
Abstract
Recent advances in vascular tissue engineering have enabled a paradigm shift from ensuring short-term graft survival to focusing on long-term stability and growth potential. We present the first experimental-computational study of a tissue-engineered vascular graft (TEVG) effectively over the full lifespan of the recipient. We show that grafts implanted within the venous circulation of mice remained patent over 2 years without thrombus, stenosis, or aneurysmal dilatation. Moreover, the gross appearance and mechanical properties of the grafts evolved to be similar to the host vein within 24 weeks, with mean neovessel geometry and properties remaining unchanged thereafter despite a continued turnover of extracellular matrix. Biomechanical diversity manifested after 24 weeks, however, via two subsets of grafts despite all procedures being the same. Computational modeling and associated immunohistological analyses suggested that this diversity likely resulted from a differential ratio of collagen types I and III, with lower I to III ratios promoting grafts having a compliance similar to the native vein. We submit that TEVGs can exhibit the desired long-term mechanobiological stability; hence, we must now focus on evaluating growth potential and optimizing scaffold properties to achieve compliance matching throughout neovessel development.
Introduction
C
Mouse models provide important insight into mechanisms underlying TEVG function2,3 and are useful test beds for comparing scaffold designs.4,5 We used an interposition model in the inferior vena cava (IVC) of the mouse to evaluate, for the first time, the long-term stability of a TEVG, literally over the lifespan of the recipient. Data revealed that the TEVG evolved on average to have properties similar to the native vein within 24 weeks post-implantation, which remained stable until the end of the 2-year study. Nevertheless, a phenotypic diversity arose between 24 weeks and 2 years that manifested as subsets of “stiff” versus “compliant” neovessels. Computational models and (immuno)histological analyses suggested that these differential properties largely depended on the ratio of collagen I to III, with higher ratios resulting in stiffer neovessels. Given that long-term stability has been established in this model, there is now a need to determine mechanisms that drive biomechanical diversity, particularly given that stiffer neovessels (relative to native) would likely have less favorable overall outcomes.
Materials and Methods
Graft fabrication, implantation, and harvest
Using methods established previously,3,4 TEVGs were constructed as tubular scaffolds from sheets of nonwoven poly(glycol acid), or PGA, and sealed with a 50:50 copolymer solution composed of poly(ɛ-caprolactone and
All animal protocols were approved by the Yale University Institutional Animal Care and Use Committee. TEVGs were implanted as IVC interposition grafts in eight female CB-17 SCID/bg mice at 8 weeks of age using sterile microsurgical techniques. 3 These mice enabled direct comparisons with our prior studies; moreover, using female mice avoids the more complex surgery in males who have two testicular arteries that branch off the abdominal aorta and are difficult to isolate and dissect. Graft patency was assessed longitudinally in vivo using a high frequency ultrasound biomicroscopy system. 4 TEVGs were harvested 2 years after implantation and subjected to mechanical testing and (immuno)histological analysis.
Mechanical testing
Lengths of the centrally positioned TEVG and distal and proximal segments of adjacent IVC were measured before and following excision, which was performed from 1–2 mm below the renal bifurcation to 1–2 mm above the iliac bifurcation. The overall composite specimens were thus 5–7 mm long. The 1–2 mm long segments of adjacent IVC facilitated the requisite cannulation of the specimens with custom drawn glass micropipets. 4 The composite specimens were tested using a custom computer-controlled biaxial device designed specifically for murine vessels. 6 Tests were performed as described previously, 4 with cyclic pressure-diameter data collected from 1 to 20 mmHg at individual in vivo axial stretches for both the TEVG and adjacent proximal IVC. In addition to quantifying pressure-diameter behaviors, area compliance (ΔA/ΔP, where A is the luminal area and P the luminal pressure) was computed at successive pressures as described previously. 4
Histology and immunohistochemistry
Following testing, specimens were fixed in 10% neutral buffered formalin for 24 h, embedded in paraffin, and sectioned (4-μm thickness) serially. 3 Representative sections were analyzed within five regions: proximal IVC, proximal anastomosis, TEVG, distal anastomosis, and distal IVC. For standard histology, sections were stained with hematoxylin and eosin (H&E), Masson's trichrome (TRI), or picro-sirius red (PSR). TRI stained samples were analyzed using a custom Matlab code that quantifies pixels associated with cytoplasm and collagen. 7 A similar program was used to calculate relative distributions of small and large collagen fibers in PSR-stained sections. 5 For immunohistochemistry, sections were deparaffinized, rehydrated, and blocked for endogenous peroxidase activity and nonspecific background staining. Next, they were incubated overnight with primary antibodies: rabbit anti-collagen I (1:250; Abcam), rabbit anti-collagen III (1:250; Abcam), or rat anti-F4/80 (1:1000; Abcam). Primary antibody binding was detected using species-appropriate biotinylated secondary antibodies, followed by incubation with horseradish peroxidase streptavidin and subsequent chromogenic development with 3,3-diaminobenzidine (Vector). Nuclei were counterstained with hematoxylin. Area fractions were quantified at 10× using ImageJ software.
