
Abstract
Although hydrostatic pressure (HP) is known to regulate chondrogenic differentiation of mesenchymal stromal/stem cells (MSCs), improved insight into the mechanotransduction of HP may form the basis for novel tissue engineering strategies. Previously, we demonstrated that matrix stiffness and calcium ion (Ca++) mobility regulate the mechanotransduction of HP; however, the mechanisms, by which these Ca++ signaling pathways are initiated, are currently unknown. The purinergic pathway, in which adenosine triphosphate (ATP) is released and activates P-receptors to initiate Ca++ signaling, plays a key role in the mechanotransduction of compression, but has yet to be investigated with regard to HP. Therefore, the objective of this study was to investigate the interplay between purinergic signaling, matrix stiffness, and the chondrogenic response of MSCs to HP. Porcine bone marrow-derived MSCs were seeded into soft or stiff agarose hydrogels and subjected to HP (10 MPa at 1 Hz for 4 h/d for 21 days) or kept in free swelling conditions. Stiff constructs were incubated with pharmacological inhibitors of extracellular ATP, P2 receptors, or hemichannels, or without any inhibitors as a control. As with other loading modalities, HP significantly increased ATP release in the control group; however, inhibition of hemichannels completely abrogated this response. The increase in sulfated glycosaminoglycan (sGAG) synthesis and vimentin reorganization observed in the control group in response to HP was suppressed in the presence of all three inhibitors, suggesting that purinergic signaling is involved in the mechanoresponse of MSCs to HP. Interestingly, ATP was released from both soft and stiff hydrogels in response to HP, but HP only enhanced chondrogenesis in the stiff hydrogels, indicating that matrix stiffness may act downstream of purinergic signaling to regulate the mechanoresponse of MSCs to HP. Addition of exogenous ATP did not replicate the effects of HP on chondrogenesis, suggesting that mechanisms other than purinergic signaling also regulate the response of MSCs to HP.
Introduction
T
Previously, calcium signaling has appeared necessary for the mechanotransduction of various loading modalities such as tension, compression, and fluid flow.14–23 Recently, we demonstrated that calcium ion (Ca++) mobility is required for the mechanotransduction of HP by MSCs. 24 In particular, both voltage-gated calcium channels (VGCCs) and Ca++ release from intracellular calcium stores were found to play key roles in the chondrogenic response of MSCs to HP. 24 However, how these Ca++ signaling pathways are initiated remains unclear. One possible mechanism is the purinergic pathway, in which adenosine triphosphate (ATP) is released from the cell in response to mechanical signals and then acts as a paracrine/autocrine factor to increase intracellular Ca++ concentrations (Fig. 1). Compression of articular chondrocytes was found to release ATP into the pericellular environment, and hemichannels have been proposed as the conduit, through which ATP exits the cell.25–28 Hemichannels, which combine to form gap junctions, are composed of connexin subunits and allow nonselective permeability to molecules smaller than 1 kDa. 29 Once ATP is released, it binds to cell-surface purine receptors (P-receptors), which, depending on the specific P-receptor type, triggers a Ca++ signaling cascade using either direct or indirect methods. P2X receptors act as ATP-gated ion channels, while P2Y receptors are coupled to G-proteins and elicit a Ca++ response through inositol triphosphate (IP3)-mediated release from the sarcoendoplasmic reticulum calcium stores (SERCS) (Fig. 1). 30 Once Ca++ enters the cell, it can initiate a variety of signaling pathways, including an amplification of the Ca++ signal through activation of VGCCs and through calcium-induced Ca++ release from intracellular calcium stores.

