
Abstract
Although regeneration of human cartilage is inherently inefficient, age is an important risk factor for osteoarthritis. Recent reports have provided compelling evidence that juvenile chondrocytes (from donors below 13 years of age) are more efficient at generating articular cartilage as compared to adult chondrocytes. However, the molecular basis for such a superior regenerative capability is not understood. To identify the cell-intrinsic differences between juvenile and adult cartilage, we have systematically profiled global gene expression changes between a small cohort of human neonatal/juvenile and adult chondrocytes. No such study is available for human chondrocytes although young and old bovine and equine cartilage have been recently profiled. Our studies have identified and validated new factors enriched in juvenile chondrocytes as compared to adult chondrocytes including secreted extracellular matrix factors chordin-like 1 (CHRDL1) and microfibrillar-associated protein 4 (MFAP4). Network analyses identified cartilage development pathways, epithelial–mesenchymal transition, and innate immunity pathways to be overrepresented in juvenile-enriched genes. Finally, CHRDL1 was observed to aid the proliferation and survival of bone marrow-derived human mesenchymal stem cells (hMSC) while maintaining their stem cell potential. These studies, therefore, provide a mechanism for how young cartilage factors can potentially enhance stem cell function in cartilage repair.
Introduction
D
Clinically, it has been long known that cartilage repair can be effective in young patients, while in adults, the tissue formed after injury or trauma is inferior to normal functioning cartilage. A concise study in rabbits clearly showed superior repair in young rabbits as compared to adult. 4 Similarly, an in utero cartilage defect study in lamb showed efficient repair. 5 More recent reports have demonstrated that human juvenile cartilage cells (from donors below 13 years of age) are more efficient at generating functional cartilage as compared to the adult cells.6,7 Juvenile chondrocytes have also been demonstrated to modify adult chondrocytes in coculture experiments8,9 suggesting that paracrine factors secreted from the young chondrocytes may have a therapeutic benefit. Upon injection into the intervertebral disc of rats, juvenile chondrocytes have been seen to exhibit a more active remodeling process than controls; 10 however, the use of human juvenile chondrocytes for regenerative purposes is likely to be limited by donor supply. Coculture studies, using a combination of juvenile chondrocytes with adipose-derived stem cells (ADSCs) to overcome this hurdle, have yielded some promising results, suggesting that small numbers of juvenile chondrocytes in combination with ADSCs can be used to induce robust cartilage formation. 11
Although multiple studies have documented the superior potential of juvenile chondrocytes, the molecular basis of such superior regeneration has not been explored. Recent studies in bovine and equine young and adult cartilage have begun to address this question; however, a precise study of the molecular factors and biological pathways that underpin these differences in humans has not yet been documented. Part of the challenge for the lack of such studies is the difficulty in obtaining healthy human cartilage especially from young donors. To address this, we have examined the gene expression profiles of a small cohort of human juvenile and adult chondrocytes, and of human mesenchymal stem cells (hMSCs) utilizing exon microarrays. Analyses of the global gene expression differences has allowed us to identify factors that distinguish juvenile chondrocytes from adult chondrocytes, including paracrine factors secreted exclusively by the juvenile chondrocytes as compared to the adult chondrocytes. In addition, we explore the potential roles that the identified factors may play in the enhanced ability of juvenile chondrocytes for cartilage repair and regeneration.
Materials and Methods
Chondrocyte isolation and culture
Adult (34 years) and juvenile articular chondrocytes (24 weeks fetus, 6 months, 18 months, and 6 years) were purchased from Lonza (Clonetics™; Lonza Walkersville, Inc.) and cultured in Chondrocyte Growth Medium (Clonetics CGM™, Lonza Walkersville, Inc.). Articular cartilage was harvested from grossly normal cartilage pieces discarded during notchplasty or debridement from patients undergoing anterior cruciate ligament reconstruction with no history of OA symptoms (18, 27, and 34 years of age) and from OA patients undergoing total joint replacement (64 and 79 years of age) under the approved Human Subjects Institutional Review Board protocols. Cartilage was dissected and the chondrocytes dissociated and cultured in high-density monolayers for limited passages, as described previously. 12
Human mesenchymal stem cells were purchased from Lonza (Lonza Walkersville, Inc., three samples from adult donors of 19, 27, and 30 years of age) and cultured in Mesenchymal Stem Cell Growth Medium (MSCGM™; Lonza Walkersville, Inc.).