Data analysis and calibration
All data are presented as mean±standard error of the mean. Differences between groups were analyzed using a one-way analysis of variance (ANOVA) or paired t-tests, as appropriate. Bonferroni corrections were used to account for multiple statistical comparisons. Statistical significance was indicated by p-values <0.05.
To compare the data directly with those collected previously,
4
and given the inherent subjectivity and experimental difficulty of estimating unloaded dimensions of native veins due to their tendency to collapse at low pressure, the in vivo configuration at 2 mmHg was used as an experimental reference configuration.
8
Data from Ref.
9
were then used to calculate deformations from a previously established unloaded reference configuration to the experimental reference configuration at 2 mmHg. Mechanical response curves were calculated using the total deformation gradient,
Predictive model for in vivo neovessel development
Our growth and remodeling (G&R) approach allows a classical formulation of the wall mechanics, including equilibrium and stress constitutive relations
10
: div
Here, ρα (0) is the initial apparent mass density (i.e., constituent mass per mixture volume) of constituent α and ρ(s) is the mass density of the mixture. The three constitutive functions are as follows: mα (τ)>0, the rates of mass density production of constituent α;
Details on specific constitutive relations used to describe and predict the evolution of TEVGs in the murine venous circulation are in Ref.12,13. These relations capture the degradation of the initial polymeric scaffold and subsequent turnover of extracellular matrix, which was driven by both a foreign body inflammatory response and mechanosensing. Here, we used this G&R model to extend our prior predictions of in vivo neovessel development over 24 weeks to now describe development over the entire lifespan of the recipient. Although we first used parameters identified previously,12,13 given the variability in the experimental data from 24 weeks to 2 years, parametric studies were used to identify values that captured the observed “compliant” and “stiff” subsets of neovessels. Different parameter values were assumed to account for the potential phenotypic diversity of cells populating the neovessels and to phenomenologically account for subsequent changes in extracellular matrix due to yet unknown but inevitable differences in chemokine, cytokine, growth factor, and protease profiles.
Briefly, matrix production was modeled as
To model the loss of matrix due to basal time-, tension-, and inflammation-dependent degradation, we let fibrillar collagen degrade via first-order type kinetics:
Mechanical contributions of the polymeric scaffold and constituents of the extracellular matrix were described as usual,12,13 namely, with a neo-Hookean relation for the scaffold and amorphous extracellular matrix and Fung-type exponentials for the collagen and passive smooth muscle. It appears that the small amounts of observed elastin 4 were not functional structurally (unpublished findings; see also Ref. 5 ), hence elastin mechanics was not modeled explicitly. For each parametric study, mechanical tests were simulated and material stiffness was calculated 15 throughout the evolution of the TEVG. 13
Results
Serial ultrasound at 2, 6, 12, 24, and 104 weeks revealed that all TEVGs (n=8) remained patent without thrombosis, stenosis, or aneurysmal dilatation until harvest at 2 years (not shown). Gross examination at explantation further revealed a transformation of the TEVGs into neovessels, with a remarkable final resemblance between the grafts and native adjacent veins (Fig. 1A). Histologically assessed TEVG wall thickness significantly decreased from 24 weeks to 2 years (from 149±12.64 to 64.30±4.49 μm). Although the TEVG remained thicker than the native IVC (21.98±0.80 μm) at 2 years, this difference was not statistically significant (Fig. 1B).
