
Abstract
Human periodontal ligament stem cells (hPDLSCs) are considered potential cellular carriers for gene delivery in the field of tissue regeneration. This study tested the osseoregenerative potential of hPDLSCs transduced with replication-deficient recombinant adenovirus (rAd) containing the gene encoding bone morphogenetic protein-2 (BMP2; hPDLSCs/rAd-BMP2) in both in vivo and in vitro osteogenic environments. After the optimal condition for rAd-mediated transduction was determined, hPDLSCs were transduced to express BMP2. In vivo bone formation was evaluated in a critical-size rat calvarial bone defect model that more closely mimics the harsher in vivo milieu for bone regeneration than subcutaneous transplantation model. As support materials for bone regeneration, block-type biphasic calcium phosphate (BCP) scaffolds were combined with hPDLSCs and/or BMP2 and transplanted into critical-size bone defects in rats. Experimental groups were as follows: BCP scaffold control (group 1 [Gr1]), scaffold containing recombinant human BMP2 (rhBMP2; group 2 [Gr2]), scaffold loaded with normal hPDLSCs (group 3 [Gr3]), scaffold combined with both normal hPDLSCs and rhBMP2 (group 4 [Gr4]), and scaffold loaded with hPDLSCs transduced with rAd-BMP2 (hPDLSCs/rAd-BMP2; group 5 [Gr5]). Our data showed that new bone formation was highest in Gr2. Less mineralization was observed in Gr3, Gr4, and Gr5 in which hPDLSCs were transplanted. In vitro transwell assay demonstrated that hPDLSCs exert an inhibitory activity on BMP2-induced osteogenic differentiation. Our findings suggest that the in vivo bone regenerative potential of BMP2-overexpressing hPDLSCs could be compromised in a critical-size rat calvarial bone defect model. Thus, further investigations are required to elucidate the underlying mechanisms and to develop efficient techniques for improved tissue regeneration.
Introduction
R
Among a number of signaling molecules, bone morphogenetic protein-2 (BMP2) has acquired considerable attention in this field. As a growth factor, BMP2 plays an essential role in bone formation by regulating osteoblast differentiation. 5 Although the delivery of BMP2 through synthetic scaffolds was shown to promote bone repair, 6 various adverse effects can occur, such as severe inflammatory soft-tissue swelling, ectopic bone formation, and the emergence of bone voids. 7 Most of these adverse effects may be associated with the direct delivery of recombinant BMP2. BMP2 is rapidly cleared from the local sites of transplantation likely due to fast release and short half-life, thus unable to efficiently act on neighboring cells. 8 The high-dose application of recombinant BMP2 to prolong its bioavailability and enhance its efficacy possibly increases the risk of adverse effects.
The release of growth factors depends on the inherent property of its carrier. 9 An ideal carrier should possess appropriate biochemical properties with adjustable release of growth factors at local sites of transplantation, thereby maximizing the efficacy and minimizing the toxicity. In this context, stem cell-based gene therapy appears to be an attractive alternative to the conventional delivery system. A recent strategy of gene transfer is to genetically modify multipotent stem cells with viral or nonviral vectors encoding the desired growth factor. These genetically modified stem cells can provide sustained bioavailability of the growth factor within the local bone defect area, leading to an enhanced bone formation. 10
Stem cells of dental origin, for example, periodontal ligament (PDL) stem cells (PDLSCs) are considered efficient gene delivery carriers. They have been demonstrated to have a proliferative activity for self-renewal and multipotency to differentiate into various cell types. 11 PDLSCs also exhibit a potent immunomodulatory activity. 12 These mesenchymal stem cell (MSC)-like properties make them an excellent source for MSC-based cell therapeutics. Accordingly, we investigated the potential of engineered human PDLSCs (hPDLSCs) in our previous study. 13 Replication-deficient recombinant adenovirus (rAd) was used to introduce a transgene containing the genetic information for the expression of BMP2 into hPDLSCs. BMP2 transduction did not affect stem cell characteristics of hPDLSCs. Sustained release of BMP2 was more potent than single high dose of recombinant human BMP2 (rhBMP2) in bone regeneration.
Nonetheless, the promising results of our previous study were limited by in vivo experimental design; the effects of BMP2-expressing hPDLSCs on bone formation were examined only in dorsal subcutaneous pockets of immunodeficient mice rather than in a bone defect animal model. Therefore, it is necessary to test this method in a bone defect model to confirm the osteogenic potential of the transduced hPDLSCs. Moreover, in our previous study, only weak osteoinduction was observed despite treatment with BMP2 when PDLSCs were present, suggesting an inhibitory effect of hPDLSCs on BMP2-induced bone regeneration. 13 Our observation is consistent with some recent studies. Thus, further study is required to investigate whether PDLSCs are inhibitory or not in BMP2-induced osteoinduction.14,15
In this study, we determined the optimal rAd concentration for hPDLSC transduction and evaluated the effects of BMP2-expressing hPDLSCs on bone formation in a rat critical-size bone defect model using a biphasic calcium phosphate (BCP) block; this animal model was chosen for harsher milieu for bone regeneration. Finally, we evaluated the effects of soluble factors released by hPDLSCs on the osteogenic differentiation of human bone marrow-derived mesenchymal stem cells (hBMSCs).
Materials and Methods
hPDLSC isolation
Third molars extracted from a healthy patient who visited the Division of Periodontology, Department of Dentistry of Inha University Hospital, Incheon, Korea were used to obtain hPDLSCs. The applied isolation protocol was approved by the Ethics Committee of Inha University Hospital (Approval No. IUH IRB 12-150). The subject enrolled in this study was informed about how her teeth were to be used, after which she provided written informed consent. The isolation protocol used in this study has been described previously.
