
Abstract
Since vascular smooth muscle cells (VSMCs) display phenotypic plasticity in response to changing environmental cues, understanding the molecular mechanisms underlying the phenotypic modulation mediated by a three-dimensional (3D) scaffold is important to engineer functional vasculature. Following cell seeding into 3D scaffolds, the synthetic phenotype is desired to enable cells to expand rapidly and produce and assemble extracellular matrix components, but must revert to a quiescent contractile phenotype after tissue fabrication to impart the contractile properties found in native blood vessels. This study shows that 3D electrospun fibrous scaffolds regulate human coronary artery smooth muscle cells (HCASMCs) toward a more synthetic phenotype characterized by reduced contractile markers, such as smooth muscle alpha-actin and calponin. The reduction in contractile markers expression was mediated by endogenously expressed proinflammatory cytokine interleukin-1β (IL-1β). 3D topography transiently induces concomitant upregulation of IL-1β and MAPK ERK1/2 through nuclear factor-κB-dependent signaling pathway. An early burst of expression of IL-1β is essential for suppression of the homeobox transcription factor Gax and related cyclin-dependent kinase inhibitor p21Cip1, which are key regulators for cells exiting from cell cycle. Our findings provide new insights for understanding signaling mechanisms of HCASMCs in electrospun 3D fibrous scaffolds, which have considerable value for application in vascular tissue engineering.
Introduction
O
Although many and complex factors have been reported to control SMC phenotype, cytokines are known to be potent inducers of SMC phenotypic change.5,6 Specifically, it has been suggested that interleukin-1β (IL-1β) produced and recognized by VSMCs regulates their phenotype.7,8 For instance, IL-1β plays a synergistic role in platelet-derived growth factor (PDGF)-BB-induced phenotype change of cultured human aortic smooth muscle cells. 9 IL-1β-stimulated migration of SMCs mediated by PDGF, fibroblast growth factor-2, and transforming growth factor-β (TGF-β). 10 Since IL-1β is a prosynthetic cytokine for VSMCs, it is implicated in the repression of multiple SMC differentiation marker genes and induces phenotypic modulation of SMCs to a distinct inflammatory state through nuclear factor-κB (NF-κB)-dependent mechanisms. 11 In our previous studies, we demonstrated that 3D topography of scaffolds induced the synthetic phenotype of human coronary artery smooth muscle cells (HCASMCs) and increased elastin synthesis in an activated MAPK ERK1/2-dependent manner. 12 Importantly, upregulation of elastin transcription by activated ERK1/2 was dependent on endogenously expressed IL-1β.12,13 Although the detailed mechanism by which scaffold topography initiate intracellular signaling for phenotypic modulation of SMCs is unknown, we suspect that IL-1β acts by suppressing the nuclear transcription factor Gax through NF-κB signaling to induce a synthetic VSMC phenotype needed for matrix deposition during engineered tissue maturation.
Largely confined to the cardiovascular system, nuclear transcription factor Gax is known to be rapidly downregulated when fully differentiated, quiescent, and contractile SMCs in culture enter the cell cycle. 14 In contrast, Gax expression is upregulated by conditions that favor differentiation and cell cycle arrest which involves the inhibition of NF-κB activation and activation of cyclin-dependent kinase (CDK) inhibitor p21Cip1. 15 The transcription factor Gax activates p21Cip1 in vascular endothelial cells, 16 which prevents progression through the cell cycle by blocking CDK from binding to cyclins, a step that is essential for G1/S transition. Overexpression of p21Cip1 reduces human VSMC migration in vitro indicating that p21Cip1 is not only a negative regulator of the cell cycle, but also has important inhibitory effects on VSMC migration. 17 Sustained activation of ERK1/2 by 3D topography of scaffolds in our previous studies 12 led us to suspect the involvement of key nuclear transcription factors in this evolutionarily conserved signaling pathway-mediated elastin biosynthesis through IL-1β. While nuclear transcription factors that function to coordinate cell growth and motility are relatively well described, 18 the role of IL-1β and Gax in VSMC phenotype regulation in 3D cultures is unknown. Identifying signaling events involved in phenotype regulation during vascular tissue engineering significantly accelerate success in tissue fabrication.
In the present work, we investigate the role of transcription factor Gax and ERK pathway in the expression of IL-1β by HCASMCs cultured on electrospun fibrous scaffolds. Our finding provides new insights to better understand signaling mechanisms of SMCs in electrospun 3D fibrous scaffolds.