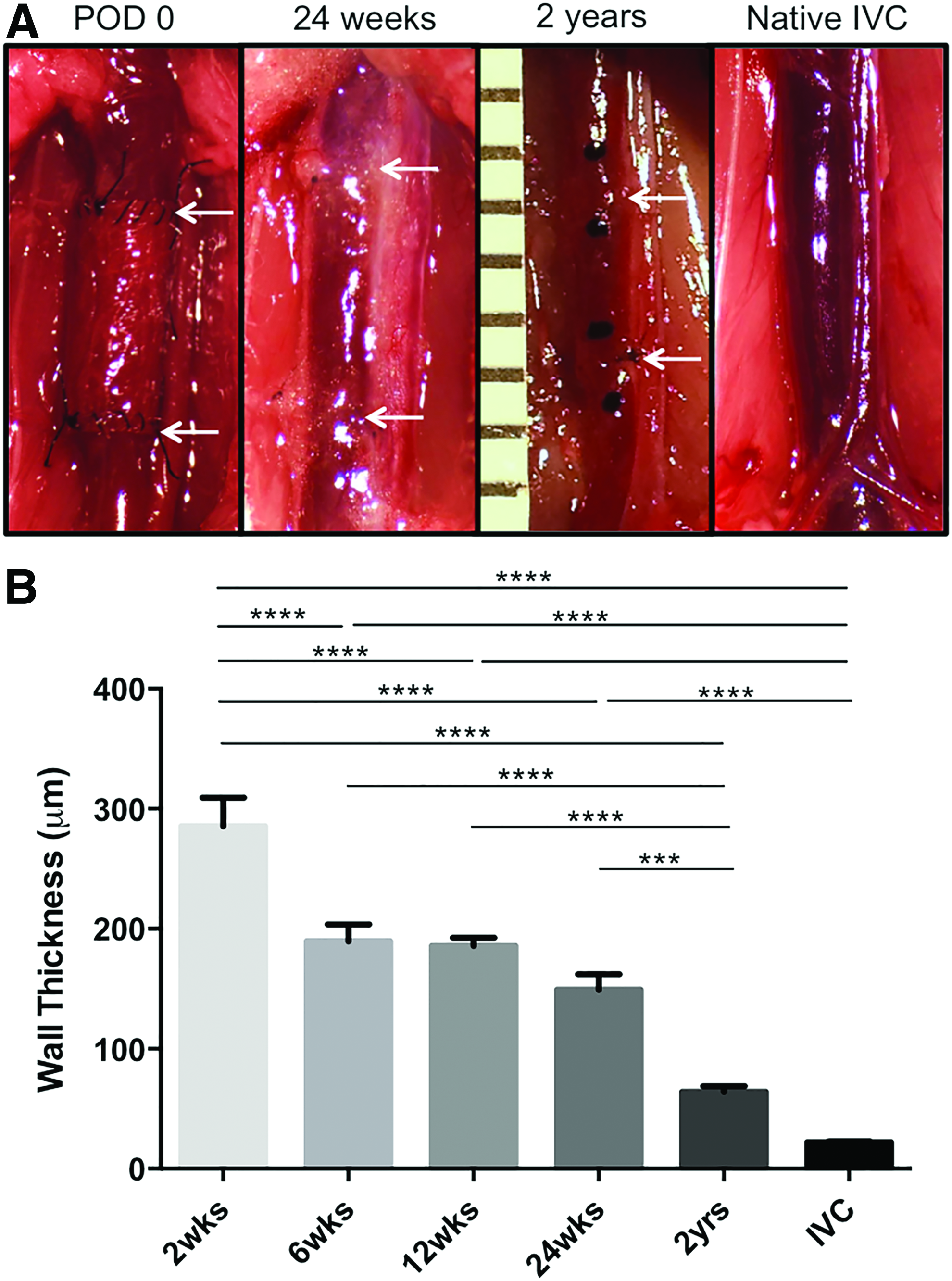
The in vivo axial stretch of the TEVG dramatically changed between 2 and 6 weeks (during which time the polymer degraded), but remained nearly unchanged thereafter. In contrast, the adjacent IVC showed a delayed decrease in axial stretch at 6 weeks that was partially, but not fully, recovered by 24 weeks. It, too, remained nearly unchanged thereafter. These findings (Fig. 2) suggested that the biaxial remodeling of both the graft and host tended toward a steady state by 24 weeks, though their early remodeling rates differed.

Changes in axial retraction upon transection revealed a biaxial development and remodeling, respectively, of the TEVGs and adjacent IVCs.
The “short-term” development of these TEVGs involves an ongoing degradation of scaffold concurrent with the deposition and remodeling of extracellular matrix. Notably, the mechanical properties of the neovessel evolve such that they are characterized less and less by the polymer-dominated stiffness, with a progressive increase in distensibility that approaches that of the native IVC at 24 weeks. 4 The present “long-term” pressure-diameter data, collected at the evolved in vivo axial lengths, revealed that the 2-year TEVGs exhibited an ∼37% increase in diameter, on average, when pressurized from 0 to 20 mmHg. In comparison, 24-week TEVGs exhibited ∼31% increase, whereas native IVCs exhibited ∼41% increase. 9 Notwithstanding the statistically insignificant differences between the 24-week and 2-year results (Fig. 3), there was greater specimen to specimen variability in distensibility at 2 years.

