
Abstract
Chondrocyte-based cartilage repair techniques require control of articular chondrocyte expansion ex vivo. Articular chondrocytes have limited availability, and prolonged culturing to obtain a cell number sufficient for clinical use often results in phenotypic alterations and increased costs. In this study, we applied a screening library consisting of micrometer-sized topographical features, termed biosurface structure array (BSSA), to identify specific topographical microstructures affecting the proliferation of human chondrocytes in passage 1 (P1) or 2 (P2). The BSSA library comprised 10 patterns and 16 combinations of pillar size (X) and interpillar gap size (Y). Specific microstructures significantly increased the chondrocytes' proliferative responsiveness in term of patterns, X and Y for P2 compared with P1. The P1 and P2 chondrocytes responded independently to similar patterns after 4 days of culturing, whereas only chondrocytes at P2 responded to specific microstructures with Y = 1 μm and X = 2, 4 μm by a 2.3- and 4.4-fold increased proliferation, respectively. In conclusion, these findings indicate that specific surface topographies promote chondrocyte proliferation and may, indeed, be a tool to control the behavior of chondrocytes in vitro.
Introduction
D
Reports on micro- and nanotopographical influence on morphology, adhesion, genomic regulation, proliferation, or differentiation and a growing interest in using articular chondrocytes in tissue engineering have sparked investigation of the topographical guidance of chondrocytes during culture.8–11 To date, few topographical investigations of chondrocyte culture have been reported. It has been observed that chondrocytes are affected by surface roughness and that micrometrically grooved topographies can alter cell adhesion, cytoskeleton, and migration of chondrocytes depending on their cell passage.8,12,13 Moreover, the aggregation of primary chondrocytes is stimulated when cultured on microgrooved substrates, and hyaluronan micropatterned surfaces preserved the chondrogenic phenotype compared with polyethylene teraphthalate surfaces. 14 These studies demonstrated alteration in mature chondrocyte behavior owing to biomechanical cues.