Materials and Methods
Fabrication of electrospun fibers and preparation for cell culture
To fabricate 3D fibrous scaffolds, solutions of poly(carbonate urethane) (PCU) (Bionate® 55D; DSM Biomedical) with 12% w/v concentrations were prepared in dimethylformamide. Electrospinning was carried out using a high voltage DC power supply (ES30P; Gamma high voltage) at 15 kV voltage, 0.1 mL/h flow rate, 22-gauge needle and 8 cm needle-to-collector distance for 7 h, onto a static collector covered with aluminum foil and nonwoven mat (to facilitate the peeling of the mat from the collector). The morphology of PCU fibers was evaluated using scanning electron microscopy (SEM; S-3400N Hitachi). For SEM, the electrospun fibrous mats were sputter coated with gold/palladium (K550X sputter coater; Emitech Ltd.) and scanned at a working distance of 10 mm and a constant accelerating voltage of 20 kV. Analysis of the SEM was performed with ImageJ software (NIH). For cell cultures, the 3D fibrous mats were cut into desired size and affixed into 12-well cell culture plates using two Teflon O-rings to make a sandwich, sterilized for 30 min with 70% ethanol followed by three washes with Hank's Balanced Salt Solution (HBSS), and then changed to normal cell culture media to equilibrate for 30 min at 37°C.
Cell culture, treatment with human TGF-β1
Primary HCASMCs were purchased from Lonza and cultured according to the supplier's instructions in Medium 231 and smooth muscle growth supplement, supplemented with 100 units/mL penicillin G sodium, 100 μg/mL streptomycin sulfate, and 2 mM
Fluorescence staining and laser scanning confocal microscopy
HCASMCs were seeded on 3D fibrous scaffolds for 4 days and fixed with 4% (w/v) paraformaldehyde (EMD Chemicals, Inc.) for 20 min at room temperature followed by permeabilization for 5 min in phosphate-buffered saline (PBS) containing 0.1% (v/v) Triton X-100. Cells were incubated with Alexa™ Fluor 488-conjugated phalloidin (dilution 1:100; Invitrogen) in 1% bovine serum albumin/PBS for 1 h. DAPI (300 nM in PBS; Invitrogen) was used to label nuclei. Samples were mounted on slides in SHUR/Mount™ (TBS) and analyzed with a Zeiss LSM 510 confocal microscope (Zeiss) equipped with an argon/neon as well as UV lasers.
RNA isolation and qRT-PCR analysis
qRT-PCR combined with reverse transcription were used to quantify gene expression in HCASMCs grown on 2D surfaces and 3D fibrous scaffolds. Total RNA from HCASMCs was extracted using TRIzol reagent (Invitrogen) following the manufacturer's protocol. Complementary DNA was synthesized using 1 μg of total RNA primed with random primers as described in the Promega™ Random Hexamers protocol (Fisher Scientific). qRT-PCR was conducted in 10 μL reaction volumes, using a CFX96™ Real-Time System (C1000 Touch Thermal Cycler; Bio-Rad) and gene expressions of human Gax, IL-1β, SM α-actin, calponin, and GAPDH were then determined with iQ™ SYBR® Green Supermix (Bio-Rad) according to the recommended protocol of the manufacturer. The sequences of primers were designed using Primer3Web and listed in Table 1. Cycling parameters were optimized as follows: denaturation 95°C (15 s), gradient annealing 53/58°C (60 s), extension 72°C (30 s), and running for 40 cycles. The mRNA expression in HCASMCs was normalized to housekeeping gene GAPDH with at least three repeats per experimental group and expressed as a relative ratio using the Bio-Rad CFX Manager 3.0 software.
qRT-PCR, quantitative real-time polymerase chain reaction.