16
In brief, the root surface of the extracted tooth was scraped to obtain periodontal tissue, which was then digested using 2 mg/mL collagenase (Wako Pure Chemical Industries, Tokyo, Japan) and 1 mg/mL dispase (Gibco, Grand Island, NY). The cells were collected and seeded in a T75 cell culture flask (BD Falcon Labware, Franklin Lakes, NJ). A growth medium comprising the following ingredients was used to culture the cells: alpha minimum essential medium (Gibco), 15% fetal bovine serum (Gibco), 2 mM
Preparation of adenoviral vectors and hPDLSC transduction
rAd was manipulated to express either enhanced green fluorescent protein (EGFP; rAd-EGFP) or BMP2 (rAd-BMP2) using the AdEasy Vector System (Qbiogene, Carlsbad, CA) according to the manufacturer's instructions (Fig. 1). The hPDLSCs were then transduced using rAd-EGFP or rAd-BMP2 at various doses (0, 10, 20, and 50 multiplicities of infection [MOI]), and labeled hPDLSCs/rAd-EGFP and hPDLSCs/rAd-BMP2, respectively.
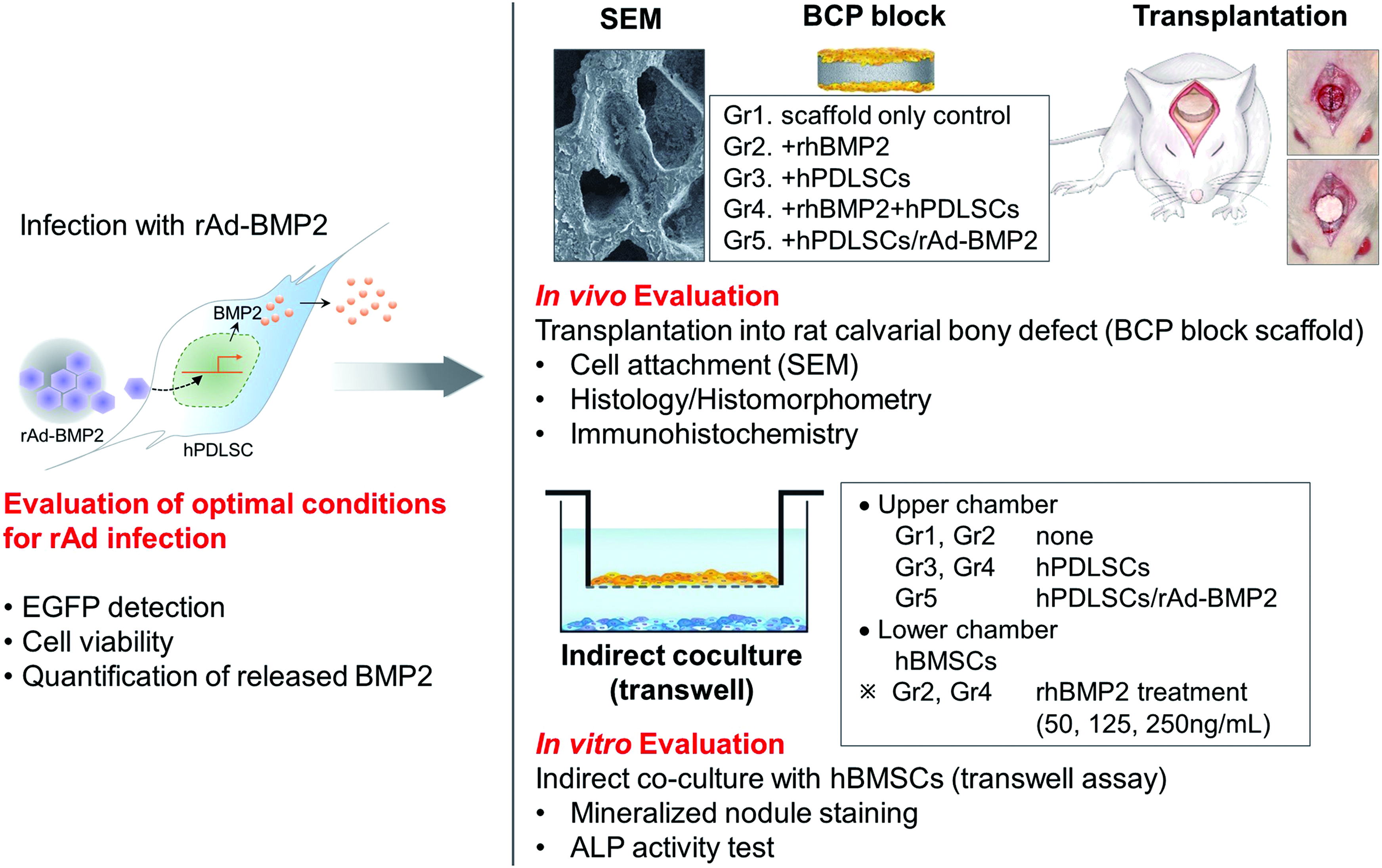
Schematic diagram of the study. hPDLSCs obtained from human third molars were transduced using rAd to produce BMP2. The most efficient rAd concentration for the transfection was determined, and the osseoregenerative characteristics of hPDLSCs and hPDLSCs/rAd-BMP2 were analyzed in both in vivo and in vitro osteogenic environments. ALP, alkaline phosphatase; BCP, biphasic calcium phosphate; EGFP, enhanced green fluorescent protein; hBMSCs, human bone marrow-derived mesenchymal stem cells; hPDLSCs, human periodontal ligament stem cells; rAd-BMP2, replication-deficient recombinant adenovirus containing the gene encoding bone morphogenetic protein-2; SEM, scanning electron microscope. Color images available online at www.liebertpub.com/tea
Briefly, the rAd particles (0, 10, 20, or 50 MOI) and 100 μM FeCl3 were separately diluted in a serum-free medium. These two solutions were then mixed at a ratio of 1:1 and incubated for 30 min at room temperature. To analyze the effect of FeCl3, the same rAd particle solutions without FeCl3 were prepared as negative controls. The hPDLSCs, precultured in 100-mm dishes (Nunc, Roskilde, Denmark), were washed with phosphate-buffered saline (PBS) and then incubated in each prepared virus-containing solution for 1 h, after which the solutions were replaced with a fresh serum-containing growth medium.
Flow cytometry for EGFP
hPDLSCs/rAd-EGFP transduced at various viral concentrations (0, 10, 20, and 50 MOI) with or without FeCl3 treatment were respectively harvested after a 24-h culture period using 0.5 mM EDTA (Gibco). After washing twice with a minimal serum-containing staining buffer (BD Biosciences), the fluorescent expression of EGFP was analyzed in each group using a flow cytometer (FACSCalibur; BD Biosciences).