Microarrays and gene expression analyses
Total RNA was purified using the Qiagen RNeasy Plus Mini Kit (Qiagen). For quantitative polymerase chain reaction (qPCR), cDNA was synthesized using the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems) and the USB VeriQuest SYBR Green qPCR Master Mix (Affymetrix) was used with gene-specific primers. All analyses were performed using the ddCT method and expression was normalized to GAPDH or 18S. SYBR green primer sequences were as follows: Human GAPDH F—TGTCCCCACTGCCAACGTGTC, R—AGCGTCAAAGGTGGAGGAGTGGGT; CHRDL1 F–TGCGAGTACAATGGGACAAC, R—GTTGCCGATTCTGAAAGAGC; OMD F—GTGTCAGTGAATGCTTCTGTCC, R—GAGTTGCTGAATGTGCATCG; MFAP4 F—TGAGAACAACACGGCCTATG, R—TCAAAGCCTGCCACAAAGAG; SHOX F—CAACGTGGAAAAGGCGTTAC, R—TCCCCTTCAGAAACACAACC.
For microarray expression analyses, purified RNA was run on a Human Gene 1.0 ST Array (Affymetrix). Two independent replicate samples for hMSCs and adult chondrocytes and three independent replicate samples for juvenile chondrocytes were run on Human Gene 1.0 ST Arrays (Affymetrix). Data analysis was performed using dChip 13 as described by the manual, and network analysis of differentially expressed genes was performed using MetaCore (Thomson Reuters).
Immunohistochemistry
Juvenile and adult chondrocytes were fixed in 4% paraformaldehyde (Sigma) then permeabilized in cold methanol (Sigma), before blocking in phosphate-buffered saline (PBS) containing 1% bovine serum albumin, 10% fetal bovine serum, and 0.4% Triton X-100. Cells were incubated with primary antibody overnight (anti-CHRDL1 1:100 [Bioss USA] and anti-MFAP4 1:200 [Santa Cruz]). The following day cells were washed in PBS and incubated for 1 h in secondary antibody (Alexa 594 goat anti-rabbit 1:250 [Invitrogen]) and cellular DNA was counterstained with DAPI (Life Technologies).
FACS analyses
Cells were dissociated to a single-cell suspension using TrypLE and fixed in BD Cytofix buffer at 1 × 107 cells per 1 mL for 20 min at room temperature. Cells were stained by incubating with antibodies of mesenchymal stem cell markers (anti-human CD90-FITC, and anti-human CD105-Alexa647; BD Biosciences)14,15 for 30 min. Primary antibodies were diluted according to the manufacturer's instructions. Cells were scanned using a LSR II flow cytometer and analyzed with FlowJo software.
Treatment with juvenile factors
Chondrocytes and hMSCs were plated at 1000 cells per well in duplicates in 48-well plates and cultured for 24 h in complete medium. After 24 h, cells were treated with control or medium containing 100 nM of recombinant human CHRDL1 (R&D Systems) or 25 nM of recombinant human MFAP4 (Abcam), with media and recombinant protein replacement every day for 4 days.
Cell proliferation assay
Cell viability was assayed daily in the recombinant protein treatment assays with the PrestoBlue Cell Viability Reagent (Life Technologies) following the manufacturer's instructions. PrestoBlue reagent was added to the cell culture medium, and the cells were incubated at 37°C for 30 min. Fluorescence intensity of the PrestoBlue was measured at 690 nm (650 nm excitation wavelength) with a microplate reader (Molecular Devices). Fluorescence level was expressed in Relative Fluorescence Units (RFU).