The aim of this study was to screen in a combinatorial way the influence of distinct topographical microstructures on the proliferation of human chondrocytes in vitro. To perform this study, we used a polydimethylsiloxane (PDMS) library called biosurface structure array (BSSA) composed of 161 distinct microstructures and human articular chondrocytes harvested from the intercondylar groove in the distal femur. The BSSA library was systematically organized into 10 distinct geometric series (A–J), which were replicated with 16 unique combinations of pillar size (X) and interpillar size (Y) (Fig. 1). Distinct surface topographical parameters of this array have previously been shown to deliver biomechanical cues that enhance mineralization of a preosteoblastic murine cell line, control proliferation and osteogenic differentiation of human dental pulp-derived stem cells, and to change primary human fibroblast proliferation.9,15,16 We hypothesized that specific surface topographies may promote proliferation of human chondrocytes.
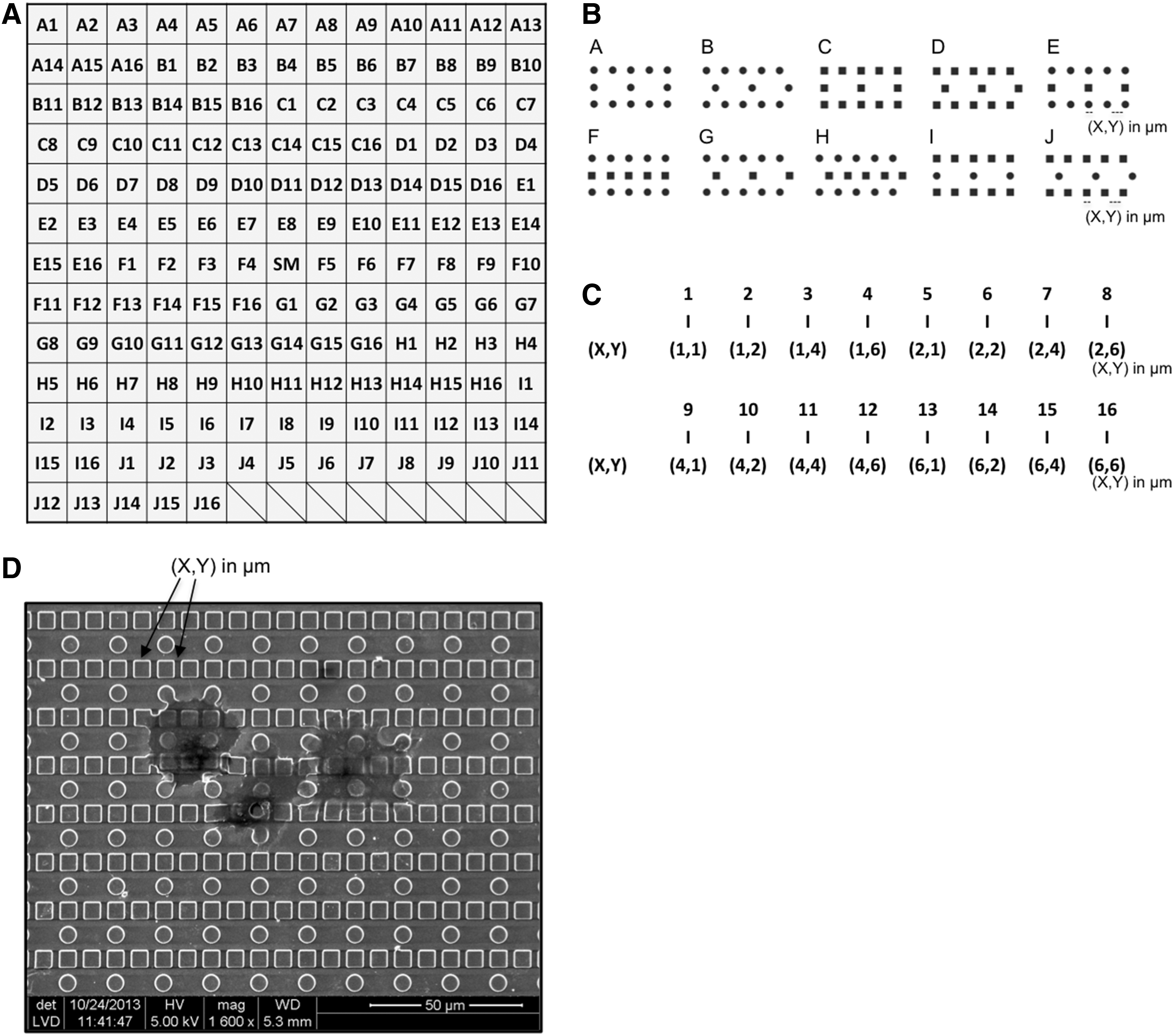
The biosurface structure array (BSSA) library design.
Materials and Methods
BSSA library design and preparation
The 2D PDMS screenings wafer, BSSA library, consisted of topographical patterns organized in 11 different geometric series (A–K), each with distinct surface microtopographies. Each topographical pattern is represented by an area of 3 × 3 mm2. Series A–J consisted of 16 unique combinations of circular and squared pillars with variable dimensions of (X, Y) pillar size (X = 1, 2, 4, 6 μm) and interpillar gap size (Y = 1, 2, 4, 6 μm). The series A-B, C-D, E-G, F-H, and I-J represented the same combination of pillar geometry but with displaced arrangement. The K structures are composed of discontinued line patterns with varying spacing and were excluded from the present study. Each library contained a smooth unstructured control (SM) placed in the middle of the library (Fig. 1).
For the preparation of the PDMS BSSA, a silicon master structure was used. The fabrication of this master has previously been described. 15 The specific study utilized a negative BSSA master, where the 160 different structures were prepared as holes with a depth of 1.6 μm. Before initiating the replica molding process, the BSSA master was fluorinated to ensure easy release of the PDMS replicas. For the fluorination process, the BSSA master was treated with ultraviolet/ozone (BioForce, Nanosciences, Inc.) for a period of 30 min before exposing the wafer to an atmosphere comprising evaporated 1H,1H,2H,2H-Perfluorooctyltriethoxysilane (Sigma-Aldrich) inside a vacuum desiccator. After 4 h, the BSSA master was removed from the desiccator and sonicated in ethanol for a period of 10 min followed by sonication in milli-Q water for an additional 10 min. Finally, the master was dried under a stream of nitrogen.