Analysis of BMP2 secretion and cellular activity
BMP2 secretion was analyzed quantitatively using an Enzyme-Linked Immunosorbent Assay (ELISA) Kit (Quantikine; R&D Systems, Minneapolis, MN) according to the manufacturer's instructions. For this purpose, hPDLSCs/rAd-BMP2 transduced at various viral concentrations (0, 10, 20, and 50 MOI) with or without FeCl3 treatment were cultured. The culture medium was refreshed on days 0 and 2, and the culture supernatants were collected on days 1 and 3.
An additional analysis of cellular activity was performed in parallel with the ELISA test using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) assay (Amresco, Solon, OH). The hPDLSCs/rAd-BMP2 were prepared in the same way as described above for the BMP2 ELISA test, and then treated with the MTT solution (0.5 mg/mL; Amresco) for 3 h followed by dimethyl sulfoxide (Amresco) for 30 min. The optical densities were measured at 540 nm using a microplate reader (Molecular Devices, Sunnyvale, CA).
In vivo transplantation of a block-type scaffold loaded with BMP2, hPDLSCs, or hPDLSCs/rAd-BMP2 in a rat calvarial defect model
Sprague-Dawley rats (n = 40) weighing 230–300 g were used in this study. The surgical procedures and experimental protocols for animal care applied in this study were approved by the Institutional Animal Care and Use Committee of the Inha University, Incheon, Korea (Approval No. Inha 120516–141). The cylinder-shaped BCP block scaffolds (MBCP+, Biomatlante; 3 mm in height and 8 mm in diameter) were prepared before the grafting procedure. The scaffolds were assigned to one of five experimental groups according to the transplantation material used, as follows: BCP scaffold control for group 1 (Gr1; negative control), scaffold containing rhBMP2 (Cowellmedi, Pusan, Korea) for group 2 (Gr2; positive control), scaffold loaded with normal hPDLSCs for group 3 (Gr3), scaffold combined with both normal hPDLSCs and rhBMP2 for group 4 (Gr4); and scaffold loaded with hPDLSCs/rAd-BMP2 for group 5 (Gr5). The scaffolds were soaked in PBS without or with 5 μg of rhBMP2 overnight at 37°C. The hPDLSCs (Gr3 and Gr4) and the hPDLSCs/rAd-BMP2 (Gr5) were correspondingly loaded onto the prepared scaffolds (1 × 106 cells/scaffold) and incubated for 2 h at 37°C before the surgery. A circular critical-size calvarial bone defect with a diameter of 8 mm was created using a trephine drill (ACE Surgical Supply Company, Brockton, MA). The prepared scaffolds were then grafted onto the defect. The rats were euthanized at 2 or 8 weeks after the graft procedure for further histological and immunohistochemical assessment.
Scanning electron microscopy
BCP block materials consisting of hydroxyapatite and β-tricalcium phosphate at a ratio of 20:80 were used as scaffold (MBCP+; Biomatlante, Vigneux de Bretagne, France; 3 mm in height and 8 mm in diameter) in this study. After the scaffolds were soaked in PBS overnight at 37°C, hPDLSCs and hPDLSCs/rAd-BMP2 (1 × 106 cells/scaffold) were then respectively loaded onto the prepared scaffolds and incubated for 2 h at 37°C. These block materials were observed with the aid of a scanning electron microscope (SEM, S-4200; Hitachi, Tokyo, Japan).
Histology and histomorphometric analysis
The calvarial specimens were fixed in 4% formalin for 1 day and then decalcified with 10% EDTA (pH 7.4). After multiple dehydration cycles in ethanol at various concentrations, the specimens were embedded in paraffin. The central-most parts of the cylindrical specimens were then cut into longitudinal sections (thickness: 5 μm) and stained with Hematoxylin and Eosin. The histological sections were observed using a light microscope (Axio Scope A1; Carl Zeiss, Oberkochen, Germany).
Histomorphometric analyses were performed using a software-based, semiautomatic, digital image analysis system (Image-Pro Plus; Media Cybernetics, Silver Spring, MD). Each slide from each of the five groups was examined to identify and measure the area occupied by newly formed bone (NB) and the total augmented area (AG). The quotient resulting from dividing NB by AG was calculated and is expressed as a percentage.
Immunohistochemistry
The same 5-μm-thick sections used for the histological analysis were deparaffinized and treated with 10 mM sodium citrate (pH 6.0) at 90–95°C for 10 min to retrieve antigens from the cytoplasm of the cells. Nonspecific staining was blocked by sequentially incubating the sections in a diluent solution (Invitrogen) for 1 h and then in 0.2% Tween 20 (Amresco) in PBS for 20 min at room temperature. Incubation overnight at 4°C followed, with rabbit anti-human or anti-rat antibodies specific to each of the following biomarkers: human mitochondrial ribosomal protein L11 (hMito; Abcam, Cambridge, United Kingdom), BMP2 (Abcam), osteocalcin (OCN; Santa Cruz Biotechnology, Santa Cruz, CA), and Chordin (Novus Biologicals, Littleton, CO). On the following day, the sections were incubated with fluorescein-isothiocyanate-conjugated goat anti-rabbit immunoglobulin G (Santa Cruz Biotechnology) as the secondary antibody. Counterstaining was performed with a fluorescent mounting medium containing 4′,6-diamidino-2-phenylindole (DAPI; Vector Laboratories, Burlingame, CA). The immunofluorescently labeled sections were observed using a confocal laser scanning microscope (Leica Microsystems, Heidelberg, Germany).