In vitro diameter-pressure responses showed a gradual increase in distensibility of the TEVG over the 2-year implantation. Note the transition from a stiff scaffold-dominated mechanical behavior at 2 weeks to a more distensible, neotissue-dominated mechanical behavior that resembles that of the native IVC at 2 years. Finally, note that the adjacent vein at 2 years was slightly, but not significantly, less distensible than the native IVC and thus similar to the evolved neovessel. Data for the native IVC are from Lee et al. 9 and data on TEVGs from 2 to 24 weeks from Naito et al. 4 Values are mean±SEM. Color images available online at www.liebertpub.com/tea
Figure 4 shows the area compliance of individual TEVGs explanted at 24 weeks and 2 years as a function of luminal pressure, noting again that IVC pressure is ∼2 mmHg in vivo. At this physiological value, the 2-year TEVGs exhibited an average compliance similar to both the 24-week TEVGs and native IVCs (0.20±0.14 vs. 0.19±0.03 and 0.22±0.02 mm2/mmHg, respectively). Again, however, there was greater variability in the 2-year compared with the 24-week explants, with the two subsets of neovessels exhibiting either a compliant or a stiff behavior.
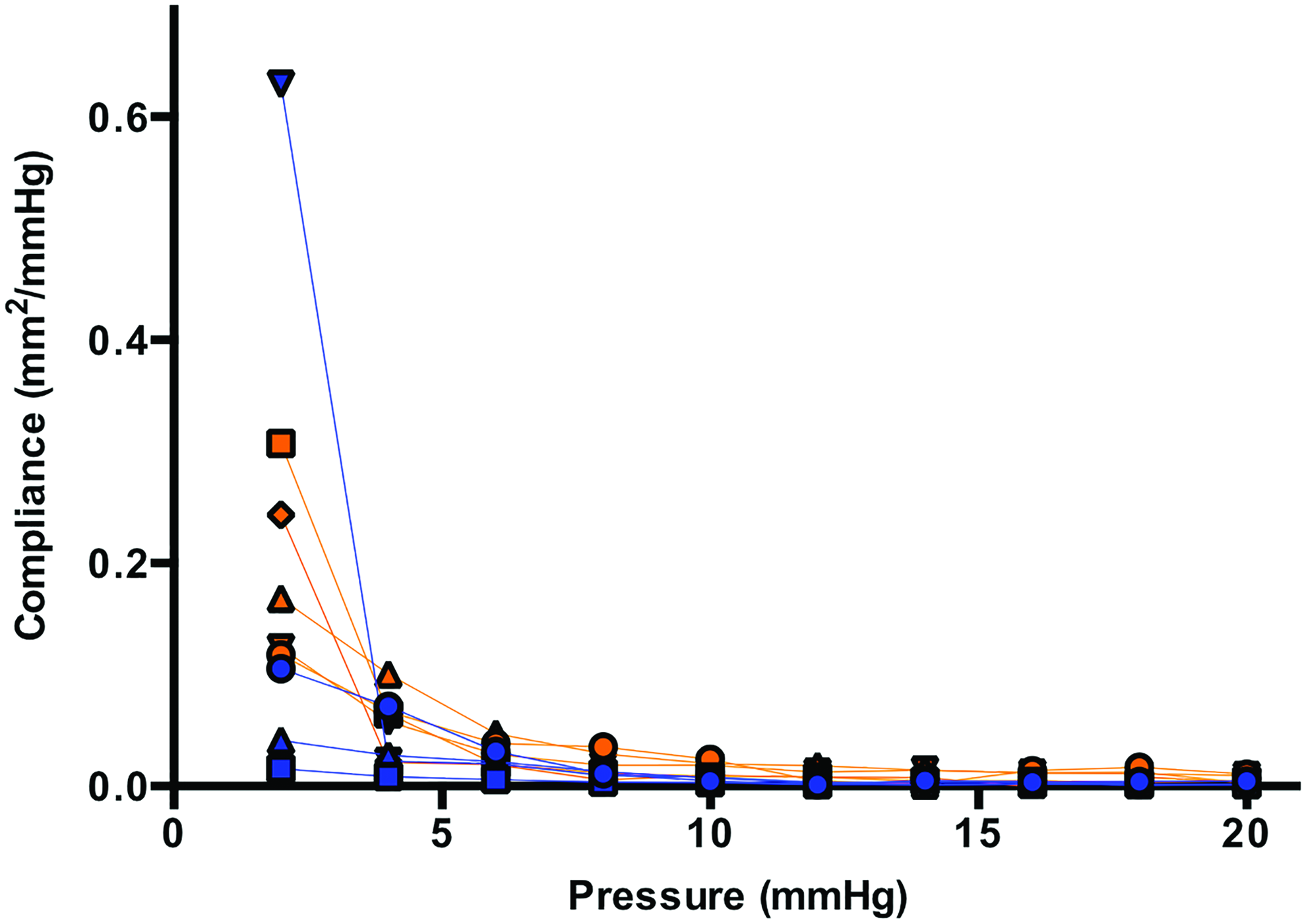
Area compliance of individual TEVGs explanted at 24 weeks (orange symbols; data from Naito et al., 4 ) and 2 years (blue symbols), each well after complete degradation of the polymeric scaffold. Note the large variability in compliance at the physiological pressure of 2 mmHg in the 2-year specimens when compared with grafts explanted after 24 weeks. The overall mean compliance of the TEVGs did not significantly differ from that of the native IVC at 24 weeks, however, and remained essentially unchanged for the remaining lifespan of the mouse. Data for the 24 week TEVGs are from Naito et al. 4 Color images available online at www.liebertpub.com/tea
Immunohistochemistry revealed negligible macrophages in the 2-year neovessels and adjacent veins, as expected. When viewed under polarized light, the PSR-stained histological sections revealed (cf. Ref. 5) a similar distribution of large (red/orange) versus small (yellow/green) fibers at the proximal and distal vein cuffs, proximal and distal anastomoses, and within the grafts, with no significant differences in area fraction between large and small fibers among the five regions (Fig. 5A). Consistent with the PSR-based finding of more large-diameter, mature collagen fibers and less small-diameter, immature collagen fibers at 2 years, immmunohistochemical analyses revealed a greater deposition of collagen I than collagen III throughout the TEVG (Fig. 5B). Also consistent with the PSR findings, there were no significant differences in overall collagen area fraction among the TEVGs, proximal and distal anastomoses, and adjacent vein cuffs.