The material used for the PDMS BSSA substrates was SYLGARD® 184 (Dow Corning). Before casting the PDMS libraries, 10% (w/w) curing agent was added to the elastomer, followed by thorough mixing. Finally, the mixture was degassed to remove air bubbles using a vacuum desiccator.
For the replica molding process, the fluorinated master was placed in an aluminum container with the structured side facing upward. Subsequently, 15 g of the degassed PDMS mixture was added to the container and initial curing was performed by placing the container at 140°C for 20 min. The cured PDMS was then cut with a scalpel along the sides of the master, and the replica was carefully pealed from the master. The PDMS replica was rinsed by sonication in ethanol and subsequently in water, for periods of 10 min each, followed by drying under a stream of nitrogen. To ensure that the replicas were fully cured, they were placed in Petri dishes and heated to a temperature of 70°C for a period of 24 h. The mechanical properties of the terminally cured substrates were evaluated by tensile mechanical testing (Alwetron TCT5; Lorentzen & Wettre) and found to have an elastic modulus of 3.12 ± 0.02 MPa.
Primary human chondrocyte extraction, proliferation, and seeding on BSSA libraries
Articular cartilage biopsies were collected from the intercondylar groove in the distal femur from six healthy patients undergoing anterior cruciate ligament reconstruction. Patients' written consent was obtained, and the protocol was approved by the Local Ethical Committee under the Danish National Committee on Research Ethics (#M-2008-008). Cartilage biopsies (n = 3 for each passage) were collected in phosphate-buffered saline (PBS) with 10% fetal bovine serum (FBS) and 1:100 penicillin–streptomycin (Sigma-Aldrich). Each cartilage biopsy was minced into smaller pieces and enzymatically digested with 0.15% collagenase II (Invitrogen, Life Technologies) in Dulbecco's modified Eagle's medium/F12 (DMEM/F12) (GlutaMAX; Gibco, Life Technologies), with 10% FBS for 20–22 h at 37°C. Isolated chondrocytes were seeded in a 25 cm2 culture flask using DMEM/F12 supplemented with 10% FBS, 1 ng/mL transforming growth factor-β3 (TGF-β3; R&D Systems), 5 ng/mL basic fibroblast growth factor (bFGF; Gibco), and 1:100 penicillin–streptomycin and cultured at 37°C in a humidified atmosphere of 5% CO2 until confluence.17,18 The chondrocytes were either used in passage 1 (P1) or reseeded in a 25 cm2 culture flask to be used in P2. Immediately before cell seeding, the BSSA libraries were sterilized by soaking in 70% ethanol for 30 min and rinsing thrice in PBS for 5 min. Following sterilization, the BSSA libraries were coated with 0.5 μg/cm2 fibronectin (Sigma-Aldrich). Chondrocytes, P1 or P2, were seeded on the BSSA libraries at 10,000 cells/cm2 in DMEM/F12 supplemented with 10% FBS and 1:100 penicillin–streptomycin. Dishes were carefully transferred to the incubator, at 37°C, in a humidified atmosphere of 5% CO2.
DNA staining of chondrocyte proliferation (cell number)
After 1 and 4 days of cultivation on the BSSA, chondrocytes were rinsed in PBS, fixed in 70% ethanol, and permeabilized with 0.1% Triton X-100, 50 mM Tris-HCl, 150 mM NaCl, 5 mM EDTA, gelatin 0.1%, and 0.05% P-40. The chondrocytes were incubated with DRAQ5™ (Biostatus Limited) 1:2000.19,20 The BSSA libraries were rinsed in PBS and left for drying for at least 24 h at 4°C. Quantitative detection of cell number was performed using Odyssey® SA far-red Imaging (LI-COR Biosciences) at 700 nm and Image Studio Software version 2.1.10 (LI-COR Biosciences).
Filamentous actin stain
Chondrocytes cultured for 2 days were fixed in 4% formaldehyde (VWR International Ltd.), permeabilized with 0.5% Triton X-100 in PBS, blocked with 1% bovine serum albumin (BSA; Sigma-Aldrich), and incubated with Alexa Fluor 488® phalloidin at 1:100 (Life Technologies) in 1% BSA for 1 h at room temperature. Chondrocytes were rinsed in PBS and counterstained with 4′,6-diamidino-2-phenylindole (DAPI) at 1:1000 (Life Technologies) in 1% BSA. Fluorescence was visualized with Olympus BX50 fluorescence microscopy (Olympus Denmark A/S).