Transwell assay of hPDLSCs or hPDLSCs/rAd-BMP2 with hBMSCs during osteogenic differentiation
The protocol used in this study for hBMSC isolation, identification, and cultivation has been described previously. 17 A transwell coculture was performed to evaluate the paracrine effects of hPDLSCs or hPDLSCs/rAd-BMP2 on hBMSCs. An upper chamber was separated from a lower chamber (six-well plate) by a semipermeable membrane with a pore diameter of 0.4 μm (Fig. 1). The lower chamber comprised hBMSCs cultured in osteogenic differentiation medium (ODM), which comprised normal growth medium (NGM) to which 10−8 M dexamethasone (Sigma-Aldrich) and 1.8 mM KH2PO4 (Sigma-Aldrich) had been added. The ODM was refreshed at 3-day intervals. The upper chamber was assigned to one of the five experimental groups corresponding to those used for the in vivo experiment: control (Gr1; negative control), rhBMP2 (Gr2; positive control), hPDLSCs (Gr3), rhBMP2 and hPDLSCs (Gr4), and hPDLSCs/rAd-BMP2 (Gr5). hPDLSCs at passage 4 and hBMSCs at passage 13 were used. The hPDLSCs and hPDLSCs/rAd-BMP2 in the upper chamber were first cultured in NGM, and then switched to ODM at the start of the coculture experiment with the lower chamber. Two of the groups, Gr2 and Gr4, were subdivided into three subgroups each (Gr2/4-a, -b, and -c), which were provided with rhBMP2 at different concentrations: Gr2/4-a, 50 ng/mL; Gr2/4-b, 125 ng/mL; and Gr2/4-c, 250 ng/mL.
Mineralized nodule formation
After 2 and 4 weeks of coculture, the mineralized nodules formed by hBMSCs in the lower chamber of each group were stained and quantified for comparison. The nodules were stabilized by applying 4% paraformaldehyde for 10 min, and then stained using 2% Alizarin Red S (pH 7.2; Sigma-Aldrich). After washing with PBS, the stained nodules were treated with 10% cetylpyridinium chloride (2 mL/well) overnight to extract the staining agent. The optical densities of each group were then measured at 540 nm using a microplate reader (Molecular Devices).
Alkaline phosphatase activity
To confirm the results of the preceding mineralized nodule formation experiment, a supplementary examination of alkaline phosphatase (ALP) activity of the hBMSCs was performed after 1, 3, 7, and 14 days of coculture. In brief, the cells were lysed with Triton X-100 (Sigma) for 30 min, after which centrifugation was performed at 13,000 rpm for 10 min to collect the supernatant. This sample was then added into an ALP test solution (Sigma) and incubated for 5 min at 37°C according to the manufacturer's instructions. The optical density of each group was read at 405 nm using a microplate reader (Molecular Devices). A more accurate comparison was shown by the ALP activity values of each group relative to their respective total amount of protein (i.e., the ALP activity per mg protein). The total protein amount was measured using a protein assay kit (BCA Protein Assay Kit; Pierce Biotechnology, Rockford, IL) according to the manufacturer's instructions.
Statistical analyses
The data retrieved during this study were evaluated statistically using the Wilcoxon rank-sum test and Kruskal–Wallis test with SPSS software (version 19.0; SPSS Predictive Analytics, Chicago, IL). Tukey's post-hoc multiple-comparison tests were also performed to reveal significant differences between groups. The cutoff for statistical significance was set at p < 0.05.
Results
Transduction efficiency
First of all, we examined whether FeCl3 treatment enhances the adenoviral transduction of PDLSCs since Fe3+ has been reported to enhance the viral transduction efficiency of MSCs. 18 The EGFP fluorescence was significantly higher for rAd-EGFP-transduced hPDLSCs (hPDLSCs/rAd-EGFP) treated with FeCl3 than for those without FeCl3 treatment at all viral concentrations, except the control group (0 MOI). The EGFP fluorescence of hPDLSCs/rAd-EGFP significantly increased in a viral dose-dependent manner, indicating an efficient transduction (Fig. 2A). The level of BMP2 expression was significantly higher for hPDLSCs/rAd-BMP2 treated with FeCl3 than for those without FeCl3, in all viral doses, except the control. In the presence of FeCl3, the increasing doses of rAd-BMP2 up to 20 MOI elevated accumulation of BMP2 released from the transduced cells up to 3 days. However, the highest dose of 50 MOI significantly reduced BMP2 release (Fig. 2B). In the absence of FeCl3, normal cellular activity such as cell proliferation was not affected by viral transduction in hPDLSCs/rAd-BMP2 during culture period. However, an overall reduction in cellular activity was observed in the FeCl3-treated hPDLSCs/rAd-BMP2. Higher viral doses caused more reduction in cell viability; statistically significant change was observed at 20 and 50 MOI (Fig. 2C). Although some cellular toxicity was observed in hPDLSCs/rAd-BMP in the presence of FeCl3, we chose the viral dose of 20 MOI plus FeCl3 treatment as the optimal condition for efficient BMP2 production in adenovirally transduced hPDLSCs; this condition was used for the subsequent experiments in this study.