Western blot analyses
Western blotting was performed to evaluate the levels of protein expression in HCASMCs. Briefly, cells were lysed in buffer containing 150 mM NaCl, 10 mM Tris-HCl (pH 7.4), 1 mM EDTA, 0.5% Nonidet P-40, and 1% Triton X-100 and supplemented with protease inhibitor mixture (1 tablet/10 mL of buffer; Roche Applied Science) and phosphatase inhibitors (1 mM NaF and 1 mM Na3VO4). Protein concentrations were determined by the BCA Protein Assay Kit (Thermo Scientific), and 20 μg of total protein lysate was resolved on 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and subsequently transferred at 90 V for 1 h to nitrocellulose membrane. Transfer efficiency and homogeneous loading was assessed by Ponceau S red stain. Nitrocellulose membranes were blocked with 5% nonfat dry milk in PBS for 1–3 h before incubation with anti-phospho-IκB-α (1:500), anti-IκB-α (1:500), anti-IL-1β (1:500), SM α-actin (1:1000), calponin1/2/3 (1:1000) from Santa Cruz Biotechnology; anti-phospho-ERK (1:1000) and anti-ERK (1:1000) from Cell Signaling Technology; anti-p21Cip1 (1:500; Thermal Scientific), anti-integrin α5 (1:1000; BD Biosciences), and anti-GAPDH (1:5000, Millipore) overnight at 4°C. The blots were then probed with horseradish peroxidase-conjugated secondary antibodies followed by the ECL detection (Thermo Scientific) according to the manufacturer's specifications.
siRNA transfection of human IL-1β in HCASMCs
HCASMCs were passaged in growth medium without antibiotics such that they will be 60–80% confluent in a 10-cm culture dish at the time of transfection. Scrambled siRNA or IL-1β siRNA (200 pmol; Santa Cruz) were mixed with 1 mL Opti-MEM reduced serum medium (Invitrogen), then gently mixed with another 1 mL of Opti-MEM reduced serum medium containing 15 μL of RNAiMAX solution (Invitrogen); after incubating this mixture at room temperature for 20 min, this complex was added to the culture dish of 50–80% confluent cells. After 24 h, the siRNA transfected HCASMCs were trypsinized and seeded either on cell culture 2D dishes or 3D fibrous scaffolds. HCASMCs were harvested for testing transfection efficiency or detecting the effect on HCASMC gene expression by western blot and/or qRT-PCR after 24 h incubation.
Statistical analyses
Data are presented as the mean±SD (standard deviation) of three experiments conducted in triplicates. All statistical analyses were performed using GraphPad Prism 5 software. Differences between two groups were compared by two-tailed, unpaired Student's t-test. For multiple group comparisons, two-way analysis of variance followed by Bonferroni's post-test was used. Values of p<0.05 were considered statistically significant.
Results
3D fibrous scaffold promotes early burst of IL-1β expression, but represses differentiation markers of HCASMCs
To mimic the fibrous ECM microenvironment, we fabricated 3D PCU fibrous scaffolds for culturing HCASMCs (Fig. 1A, B). The rationale for using nondegradable PCU fibrous scaffolds is to avoid the potential effects of degradation products that may confound targeted evaluation of cell phenotype and signaling under defined conditions. Before signaling studies, we evaluated cell morphology and attachment on the 3D fibrous mat to demonstrate that cells had intact cytoskeleton as shown by the confocal microscopy image (Fig. 1C). Since our previous study suggested that scaffold topography could be an important environmental cue for promoting elastin synthesis by potentially regulating HCASMCs phenotype, 12 we investigated if 3D topography in electrospun fibrous scaffolds could regulate SMC differentiation marker genes. As shown in Figure 2A and B, the two marker genes SM α-actin and calponin were significantly downregulated by SMCs in 3D fibrous scaffolds compared with 2D surfaces (p<0.05). Pretreatment with human TGF-β1 upregulated these marker gene expressions in 2D cultures, phenotype transition to a more contractile state, but it had no effect on 3D fibrous environment (p>0.05). In addition to gene expression, protein expression levels of the two differentiation markers were downregulated by SMCs in 3D fibrous scaffolds as confirmed by western blot analysis (Fig. 2D). In contrast to these differentiation marker genes, a burst of expression of cytokine IL-1β gene was observed on the fibrous scaffolds compared to 2D counterparts (Fig. 2C). Interestingly TGF-β1 did not have any effect on both 3D and 2D cultures. To further confirm our observations, the protein levels of IL-1β were also determined by western blot analysis. Consistent with the gene expression, IL-1β protein was rapidly upregulated by HCASMCs in 3D fibrous scaffolds after 24 h culture, but surprisingly declined by 48 h (Fig. 2E) indicating a possible temporal effect. These findings suggest that the topography of 3D fibrous scaffolds promotes proliferation of HCASMCs to form a dense cellular layer in more synthetic phenotype and cytokine IL-1β might be involved in regulation of phenotypic transition thereafter.

