
Abstract
To utilize three-dimensional (3D) engineered human skeletal muscle tissue for translational studies and in vitro studies of drug toxicity, there is a need to promote differentiation and functional behavior. In this study, we identified conditions to promote contraction of engineered human skeletal muscle bundles and examined the effects of transient inhibition of microRNAs (miRs) on myogenic differentiation and function of two-dimensional (2D) and 3D cultures of human myotubes. In 2D cultures, simultaneously inhibiting both miR-133a, which promotes myoblast proliferation, and miR-696, which represses oxidative metabolism, resulted in an increase in sarcomeric α-actinin protein and the metabolic coactivator PGC-1α protein compared to transfection with a scrambled miR sequence (negative control). Although PGC-1α was elevated following joint inhibition of miRs 133a and 696, there was no difference in myosin heavy chain (MHC) protein isoforms. 3D engineered human skeletal muscle myobundles seeded with 5 × 106 human skeletal myoblasts (HSkM)/mL and cultured for 2 weeks after onset of differentiation consistently did not contract when stimulated electrically, whereas those seeded with myoblasts at 10 × 106 HSkM/mL or higher did contract. When HSkM were transfected with both anti-miRs and seeded into fibrin hydrogels and cultured for 2 weeks under static conditions, twitch and tetanic specific forces after electrical stimulation were greater than for myobundles prepared with HSkM transfected with scrambled sequences. Immunofluorescence and Western blots of 3D myobundles indicate that anti-miR-133a or anti-miR-696 treatment led to modest increases in slow MHC, but no consistent increase in fast MHC. Similar to results in 2D, only myobundles prepared with myoblasts treated with anti-miR-133a and anti-miR-696 produced an increase in PGC-1α mRNA. PGC-1α targets were differentially affected by the treatment. HIF-2α mRNA showed an expression pattern similar to that of PGC-1α mRNA, but COXII mRNA levels were not affected by the anti-miRs. Overall, joint inhibition of miR-133a and miR-696 accelerated differentiation, elevated the metabolic coactivator PGC-1α, and increased the contractile force in 3D engineered human skeletal muscle bundles.
Introduction
C
Muscle maturation can be promoted by transient regulation of the level of microRNAs (miRs), short (20–22 nt) noncoding RNA strands that act through posttranscriptional regulation of genes.7,8 Several microRNAs are extremely important in the regulation of muscle proliferation, and differentiation and their overexpression or inhibition can have profound effects on myogenesis.9,10 MiR-1 and miR-206 promote myoblast differentiation, and miR-133a promotes proliferation. Koning et al. demonstrated increased myoblast differentiation when miR-1 and miR-206 were transiently overexpressed in primary human skeletal myoblasts (HSkM). 9 We previously showed that inhibiting miR-133a alone in C2C12 murine myoblasts 10 and primary HSkM 11 significantly accelerated differentiation. Aoi et al. identified stretch-sensitive miR-696, which regulates the expression of an important and potent metabolism coactivator, PGC-1α, and observed heightened levels of PGC-1α protein levels in anti-miR-696 C2C12 samples 24 h after transfection. 12 Heightened expression of PGC-1α, which is encoded by the gene PPARGC1A, 13 has been linked to increased oxidative energy metabolism and type I (slow twitch) myofiber formation.13–15
Recently, we extended our results with murine C2C12 cells and showed that transient inhibition of miR-133a alone accelerated myogenesis in two-dimensional cultures of primary human myoblasts. 11 In this study, we tested the hypothesis that jointly inhibiting miR-133a and miR-696 simultaneously accelerated differentiation in 2D cultures and engineered 3D human skeletal muscle tissue (myobundles) with increased type I myosin heavy chain (MHC) expression compared to cells transfected with a scrambled sequence. Regulating the MHC expression and extent of maturation provides better control of the engineered muscle tissue. 2D cultured muscle was used to study proteins and gene expression in response to miR inhibition to enable comparison with other published work on miRs and assess similarities and differences with 3D culture. We measured changes in contractile forces production, fiber type, and metabolic genes following miR inhibition in a more native-like environment containing tension and fiber alignment.
Methods and Materials
Primary HSkM isolation
Primary HSkM were isolated from biopsies of the vastus lateralis of healthy middle-aged volunteers under conditions of informed consent and approval of the Duke Investigational Review Board as presented previously. 11 One 30–50 mg biopsy yielded 15–20 million cells. Four isolations of HSkM from separate individuals were used for these studies. Due to differential adhesion, the average purity was 88.0% ± 6.1% as assessed by desmin staining of four donors after thawing frozen cultures.
Cell culture
Primary HSkM cultures were expanded as described previously.
11
Myoblasts seeded at 1,500 cells/cm2 in uncoated tissue plastic flasks were used at passages 3–6. A growth medium (GM) containing low glucose (1 g/L
MicroRNA transient transfection
Single-stranded antisense microRNA molecules specific for the miR-133a (UUUGGUCCCCUUCAACCAGCUG; Applied Biosystems) and miR-696 (GCGUGUGCUUGCUGUGGG; Applied Biosystems) sequence were used with siPORT NeoFX Transfection Agent (Ambion) to transfect cells. Anti-miR Negative Control #1 (Applied Biosystems), a single-stranded random microRNA sequence, was used as the control. Myoblasts were transfected at ∼80% confluence. siPORT NeoFX Transfection Agent was diluted with OptiMEM I (Gibco) and mixed with 50 nM of single-stranded anti-miR (anti-miR-133a, anti-miR-696, or anti-miR-negative control) as previously described. 11 Transfection mixtures were allowed to incubate in tissue culture flasks at 37°C for 10 min, then added to cells with the GM. Cells were transfected at 37°C and 5% CO2 for 18 h, after which cells were rinsed, trypsinized, and myobundles prepared. To assess the transfection efficiency and half-life of the anti-miR, Cy3 and FAM dye-labeled anti-miR Negative Control #1 (Applied Biosystems) were used as described above. Transfected HSkM containing Cy3 or FAM labels were viewed under fluorescence and quantified as the proportion of total number of cells in the field view.
