles). We could observe that some of the cementoblast-like cells penetrated into the dentinal tubules on the dentin matrix (Fig. 6D, triangles). At the beginning of cementoblast maturation on the dentin surface, cementoblasts extend numerous tiny cytoplasmic processes into the loosely arranged and not yet mineralized dentinal matrix to position the initially secreted collagen fibrils of the cementum matrix. 37 It means that cementoblasts have the capacity to modify the cellular structure according to the surrounding tissue to make preparations for cementum formation, as we have found here. Furthermore, immunohistochemical staining showed strongly expressed OSX and CEMP1 in cementoblast-like cells of rhPAI-1-treated group, but hardly any expression in the control group (Fig. 6E–H). More importantly, at 10 weeks, rhPAI-1 significantly induced hPDLSCs to regenerate a thick layer of cementum-like tissue on the surface of dentin matrix, whereas we could not detect any such tissue in the control group (Fig. 6I, J). In addition, Col1 was strongly expressed in newly formed cementum-like tissue in the rhPAI-1-treated group compared to the control group (Fig. 6K, L). Taken together, these results suggest that rhPAI-1 markedly induces cementogenic differentiation of hPDLSCs, and the HA/TCP graft significantly improves periodontal tissue generation. Therefore, rhPAI-1 may solve the problem of attachment of the PDL fibers, which support tooth to resist masticatory load, in periodontitis.

Effects of rhPAI-1 on cementum regeneration on the dentin surface.
Recombinant human PAI-1 facilitates generation of PDL
PDL is the fibrous connective tissue that plays important roles in attaching tooth to the surrounding alveolar bone and absorbing heavy forces to cushion the impact. 36 At 10 weeks after transplantation, we examined whether rhPAI-1 had the ability to induce hPDLSCs to generate PDL. Picrosirius red staining showed that rhPAI-1-induced PDL fibers inserted into newly formed cementum-like tissue layer on the surface of HA/TCP and dentin matrix, whereas the PDL fibers in the control group were disorganized (Fig. 7A–D). Furthermore, higher magnification images clearly showed that the PDL fibers were almost perpendicularly inserted into newly formed cementum-like tissue in the rhPAI-1-treated group, known as Sharpey's fibers (Fig. 7F). In contrast, in the control group, the PDL fibers were orientated primarily parallel to the dentin surface (Fig. 7E). To confirm the orientation of PDL fibers, we obtained high-resolution images using SEM. The rhPAI-1-treated group showed the PDL fibers inserted into newly formed cementum-like tissue on the dentin surface (Fig. 7H, J), whereas the PDL fibers were parallel to the dentin surface in the control group (Fig. 7G, I). In addition, positive control of beagle dog tooth root showed dentin, cementum, cementoblasts, PDL fibers, and Sharpey's fibers to prove the generated periodontal tissue (Fig. 7K, L).

Recombinant human PAI-1 induced generation of PDL in vivo.
Discussion
PAI-1 is known to function as the primary physiological inhibitor of two main mammalian plasminogen activators, tPA and uPA. PAI-1 can be synthesized by various cells, including hepatocytes, adipocytes, glomerular epithelial cells, tubular epithelial cells, vascular endothelial cells, vascular smooth muscle cells, platelets, monocytes, and macrophages. 38 A potential role of PAI-1 in bone remodeling was reported when osteoblastic cells from PAI-1-deficient mice were found to display a significantly enhanced potential to degrade nonmineralized bone-like matrix. 39 However, the effects of PAI-1 on cementogenic differentiation in hPDLSCs have never been elucidated. Our present studies showed that rhPAI-1 stimulated cementogenic differentiation of hPDLSCs in vitro and in vivo. Above all, rhPAI-1 has a great ability to promote differentiation of hPDLSCs into cementoblast-like cells and generation of cementum-like tissue with inserted PDL.
Our in vitro results indicated that rhPAI-1 increased the mRNA expression levels of ALP, Col1, OPN, OCN, BSP, and Runx2 in hPDLSCs. These genes are known to be shared in osteoblasts and cementoblasts. Consistent with these results, the protein expression levels of Runx2 and OSX were also increased by rhPAI-1 treatment. It has been reported that Runx2 is an osteoblast-specific gene that regulates the initiation of osteogenesis. 40 However, the results of this study suggest that Runx2 and OSX may participate in rhPAI-1-induced cementogenic differentiation. We also found that rhPAI-1 significantly induced CEMP1 and F-spondin expression. These data suggest that some of these transcription factors might be important for cementogenic differentiation induced by rhPAI-1.