Extracellular matrix—proteoglycans
Chondrocytes cultured for 2 days were rinsed with PBS, fixed with cold 4% formaldehyde, and stained with alcian blue solution pH 2.6 (Sigma-Aldrich) overnight. Excess staining was removed, and the BSSA library was washed with 3% glacial acetic acid (MERCK KGaA), followed by a rinse in PBS. Images were obtained with Motic AE21 inverted microscopy.
Total RNA extraction and quantitative RT-PCR
Total RNA was extracted using TaqMan® Fast Cells-to-Ct™ (Ambion; Life Technologies), according to the manufacturer's instructions. Before total RNA extraction, each 3 × 3-mm2 sample was cut out and washed in PBS. Each sample was lysed, and reverse transcriptase polymerase chain reaction (PCR) was performed on the cell lysate, which was transcribed into cDNA using TaqMan Fast Cells-to-Ct (Ambion). Quantitative real-time RT-PCR was performed on a 7500 Fast Real-Time PCR system (Applied Biosystems) using TaqMan Fast Universal PCR Master Mix (Applied Biosystems) and TaqMan Gene Expression Assays (Applied Biosystems) with the following assay: sex-determining region Y box 9 (SOX9) Hs00165814_m1. Standard enzyme and cycling conditions for the 7500 Fast System were used. Each biological sample was run in technical duplicates for each gene. Data analysis was performed using 7500 Fast System Sequence Detection Software version 3.1 (Applied Biosystems). Target gene expression was normalized to the housekeeping genes beta-2-microglobulin (B2M) Hs99999907_m1 and ribosomal protein L13a (RPL13a) Hs03043885_g1 based on BestKeeper values.17,21
Statistics
For comparisons of proliferative data, normality was checked using Q-Q plots and statistical analysis was performed using three-way ANOVA (pattern*X*Y), with p-values <0.05 considered significant. For data concerning gene expression, one-way ANOVA with p-values <0.05 were considered significant. Data are presented as mean ± standard error of the mean of three independent experiments in triplicates (n = 3). Stata Statistical Software: release 13.1 (StataCorp LP) was used.
Results
Far-red DNA staining and cell proliferation
Validation of attachment point was based on population doubling analyses showing no detectable proliferation within the first 24 h of culture (data not shown). Day 1 was therefore selected as the attachment point for this experimental setup. Proliferation was assessed as cell number after 4 days of culture subtracted the cell number on day 1 (attachment).
Seeding and distribution of the chondrocytes onto the BSSA library were visualized using far-red DNA staining after 1 and 4 days of culture (Fig. 2A, B). On day 4, the screening of the 160 different topographies revealed specific microstructures with higher DNA intensity staining (Fig. 2B).

Far-red DNA imaging of chondrocytes on the BSSA library.
To investigate the effect of dimensions (X, Y), data with varying patterns and the same X and Y were clustered. The first screening analysis was made of the attachment point for chondrocytes at P1. It revealed no significant attachment preferences with regard to the different series A–J, X, or Y (Fig. 3A, E). The second screening for proliferation of chondrocytes at P1 after 4 days of culture identified a significant effect of patterns (p < 0.0005). This is depicted in Figure 3B by an ordered orientation of the 10 different pattern series A–J compared with the previous screening in Figure 3A. X or Y had no significant effect on the proliferation (Fig. 3F). The third screening of attachment point for chondrocytes at P2 revealed a significant effect of the dimensions X and Y (p < 0.0007) (Fig. 3G), which suggests an adapted attachment for X and Y for chondrocytes at higher passages. The patterns had no significant effect at this time point (Fig. 3C). The fourth screening of proliferation of chondrocytes at P2 after 4 days of culture identified a significant effect of both patterns (p < 0.0001) and dimensions X and Y (p < 0.001) (Fig. 3D, H). The pattern series I and J had a higher proliferative effect than the pattern series A and B (Fig. 3D). Furthermore, a clear systematic effect of Y, and to a lesser extent X, significantly influenced the proliferation (Fig. 3H). Proliferation at the microstructure dimensions X = 2, 4 μm and Y = 1 μm was increased by 2.3- and 4.4-fold, respectively, compared with the smooth unstructured control (SM) microstructure, while dimension X = 2, 4 μm and Y = 6 μm were only increased 0.8- and 1.4-fold, respectively. All 16 dimensions increased the proliferation compared with SM, which indicates an effect of exterior stimulation of proliferation at this time point. A subset of dimensions, X = 2, 4 μm and Y = 1, 6 μm in the pattern series H and I with chondrocytes at P2, was selected for further analyses owing to the high and low proliferative effect.