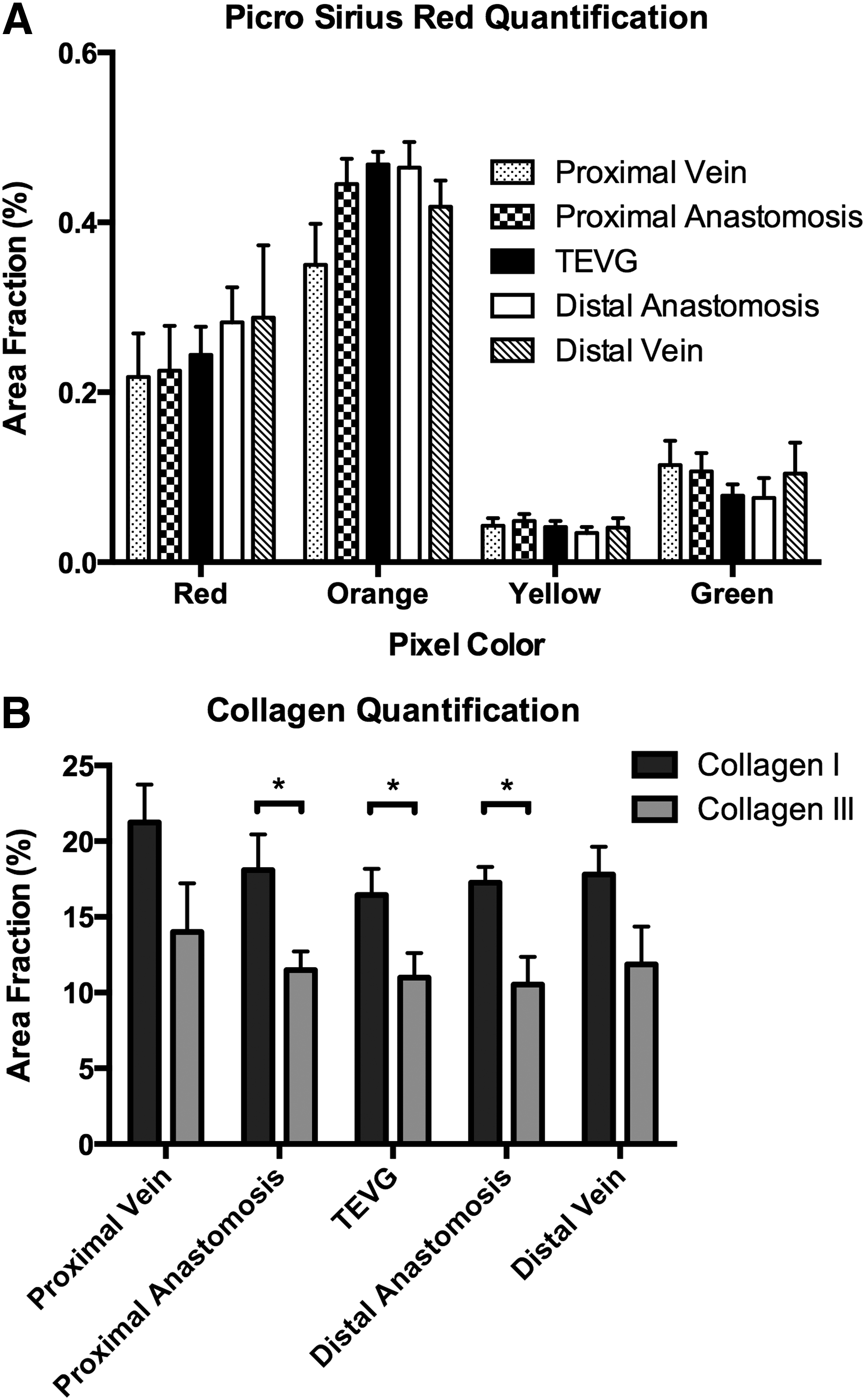
Collagen content.