Elements of the purinergic signaling pathway and their respective pharmacological inhibitors. Color images available online at www.liebertpub.com/tea
Previously, we demonstrated that matrix stiffness regulates the mechanotransduction of HP through cell–matrix interactions.6,7 MSCs were seeded in agarose hydrogels with varying stiffness; the pericellular matrix was more developed, cytoskeletal organization was altered, and focal adhesions were increased in the stiff gels relative to the soft gels, with MSCs in the soft gels exhibiting a stronger chondrogenic phenotype. However, the pro-chondrogenic response to HP was only observed in the stiffer gels. 7 Furthermore, this mechanoresponse was completely suppressed with the addition of soluble RGDS peptide, an integrin blocker. Together with other studies, these data suggest that cellular interactions with a well-developed pericellular matrix are crucial to HP mechanotransduction, particularly in hydrogel scaffolds such as agarose.6,7
The first objective of the current study was to investigate whether purinergic signaling plays a role in the chondrogenic response of MSCs to HP by utilizing pharmacological inhibitors to block various components of the purinergic pathway. Based on our previous studies investigating the influence of matrix stiffness on the mechanotransduction of HP 7 , the current study further aimed to examine the interplay between matrix stiffness and purinergic signaling by investigating the release of ATP in soft and stiff hydrogels exposed to HP. In addition, the upregulation of chondrogenic markers through the mechanotransduction of HP has been associated with an altered vimentin architecture, and inhibition of HP mechanotransduction also blocks vimentin reorganization.7,24 Therefore, the interplay between purinergic signaling and HP-mediated vimentin remodeling was investigated. Finally, we sought to determine whether the chondrogenic mechanoresponse to HP could be mimicked by stimulating the purinergic pathway with the addition of exogenous ATP.
Materials and Methods
Materials
Porcine femurs were procured from a local abattoir, Martin's Custom Butchering. Dulbecco's modified Eagle's medium, insulin-transferrin-selenium, Alexa Fluor® 568 Donkey Anti-Goat IgG Antibody, and the PicoGreen assay were purchased from Life Technologies. Fetal bovine serum (FBS) was purchased from Omega Scientific. Penicillin-streptomycin was purchased from Corning, VWR. Agarose, media supplements, purinergic inhibitors, luciferin-luciferase assay, anti-Mouse IgG biotin conjugate, monoclonal anti-vinculin FITC conjugate, and all other chemicals (unless otherwise noted) were purchased from Sigma-Aldrich. Transforming growth factor-β3 (TGF-β3) was purchased from PeproTech. Sodium pyruvate was purchased from Lonza, VWR. Ethanol was purchased from Acros Organics, VWR. Guanidine-HCl was purchased from Calbiochem, VWR. Bovine serum albumin (BSA) and paraformaldehyde were purchased from Fisher Scientific. The pressure vessel was purchased from Parr Instrument Company. Isopropanol was purchased from Alfa Aesar, VWR. The primary antibody for immunohistochemistry was purchased from Abcam. The ABC reagent and DAB peroxidase were purchased from Thermo Scientific, VWR. Anti-vimentin FITC and anti-alpha tubulin eFluor® 615 were purchased from eBioscience, Inc. The P2Y2 Antibody (A-20) was purchased from Santa Cruz Biotechnology.
Cell isolation, expansion, and encapsulation
Porcine bone marrow was harvested from the femoral diaphysis of pigs (∼50 kg) under sterile conditions. MSCs were isolated and expanded according to a modified method developed for human MSCs.