Engineered human muscle bundle construction
3D myobundles were constructed using 45 μL polydimethylsiloxane (Dow Corning Corporation) molds similar to our previously described methods.16,17 Velcro was used as anchor points at either end of the mold. A hydrogel mixture (45 μL per myobundle) of 20% Matrigel (BD Biosciences), 4 mg/mL fibrinogen (Sigma), 50 U/mL thrombin (Sigma), and passage 5–7 HSkM (5, 10 or 15 × 106 cells/mL) gelled at 37°C and 5% CO2 for 1 h before addition of GM. At 48 h postseeding, all gels had compacted and appeared opaque. At this time, the GM was exchanged for the DM (day 0); the DM was exchanged every 48 h for the 2-week static culture period.
Immunofluorescence
To assess for the presence of striations and type I and II MHC, samples cultured for 14 days postdifferentiation were fixed and immunostained as previously described. 11 The following primary antibodies were used: sarcomeric α-actinin (SAA, 1:800; Sigma), slow MHC (1:300; AbCam ab11083, reacts only with human adult type I MHC), and fast MHC (1:300; AbCam ab91506, reacts with human and mouse type IIA, IIX/B MHC). Confocal images (Zeiss model 510) were taken at varying depths throughout the myobundles. Resulting images were assessed with ImageJ software for fiber diameter, percent of nuclei in fibers (fusion index), and fiber type. For fiber diameter measurements, four measurements were taken per field of view, and three fields of view were taken for each experiment. Typically, each field contains 30–50 fibers. Using the ImageJ “Color Pixel Counter” feature, pixels stained for SAA were divided by total pixels per field of view to calculate area fraction of myofibers, which was used to calculate specific force per fiber area.
Quantitative real-time polymerase chain reaction
Real-time polymerase chain reaction (RT-PCR) was performed as described previously. 11 Total mRNA was isolated from 2D cultures 6 and 14 days after onset of differentiation and from 3D myobundles 14 days after onset of differentiation using the Aurum Total RNA Mini Kit (Bio-Rad). RNA concentration and quality were measured using the NanoDrop ND-1000 spectrophotometer (NanoDrop). For RNA expression, cDNA was prepared using the iScript cDNA Synthesis Kit (Bio-Rad) following manufacturer's instructions. Primers for PPARGC1A, slow and fast MHC, HIF-2α, COX II, and GAPDH (endogenous control) were custom ordered (IDT, Table 1). The 2−ΔΔCT method was used to assess mRNA expression. Total RNA from 11-day-old mouse embryo was used as the reference (Clontech).
MHC, myosin heavy chain.
Western blot
At day 14, protein was isolated from cells as described previously. 11 15 μg of isolated protein was loaded into each lane on a 12% Ready Gel Tris-HCl Precast Gel (Bio-Rad) for sodium dodecyl sulfate–polyacrylamide gel electrophoresis. Primary antibodies for SAA (1:2500, clone EA-53; Sigma), 60 kDa glycosylated mitochondrial protein (1:2500, MTC02; AbCam), PGC-1α (1:2000; Calbiochem), fast MHC (1:10,000; Abcam), slow MHC (1:1000; Abcam), and MF20 (1:10,000 Developmental Studies Hybridoma Bank, University of Iowa), which recognizes all MHC isoforms and β-actin (1:1000; Santa Cruz Biotechnology), were used to probe the PVDF membranes. Protein expression was normalized to the β-actin protein band.
Measurement of isometric contractile force
Isometric contractile force was determined for myobundles 14 days after onset of differentiation as previously described.16,17 Force output was recorded for electrical stimuli at 1, 5, 10, 20, and 40 Hz at 5.9 V/cm in 5% increments of length from 100% to 120% of initial construct length (8 mm). Force traces were recorded using LabVIEW and exported to Microsoft Excel files. For analysis, Microsoft Excel files were loaded into a custom-made MATLAB program. Photos taken at 100% length before force testing were assessed in ImageJ for myobundle diameter; specific forces were calculated by dividing force output (mN) by cross-sectional area (mm2) assuming a circular cross-section of myobundles.
Statistical analysis
Data are presented as mean ± standard error (SEM) unless otherwise noted. The sample size n refers to the number of myoblast isolations used in the experiment. Each experiment was performed twice with myoblasts from each isolation. Two bundles were averaged together for each datum value at a given condition in a single experiment. The value n reported in the figure legend represents the number of replicate experiments performed at each condition. ANOVA and Fisher's PLSD or Tukey's post hoc tests were performed using STATVIEW 5.0 statistical analysis package. HSkM isolated from separate individuals were considered distinct n values. A value of p < 0.05 was considered significant.