Results
Juvenile chondrocytes have gene expression patterns distinct from both adult chondrocytes and MSC
Striking phenotypic and functional differences have been previously reported between juvenile and adult human chondrocytes.6,16 However, to date, no differential molecular regulators or pathways have been identified for the young and adult chondrocytes. The major difficulty for such analyses is the paucity of normal human cartilage samples. As a first attempt to identify juvenile chondrocyte-specific factors that provide them with the characteristics different from adult chondrocytes, we have utilized a small cohort of juvenile and adult chondrocytes consisting of juvenile chondrocytes isolated from three different donors and adult chondrocytes isolated from two different donors (see Materials and Methods section for details). In addition, these juvenile and adult chondrocyte samples were previously characterized by us 16 and demonstrated the characteristic functional differences such as increased extracellular matrix (ECM) production by the juvenile chondrocytes compared to adult chondrocytes. To investigate the molecular differences between the juvenile and adult chondrocytes, we utilized exon microarrays to determine their global gene expression profiles. For the microarray analysis, total RNA was extracted from chondrocytes after limited cell culture and expansion (1–2 passages in 2–4 days) in high-density monolayers to prevent dedifferentiation, as described previously. 17 Human Gene 1.0 ST Arrays (Affymetrix) were utilized and data analysis was performed using dChip. 13
First, we analyzed the gene expression profiles to compare the juvenile chondrocyte samples from 18-month-old, 24-week-old fetus, and 6-month-old. As shown in Supplementary Fig. S1 (Supplementary Data are available online at www.liebertpub.com/tea), a comparison of chondrocytes from 18-month-old with 24-week-old shows a 94% overlap. Similarly, chondrocytes from 18-month-old/6-month-old and 24-week-old/6-month-old show a 93.5% overlap out of 22,022 genes analyzed, demonstrating that there is overall a very high similarity between these samples. The complete gene lists from these comparisons are included as Supplementary Table S1. To identify the differentially expressed genes in the juvenile chondrocytes, we then compared them to the adult chondrocytes (schematic Fig. 1A). Genes were ranked according to the average fold change in transcript expression in all the juvenile chondrocyte samples averaged together compared to the transcript expression in the averaged adult chondrocyte samples. Analysis of genes with a 1.5-fold or greater increase in expression identified 597 number of genes that were upregulated in the juvenile chondrocytes compared to the adult chondrocytes (Supplementary Table S2). The top 20 genes are listed in Figure 1B. Network analyses of all the genes with a 1.5-fold or greater increase in expression in juvenile versus adult chondrocytes interestingly identified pathways already implicated in early chondrocyte development, including epithelial–mesenchymal transition and cartilage development pathways (Fig. 1C). We also performed network analysis on the genes with a 1.5-fold or greater decrease in the juvenile chondrocytes when compared to the adult chondrocytes which, as expected, revealed pathways known to be upregulated in mature chondrocytes (Supplementary Fig. S2).
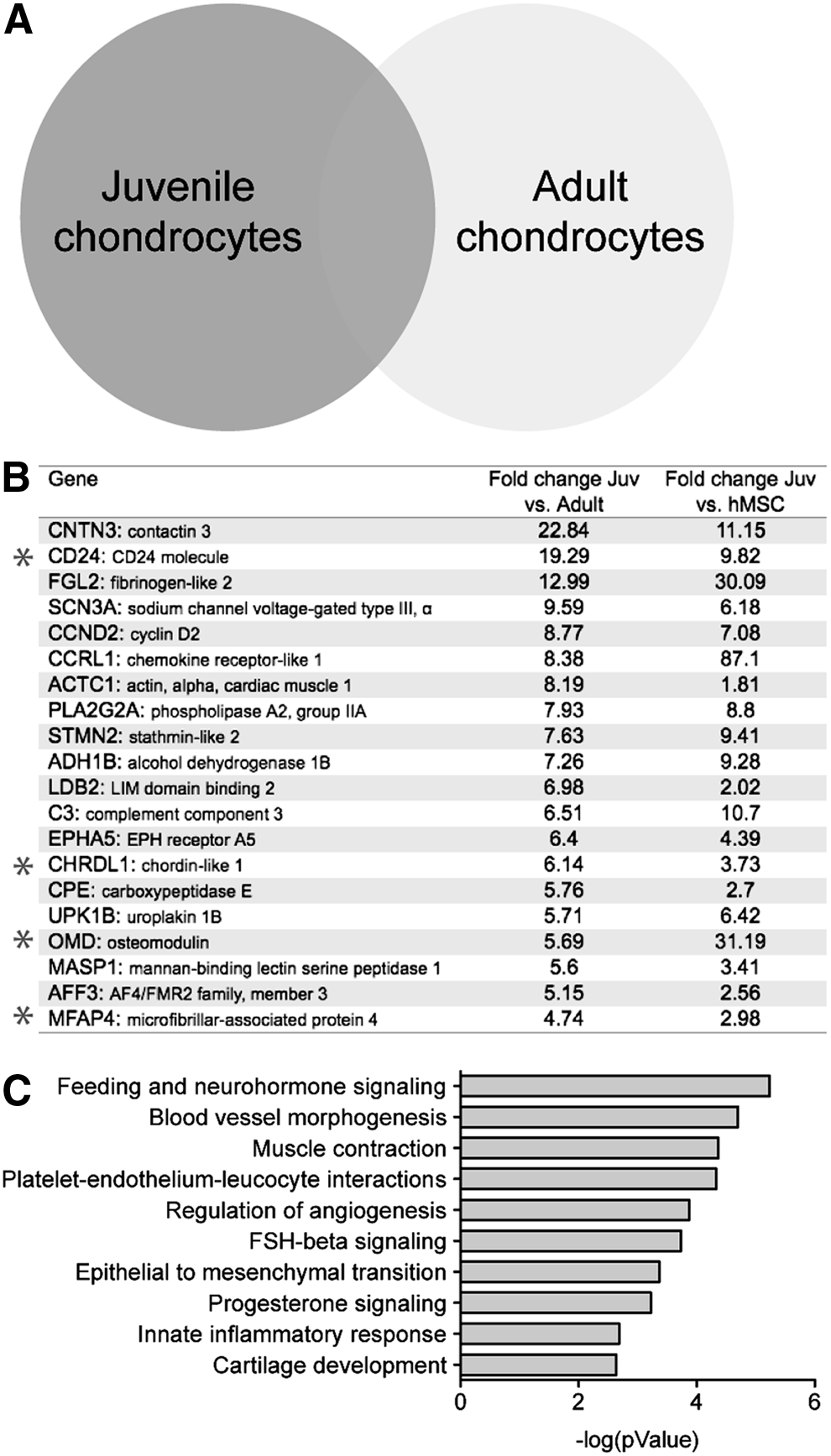