Using the computational G&R model, the previously identified set of model parameters that fit the mean TEVG data up to 24 weeks postimplantation12,13 similarly predicted the mean data up to 2 years (Fig. 6, middle row), hence capturing salient features of long-term in vivo neovessel development. Yet, given that the 2-year data suggested two distinct subsets of neovessels, we also determined separate sets of parameters to capture these differences. Simply changing material properties for the collagen produced during the mechano-mediated period (e.g., γ2,strs with a slight lowering of γ1,strs; Table 1) yielded best-fit simulations for the “stiff” neovessels (Fig. 6, top row and Table 1). In contrast, multiple changes were needed to best fit the “compliant” neovessels (Fig. 6, bottom row). First, the parameters modeling the mechano-mediated collagen were decreased to γ1,strs=2.65 and γ2,strs=2.65 (Table 1); second, the half-life of constituents produced following the attenuation of the inflammatory period (when ɛ (s)=1) was reduced to

Circumferential tension-stretch behaviors for computationally simulated (dark lines) and implanted TEVGs (open symbols) at 1 day

Differences in fibrillar collagen composition appeared to determine late-term neovessel mechanical properties.
Best-fit parameters were identified to fit the expected mechanical responses for the mean data as well as upper (stiffest) and lower (most compliant) bounds for the experimental data.
Finally, we computed evolving values of biaxial material stiffness, at the individual in vivo conditions, for the three different sets of best fit model parameters: those resulting in the stiff, mean, or compliant behavior at 2 years (Fig. 8). The stiff neovessels were predicted to have the highest peak stiffness (around 200 days; phase C in the figure), which remained high over the murine lifespan. The parameters that best fit the mean TEVG behavior yielded a peak in stiffness at about the same time (phase C) that was nevertheless lower and followed by a decline to a new preferred remodeled state that was closer to the compliant than the stiff data sets. As expected, the parameters associated with the most compliant neovessels yielded the lowest stiffness at all times following polymer degradation. In particular, decreasing the half-life of the collagen produced during the inflammatory period following polymer degradation hastened the transition to the preferred remodeled state (phases C to D). Furthermore, the remodeled state of the most compliant neovessels approached that of the native IVC, shown by filled circles for comparison (Fig. 8), despite the initial polmer-dominated stiffness being the same in all cases (phase A).
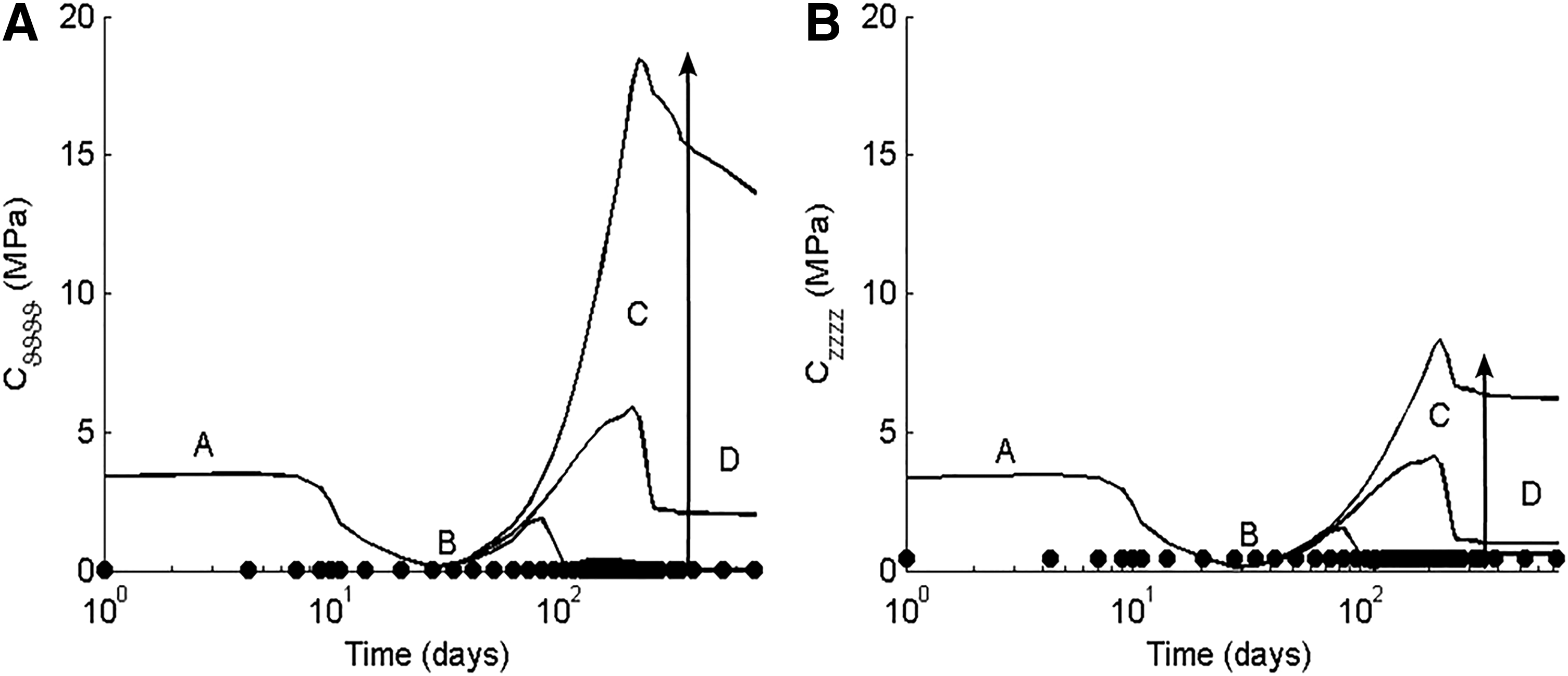
Parametric study. Values of circumferential (panel
Discussion