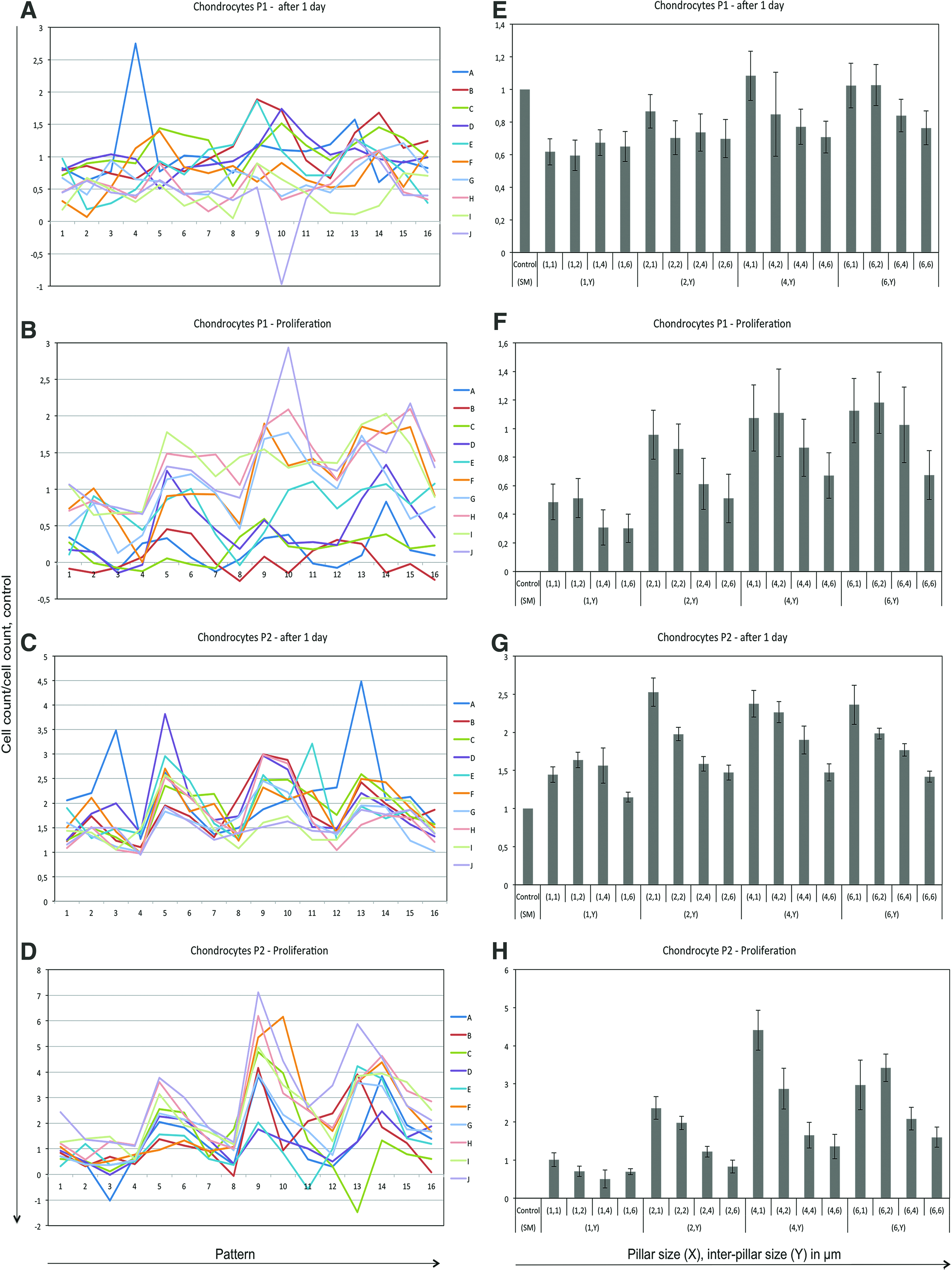
Validation of proliferation over time based on cell number on P1 and P2 chondrocyte. Attachment point on day 1
F-actin stain of cell morphology and cell number
The morphology structure of the actin filaments of chondrocytes at P2 cultured for 2 days on topographical dimension, X = 2, 4 μm, Y = 1, 6 μm, and SM were visualized (Fig. 4). On these five selected microstructures, chondrocytes had thick actin stress fibrils. We observed a minor morphological difference between chondrocytes cultured on microtopography compared with SM: the cytoskeleton of chondrocytes on X = 2, 4 μm and Y = 1, 6 μm appeared more spread out and had a polygonal morphology with multiple filopodia per cell, whereas the chondrocytes cultured on the SM were more elongated. The results also indicated a difference in the number of chondrocytes per microstructure. At dimension X = 2, 4 μm and Y = 1 μm, the number of chondrocytes seemed to be increased compared with X = 2, 4 μm and Y = 6 μm. At X = 2, 4 μm and Y = 1, 6 μm, the number of chondrocytes seemed to be increased compared with SM. These results supported the findings of a positive proliferative effect of these dimensions shown in Figure 3H.
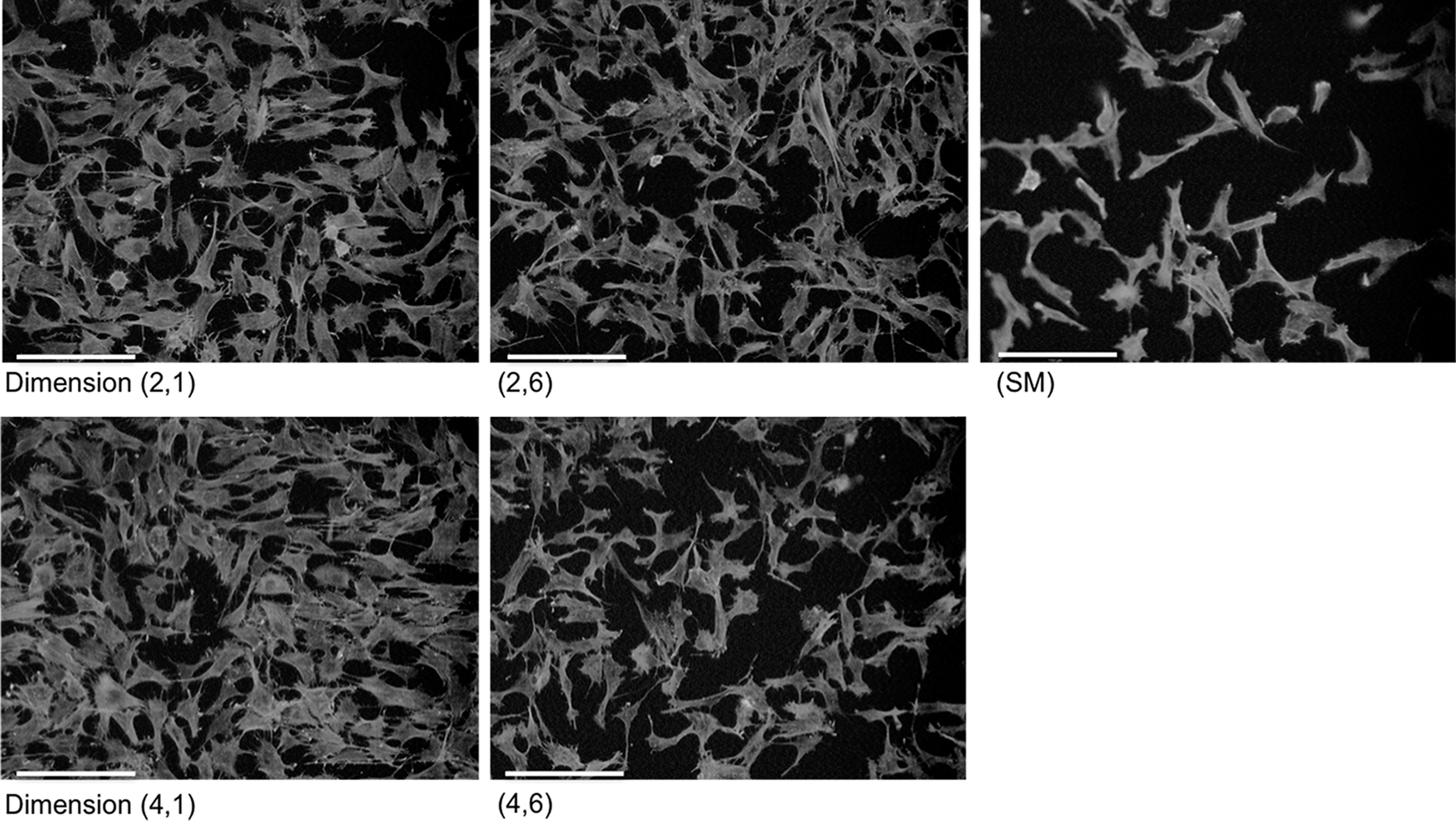
Fluorescent F-actin stain of P2 chondrocytes cultured on dimension X = 2, 4 μm, Y = 1, 6 μm, and SM for 2 days on series I. Original images of green staining were converted into grayscale for better visualization. Scale bar = 150 μm.
Projected pillar area