Scanning electron microscopy images showing three-dimensional (3D) structures of poly(carbonate urethane) (PCU) fibrous scaffolds
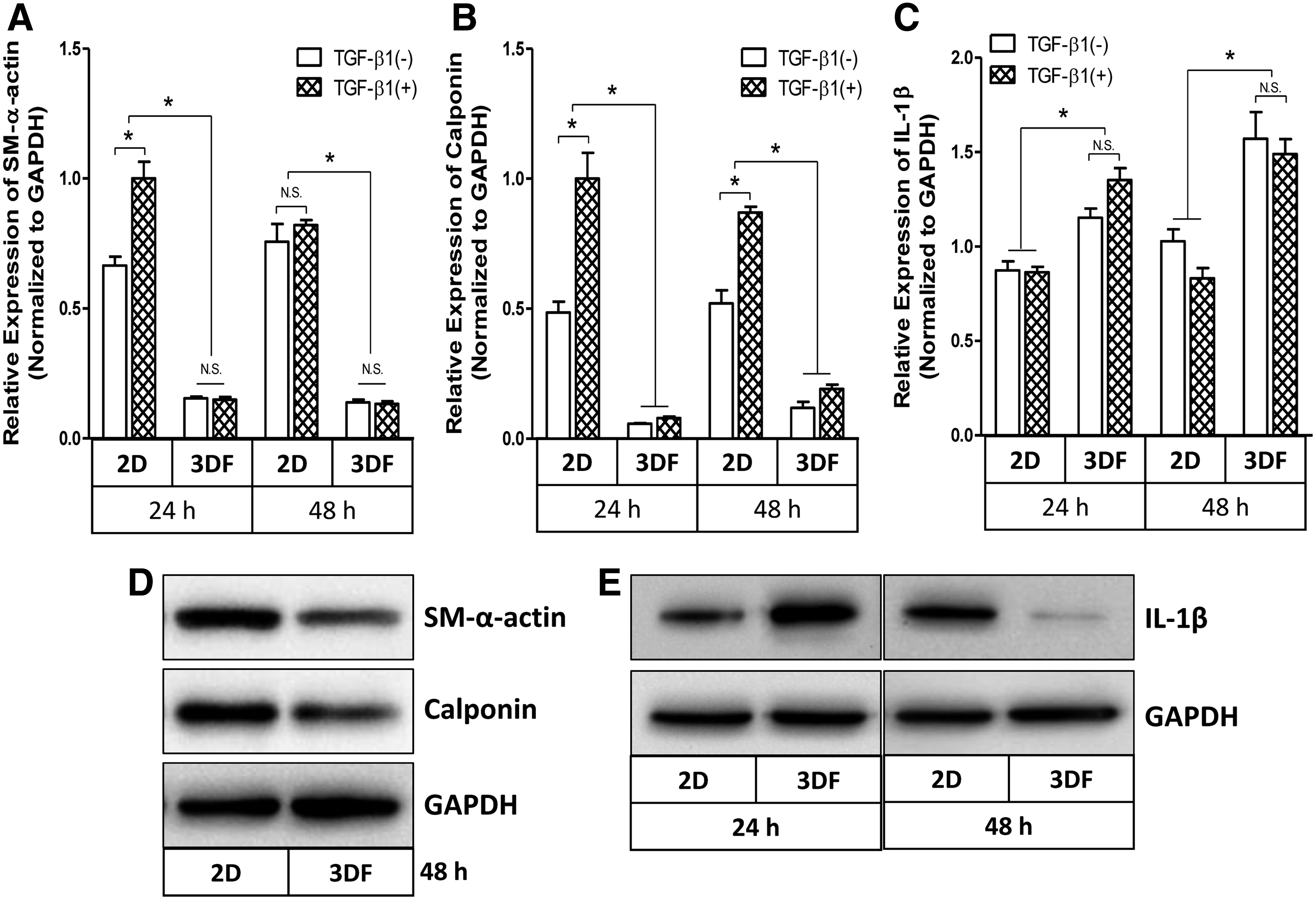
Downregulation of mRNA expression of smooth muscle differentiation marker gene SM α-actin
Scaffold-induced temporal repression of IL-1β expression is linked to a rapid downregulation of MAPK ERK1/2
ERK1/2-associated signaling pathway may play an important role on expression of IL-1β in HCASMCs cultured within 3D fibrous scaffolds. To ascertain the involvement of ERK1/2 activation in the downregulation of IL-1β in 3D fibrous scaffolds, we performed western blot analysis. Downregulation of IL-1β expression by HCASMCs in 3D fibrous scaffolds was first confirmed for two time points (Fig. 3A, B). Figure 3A and B also showed a concurrent downregulation of ERK1/2 phosphorylation in HCASMCs seeded within 3D fibrous scaffolds in a time-dependent manner. By comparing with 2D cultures, we observed that phosphorylation of ERK1/2 was reduced in 3D cultures, but not in 2D (Fig. 3C, D). Although pretreatment with TGF-β1 increased phosphorylation of ERK1/2 in a 2D environment, it had no effect on 3D cultures. Consistent with our previous observation, 12 the current results suggest that activated ERK1/2 is needed for expression of IL-1β and may, in part, be responsible for the downregulation of IL-1β expression in 3D fibrous scaffolds.