There were three main findings of this study. First, PGA-P(CL/LA)-based TEVGs implanted in the murine IVC reached, on average, a nearly steady state level of neovessel development by 24 weeks postimplantation that remained essentially unchanged to 2 years of age. Second, although the mean biaxial mechanical behavior of the neovessels approached that of the native IVC, significant biomechanical diversity manifested in terms of graft stiffness, particularly after 24 weeks of implantation. Third, computational modeling suggested that the observed phenotypic diversity likely resulted from differences in evolving collagen properties, which suggested the need for particular a posteriori experimental assays that indeed confirmed the model predictions. Toward this end, note the following.
First, normal vascular wall and inflammatory cells respond to changes in biomechanical environment primarily by remodeling the extracellular matrix to promote a mechanical homeostasis. 16 This process appears to extend to neovessel development, with the TEVGs and adjacent IVCs remodeling biaxially. 4 Herein, we further showed that the mean biaxial biomechanical development and remodeling of our TEVGs had essentially stabilized by 24 weeks; circumferential and axial properties remained nearly unchanged for the remainder of the murine lifespan. That is, quantification of the in vitro pressure-diameter responses of the TEVGs, at their individual evolving in vivo axial stretches, and calculation of the area compliance as a function of luminal pressure both revealed, on average, only minor increases in distensibility at 2 years relative to the 24-week explants, with an overall mean circumferential behavior that did not significantly differ from that of the native IVC. Quantification of axial retraction for both the TEVG and adjacent vein revealed a similar stability in axial behavior from 24 weeks to 2 years.
We recently introduced a computational G&R model to describe the evolution over 24 weeks of a TEVG implanted in the murine venous circulation that incorporates the monotonic loss of initial polymeric scaffold and subsequent extracellular matrix kinetics driven by both a foreign body inflammatory response and mechanosensing 12 (cf. Fig. 3). Herein, we used this model to extend the predictions from 24 weeks to 2 years postimplantation. As seen in Figure 6 (middle row), the prior model captured the subsequent mean behavior despite the continued turnover of matrix primarily due to the finite half-life of vascular collagen. 17 This finding was possible because of the long-term numerical stability of the computational model, but more importantly because of the modeled concept of homeostatic turnover of matrix under unchanging mechanical loading. 18 The experimental findings did not reveal any age-related effects on the mechanical properties of either the neovessel or the adjacent IVC, which is interesting given that mouse arteries can experience an aging-related stiffening 19 similar to that in humans. 20 Nevertheless, the current finding that there was no appreciable change in neovessel mechanical properties from 24 weeks to 2 years postimplantation is remarkable and extremely encouraging regarding the long-term mechanobiological stability of this TEVG.