The cell number was plotted as a function of the relative projected area of the pillars (area on top of the pillars as a fraction of the total unit area) of the various microstructures. This plotting was performed to investigate to which extent the simple geometric pillar surface effect influences the proliferation. We observed a trend toward an increase in cell number (proliferation) as the projected pillar area increases up until the maximum value of ∼0.5, which was independent of the specific geometry A–J (Fig. 5).

Cell number of P2 chondrocytes as a function of the projected pillar area. Cell number was plotted as a function of the relative projected area of the pillars (area on top of the pillars as a fraction of the total unit area) of the various microstructures. Different series were combined on similar patterns and same pillar surface coverage; series (A+B), (C+D), (E+G), (F+H), and (I+J). The individual graphs illustrate the degree of proliferation after 4 days for a specific pillar size, X, of 1 μm, 2 μm, 4 μm, and 6 μm over a range of interpillar gap sizes, Y, of 1, 2, 4, and 6 μm.
Proteoglycan deposition and SOX9 expression of passage 2 chondrocytes
To investigate the influence of rapid proliferation on P2 chondrocytes cultured for 4 days on pattern H with topographical dimension, X = 2, 4 μm and Y = 1, 6 μm, and SM primary chondrogenic features were considered. Although the chondrocytes were cultured in monolayer, qualitative deposition of proteoglycan was visible in each of the five dimensions (Fig. 6A) and there were no significant differences in the relative quantitative measure of chondrocyte transcription factor SOX9 gene expressions (Fig. 6B).

Proteoglycan deposition and SOX9 expression of passage 2 chondrocytes.
Discussion
Our study has provided novel results showing that specific topographical microstructures significantly promote human chondrocyte proliferation. In particular, we found that with an increasing passage number, chondrocytes became more responsive to the microstructures in terms of patterns and dimensions. Previous studies have shown that surface topography biomechanical cues affect the behavior of many mammalian cell types at a molecular level.22–24 The 2.3- and 4.4-fold increase in chondrocyte proliferation compared with smooth standard surface (SM) suggests that the pillar sizes X = 2, 4 μm and interpillar gap size Y = 1 μm are important key players in the higher proliferation. Another important finding was that independent of their passage, chondrocytes responded to the same specific pattern series with increased proliferation (e.g., H, I, and J compared to A, B, C, and D). Regarding the relative projected pillar area, we observed a trend toward higher proliferation of P2 chondrocytes with increasing relative projected area of the pillars; this trend was independent of the detailed microstructure pattern. This points toward a strategy for increasing the relative pillar area that should involve increased roughness to topographically functionalize cell substrates for chondrocyte proliferation.