Results
Transfection efficiency and lifetime of microRNAs
No significant difference was observed in the transfection efficiency for HSkM that received Cy3-1 and FAM-labeled negative control anti-miRs (90.6% ± 2.3%) and those that received a single miR (94.4% ± 2.9%; Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/tea). From a polynomial fit of data, single miR transfection had a half-life of 105.2 ± 0.7 h (n = 3) and joint miR transfection had a half-life of 104.1 ± 0.6 h (n = 3), which was not significantly different.
MicroRNA inhibition in 2D cultures accelerates myogenesis and affects PGC-1α
HSkM were transfected with anti-miR-133a, anti-miR-696, both anti-miRs, or a scrambled sequence of microRNAs. HSkM differentiation was compared 14 days after shifting to DM, a duration previously found to produce prominent striations demonstrating tissue maturity. 18 Western blots of SAA protein at day 14 postdifferentiation showed that joint inhibition of miR-133a and miR-696 produced significantly more SAA protein compared to negative control (Fig. 1A, B, *p < 0.05, n = 4). There was no significant difference in total mitochondrial protein (MT) at 14 days postdifferentiation among anti-miR treatments. Consistent with our prior results in mouse and human myotubes,10,11 inhibiting miR-133a promotes myotube maturation.

Sarcomeric α-actinin (SAA) and mitochondria protein (MT) in 2D cultures.
MiR-696 negatively regulates expression of PGC-1α protein 12 and Western blots indicated that all anti-miR treatments significantly increased PGC-1α protein expression compared to negative control (Fig. 1C, D). While treatment of myoblasts with either anti-miR-133a or anti-miR-696 did not affect PGC-1α gene (PPARGC1A) levels relative to negative control as expected for microRNAs that regulate translation, joint inhibition of anti-miR-133a and anti-miR-696 caused a doubling of PPARGC1A levels (p < 0.05, n = 3, Fig. 2), suggesting that additive effects between the two anti-miRs affect gene and protein expression of PGC-1α.

Expression of PPARGC1A gene at day 14 postdifferentiation. Joint transfection samples showed significantly increased expression of PPARGC1A gene compared to negative control. Results were normalized to housekeeping gene GAPDH (*p < 0.05, n = 3). Error bars denote SEM.