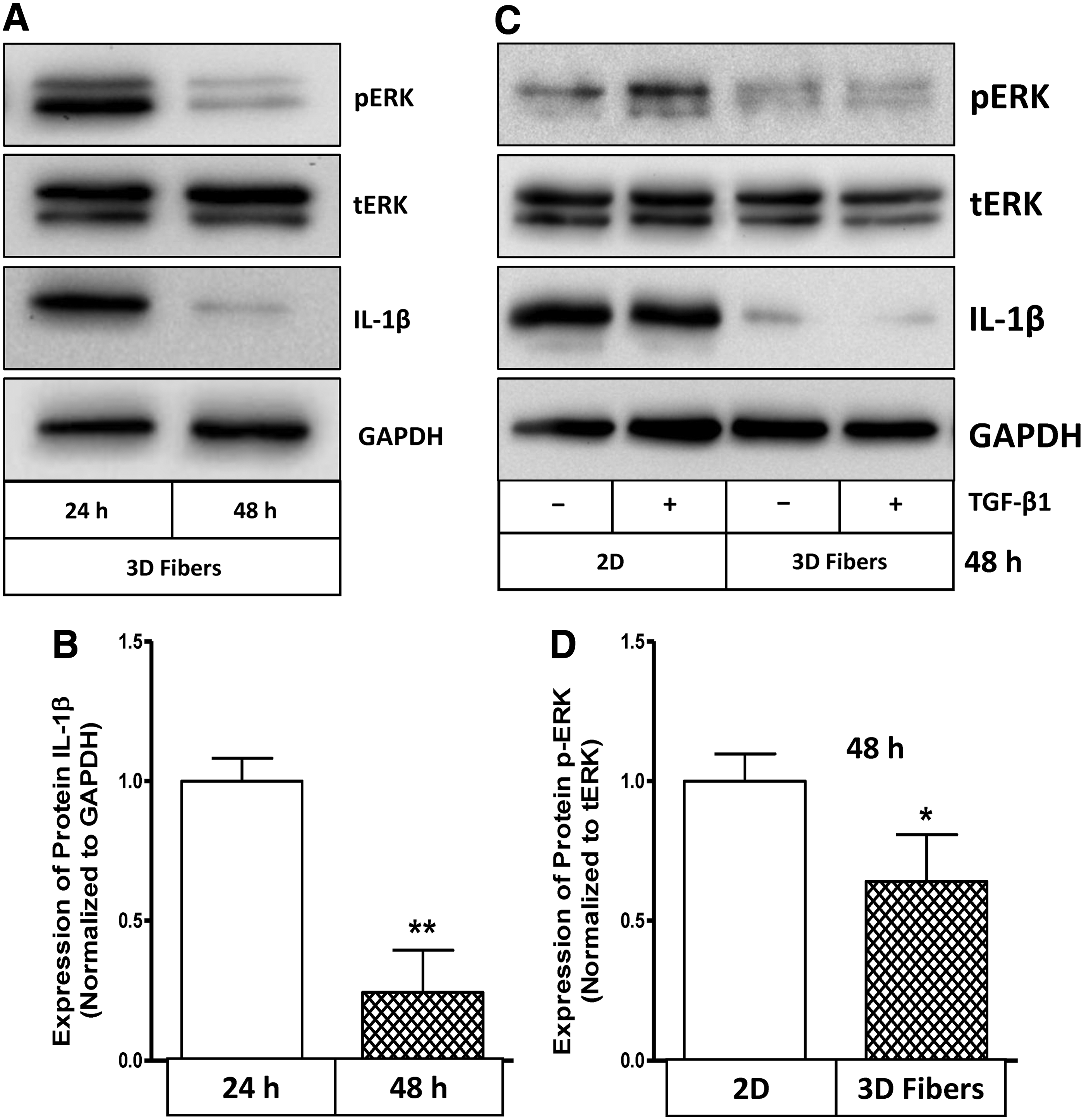
Topography of 3D fibrous scaffolds and hTGF-β1 differentially regulate IL-1β protein expression and ERK1/2 activation between 2D and 3D environments.