Juvenile chondrocytes display unique gene expression profiles.
Of the genes identified as having a 1.5-fold or greater increase in expression in the juvenile chondrocytes when compared to the adult, we focused on the genes with the most differential expression (the top 20 genes, ranked by fold change; Fig. 1B). This list of unique juvenile chondrocyte genes included several genes that had previously been implicated in chondrocyte or ECM function. Initially, we chose to further investigate paracrine factors that were differentially secreted by the juvenile chondrocytes. These factors included chordin-like 1 (CHRDL1) a known bone morphogenetic protein (BMP)-antagonist, 18 osteomodulin (OMD) a proteoglycan that is highly expressed in bone and cartilage progenitor cells, 19 and microfibrillar-associated protein 4 (MFAP4) 20 an ECM protein known to bind collagen.
Next, we utilized bone marrow-derived human mesenchymal stem cells (hMSC) from two different donors, to investigate whether the juvenile chondrocytes share any stem cell-like factors with the hMSC. We analyzed genes that had a 1.5-fold or greater increase in expression in the juvenile chondrocytes when compared to the hMSCs (Supplementary Table S3). Network analysis of the genes with a 1.5-fold upregulation in the juvenile chondrocytes revealed several pathways involved in cell cycling and repair of DNA damage (Supplementary Fig. S3A). Network analysis of the genes with a 1.5-fold or greater decrease in the juvenile chondrocytes when compared to the hMSCs identified pathways related to other lineages, indicating the commitment of the juvenile chondrocytes to the chondrogenic lineage as opposed to the multipotent hMSCs (Supplementary Fig. S3B).
Next, we analyzed the subset of genes that are upregulated in both juvenile chondrocytes and hMSCs compared to adult chondrocytes and thus may represent genes indicative of stem cell-like qualities or stemness, in juvenile chondrocytes (Fig. 2A). We identified 249 genes that are upregulated by 1.5-fold or greater in both juvenile chondrocytes and hMSCs when compared to adult chondrocytes (shown in schematic in Fig. 2A and the top 20 are listed in Fig. 2B, Supplementary Fig. S4 and Supplementary Table S4). To identify the pathways these genes represented, we performed network analyses that identified multiple developmental and reproductive pathways (Fig. 2C), suggesting that there is possibly a stem cell-like population or characteristics present in juvenile chondrocytes that may be responsible for their high regeneration potential. The interesting common genes included HoxB2, a family member of homeobox genes known to play regulatory roles in vertebrate patterning 21 and ADAMTS12, a metalloproteinase implicated in growth plate development as well as cartilage degeneration in arthritis. 22 Upon using these common 249 genes as the input list for the MetaCore gene network analysis program, we identified the Wnt pathway as the top enriched biological pathway [including related genes Tcf (Lef), Frizzled, insulin-like growth factor-1 receptor, c-Jun, and SFRP4] (Supplementary Fig. S4). Besides the Wnt pathway, which is well known to play a role in multiple tissue-specific stem cells, the other top scored networks included G-protein and Neuroglin-related networks. Published references relevant to these top three stemness genes' networks are listed in the Supplementary Table S4.