31
Cultures were expanded in hgDMEM GlutaMAX supplemented with 10% FBS and penicillin (100 U/mL)-streptomycin (100 μg/mL). After expansion (third passage), MSCs were encapsulated in agarose (Type VII) hydrogels at a density of 15 × 106 cells/mL. MSCs were mixed with agarose at ∼40°C to yield a final gel concentration of either 1% (soft) or 4% (stiff). The agarose-cell suspension was cast in a stainless steel mold and cored using a biopsy punch to produce cylindrical scaffolds (Ø 5 × 3 mm thickness). Constructs were maintained in 2.5 mL/construct of a chemically defined media (CDM) consisting of hgDMEM GlutaMAX supplemented with penicillin (100 U/mL)-streptomycin (100 μg/mL), 100 μg/mL sodium pyruvate, 40 μg/mL
Application of HP
Constructs were sealed into sterile bags with 1.5 mL of medium per construct during the daily loading period. After loading, constructs were removed from bags and returned to culture dishes containing 2.5 mL of medium per construct to allow gases to equilibrate overnight. Cyclic HP was applied in a custom bioreactor filled with water within a 37°C water bath as described previously. 5 The sealed bags exposed to HP were placed into the pressure vessel, while the FS specimens were placed in a separate pressure vessel, which was immersed in the same temperature-regulated water bath, but was not connected to the mechanical testing system. The pressure vessel was connected to a hydraulic cylinder that was loaded using a computer controlled Instron 88215 materials testing machine. The pressure inside the vessel was measured using a pressure gauge. The load applied to the hydraulic cylinder by the Instron was set such that the HP inside the vessel reached an amplitude of 10 MPa at a frequency of 1 Hz, 4 h/d, 5 d/week for 3 weeks. Half-medium exchanges were performed biweekly to replenish nutrients and pharmacological inhibitors, and media samples were collected and stored at −80°C for biochemical analysis. The constructs were cultured with the relevant pharmacological inhibitor (or without inhibitors in the control groups) for the duration of the experiment.
ATP release assay
On days 7, 14, and 21, the stiff gels (both the Ctrl and -Hemi groups) and soft gels were loaded for 15 min. Media samples were then obtained and immediately boiled for 1 min to inactivate any ATPases. The samples were then analyzed with a commercially available Luciferin-Luciferase Assay Kit and quantified using a luminescence reader (n = 9). The samples were then loaded for an additional 4 h and the assay was repeated as described.
Biochemical analysis
On day 21, constructs (n = 4) were digested with papain (125 μg/mL) in 0.1 M sodium acetate, 5 mM
Confocal microscopy and immunohistochemistry
At day 21, constructs (n = 2) were cut in half and fixed in 4% paraformaldehyde overnight at 4°C and rinsed with PBS. To examine focal adhesion formation and cytoskeletal organization, samples were permeabilized in a 1% Triton-X and 2% BSA solution for 45 min and washed in PBS. The samples were then incubated in a 1.5% BSA solution containing one of 60 μg/mL monoclonal anti-vinculin FITC conjugate, 5 U/mL rhodamine phalloidin, 1 μg/mL anti-vimentin FITC, or 1 μg/mL anti-alpha tubulin eFluor 615 for 1.5 h and then imaged using a Nikon A1R confocal microscope at 40× magnification. To investigate the architecture and distribution of the P2Y2 receptor, the purine receptor that reportedly has the most prominent role in ATP binding,25,35,36 samples were blocked with 10% donkey serum for 30 min and then incubated in 1.5% donkey serum containing 4 μg/mL of the P2Y2 antibody for 1.5 h. Next, the samples were washed in PBS, incubated with 5 μg/mL donkey anti-goat Alexa Fluor 568 in the dark for 1 h, washed with PBS again, and then imaged using a Nikon A1R confocal microscope at 40× magnification.
The remaining halves were dehydrated and embedded in paraffin wax. Constructs were sectioned perpendicular to the disc face yielding 5 μm thick sections. Collagen type II was then identified through immunohistochemistry. Sections were treated with peroxidase, followed by chondroitinase ABC in a humidified environment at 37°C for 1 h to permeabilize the extracellular matrix. Samples were then blocked with goat serum, and afterward, the primary antibody for collagen type II (mouse monoclonal) was applied for 1 h. Next, the secondary antibody (anti-mouse IgG biotin conjugate) was added for 1 h followed by incubation with ABC reagent for 45 min. Finally, the slides were developed with DAB peroxidase for 5 min. Samples were washed with PBS between each step.
Statistical analysis
Statistical analysis was performed using Prism (version 4.03; GraphPad Software). Biochemical results, both numerical and graphical, were expressed in the form of mean ±standard deviation. Differences between HP and FS samples were determined using a Student's t-test, while differences between groups cultured with or without the various inhibitors were determined using a one-way ANOVA with Bonferroni post test. A level of p < 0.05 was considered significant.