Knockdown of IL-1β mediated by siRNA transfection promotes expressions of two differentiation marker genes and blocks activation of nuclear transcription factor NF-κB
Given the downregulation of differentiation marker genes and concomitant upregulation of cytokine IL-1β in 3D fibrous scaffolds, we investigated the role of IL-1β in the repression of the differentiated SMCs phenotype. To do this, we first knocked down the gene expression of IL-1β in cultures using siRNA and assessed the transfection efficiency of knockdown (Fig. 4A, B). We then evaluated the effect of endogenous IL-1β knockdown on expression of SMC differentiation marker genes. As shown in Figure 4C and D, SM α-actin and calponin were significantly upregulated by knockdown of IL-1β mRNA suggesting that IL-1β is an upstream regulator for repression of differentiation marker genes.

Efficiency of siRNA transfection-mediated knockdown of IL-1β was determined by western blot analysis
IL-1β can modulate smooth muscle cell phenotype to a distinct inflammatory state through NF-κB-dependent mechanisms in conventional 2D cell cultures in which IL-1β represses the expression of SMC differentiation marker genes.11,19 We speculate that, on 3D fibrous mats, IL-1β might also repress marker genes through induction of NF-κB activation; and therefore declined IL-1β expression in 3D fibrous scaffolds could promote HCASMC differentiation without NF-κB activation. Inactivated NF-κB dimers are sequestered in the cytosol of cells through noncovalent interactions with a class of inhibitor proteins, called I-κB (I-κB-α is one of the members of I-κB family). Signals that induce NF-κB activity cause the phosphorylation of I-κB, their dissociation, and subsequent degradation, thereby allowing activation of the NF-κB complex. 20 Considering IL-1β was rapidly induced by HCASMCs in 3D fibrous scaffolds after culture for 24 h, we knocked down IL-1β gene mediated by siRNA transfection in HCASMCs and cultured these cells in 3D fibrous scaffolds for 24 h. As shown in Figure 5A, phosphorylated I-κB-α indicates that NF-κB activation of HCASMCs was rapidly induced within 3D fibrous scaffolds after 24 h (scrambled group as control), and that siRNA-mediated knockdown of IL-1β resulted in an inhibition of NF-κB activation through dephosphorylation of I-κB-α. The results suggested that the topography-induced IL-1β within 3D fibrous scaffolds could modulate phenotypic transition of HCASMCs through NF-κB-mediated signaling pathway.

Phosphorylation of I-κB-α and cell cycle-dependent inhibitor p21Cip1 were regulated by IL-1β and 3D topography of fibrous scaffolds.
Knockdown of IL-1β mediated by siRNA transfection triggers cell cycle inhibitor p21Cip1 involved in the activation of homeobox gene Gax
Studies reported that IL-1β may cooperate with PDGF-BB to increase cell cycle progression and promote SMC proliferation by suppression of PDGF-BB-induced cell cycle inhibitor p21Cip1. 21 Consistent with this understanding, p21Cip1 expression was significantly triggered by IL-1β knockdown in 3D fibrous scaffolds implicating a positive role of IL-1β on the suppression of p21Cip1 (Fig. 5A, B). Moreover, a time-dependent increase of p21Cip1 was also observed in HCASMCs seeded in 3D fibrous scaffolds in a longer term culture up to 96 h (Fig. 5C, D) that might be due to the downregulation of IL-1β expression in later time points. These results indicated that regulation of IL-1β by HCASMCs in 3D fibrous scaffolds may be responsible for p21Cip1-mediated signaling mechanisms.