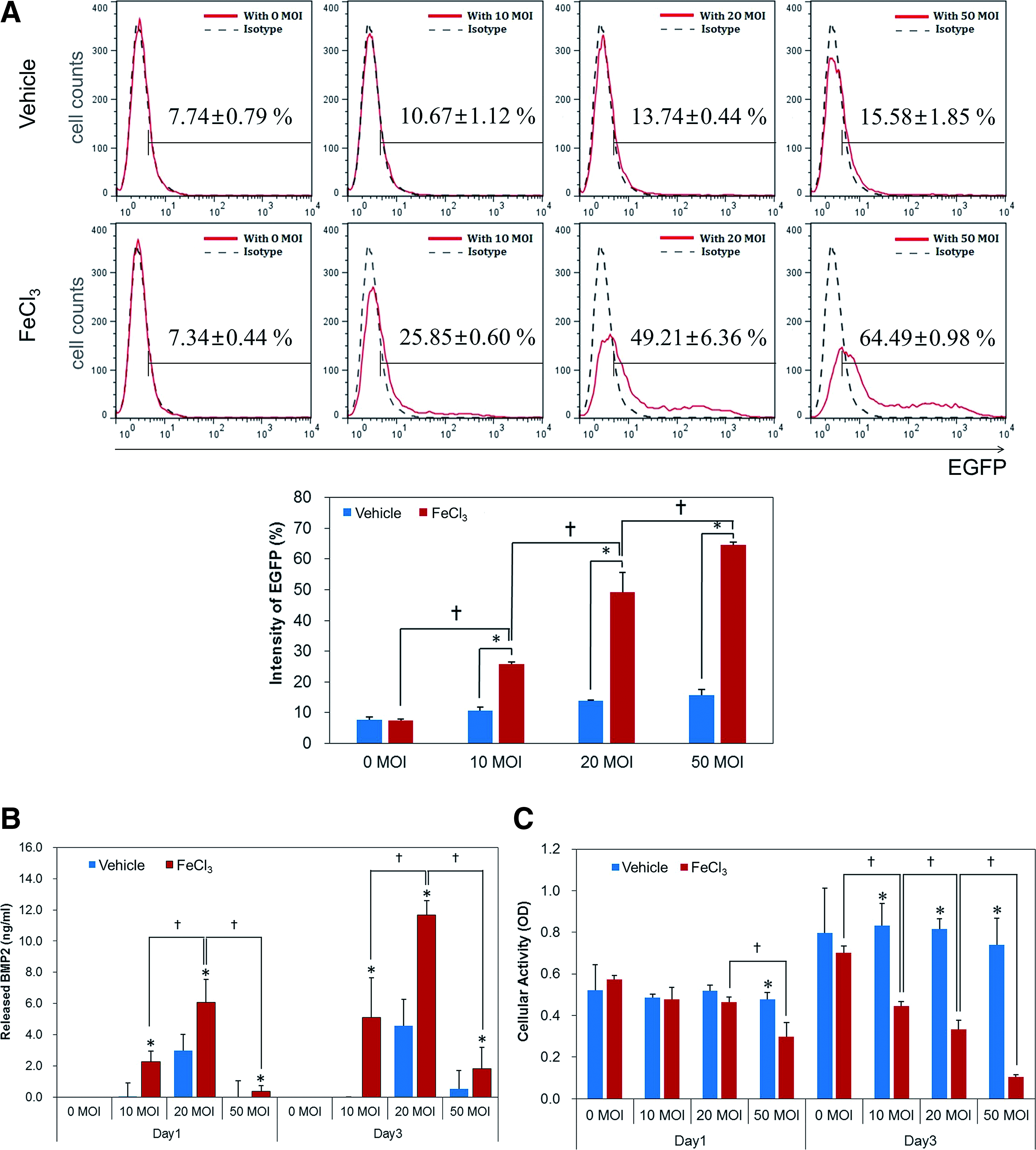
Transduction efficiency of hPDLSCs/rAd-BMP2 for various rAd concentrations.
In vivo transplantation of hPDLSCs/rAd-BMP2 and evaluation of new bone formation
We evaluated in vivo effects of hPDLSCs/rAd-BMP2 loaded with BCP scaffolds after transplantation in a rat calvarial bone defect model. Experimental groups were as follows: BCP scaffold control for Gr1, scaffold containing rhBMP2 for Gr2, scaffold loaded with normal hPDLSCs for Gr3, scaffold combined with both normal hPDLSCs and rhBMP2 for Gr4, and scaffold loaded with hPDLSCs/rAd-BMP2 for Gr5. The SEM images of the BCP scaffold revealed a surface with well-attached cells in both cell groups (hPDLSCs/rAd-BMP2 and hPDLSCs). These cells were in direct contact with the surface, and already became flat and spread out on the surface after 2 h of loading (Fig. 3A). After 2 weeks of recovery, histological analysis of each scaffold showed that mineralization was found (Fig. 3B, C) at the basal part of the scaffold facing the defect and aligned along the inner walls of the empty spaces. The mineralized areas appeared to be a part of the osteoid with low degree of mineralization, which were difficult to distinguish from the collagenous tissue around. The remaining areas were filled with connective tissue occupied by fibroblasts, blood vessels, and polymorphonuclear leukocytes (PMNs). The degree of mineralization was significantly greater in Gr2 sections than that in the other groups (Fig. 4), although it was mostly visible as rather thin bands along the inner walls of the scaffold. No significant difference in mineralization was observed among Gr1, Gr3, Gr4, and Gr5 (Fig. 4). At 8 weeks, the overall mineralization increased in both quantity and maturity, and the histological characteristics of each group became more distinguishable. The new bone formation was the highest in Gr2, followed by Gr1 and Gr4, and significantly less in Gr3 and Gr5. Mineralization with embedded osteocytes was restricted to the basal half of the scaffold, except for Gr2, in which it was spread throughout the scaffold. However, in Gr2, most of the available space inside the scaffold was occupied by bone marrow, so that the actual mineralized bone tissue surrounding it formed only a thin band along the inner walls of the scaffold. The bone marrow was packed with adipose tissue containing adipocytes with characteristic lipid droplets and eccentric nuclei. This adipose tissue was surrounded by connective tissues that were abundant in immune cell infiltrates. Similar histology was shown in Gr4, although with less pronounced adipose tissue and fewer PMNs. Fewer PMNs were also observed in Gr3 and Gr5 as compared to Gr2. In Gr5, the newly formed bone appeared to be more compact and a minimal marrow and adipose tissues were found in it (Figs. 3B, C, and 4).

Histological and histomorphometric evaluation following transplantation of hPDLSCs or hPDLSCs/rAd-BMP2 in vivo.