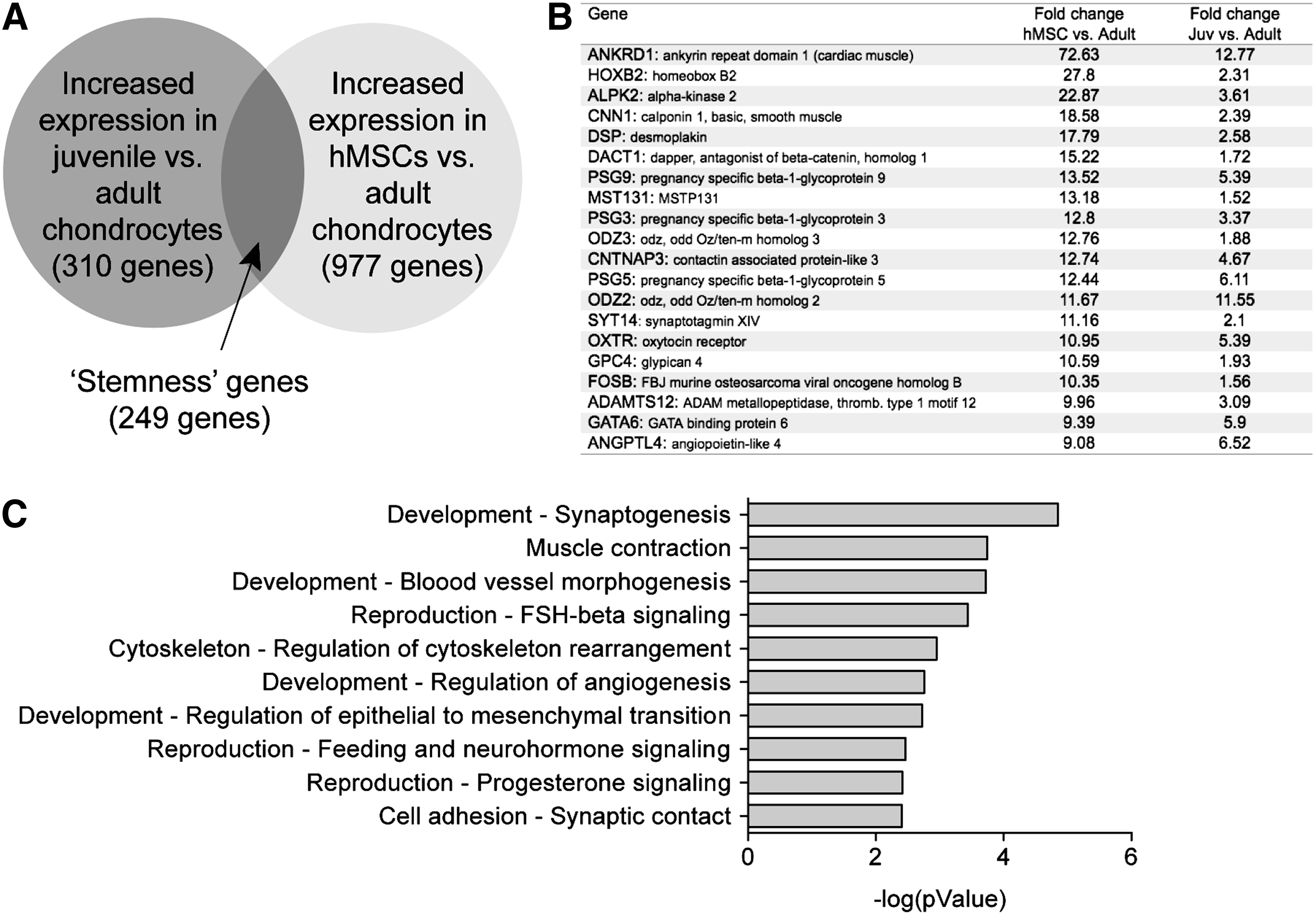
Common stemness genes upregulated in both juvenile chondrocytes and hMSCs when compared to adult chondrocytes.