Given that the ready expression of endogenous IL-1β inhibits p21Cip1 and lack of IL-1β expression could trigger rapid upregulation of p21Cip1 in turn leading to p21Cip1-dependent cell cycle arrest, migration, and inhibition of proliferation, we wanted to investigate if human homeobox gene Gax was involved in the regulation of IL-1β by HCASMCs, as GAX regulates cell cycle and proliferation through p21Cip1.16,22 An early burst of IL-1β expression was rapidly induced and hGax gene was concomitantly sequestered by HCASMCs cultured on 3D fibrous scaffolds compared with 2D cultures (Figs. 2E and 6A). However, siRNA-mediated knockdown of IL-1β significantly upregulated the expression of hGax gene suggesting that expression of IL-1β is likely responsible for hGax gene regulation. After culture for 48 h, expression of hGax gene was significantly upregulated by HCASMCs in 3D fibrous scaffolds, which corresponds to the downregulation of IL-1β when compared to 2D cultures (p<0.001; Fig. 6B). The sustained expression of hGax gene was persistent up to 96 h (Fig. 6C) that might be due to the absence of IL-1β expression. In addition, Gax is a key regulatory factor for the expression of several integrin isoforms in which the respective β chains of the αvβ3 and αvβ5 integrins are specifically downregulated by Gax. 15 Indeed, our results indicated that downregulation of α5-integrin protein expression and concomitant upregulation of hGax was induced by HCASMCs in 3D fibrous scaffolds through p21Cip1-mediated signaling pathway (Fig. 6D, E).

Regulation of homeobox hGax gene by HCASMCs in 3D fibrous scaffolds
Discussion
Cell signaling studies in 3D scaffolds are an important component in tissue engineering strategies. Understanding of signals involved in the 3D scaffold microenvironment provides a powerful tool to understand cellular responses to materials in an in vivo relevant model (i.e., 3D). In this study, we show that HCASMCs seeded on electrospun fibrous PCU scaffolds acquired a more synthetic phenotype by upregulating endogenous IL-1β (Figs. 1 and 2). We previously reported successful induction of elastin biosynthesis by HCASMCs within 3D porous PCU scaffolds using a particulate leaching approach, in which the 3D scaffold topography alone was found to be an important environmental cue to regulate HCASMCs phenotype. 13 As a promising approach, electrospinning technique is capable of generating nanoscale fibers to fabricate a scaffold which provides a suitable microenvironment for cells. In the current work, nondegradable PCU has been chosen to avoid the possibility of degraded fragments affecting signaling events initiated by the 3D fibrous structure. 23 Our studies showed that contractile markers SM α-actin and calponin were significantly downregulated by HCASMCs seeded within 3D fibrous scaffolds (Fig. 2) modulating HCASMC phenotype toward more synthetic phenotype.
The key finding of the current study is the role of endogenously expressed IL-1β in the regulation of Gax and subsequent HCASMC phenotype modulation. Recent progress in understanding tissue formation suggests that in a regenerative medicine model, the immune response, particularly the cytokine-associated inflammatory process, may play a key role in neovessel development. 24 IL-1β, as a major cytokine released during inflammatory response, has been shown to repress the expression of multiple SMC differentiation marker genes 25 and promote SMC phenotypic modulation to an inflammatory state by activating NF-κB-dependent proinflammatory genes. 11 Our current studies demonstrate that IL-1β was rapidly upregulated with concomitant downregulation of differentiation marker genes by HCASMCs in 3D fibrous scaffolds, which might be happening through an autocrine mechanism, since IL-1β can induce its own expression through a positive autoregulatory loop associated with the recruitment of NF-κB pathway. 26 NF-κB is a stress-sensitive transcription factor activated by both proinflammatory cytokine and shear stress. 27 In human coronary SMCs, it is reported that pulsatile stretch stimulates superoxide production and activates NF-κB. 28 Recent report demonstrated that SMC-selective inhibition of NF-κB signaling pathway attenuated SMC phenotypic switching and neointimal formation. 29 On the other hand, NF-κB activation initiates transcription of many genes, including cytokines, which can enhance early monocyte recruitment to the scaffold facilitating neovessel formation. 30 In this study, we show that NF-κB activation was blocked due to the absence of expression of IL-1β after culture for 48 h (Fig. 5A, B).