A previous report demonstrated that transplanted hPDLSCs generated new cementum and collagen fibers in immunocompromised mice. 3 However, the precise mechanisms by which hPDLSCs differentiate into cementoblasts and generate cementum when transplanted in immunocompromised mice are unclear. Based on our in vitro results, we hypothesized that hPDLSCs treated with rhPAI-1 would effectively differentiate into cementoblasts. Moreover, rhPAI-1 might have a great ability to generate cementum with inserted PDL fibers in such conditions. To test these hypotheses, histological analysis was conducted to elucidate the function of rhPAI-1 on cementogenic differentiation in hPDLSCs. Previous reports have demonstrated that xenogenic bone graft and growth factors, including transforming growth factor-beta (TGF-β), bone morphogenetic protein (BMP)-2, BMP-4, and platelet-derived growth factor (PDGF), improve periodontal tissue regeneration.41–44 MSC transplantation can supply large numbers of necessary cells and increase periodontal regenerative capacity.45,46 It also well known that MSCs can differentiate into the desired tissues, such as cementum, alveolar bone, dentin, and collagen fibers. Recently, it has been reported that CEMP1-positive progenitor cells localized in the PDL participate in the differentiation of progenitor cells for cementum formation. 47 Cementum formation is critical for appropriate maturation of the periodontium, both during development and regeneration of periodontal tissue. 36 Indeed, our in vivo studies showed that rhPAI-1 promoted hPDLSCs to differentiate into cementoblast-like cells and generate a large amount of cementum-like tissue, whereas the control group generated a limited amount of cementum-like tissue. Furthermore, several studies have reported that CEMP1 protein expression is localized to the cementoblast cell layer lining the cementoid surface of cementum.15,17 Immunohistochemical staining indicated that cementoblast-like cells strongly expressed CEMP1 in the rhPAI-1-treated group, whereas expression was weak in the control group. Col1 and OPN expression was much stronger in cementum-like tissue of the rhPAI-1-treated group than control group, and OSX expression was also significantly increased in the rhPAI-1-treated group compared to the control group. Therefore, OSX and CEMP1 might play an important role in cementogenesis in response to rhPAI-1. The results of the present study suggest that rhPAI-1 may promote cementoblast differentiation and cementum formation in hPDLSCs through the OSX-CEMP1 pathway. However, the mechanisms underlying rhPAI-1-induced cementogenic differentiation should be explored in future studies.
Previous studies indicated that HA/TCP had inducible effects on hard tissue formation of MSCs.48,49 Based on these results, many researchers attempted to generate hard tissues such as bone, dentin, and cementum by combining HA/TCP with MSCs. However, the clinical results for cementum formation were very poor because of the different structure and histological component between HA/TCP and tooth root dentin matrix. In this study, hPDLSCs were mixed with dentin matrix without or with rhPAI-1 and subcutaneously transplanted into the dorsal surface of immunocompromised mice. First, we demonstrated that rhPAI-1 induced hPDLSCs to differentiate into cementoblast-like cells. Most notably, the differentiated cementoblast-like cells were arranged in a line on the dentin matrix surface and penetrated into the dentinal tubules. Second, rhPAI-1 induced hPDLSCs to regenerate cementum-like tissue on the surface of dentin matrix in vivo. These results suggest that the morphology and property of differentiated cementoblast-like cells are similar to those of cementoblasts. Moreover, our data indicated that rhPAI-1-induced cementoblast-like cells showed strong expression of OSX and CEMP1 in hPDLSCs. Moreover, Col1 was significantly expressed on newly formed cementum-like tissue in the rhPAI-1-treated group, in conformity with the positive expression of Col1, OSX, and CEMP1 on cementoblasts and cementum of human tooth root (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/tea). Taken together, these results suggest that rhPAI-1 induces cementoblast differentiation and cementum formation on the dentin matrix surface in hPDLSCs.
In periodontal disease, PDL fibers are orientated primarily parallel to the root surface as a result of cementum resorption and, are therefore, unable to absorb masticatory load and prevent tooth loss. Our initial experiments demonstrated that rhPAI-1 had a great ability to induce cementogenic differentiation in hPDLSCs. As mentioned above, PDL generation is a very important problem in periodontal tissue regeneration as well as cementum. The importance of cementum formation is emphasized by regeneration of new cementum with inserted PDL fibers on the root surface. 36 In the present study, rhPAI-1-induced PDL fibers inserted into newly formed cementum-like tissue layer, whereas the PDL fibers were almost parallel to the root surface in the control group. SEM micrographs illustrated integration of PDL fibers with the mineralized matrices, including PDL inserts within the cementum. 50 Accordingly, we could clearly see that the PDL fibers were inserted into the newly formed cementum-like tissue layer in the rhPAI-1-treated group using SEM compared to the control group. These results suggest that rhPAI-1 not only induces cementum formation but also generates PDL fibers that insert into cementum for supportive function. However, it is still hard to regenerate tooth root, so a dental implant is generally substituted to provide the same function as tooth root. Till now, many cases reported that dental implants induce bone resorption as a result of masticatory overload. 51 This is because dental implant directly attaches to the surrounding alveolar bone in the absence of inserted PDL fibers, which normally resist forces to support tooth. In future studies, we will examine the function of rhPAI-1 and hPDLSCs on the surface of dental implants during periodontal tissue regeneration.
Conclusions
In this study, we investigated the effects of rhPAI-1 on cementogenic differentiation of hPDLSCs in vitro and in vivo. We found that rhPAI-1 induced hPDLSCs to generate cementum-like tissue with generation of PDL fibers that inserted into newly formed cementum-like tissue in vivo. rhPAI-1 might upregulate the levels of OSX and CEMP1 through Smad2/3 and p38 pathways, whereas specific inhibitors of Smad3 and p38 inhibited the enhancement of mineralization of hPDLSCs by rhPAI-1. Taken together, our data suggest that rhPAI-1 with hPDLSCs may be a good approach for future clinical applications in periodontal tissue regeneration and, furthermore, in tooth-root bioengineering.
Footnotes
Acknowledgment
This work was supported by a grant from the National Research Foundation of Korea (NRF) funded by the Korean government (MSIP) (No. 2011-0028922).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.