Validation of juvenile chondrocyte-specific factors
To validate the findings from the microarray study and to explore the factors of interest further, we examined the mRNA levels of the putative juvenile factors by qPCR. We used a range of samples, including additional juvenile and adult chondrocyte, besides the ones used for the initial microarray experiments for these validation studies. We also included samples from OA patients to test whether the identified juvenile factors are altered in the diseased state since some embryonic gene transcripts (Col2α1, e.g.) are known to get upregulated in OA chondrocytes. The qPCR data were in good agreement with the microarray data and identified trends in the expression patterns of these factors (Fig. 3). Compared to the expression in hMSCs, CHRDL1 was upregulated fivefold or more in all juvenile samples examined, whereas the adult and OA samples showed no change in expression (Fig. 3A). OMD gene expression was somewhat more variable; elevated levels were observed in the juvenile chondrocytes compared to hMSC, but some adult and OA samples also showed moderate upregulation (Fig. 3B). MFAP4 was upregulated ∼10-fold in all the juvenile chondrocytes, and showed progressive decreases in expression levels with age in adult chondrocytes (Fig. 3C).
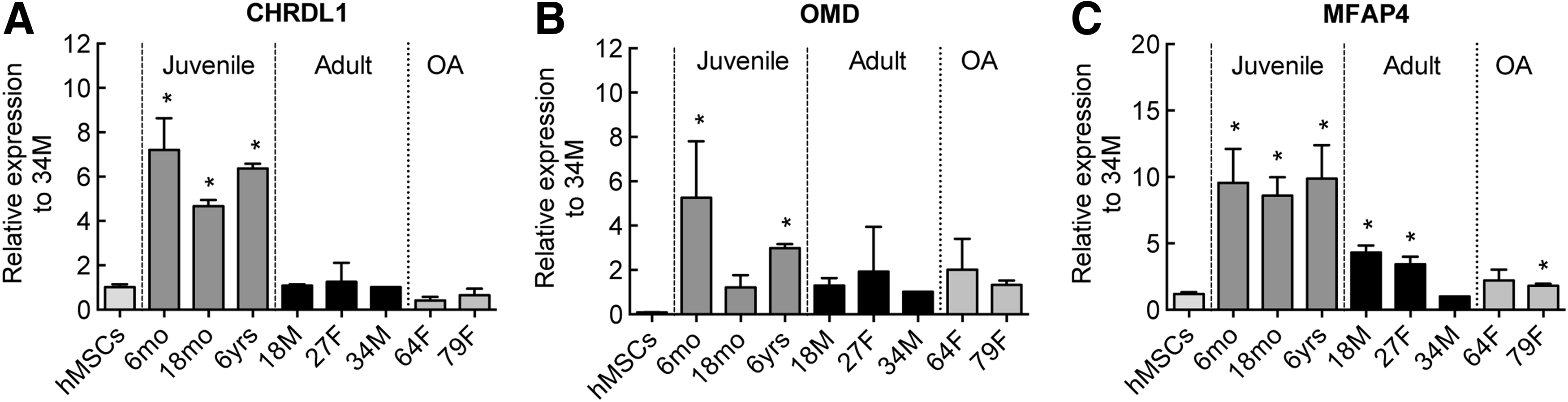
qPCR assessment of gene expression levels of juvenile chondrocyte markers selected for further analysis.
MFAP4 shows an age-dependent decrease while CHRDL1 is absent in adult chondrocytes
To assess whether the juvenile chondrocyte-specific factors we identified through our mRNA studies were upregulated at the protein level, we performed immunostaining on juvenile and adult chondrocytes. Since the expression levels of OMD were somewhat variable, we omitted this factor from further investigations. We observed that CHRDL1 was clearly expressed at the protein level in juvenile chondrocytes, whereas expression was completely absent in the adult chondrocytes (Fig. 4A). On the other hand, MFAP4 displayed more intense staining in the juvenile chondrocytes compared to the adult, but a decline in expression was observed in the adult samples (Fig. 4B), in good correlation with the mRNA expression data (Fig. 3).