We evaluated the downstream effects of anti-miRs by immunostaining for slow and fast MHC, respectively. At 14 days postdifferentiation of 2D cultures, proportion and the intensity of staining of fibers positive for slow MHC (86.9% ± 3.9%) and fast MHC (91.9% ± 2.9%) were not different. Most human skeletal muscle fibers expressed both slow and fast MHC protein isoforms, consistent with in vitro19–21 and in vivo 22 results. Furthermore, there was no significant difference between myofiber density and nuclei in fibers among all conditions.
Engineering of functional 3D human skeletal muscle bundles
Since anti-miRs increased maturation of human myotubes in 2D culture, we hypothesized that 3D tissue-engineered myobundles prepared with anti-miR-treated HSkM would exert greater contraction forces than myobundles made with HSkM receiving a scrambled miR sequence. Since the seeding density of skeletal myoblasts into a 3D scaffold or hydrogel affects maturation of muscle and generation of force,2,23,24 we first examined seeding densities from 5 to 15 × 106 HSkM/mL. While a seeding density of 5 × 106 C2C12/mL was sufficient to produce contractile forces after electrical stimulation, 25 human myobundles seeded with 5 × 106 cells/mL did not elicit electrically induced contractions. Consistent with this result, confocal images of immunostained myobundles showed negligible myoblast fusion and little organization of myofibers (Fig. 3A).
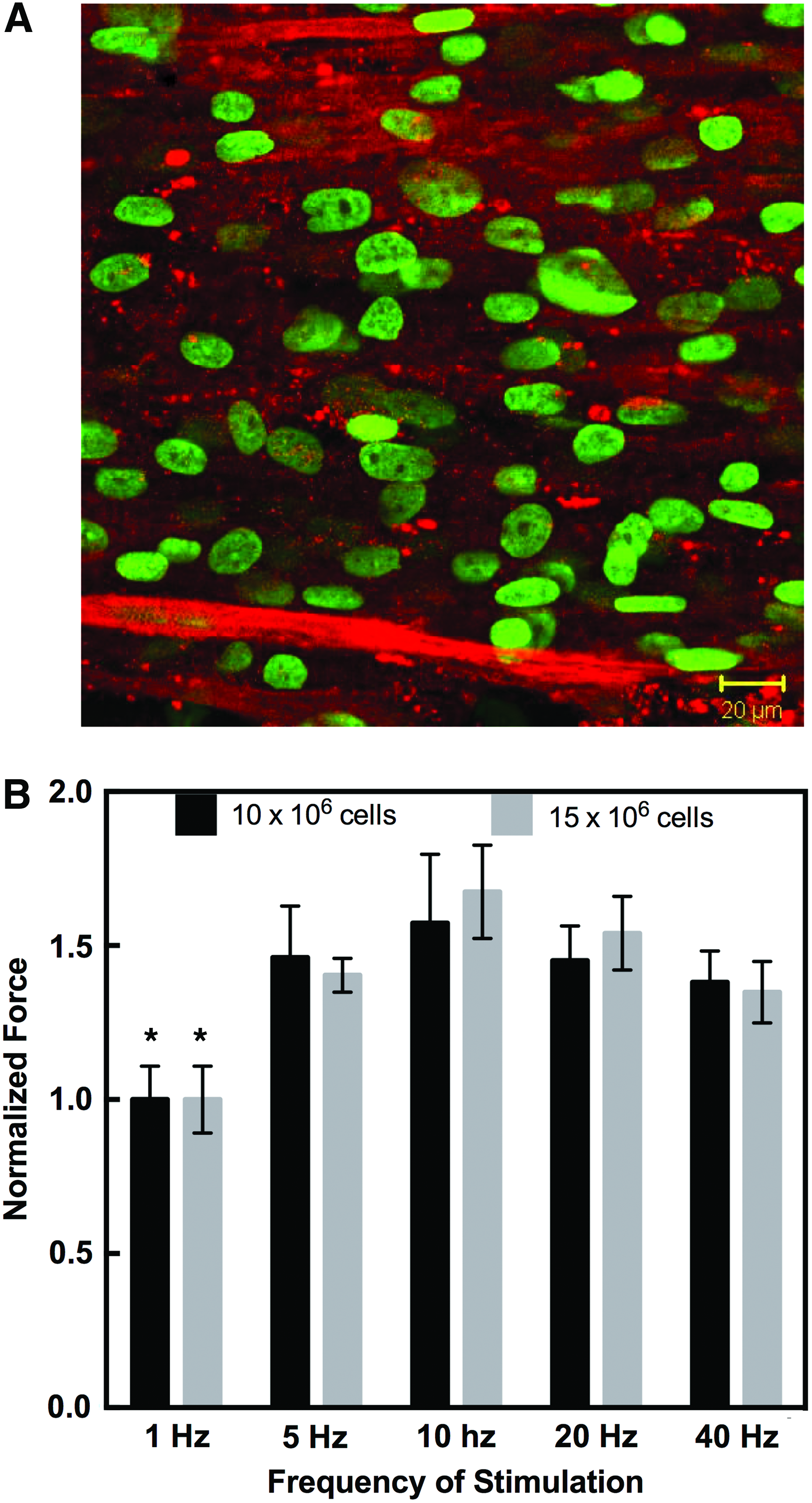
Human skeletal myoblast (HSkM) seeding density for muscle bundles. Bundles were seeded with 5 × 106 HSkM/mL, 10 × 106 HSkM/mL, and 15 × 106 HSkM/mL and cultured statically for 2 weeks.
Myobundles seeded with either 10 × 106 HSkM/mL or 15 × 106 HSkM/mL exhibited fusion and striations of myofibers and an ANOVA showed a significant effect of stimulation frequency on contractile force (Fig. 3B, *p < 0.001, n = 3). A post hoc Tukey test showed that the force at 1 Hz was different than the force at all other conditions, and no differences were detected among the other frequencies. Thus, there appears to be a threshold of HSkM seeding density into fibrin-based muscle constructs that allows for functional force output by engineered muscle. Subsequent studies were conducted with 15 × 106 HSkM/mL in fibrin/Matrigel bundle system.
Structurally, 2-week-old myobundles showed a high density of aligned myofibers containing striations (Fig. 4A–C, striations denoted by arrows) and expressed acetylcholine receptors (AChR) at the periphery of myofibers within the engineered myobundles, which are involved in neuromuscular junction formation (Fig. 4B, C, arrows).
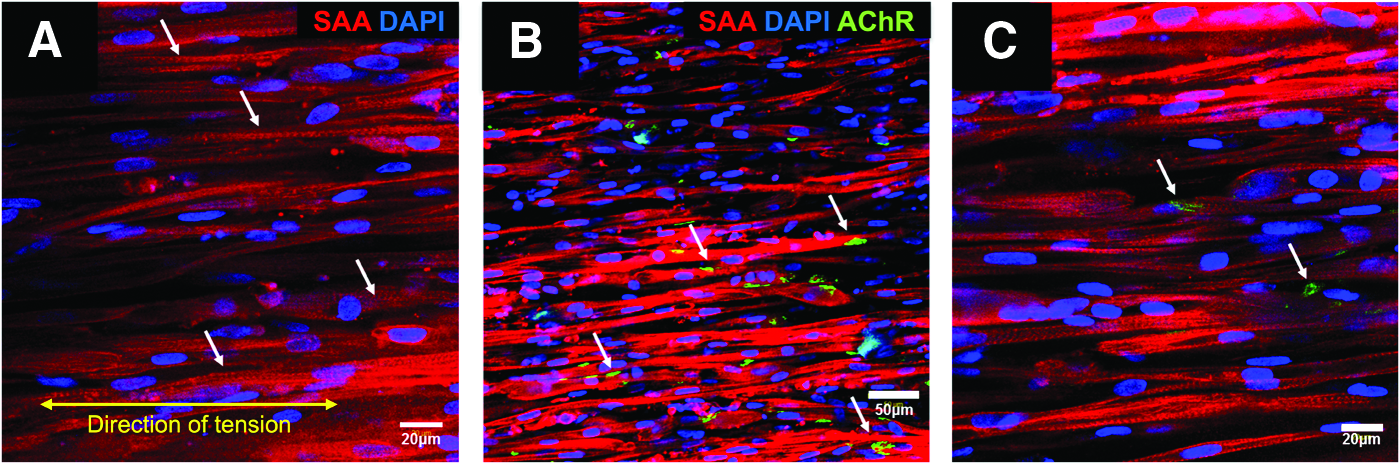
Immunostaining of three-dimensional (3D) human myobundles seeded at 15 × 106 HSkM/mL and examined 14 days after addition of differentiation medium (DM).