Results
ATP is released by MSCs in response to HP through hemichannels
The first step in determining whether purinergic signaling regulates the mechanotransduction of HP was to assess whether HP increases the amount of ATP released into the media. After 15 min of loading, no significant increase in ATP in the media was found. After 4 h of exposure to HP, extracellular ATP tended to increase in the control hydrogels that were cultured without inhibitors on days 7 and 14 and increased significantly on day 21; however, flufenamic acid (blocks hemichannels) abrogated this response at all three time points (Fig. 2).

ATP released by MSCs cultured either with or without an inhibitor of hemichannels after 4 h of exposure to HP on days 7, 14, and 21. #p < 0.10, *p < 0.05, relative to FS condition (n = 9). ATP, adenosine triphosphate; HP, hydrostatic pressure; MSCs, mesenchymal stromal/stem cells; FS, free swelling.
Purinergic signaling regulates the chondrogenic response of MSCs to HP
DNA and sGAG/DNA levels in the FS Ctrl, -Hemi, -ATP, and -PRec constructs were assessed to determine whether the pharmacological inhibitors had any influence on viability or differentiation of MSCs. None of the inhibitors had a significant effect on either the DNA or the sGAG/DNA levels (p = 0.158 and 0.060, respectively; Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/tea).
Next, to determine the effect of purinergic signaling on the response of MSCs to HP, pharmacological inhibitors of various components of the purinergic signaling pathway were utilized. In the control specimens without inhibitors, HP significantly enhanced total sGAG/DNA synthesis by 15% (Fig. 3A). Not only was ATP released from MSCs through hemichannels (Fig. 2) but also inhibition of hemichannels was found to suppress the chondrogenic response of MSCs to HP (Fig. 3A). Next, extracellular ATP was hydrolyzed and P-receptors were blocked to investigate the downstream effects of ATP release on the mechanotransduction of HP. Suppressing either ATP or P-receptors abrogated the beneficial effect of HP on total sGAG/DNA production (Fig. 3A). Immunohistochemistry indicated that HP also appeared to enhance collagen type II accumulation, but inhibiting hemichannels, extracellular ATP, and P-receptors suppressed this response (Fig. 3B).
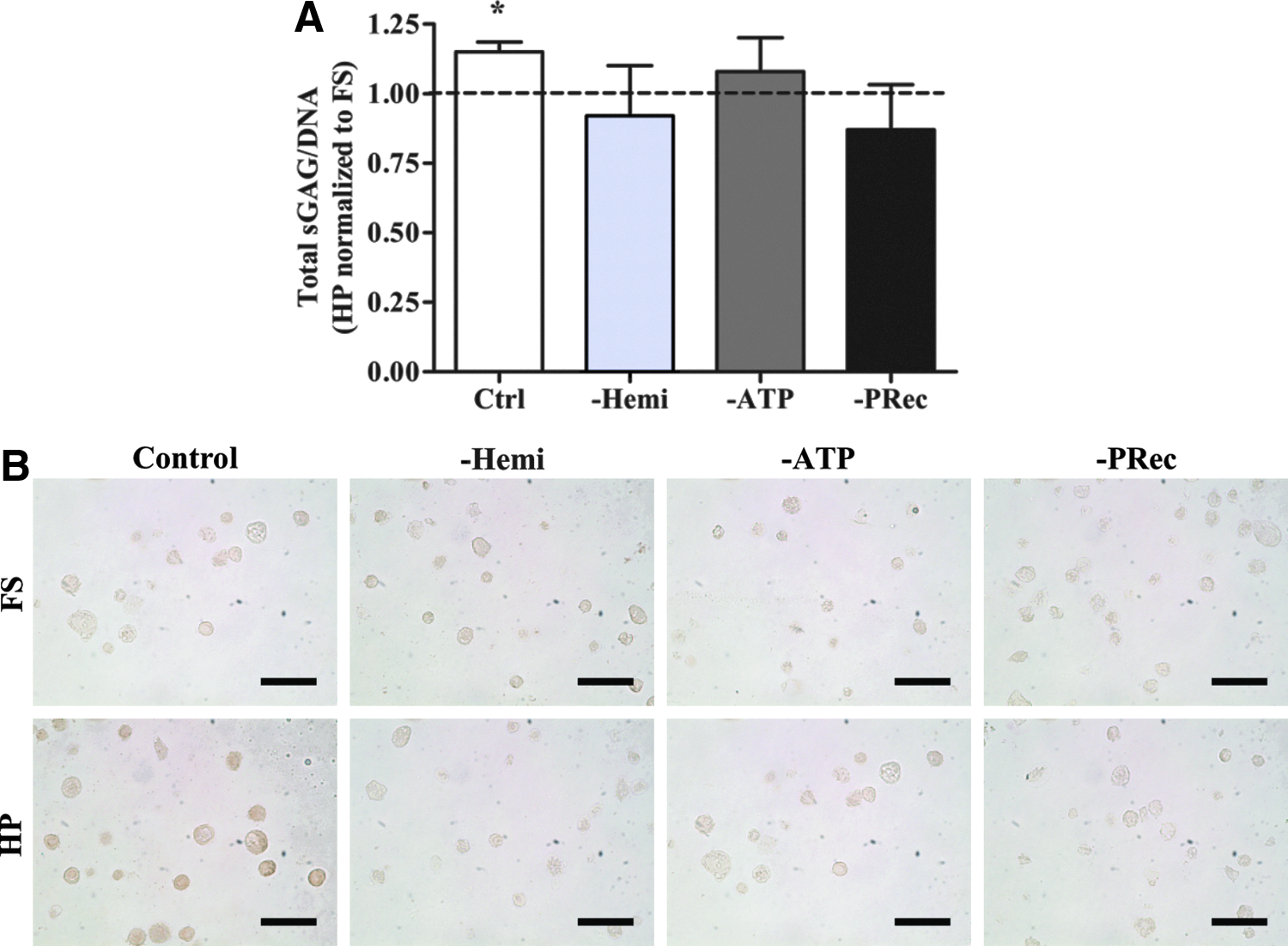
Purinergic signaling is required for changes in vimentin architecture in response to HP
Focal adhesion formation was previously determined to be necessary for mechanotransduction of HP. 7 Therefore, the distribution of focal adhesions was next examined with confocal microscopy to determine if inhibition of purinergic signaling regulates the chondrogenic response of MSCs to HP through alterations in focal adhesion formation. Neither the pharmacological inhibitors nor application of HP affected focal adhesion formation, with all groups demonstrating a punctate structure (Supplementary Fig. S2).
No effect on actin or tubulin architecture was observed with either inhibition of purinergic signaling or application of HP (Supplementary Fig. S2). However, the application of HP led to a more diffuse vimentin structure in the control group, while retaining a punctate structure in the other groups that were cultured with inhibitors (Fig. 4).