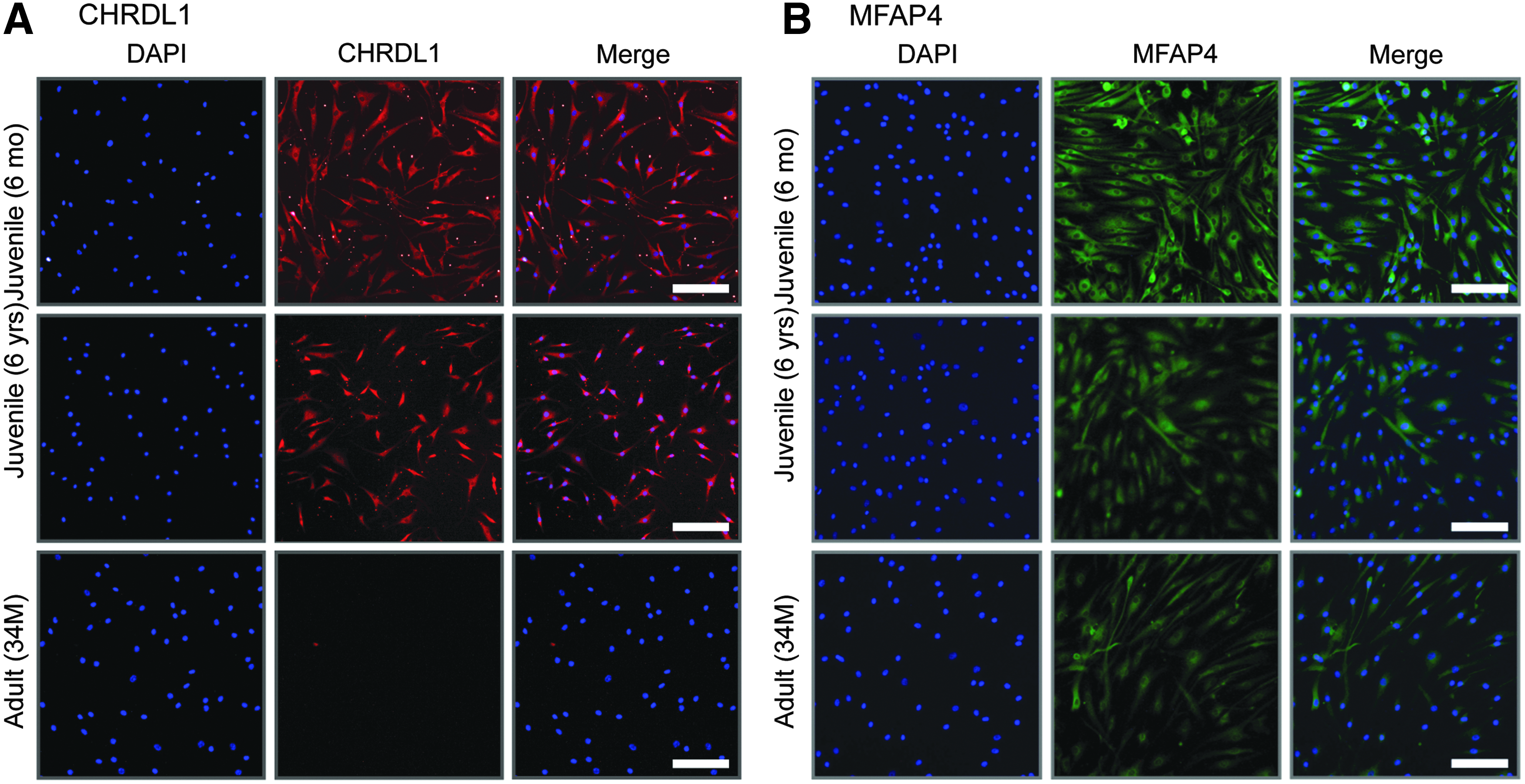
Protein expression of selected juvenile chondrocyte-specific markers.