Effect of anti-microRNA-133a and/or -696 on structure and function of 3D human myobundles
The anti-miR treatment administered to HSkM before bundle seeding affected the extent of fibrin gel compaction. Specifically, anti-miR-133a myobundles had a 22.4% ± 6.1% smaller diameter than the negative control (Supplementary Fig. S2A, p < 0.05, n = 3). Transfection of HSkM with both anti-miRs produced myobundles with a smaller diameter than myobundles produced with HSkM transfected with negative control (36.8% ± 3.4%, p < 0.005, n = 3) or anti-miR-696 (25.6% ± 3.4%, p < 0.05, n = 3). Interestingly, myofiber diameters did not differ between various miR conditions. Myobundles made with HSkM that received the negative control contained myofibers 14.3 ± 3.7 μm in diameter, whereas treatment with anti-miR-133a, anti-miR-696, or both anti-miRs together produced myofibers with diameters of 16.0 ± 3.4 μm, 13.7 ± 4.4 μm, and 13.9 ± 3.3 μm, respectively.
Myofiber density was significantly higher in anti-miR-133a constructs compared to negative control constructs (p < 0.05, n = 4, Supplementary Fig. S2B), although average percent of nuclei in fibers (fusion index) did not differ among anti-miR treatments (fusion index for negative control was 61.9% ± 3.5%, anti-miR-133a was 70.4% ± 3.5%, anti-miR-696 was 64.7% ± 5.6%, and joint transfection was 61.8% ± 5.6%, n = 3).
After 2 weeks of culture in differentiation media, myobundles prepared with myoblasts transfected with anti-miR-133a and -696 exhibited significantly higher twitch and tetanus forces than negative control (p < 0.0001, n = 4), anti-miR-133a (p = 0.0004, n = 4), and anti-miR-696 (p = 0.0002, n = 4; Fig. 5A). The contractile force induced by the negative control was similar to the values for bundles prepared with untreated myoblasts. Myobundles derived from myoblasts transfected with both anti-miR-133a and anti-miR-696 exhibited increased twitch and tetanic forces compared to other conditions (Fig. 5A). The tetanus-to-twitch ratio (TtR) was between 2.0 and 2.5, and there was no significant difference in TtR among the conditions.
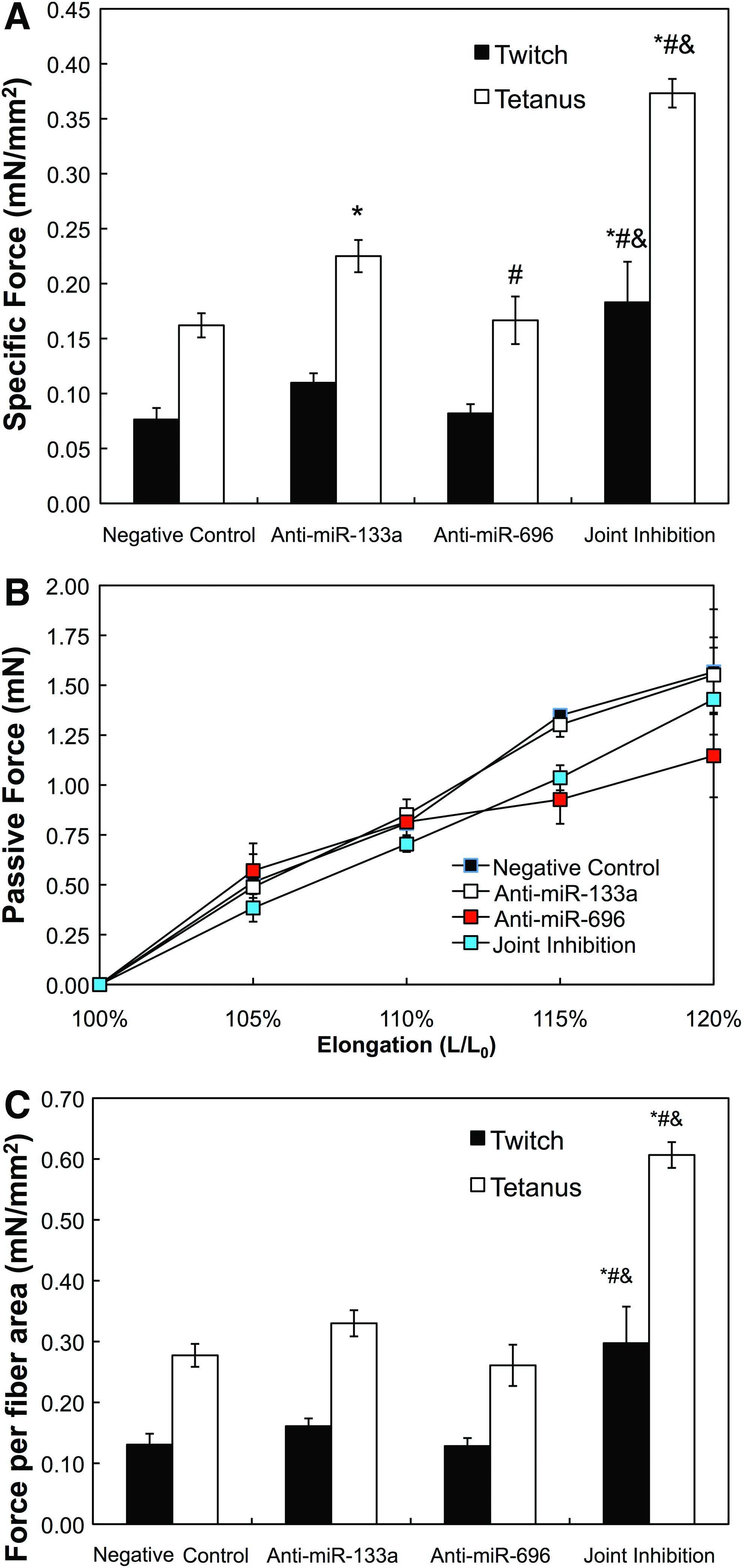
Isometric force testing of muscle bundles.