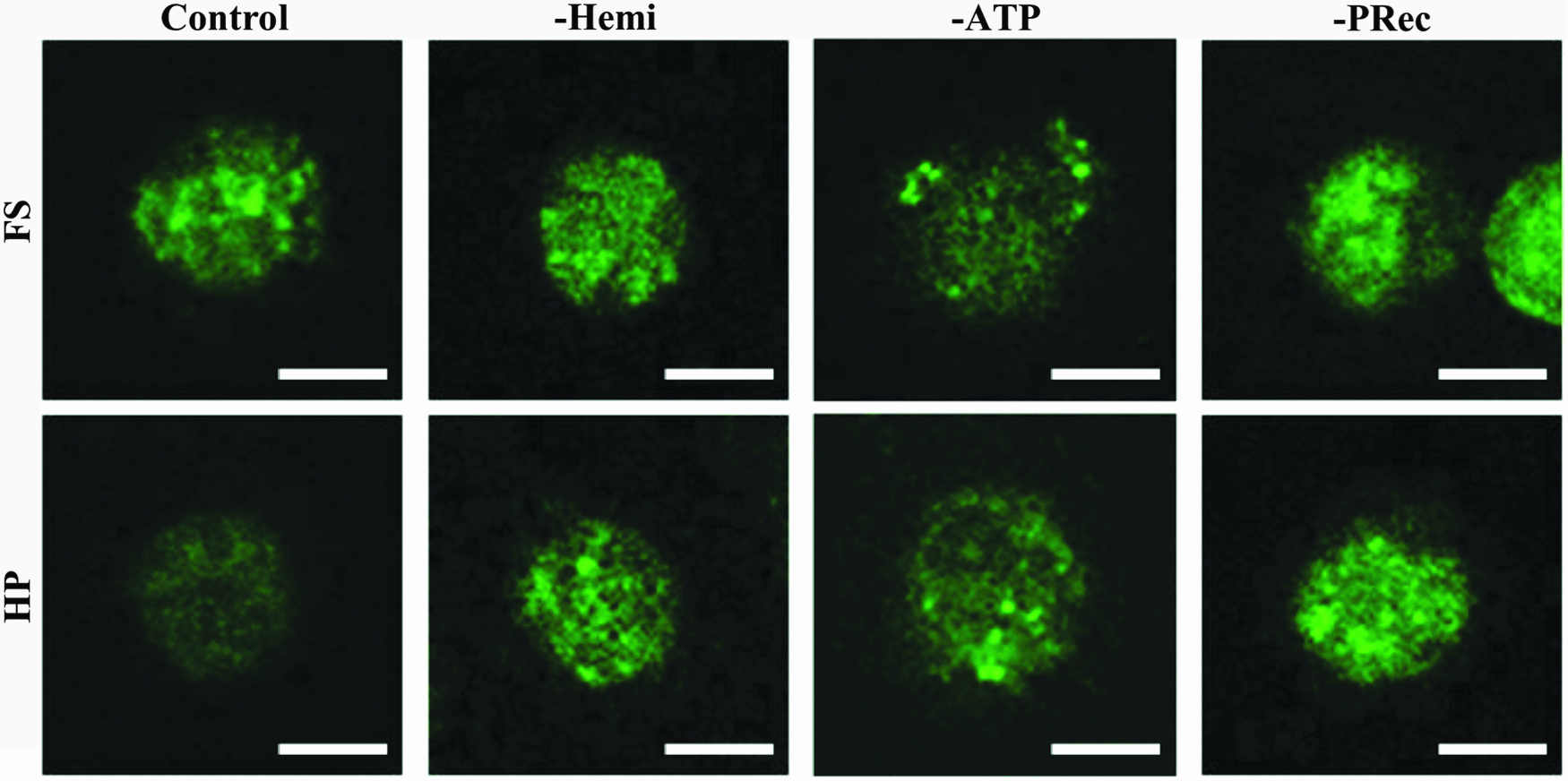
Representative confocal images of vimentin in constructs in either the HP or FS groups. Scale bars = 10 μm. Color images available online at www.liebertpub.com/tea
Purinergic signaling acts upstream of matrix stiffness in the mechanotransduction of HP
As stated previously, matrix stiffness mediates the mechanotransduction of HP 7 ; however, whether this differential response is regulated by purinergic signaling was yet to be investigated. Application of HP had no effect on ATP release in either the soft or stiff hydrogels after 15 min of loading (data not shown). However, ATP release was stimulated in both after 4 h of loading on days 7 and 14 and significantly increased on day 21 (Fig. 5A). In addition, no differences were observed in the P-receptor distribution between MSCs in the soft and stiff gels (Fig. 5B). While ATP release and P-receptor distribution appeared to be similar in the two groups, total sGAG production was only enhanced in the stiff hydrogels in response to HP (Fig. 5C).

Addition of exogenous ATP does not replicate effects of HP
Since purinergic signaling was found to be required for the mechanotransduction of HP and this pathway is thought to act upstream of Ca++ signaling, we next sought to determine if adding exogenous ATP could replicate the chondrogenic effects of HP. At the concentrations of ATP used in the current study, no differences in total sGAG/DNA were observed (Fig. 6).

Total sGAG/DNA produced in soft and stiff hydrogels cultured with or without ATP.