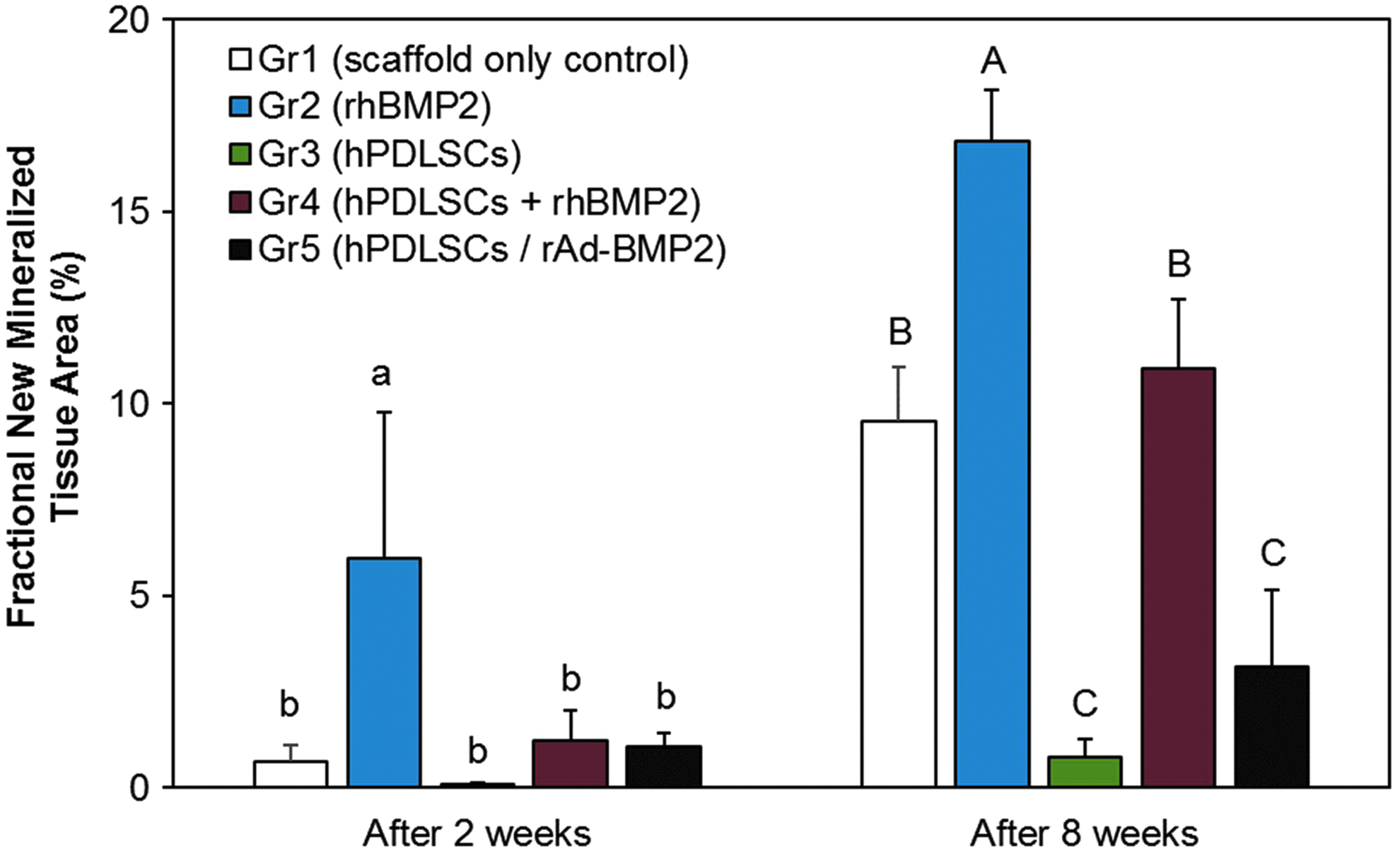
Histomorphometric analysis of in vivo new bone formation Data displaying the histomorphometric measurements of newly formed mineralized tissue in each group at 2 and 8 weeks of healing. Gr2 exhibited significantly more mineralization than any other group. At 8 weeks, the levels of new bone formation were significantly greater in Gr1 and Gr4, compared to those of Gr3 and Gr5. Alphabetic letters represent statistically homologous subgroups at a significance level of p < 0.05. Color images available online at www.liebertpub.com/tea
To demonstrate whether transplanted hPDLSCs survive and contribute to the new bone formation in vivo, their biodistribution was assessed by the existence of human mitochondria by using human mitochondria-specific marker, hMito. Immunofluorescence staining results showed that hMito was evident in Gr3, Gr4, and Gr5 at 2 and 8 weeks (Fig. 5A, B). BMP2 was detected in all groups at 2 weeks, with the staining being equally pronounced in all groups. At 8 weeks, Gr3 was negative for BMP2, whereas the other groups were positive for it (Fig. 5B). OCN at 2 weeks was highly expressed in Gr1 and Gr2, but weakly expressed in Gr4 (Fig. 5A). No OCN expression was observed in Gr3 or Gr5. At 8 weeks, all groups displayed positive staining for OCN with comparable fluorescence intensities. Chordin was weakly expressed in Gr3 and Gr4, but was highly expressed in Gr5 at 2 weeks (Fig. 5A). At 8 weeks, Chordin expression was evident in Gr3, Gr4, and Gr5 with comparable fluorescence intensities (Fig. 5B).
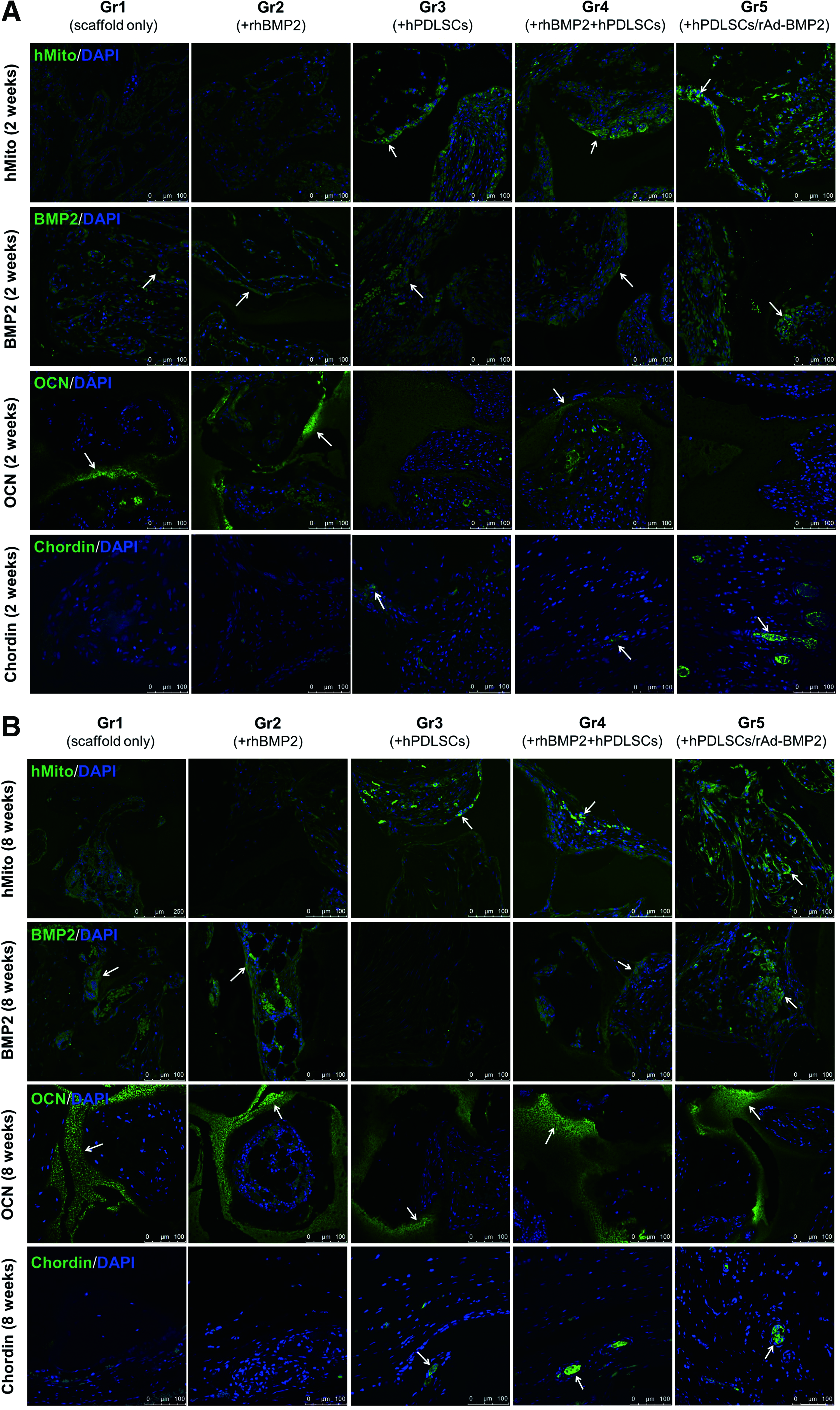
Immunohistochemistry of the transplants. The presence of various marker proteins was evaluated in the transplanted specimens obtained after 2 and 8 weeks of healing.