Since the increased passive force generation is correlated with increased extracellular matrix (ECM) presence in both native and 3D-engineered tissues,16,26–29 we measured passive force to assess if myobundle compaction observed with anti-miR treatment may be due to fibroblast contraction or ECM deposition. Passive forces increased in an approximately linear manner with increases in myobundle length (Fig. 5B). ANCOVA did not show any significant difference between rates of passive force amplitude change with increasing construct elongation, indicating that there is likely no significant difference in ECM content between microRNA treatments.
To assess whether fibroblasts in the myobundles were affected by the anti-miRs and might influence contraction or compaction of the myobundles, we prepared 3D bundles by seeding fibroblasts treated with anti-miRs into hydrogels prepared in the same manner as myobundles. After 14 days in culture, fibroblast bundle diameter was not affected by anti-miR treatment: diameters were 0.60 ± 0.02 mm, 0.60 ± 0.03 mm 0.58 ± 0.05 mm, and 0.62 ± 0.01 mm for bundles prepared with HSkM transfected with negative control, anti-miR-133a, anti-miR-696, or both anti-miRs, respectively (n = 4 for each condition).
There was no significant difference between area fraction of myofibers occupying the myobundles for conditions tested; the percent myofiber area was 58.4% ± 8.3%, 68.2% ± 7.4%, 63.9% ± 7.0%, and 61.5% ± 8.2% for bundles prepared with HSkM transfected with negative control, anti-miR-133a, anti-miR-696, or both anti-miRs, respectively (n = 4 for each condition). Using the calculated area fraction of fibers and specific force results per bundle presented above, we calculated the force produced by fiber area for each microRNA treatment condition. This analysis allowed us to account for potential variations in specific force due to anti-miR effects on ECM fraction or myofiber density among microRNA treatments. The tetanus force per myofiber cross-sectional area did not significantly differ among negative control, anti-miR-133a, and anti-miR-696 treatments of HSkM (Fig. 5C), partly due to the variation in the fractional myofiber area. However, the tetanus force per myofiber cross-sectional area was increased more than twofold by joint inhibition of miR-133a and miR-696. Twitch and tetanus force amplitudes per fiber area increased over twice in magnitude between joint inhibition and all other conditions (p < 0.05, n = 4 for all). From this analysis, the significant increase in tetanus forces for bundles made with HSkM transfected with anti-miR 133a may be partially attributed to the increased myofiber density within the engineered constructs.
We also assessed kinetic parameters of twitch and tetanus, including the time to peak twitch (TPT) and time to tetanus (TT) kinetics, and found no difference among the studied groups (TPT ranged from 160 to 190 msec for all conditions, and TT ranged from 1100 to 1125 msec). However, half-relaxation time of tetanus was significantly shorter in anti-miR-696 compared to all other groups (p < 0.01; Supplementary Fig. S3).
Effect of microRNA-133a and/or -696 inhibitors on MHC gene and protein expression of 3D human skeletal muscle bundles
Treatment of HSkM with anti-miR-696 increased gene expression of both slow and fast MHC compared to negative control (Fig. 6A; p < 0.05, n = 3). Interestingly, inhibition of both miRs resulted in a small, but statistically significant increase in fast MHC mRNA compared to negative control (p < 0.05, n = 3) and a significant decrease in slow MHC compared to anti-miR-696 treatment alone (p < 0.05, n = 3). The ratio of slow:fast MHC gene expression in the joint inhibition group was significantly lower compared to myobundles prepared by treatment of HSkM with either anti-miR-133a or anti-miR-696 (p < 0.05, n = 3, Fig. 6B). While immunostaining of myobundles did not show a difference in the percent of fiber stained with slow or fast MHC (92–97% slow and 90–100% fast; Fig. 6C), MHC staining intensity was generally consistent with RT-PCR findings, although with much lower levels of an increase in slow MHC protein than mRNA. Specifically, slow MHC intensity was higher for anti-miR-133a and anti-miR-696 treatment groups compared to negative control and joint inhibition conditions (p < 0.05, n = 3, Fig. 6C). Fast MHC intensity was not increased with treatment of anti-miR 133a or anti-miR696, but when both anti-miRs were added together, the intensity was 40–65% higher compared to other groups (p < 0.05 for all, n = 3, Fig. 6D).