Second, extreme subject to subject variability is observed and expected in the human population; it is usually ascribed to differential genetic backgrounds, ages, sexes, diets, risk factors, exercise regimes, and so forth. An unexpected observation emerging from studies using mouse models, however, is the often significant phenotypic diversity in vascular remodeling and disease progression despite the same genetic background, age, sex, diet, and environmental conditions. For example, continuous infusion of angiotensin-II in ApoE−/− mice results in lethal aortic dissections in only 10% of the animals while most others exhibit nonlethal dissections. 21 Treatment of Myh11−/− mice with L-NAME and a high salt diet results in aortic dissections in only 20% of the animals while the others exhibit no discernable phenotype. 8 In this study, we subjected female mice of the same genotype (CB-17 SCID/bg) to the “same” surgical intervention at 8 weeks of age and observed them over 2 years while maintaining them on the same diet and under the same environment conditions. Yet, even within our small group of experimental animals (n=8), two distinctly different biomechanical phenotypes emerged: stiff versus compliant neovessels at 2 years. Hence, although research has traditionally focused on mean behaviors, explicitly considering such phenotypic diversity may represent a special opportunity to use mouse models to elucidate important underling mechanisms.
Third, given the variability of the experimental observations from 24 weeks to 2 years, a series of parametric studies determined which model parameters could delineate the most versus the least distensible neovessels. We found that the long-term development of stiff neovessels could be computationally simulated by increasing the material stiffness of the collagen that was produced during the mechano-mediated period (Fig. 6, top). This phenomenological modification of collagen parameters was motivated by possible differences in collagen composition such as an increased collagen type I:III ratio or an increased number of collagen cross-links. Conversely, the development of compliant neovessels could be simulated by both decreasing the material stiffness of the collagen produced during the mechano-mediated period (i.e., a decrease in type I:III collagen ratio or decrease in cross-linking) and decreasing the half-life of the collagen produced during the inflammatory period in response to polymer degradation (Fig. 6, bottom). It is emphasized that this parameter sensitivity study/estimation was much more stringent than that usually performed in biomechanical studies (cf. Ref. 22 ). That is, parameter sensitivity studies typically focus on effects of parameter values on the material behavior at one particular time. In contrast, we sought parameters that could preserve the similar mechanical behaviors at early times and yet capture divergent behaviors at later times. Interestingly, we previously found a hint toward phenotypic diversity at 24 weeks (cf. Ref. 12 , Fig. 3), but the present 2-year experimental and computational results revealed its extent. There remains a pressing need to discover which biological or mechanical factors cause this divergent long-term behavior, which persisted over the lifespan of these mice.
We emphasize that the search for model parameters that could lead to stiff versus compliant neovessels (Table 1) was motivated solely by mechanical data (Fig. 6). It was these findings, in turn, that led to the additional histological and immunohistochemical studies to assess whether our computational predictions could be supported by experimental data. Notwithstanding the unplanned small sample sizes (n=4 for each subset of neovessels), the two independent measurements of differences in collagen composition between the stiff and compliant neovessels supported the conclusions of the computations. That is, although a larger study population will be needed for increased statistical power, neovessels that exhibited a stiffer circumferential behavior consistently had a higher ratio of large to small diameter fibers as determined by PSR colorimetric analysis and a higher ratio of collagen type I to III as revealed by immunohistochemistry (Fig. 7). There is, of course, a need to consider other aspects of the collagen organization, including possible roles of differential transmural distributions, orientations, and cross-linking of fibers in understanding fully the mechanical properties of these neovessels. Nevertheless, there are two important messages. First, computational modeling can focus experimental design and, second, there is a need to determine why otherwise similar interventions led to such phenotypic diversity, particularly since the stiff neovessels represent cases of marked compliance mismatching.
In addition to identifying underlying biological reasons for the observed phenotypic diversity, there is a need to consider mouse to mouse variations in the surgical procedure or implanted grafts that could potentially cause different outcomes. Although all surgeries were performed by the same investigator following hundreds of successful procedures, subtle differences in, for example, the axial stretch of the graft when secured by sutures at the anastomoses could affect the long-term outcome. Furthermore, minor variations in graft physical properties may have far-reaching effects on the local degradation characteristics of the polymer and its load-bearing integrity, which in turn could change the mechanical stimuli experienced by the resident cells and thereby influence local cell phenotype and matrix production. Such possibilities emphasize the importance of identifying and optimizing key physical parameters of implanted scaffolds via advances in computational modeling13,23 and of controlling fabrication techniques to minimize the differing degrees of compliance mismatch that may result secondary to the biological response following implantation.
Footnotes
Acknowledgments
This research was supported, in part, by R01-HL098228 (C.K.B.), 5T32-HL098069 (A.J. Sinusas), and NIH MSTP TG T32GM07205. The authors acknowledge Dr. Brooks Udelsman for assistance with the longitudinal ultrasound and Magdalene Yin Lin Ting for assistance with image quantification.
Disclosure Statement
C.K.B. receives grant support from Gunze Limited and Pall Corporation. The authors do not have any professional or financial conflicts of interest, however.