Transwell assay of hPDLSCs or hPDLSCs/rAd-BMP2 with hBMSCs during osteogenic differentiation
Since our data suggested the inhibitory potential of hPDLSCs in rhBMP2-mediated osteogenesis in vivo, we further investigated whether hPDLSCs negatively regulate bone formation. To this end, we set up an in vitro transwell assay in which hPDLSCs were seeded in the upper chamber and hBMSCs with high osteogenic potential were seeded in the lower chamber. We examined whether PDLSCs reduce bone nodule formation by hBMSCs during osteogenic differentiation. The intensity of Alizarin Red S stain on mineralized bone nodules from rhBMP2-treated hBMSCs were increased both in a 2-week culture and in a 4-week culture (Fig. 6A). When osteogenically induced, hBMSCs successfully produced mineralized bone nodules in response to rhBMP2. The numbers and areas of mineralized nodules were increased with increasing doses of rhBMP2 (50, 125, and 250 ng/mL). However, in Gr4, an overall reduction in the intensity of Alizarin Red S stain was evident in response to rhBMP2 in both culture periods. This decrease was especially pronounced at high rhBMP2 concentrations of up to 250 ng/mL in the presence of hPDLSCs. When hPDLSCs were cocultured in the upper chamber, the osteoinductive activity of hBMSCs in the lower chamber disappeared. Likewise, when hPDLSCs/rAd-BMP2 were cocultured with hBMSCs, mineralized bone nodules identified by Alizarin Red S staining were not observed. There were no significant difference in mineralized nodule formation between Gr3 and Gr5 (Fig. 6A). ALP activity was also measured at up to 14 days (Fig. 6B). The ALP activity of hBMSCs was significantly increased with increasing doses of rhBMP2 at day 14 (Gr2). However, in Gr4, no significant change in the ALP activity was found among three doses of rhBMP2 in the presence of PDLSCs. Compared to Gr2, the ALP activity was dramatically reduced in Gr4, which was consistent with the Alizarin Red S staining results. There was no significant difference in the ALP activity between Gr1 and Gr3. (Fig. 6B).

Transwell assay of hPDLSCs or hPDLSCs/rAd-BMP2 with hBMSCs during osteogenic differentiation.
Discussion
The first part of the investigation focused on the rAd concentration for optimal transduction of hPDLSCs for high-yield production of BMP2 from transduced cells. The rAd used in this study was preconditioned with FeCl3 to enhance the transduction efficiency. Ferric ions (Fe3+) have been reported to significantly enhance the rAd-mediated transduction of MSCs by diminishing the electrostatic repulsion caused by the negative surface charges of both the rAd and the MSCs.18,19 Consistent with the previous reports, we showed that the transduction of BMP2 and its release from the cells were significantly enhanced with FeCl3 treatment. The optimal condition we selected for rAd-mediated transduction of BMP2 in hPDLSCs was MOI 20 and FeCl3 treatment, which was used for all transduction experiments in this study.
Given that the choice of study design can significantly affect the results, we chose a harsh environmental condition for evaluating in vivo bone formation using a BCP block. Although the BCP scaffold has already been proven to be biocompatible and effective as a BMP2 carrier in onlay graft indications,3,6 some limitations have also been reported. Limited resorption in vivo and remodeling capability relative to a faster new bone formation along the BCP scaffold may obstruct cell migration and new bone formation inside it.3,20 New bone formation inside the BCP scaffold is therefore more likely affected by the cells and the growth factors loaded, compared to particulate graft sites, in which the predominant influence of neighboring endogenous cells and bioactive factors on bone formation may be misinterpreted as an effect directly induced by the loaded factors. Hence, this experimental model may provide more distinguishable and reliable results depending on the study concepts.