Discussion
Previously, Ca++ signaling was found to play a key role in the mechanotransduction of HP 24 ; however, the underlying mechanisms initiating the Ca++ signaling pathway are unclear. Purinergic signaling is activated in response to other mechanical loads, such as compression and fluid flow, and initiates Ca++ signaling.19,27,37–41 Similarly, HP was found to increase extracellular ATP through hemichannels in the current study. Furthermore, the application of HP enhanced the chondrogenesis of MSCs, while inhibition of hemichannels, extracellular ATP, and P-receptors eliminated this beneficial response. Together, these data suggest that purinergic signaling regulates the chondrogenic response of MSCs to HP.
Previously, we demonstrated a novel role for vimentin in the chondrogenic response of MSCs to HP, as vimentin architecture transitioned from a punctate appearance in FS conditions to a more diffuse organization when exposed to the mechanical load 7 . Vimentin reorganization in response to HP was also found to be dependent on integrin binding and Ca++ signaling.7,24 To determine whether the cytoskeletal response also depends on purinergic signaling, vimentin organization was investigated in the presence of inhibitors of this mechanotransduction pathway. Vimentin organization was punctate in all of the FS conditions, including in the controls and in the specimens exposed to the pharmacological inhibitors. Similar to the previous experiments, vimentin staining became more diffuse when the MSCs were exposed to HP in the control group. However, vimentin remained punctate with loading in the presence of all inhibitors of purinergic signaling utilized in this study. A relationship between purinergic signaling and vimentin organization has been observed in astrocytes; in this previous study, addition of exogenous IP3 (a secondary messenger of purinergic signaling) phosphorylated and reorganized vimentin. 42 Overall, our results suggest that vimentin reorganization is integrally correlated to the chondrogenic response of MSCs to HP and that this reorganization is dependent on purinergic signaling.
Matrix stiffness also regulates the mechanotransduction of HP 7 ; however, it was unclear whether purinergic signaling acts upstream or downstream of the effects of matrix stiffness. ATP release in response to HP and P-receptor distribution in both soft and stiff hydrogels appeared similar, yet chondrogenesis was only enhanced by HP in the stiff gels. Therefore, purinergic signaling appears to act upstream of the effects of matrix stiffness on the mechanotransduction of HP; however, the specific pathway regulated by matrix stiffness or the specific MSC phenotype promoted by a stiff matrix, which alters the effects of purinergic signaling, is unclear. P-receptors contain arginine-glycine-aspartic acid (RGD) sequences that interact with integrins, and inhibition of this linkage was found to decrease the signaling capabilities of both the P-receptors and focal adhesions. 43 Furthermore, matrix stiffness is known to modulate integrin binding7,44–46, which provides a potential mechanism for matrix stiffness to indirectly regulate P-receptor sensitivity. Therefore, although ATP was released and P-receptors were expressed in the soft hydrogels, one explanation for the lack of response to the mechanical signal in soft gels is that the P-receptors may not be “primed” to initiate a signaling cascade in the soft pericellular environment.
Understanding the mechanotransductive pathways involved in the chondrogenic response of MSCs to HP could have important implications in cartilage tissue engineering and regenerative medicine. While mechanical stimulation with HP clearly increases the chondrogenesis of MSCs, long-term mechanical loading is costly. Since purinergic signaling is thought to act upstream of Ca++ signaling19,38 and appears to act upstream of matrix stiffness in the chondrogenic response of MSCs to HP, we hypothesized that stimulating the purinergic pathway through exogenous ATP could replicate the beneficial effect of HP on chondrogenesis in the stiff hydrogels. However, the data did not support this hypothesis; daily addition of exogenous ATP had no effect on the chondrogenesis of MSCs. Previous studies have investigated the effect of exogenous ATP on chondrogenic cells with mixed results. For example, addition of exogenous ATP has been found to increase matrix synthesis of 3D high-density bovine articular chondrocytes.35,47,48 However, addition of exogenous ATP to human MSCs and bovine chondrocytes seeded in agarose hydrogels did not increase matrix synthesis of the constructs, although supplementing the media with ATP only on days 21–28 improved their mechanical properties 36 . In the current study, purinergic signaling was necessary for the mechanotransduction of HP, but the addition of exogenous ATP did not appear to be sufficient to mimic the mechanical load. HP induces additional processes other than purinergic signaling that may also be required in the chondrogenic response to HP. For example, HP inhibits the activity of the sodium-potassium (Na/K) pump and the Na/K/chloride transporter in bovine chondrocytes 49 , but this effect is not likely to be duplicated by the addition of exogenous ATP. Further studies into the effects of HP may uncover additional pathways that are activated by the load and that are necessary to elicit a chondrogenic response.