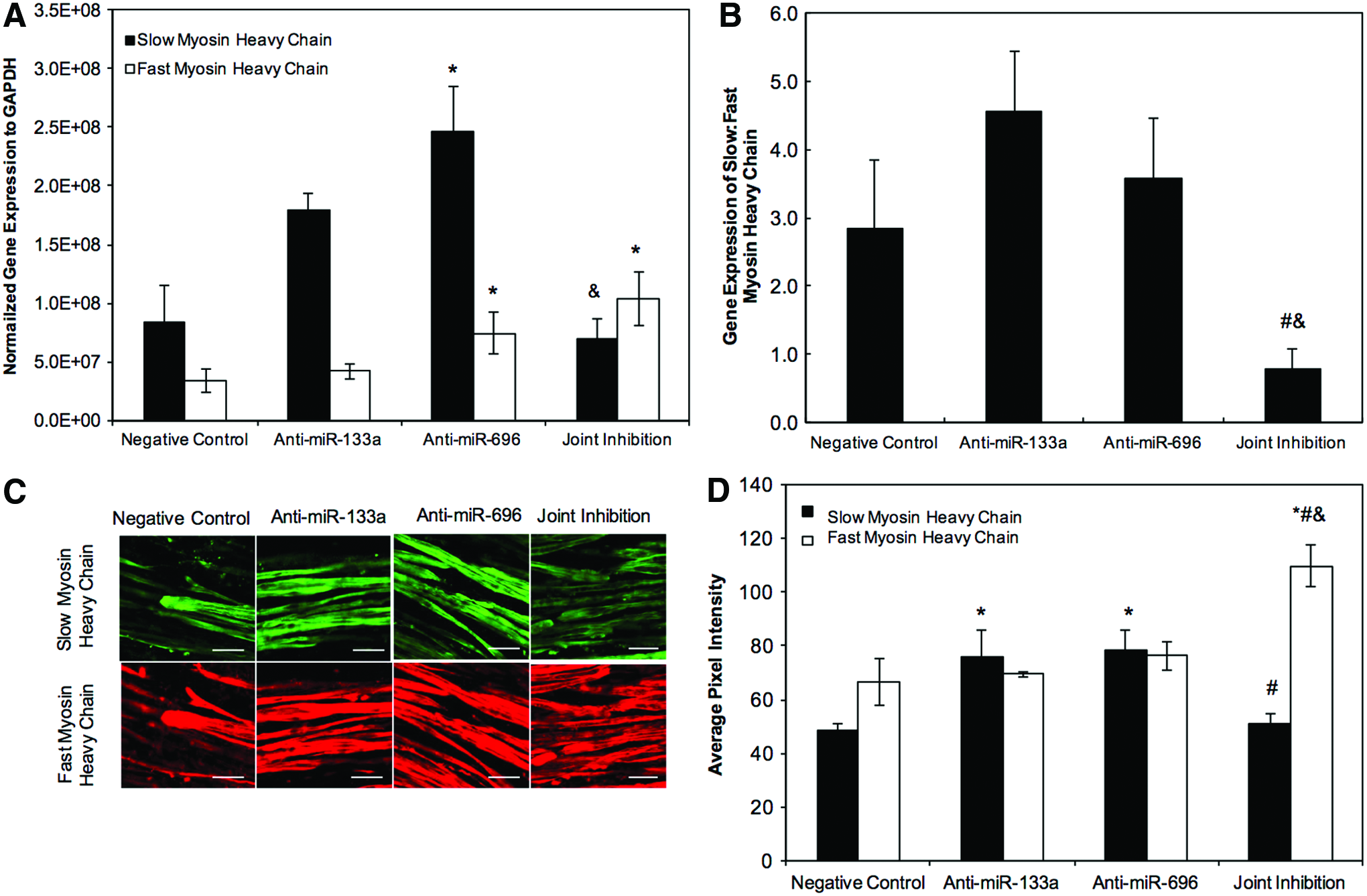
Myosin heavy chain (MHC) isoforms in 3D human myobundles seeded at 15 × 106 HSkM/mL and examined 14 days after addition of DM.
Western blots (Supplementary Fig. S4) showed modest increases in all myosin isoforms (16%) and slow MHC (34%) relative to negative control for cells treated with anti-miR 133a or anti-miR696. The changes in response to each anti-MiR were smaller than the 58% increase observed by immunofluorescence. Both the Western blots and immunofluorescence showed no change in slow MHC when both anti-miRs were added together. Western blots did not show a change in fast MHC for any anti-miR treatment. Together, the results suggest that anti-mIR-133a or anti-miR-696 treatment lead to modest increases in slow MHC, but no consistent increase in fast MHC.
Similar to results in 2D, only myobundles prepared with myoblasts treated with anti-miR-133a and anti-miR-696 produced an increase in PGC-1α mRNA (Fig. 7A). Since PGC-1α regulates mitochondrial gene expression, we examined the effect of the anti-miRs on two PGC-1α targets, HIF-2α 30 and COXII. 31 The mRNA levels of HIF-2α mRNA showed an expression pattern similar to that of PGC-1α mRNA, (Fig. 7B), but COXII mRNA levels were not altered by the anti-miRs. Thus, the anti-miRs are able to regulate the level of PPARGC1a such that some of the PGC-1α target genes are elevated.

PGC-1α
Discussion
In this study, we showed that inhibition of miR-133a and/or miR-696 promotes myogenic differentiation, changes in protein/gene expression that may alter metabolism, and increased force production in 2D- and/or 3D-engineered human muscle cultures. Specifically, inhibition of either miR-133a or miR-696 increased PGC-1α protein 14 days after onset of 2D differentiation, while inhibition of these two microRNAs simultaneously in 2D-cultured myotubes also increased PGC-1α gene (PPARGC1A) expression and SAA protein expression. In 3D-engineered human skeletal muscle myobundles, joint inhibition of miR-133a and miR-696 led to an increase in specific contractile force and fast MHC expression, as well as PPARGC1A and HIF-2α gene expression.