As the proliferative responsiveness of chondrocytes to topography is influenced by passage and culture time, it is, therefore, tempting to hypothesize that higher passage chondrocytes are more responsive to biomechanical cues than early cultured chondrocytes. The validity of our proliferation for P2 chondrocytes was supported by qualitative illustration of F-actin staining, where the chondrocyte number increased depending on the combination of X and Y. A previous study supports these findings by also showing that specific X and Y influenced fibroblast proliferation. 16 However, no plausible hypotheses presently explain the mechanism involved in the cell–topography interaction on proliferation. 11
For the selected subset of dimensions, X = 2, 4 μm and Y = 1, 6 μm representing high and low proliferative effect of P2 chondrocytes, proliferation varied while differentiation remained rather similar. High proliferation in P2 chondrocytes may have led to a more dedifferentiated stage. However, the chondrocytes in P2 were able to deposit visible proteoglycans. This observation was also supported by the expression of SOX9, a chondrocyte transcription factor essential for chondrogenic differentiation, where no significant differences between the five selected microstructures (including smooth surface control) were observed.25,26 Thus, this observation also implies that the increased proliferation in general did not result in reduced gene expression of SOX9 compared with smooth standard surface SM, which indicates that P2 chondrocytes on the specific microstructure still express and hold the potential of a chondrogenic phenotype. Collectively, our data show that specific microstructures increase the proliferation of human chondrocytes without deviating their differentiated state comparable with SM.
Mimicking the in vivo natural milieu is an essential principle in cell culturing, and one major contributor to such culturing is growth factors. Expansion of chondrocytes is currently performed in monolayer cultures where proliferation is stimulated by adding growth factors (i.e., bFGF, TGF-β1 or -β3, and insulin-like growth factor-1 [IGF-1]).27–29 These agents are costly, and a reproducible culture milieu depends largely on manufactures, batch number, and expiration of the product. In the present study, no growth factors were added when culturing chondrocytes on the BSSA library. Hence, the only stimulator of proliferation was the detailed microstructure. To which extent microstructure roughness functions as a proliferation cue is difficult to determine at this moment. The present study proposes an alternative strategy to the traditional monolayer expansion culture through tailored polymer surfaces composed of specific patterns, pillars, and gaps. The production methodology of the culture surface provides for an easily standardized large-scale production compared with the higher variability in medium preparation containing multiple agents such as growth factors.
The main focus of this article was screening of chondrocyte proliferation, and we did not address hypotheses regarding the effect of attachment and adhesion processes and detailed differential mechanisms on the topography, nor did we address the role of topography in chondrogenic differentiation. Technically, the combinatorial library design and small unit area of 3 × 3 mm2 per microstructure reduced the opportunities for analysis regarding chondrogenicity, substantial phenotypic evaluation, and any potential protein signaling mechanisms involved. Further investigations must be performed to elucidate the obvious trend on proliferation where focus is on larger microstructured culture areas and prolonged culture time to determine the potential effects for culture of chondrocytes on topography intended for cell-based cartilage repair.
In conclusion, this study demonstrated that specific topographical microstructures increased the proliferation of human chondrocytes. Specifically, the responsiveness of the chondrocyte proliferation to pattern, pillar size, and interpillar gap size increased significantly as the passage of the chondrocytes was higher. The increased proliferation did not lead to deviate differentiation as evaluated by proteoglycan deposition and SOX9 gene expression compared with the smooth surface. These findings indicate that specific surface topographies promote chondrocyte proliferation and may prove to be an important tool to controlling chondrocyte behavior in vitro. Hence, we suggest that tailored surface topography is a simple alternative and a reproducible method for ex vivo expansion of primary human chondrocytes for practical and clinical cell-based cartilage repair strategies.
Footnotes
Acknowledgments
The authors gratefully thank The Danish Rheumatism association for the financial support, Grant No. R106-A2368 and R116-A2791, the Interdisciplinary Nanoscience Centre (iNANO), Aarhus University, Denmark, for producing the BSSA library wafers, and the excellent technical assistance provided by Anna Bay Nielsen, Orthopedic Research Laboratory, Aarhus University Hospital, Denmark.
Disclosure Statement
No competing financial interests exist.