Previously, we reported that hPDLSCs/rAd-BMP2 produced the most extensive new bone formation when subcutaneously transplanted in mice. 13 Unexpectedly, Gr5 containing hPDLSCs/rAd-BMP2 did not effectively regenerate bone in quantity in the critical-size calvarial defect model (Figs. 3 and 4). Likewise, hPDLSC-loaded groups, including Gr3 and Gr4, also exhibited significantly less new bone formation compared to hPDLSC-free Gr1 and Gr2. The reduced mineralization in Gr3, Gr4, and Gr5 suggests the inhibitory regulation of the osteogenic environment possibly caused by soluble factors released from hPDLSCs or hPDLSCs/rAd-BMP2 (Fig. 6). It has been shown that hPDLSCs produce negative regulators of osteogenesis, such as Chordin and PDL-associated protein-1 or asporin (PLAP1/asporin).14,15,20,21 Mizuno et al., demonstrated that gene expression profile of MSCs is influenced by soluble factors released from hPDLSCs when both cell types are cocultured. 14 Among the soluble factors, Chordin was suggested to specifically bind BMP2, thereby suppressing the BMP2-induced mineralization of MSCs. Our results also showed that Chordin was expressed in hPDLSC-loaded groups, including Gr3, Gr4, and Gr5 (Fig. 5). It has been demonstrated that BMP2 upregulates PLAP-121 and its overexpression interferes with both natural and BMP2-induced mineralization in the presence of PDL cells. 22 Consistent with these findings, both the mineralization of nodules and ALP activity of rhBMP2-treated hBMSCs were significantly reduced in the presence of hPDLSCs (Fig. 6). Moreover, fewer mineralized nodules were formed when hBMSCs were cocultured with hPDLSCs/rAd-BMP2.
The cellular heterogeneity may also contribute to the reduction in mineralization of Gr5 (Figs. 4 and 6). The transduction efficiency for the hPDLSCs/rAd-BMP2 was only 49.24% ± 6.36% (mean ± SD), indicating that the other 50% of the cells were not transduced. Since these nontransduced hPDLSCs proliferate more highly than the transduced cells, it is likely that nontransduced cells predominate in the population over time, resulting in less osseous differentiation. Although it seems unlikely that these nontransduced hPDLSCs significantly affect the overall results in this study, they may be worthy of further investigation and discussion.
Limited recovery time up to 8 weeks in the in vivo transplantation experiments may provide another plausible explanation for the reduced mineralization in Gr5. The immunohistochemical results indicate that the mineralization process was slower in the hPDLSC-loaded groups (Gr3, Gr4, and Gr5) than in the hPDLSC-free groups (Gr1 and Gr2). At 2 weeks of recovery, Gr1 and Gr2 exhibited strong fluorescent signals for OCN, whereas weak fluorescent intensity was detected in Gr4 (Fig. 5). These fluorescent signals with a comparable intensity were shown at 8 weeks of recovery. Expression of OCN, an osteoblast marker, is known to be accumulated at the late stage of osteogenic differentiation. 23 It is speculated that the mineralization is faster in the groups with no hPDLSCs, whereas those loaded with hPDLSCs slowly develop their osteogenic milieu. Accordingly, rhBMP2 in Gr4 at 2 weeks of healing could have slightly compensated for this slow development, exhibiting some weak signals of OCN expression.
A possible immunologic response caused by human stem cells transplanted in rats was excluded when planning for in vivo experiment. The rationale for this exclusion was provided by several studies that have investigated the immunomodulatory properties of MSCs12,24 and PDLSCs, 25 and their in vivo effects through allogeneic or xenogeneic administration in rats, rabbits, or dogs.26–30 None of the animals tested reportedly exhibited significant adverse immunoreactions. Moreover, we found a number of PMN only in Gr2, but a few of immune cell infiltrates in hPDLSC-loaded Gr3 and Gr5, consistent with our previous study. 13 We also detected hMito in all hPDLSC-transplanted groups (Gr3, Gr4, and Gr5) at both 2 and 8 weeks, indicating that hPDLSCs survived in the recipients even after 8 weeks of transplantation.
The histological specimens in this study displayed similar patterns to those found previously.13,31 Although the amount of new bone formation was highest in Gr2, Gr2 also exhibited a specific histology characterized by abundant adipose tissue in the marrow space, with only a relatively thin layer of new bone surrounding it. Such findings are consistent with several studies demonstrating that BMP2 restrains the formation of collagen,32,33 but promotes adipogenesis.34,35 In contrast, well-organized new bone formation without adipose marrow spaces or immune cell infiltrates was found in Gr5 (Fig. 3C).
Bone defects during tissue repair and differentiation into actual bone tissue are fundamentally affected by various reactive factors and the extracellular matrix components that communicate with each other to ultimately regulate the local regeneration process. 36 The importance of such a microenvironment in stem cell-regulated tissue regeneration has been suggested.23,37,38 hPDLSCs originate from the periodontal attachment apparatus, where they communicate with stromal cells to regulate alveolar bone metabolism. Since there is a fundamental difference between ex vivo-expanded stem cells and endogenous cells, genetically engineered hPDLSC-mediated regeneration of bone tissue requires further investigations to control the local microenvironment. However, the interaction and regulation between hPDLSCs and the microenvironment of their transplants remain unclear yet.
Meanwhile, our previous and present studies suggest that the osteoinductive potential of hPDLSCs varies depending on the experimental models; hPDLSCs promoted BMP2-induced osteogenesis in subcutaneous transplantation, 13 but negatively regulated bone formation in a critical-size bone defect. These pleiotropic effects of hDPLSCs on in vivo osteoinduction imply the possible risks of simple application of stem cells according to their in vitro stem cell properties. Our findings indicate that more extensive investigations in a variety of pathophysiological conditions are required before clinical applications.
In this study, we attempted to elucidate the regulatory function of hPDLSCs and the potential of hPDLSCs/rAd-BMP2 as a carrier for bone regeneration in osteogenic milieu. We found that the in vivo bone regenerative potential of BMP2-overexpressing hPDLSCs could be compromised in a critical-size rat calvarial bone defect model, suggesting the complexity of osteoinductive function of hPDLSCs depending on in vivo tissue environment. Further investigation is needed to better understand the regenerative properties of hPDLSCs and to make their practical use for tissue repair and tissue engineering.
Footnotes
Acknowledgments
The authors are grateful to Young-Chul Sung at Pohang University of Science and Technology (POSTECH) for his kind gift of replication-deficient recombinant adenoviral vector (rAd) expressing EGFP and BMP2, and to Cowellmedi for providing rhBMP2. A special appreciation is dedicated to Dr. Im-Hee Jung for providing excellent graphic works. This research was supported by a Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2012R1A1A2006818) and the Ministry of Science, ICT & Future Planning (NRF-2015R1A2A2A01004589).
Disclosure Statement
No competing financial interests exist.