We used anti-miRs to modulate human skeletal muscle function due to their potent effects on myogenesis, 32 endogenous presence, and localization to the cytoplasm. 11 MicroRNAs were effectively delivered to the cytoplasm using liposomal transfection. Transient transfection leads to lower cell cytotoxicity 33 and prevents prolonged overexpression or inhibition of genes that may have deleterious effects on function. 34 The half-lives of single and joint microRNA inhibition were over 4 days, which was sufficient for anti-miRs to exert their effects on the myofiber development. As we observed with C2C12 cells that were treated with anti-miR-133a, 10 the anti-miR effects can persist for 2 weeks after transfection.
While our cultures are predominantly composed of HSkMs, fibroblasts are present and could contribute to the passive force and differences in myobundle compaction observed after anti-miR treatment. Fibroblasts help stabilize engineered muscle tissue and produce important ECM. 35 While Duisters et al. reported that levels of miR-133 were 15-fold greater in rat cardiac myocytes compared to rat fibroblasts, 36 the passive force in skeletal muscle myobundles was not sensitive to the anti-miR treatment (Fig. 5B) and transfection of fibroblasts with the anti-miRs did not affect compaction of bundles prepared only with fibroblasts. While we cannot exclude possible interactions between the fibroblasts and myofibers in the presence of the anti-miRs, these results suggest that differences in myobundle diameter and contractile force were due primarily to the myofibers.
Culture conditions in 2D and 3D differ quite drastically: the generation of unidirectional tension in 3D caused orientation and alignment of myofibers and promoted AChR clustering, while 2D cultures usually lack necessary stimuli to elicit this level of organization yielding random orientation of myofibers. Interactions between skeletal myoblasts and their surrounding ECM proteins contribute to skeletal myogenesis and function and may facilitate greater maturation in 3D versus 2D. 37 Nonetheless, our prior study 11 and this work with muscle cells cultured in 2D do show consistent enhancement of differentiation, which we also observed in 3D.
MicroRNAs inhibit mRNA translation either by blocking the initiation of translation, which does not affect mRNA levels, or by accelerating mRNA degradation, which leads to reduced mRNA levels. 38 Aoi et al. found that levels of PPARGC1A were unchanged for miR-696 overexpression and inhibition. 12 Likewise, in 2D and 3D culture, PPARGC1A expression was unchanged following miR-133a and miR-696 inhibition. Results are consistent with miR-133a and miR-696 inhibiting the initiation of PGC-1α translation. In contrast, joint inhibition of miR-133a and miR-696 significantly increased PPARGC1A gene expression. One possible explanation is that together miR-133a and miR-696 promote PPARGC1A degradation, so that when removed, PPARGC1A levels increase. However, this increased level of gene expression by joint inhibition of miR 133a and 696 did not produce more PGC-1α than inhibition of either microRNA alone, possibly because anti-miR 696 blocked translation or PGC-1α was under regulation of other targets of these microRNAs.
MiR-133a acts at an early stage in the differentiation pathway, whereas miR-696 acts at a later stage. 39 Inhibition of miR-133a represses serum response factor, while miR-696 regulates the metabolism coactivator, PGC-1α. However, unlike transcription factors, microRNAs tend to act through modest changes in the expression of one or more regulatory proteins. 40 Consistent with this view, we did not observe an increase in fast or slow myofibers or mitochondrial genes after individual microRNA treatment; joint inhibition led to significantly higher levels of SAA protein compared to negative control levels after 14 days in 2D culture suggesting increased levels of organization and muscle maturity within engineered tissues. 41
In our 3D studies, joint inhibition of miR-133a and miR-696 yielded higher twitch and tetanus forces compared to all other tested conditions. We found that responses to miR-133a inhibition are qualitatively consistent with those reported by Liu et al. in an in vivo mouse model where miR-133a inhibition led to an increase in type I fibers and slow MHC expression. 34 In addition, miR-696 inhibition caused an increase in MHC I fiber protein expression as observed by others.13,31
While fiber type has some influence on the speed and force of contraction, 41 skeletal muscle contraction depends, in a complex manner, upon the contractile proteins, calcium handling, and mitochondrial function. Furthermore, we observed only partial maturation of the bundles. Fiber types differ in MHC and troponin isoform expression, calcium handling proteins, and metabolic proteins. 41 Culture conditions (e.g., serum, glucose, and the presence of specific antibiotics) affect each of these functions in different ways.42,43 In this study, anti-miR treatments did not affect the kinetics of force relaxation, suggesting that calcium handling was not altered. Both miR-133a and miR-696 have a defined set of targets, so other miRs (or anti-miRs) and culture conditions may be needed for further differentiation to a mature muscle phenotype.
In summary, these results indicate that addition of multiple microRNAs can enhance expression of specific mature forms of muscle proteins and enhance contractile force. Combining microRNAs with electrical and/or mechanical stimulation may further enhance differentiation in vitro toward a mature muscle phenotype for function in vitro assessment of muscle. With further optimization, transient transfection of engineered muscle or local delivery of microRNAs or anti-miRs could be used to accelerate the repair process.
Footnotes
Acknowledgments
The authors would like to thank the following at Duke University: Dorothy Slentz for help in development of the primary myoblast isolation protocol; Mark Juhas, Lauran Madden, and Chris Jackman for guidance and help with the functional force testing of muscle constructs; Samanatha Lasater for assistance with RT-PCR; and Cristina Fernandez for post hoc testing of mitochondrial genes. The work was supported by NIH Grants R01AR055226 and R01AR065873 from the National Institute of Arthritis and Musculoskeletal and Skin Disease to N.B. and NIH Grant UH2/UH3TR000505 and the NIH Common Fund for the Microphysiological Systems Initiative to G.A.T.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.