One limitation of the current study was that we were not able to exactly replicate the ATP exposure of the cells during mechanical loading with the exogenous ATP protocol. Therefore, it is possible that more accurate duplication of the dose, timing, and duration of exogenous ATP supplementation may enhance chondrogenesis in unloaded agarose culture. Due to the short half-life of ATP, a single high-dose supplementation was added to the media, as was done in previous studies.36,47 Potentially, utilization of a computer-controlled perfusion system that can supply lower doses of ATP continuously would better replicate the effects of HP. However, exogenous ATP was added daily, consistent with our HP protocol. An additional limitation is that the pharmacological inhibitors may have had some effect on the behavior of the MSCs, as additional samples would likely indicate that the inhibitors caused a significant difference in the sGAG/DNA levels. This was particularly true for the −PRec group, which was the most different from the control in unloaded conditions. Our previous research has shown that integrin binding to a well-developed pericellular matrix is required for MSCs embedded in agarose to transduce HP.7,24 A major concern, therefore, was that the inhibitors would suppress chondrogenesis such that the cells would no longer transduce the mechanical signal due to an insufficiently developed matrix. As the sGAG/DNA was elevated in the −PRec group, this concern was alleviated.
Overall, purinergic signaling is necessary in the mechanotransduction of HP, as it appears that ATP is released through hemichannels and, subsequently, activates P-receptors in response to the mechanical load. These results also help to explain our previous work, in which the VGCCs, SERCS, and downstream molecules in the Ca++ signaling pathway were determined to be critical in the chondrogenic mechanoresponse to HP. 24 The activated P-receptors allow an influx of Ca++, 30 which could depolarize the membrane and can activate the VGCCs. Once activated, VGCCs have been shown to increase intracellular Ca++ concentration directly and indirectly through calcium-induced calcium release from SERCS. 30 Increased intracellular Ca++ concentration would likely initiate Ca++ signaling cascades, eventually altering gene expression and the differentiation of MSCs (Fig. 7). Matrix stiffness also regulates this pathway, possibly by altering P-receptor sensitivity or by some other mechanism that is downstream of P-receptor activation. The mechanical environment of the cells may also modulate the response to HP through mechanisms that are currently unknown, as exogenous ATP and a stiff matrix were not sufficient to elicit a chondrogenic response on their own. Finally, a deeper understanding of the mechanotransductive pathways activated by MSCs in response to HP and the interactions between these pathways may elucidate pharmaceutical targets to reap the benefits of HP without the costly application of mechanical loading.

The purinergic and calcium signaling pathways proposed to act in the mechanotransduction of HP. (1) HP initiates release of ATP through hemichannels. (2) ATP activates P-receptors to allow Ca++ to enter the cell directly or through IP3-mediated release from SERCS. (3) The increase in intracellular Ca++ concentration triggers VGCCs, which in turn (4) induce calcium-induced calcium release from SERCS. (5) Ca++ then binds to downstream targets, which are known to trigger a variety of signaling cascades that (6) alter gene expression and differentiation. SERCS, sarcoendoplasmic reticulum calcium stores; VGCCs, voltage-gated calcium channels. Color images available online at www.liebertpub.com/tea
Footnotes
Acknowledgments
This study was supported by a Naughton Graduate Fellowship and the National Science Foundation under Grant No. 1335007 (BMMB/CMMI).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.