SMCs phenotype changes in response to a variety of growth factors, and entrance to the cell cycle is regulated by homeobox Gax gene, which is a negative regulator of proliferation, and exerts remarkable inhibitory effects on the proliferation of VSMCs in vitro.22,31 Gax has been shown to be rapidly downregulated when cultured VSMCs re-enter the cell cycle upon stimulation by serum or PDGF in vitro 32 and downregulated in vivo in response to vascular injury-induced changes in vessel wall morphology that leads to excessive cellular proliferation. 33 Overexpression of Gax can inhibit cell proliferation in a CDK inhibitor p21Cip1-dependent manner 22 and particularly, activated p21Cip1 can be inhibited by IL-1β in response to PDGF-BB in SMCs. 21 In contrast with CDKs, which are positive regulators of cell cycle progression, the CDK inhibitor p21Cip1 is a critical negative regulator of the cell cycle that provide counterbalance to increased cyclin/CDK activities. 17 In 3D scaffolds, an early burst of IL-1β by HCASMCs could be a major suppressor for the expression of p21Cip1 and thus promotes HCASMCs to remain in a more synthetic phenotype. While confluence of cells was rapidly achieved in fibrous scaffolds, withdrawal of IL-1β expression could result in rapid upregulation of p21Cip1. Indeed, our results clearly showed that protein levels of p21Cip1 were initially sequestered and increased by HCASMCs in scaffolds in a time-dependent manner (Fig. 5C, D). Expectedly, transcription factor Gax showed the similar pattern of expression as p21cip1 suggesting a role of transcriptional regulator (Fig. 6). Although overexpression of Gax gene inhibits inflammatory cytokine expression including IL-1β and attenuates vascular remodeling as has been reported, 34 it seems not the case in this study because siRNA transfection-mediated knockdown of IL-1β significantly upregulated the expression of Gax gene and p21cip1 in 3D environments (Figs. 5A and 6A). Our results strongly suggest that IL-1β most likely acts as a regulator upstream of Gax gene-mediated signaling pathway. This was also supported by a time-course experiment in which the gene expression of Gax was upregulated by 3D topogarphy in a time-dependent manner (Fig. 6C). Therefore, our results demonstrate that higher levels of expression of IL-1β at an early stage of cell seeding are essential for the suppression of homeobox transcription factor Gax in a CDK inhibitor p21Cip1-dependent manner that facilitates HCASMCs remaining in a more synthetic phenotype in 3D fibrous scaffolds. Consistent with the reports by others, Gax is also a key regulatory factor for the expression of several integrin isoforms as the respective β chains of the ανβ5 integrins are specifically downregulated by Gax. 15 The upregulation of Gax and concomitant downregulation of protein expression of α5-integrin has been observed in 3D fibrous scaffolds (Fig. 6B, D). Notably, NF-κB is shown to bind directly to the α5β1-integrin promoter region in 3D culture of tumor cells, resulting in β1 integrin overexpression, which reveals a β1-integrin–NF-κB-β1-integrin loop-like activation pathway. 35 There also has been evidence that cell cycle arrest appeared important especially for Gax overexpression-induced inhibition of cell migration associated with the downregulation of integrin.14,15 Thus, IL-1β withdraw-induced inactivation of NF-κB could be responsible for the downregulation of α5-integrin in this study. It is noteworthy that our experiments were conducted for relatively short times; between 1 day (24 h) and 4 days (96 h) because we wanted to study signaling events early in the cell–fibrous scaffold interaction process. Beyond 4 days, we suspected that ECM proteins produced and secreted by cells may change the microenvironment and hence the signaling process that is mediated by the fibrous scaffolds. Future studies will investigate time points beyond 4 days.
In summary, we found that autocrine IL-1β signaling is an integral part of HCASMCs phenotype transition by regulating the activation of transcription factor Gax. The initial suppression of hGax gene induced by IL-1β blocked cell cycle inhibitor p21Cip1 through activated NF-κB-dependent signaling pathway, which promotes dedifferentiation and migration of HCASMCs in 3D fibrous scaffolds. Furthermore, we showed that suppression of IL-1β activated hGax gene expression followed by the activation of p21Cip1-associated signaling pathway and, in turn, inhibited cell migration by blocking the protein expression of α5-integrin.
Footnotes
Acknowledgment
This work was supported by the Heart and Stroke Foundation of Canada (HSFC) grant. Somiraa Said is acknowledged for taking the SEM images of the fibrous scaffolds.
Disclosure Statement
No competing financial interests exist.