CHRDL1 stimulates an increase in hMSC proliferation, but does not affect proliferation of adult chondrocytes
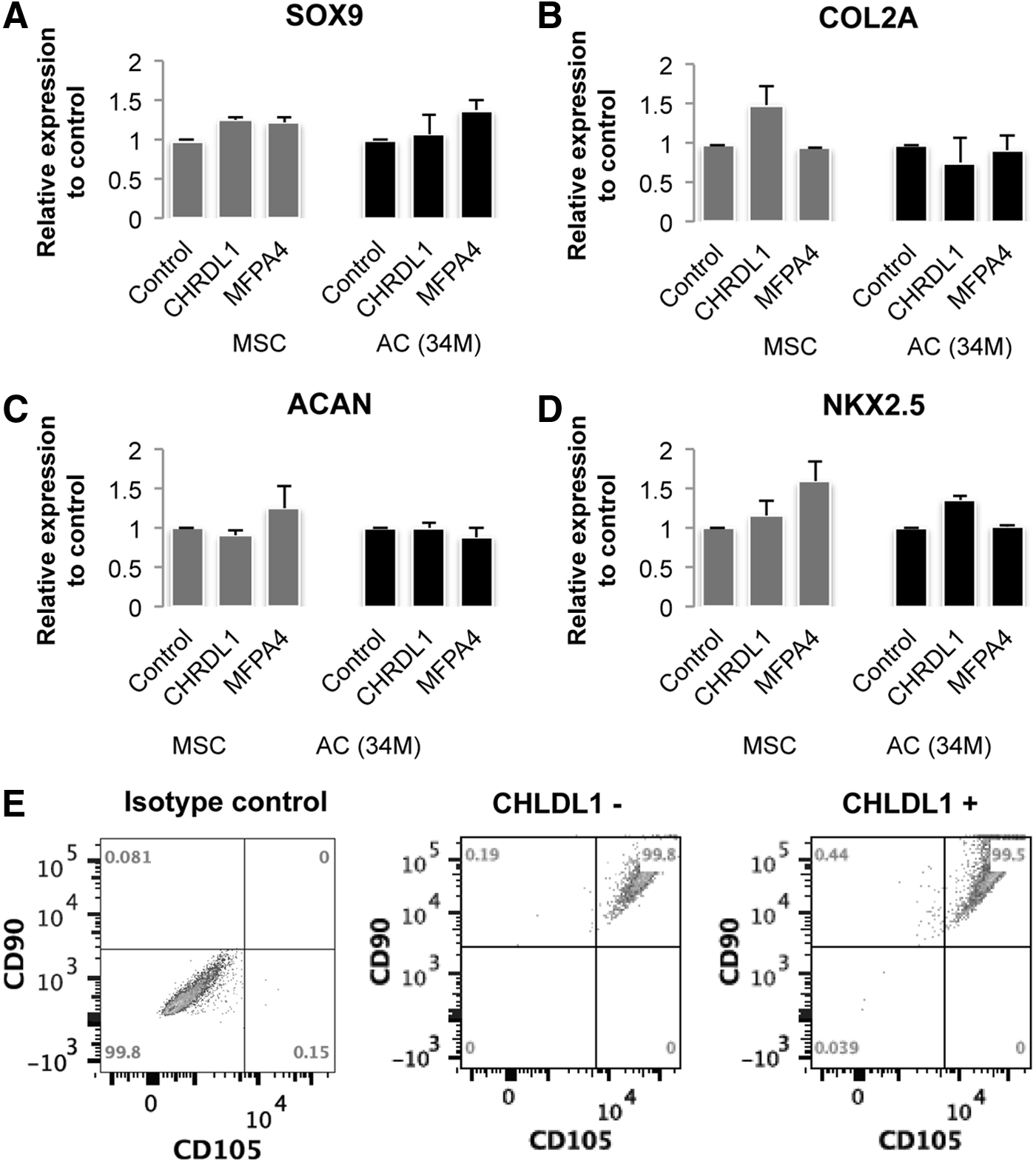
Next, we explored a possible mode of action of CHRDL1 and MFAP4 in young cartilage, specifically testing if either of these factors promoted stem cell survival or proliferation. We exposed hMSCs and adult chondrocytes to either soluble recombinant human CHRDL1 or MFAP4 and observed the effect on the proliferation of hMSCs and adult chondrocytes. When cultured for 4 days in the presence of CHRDL1, hMSCs showed a 3- to 4-fold increase in cell proliferation, whereas no effect was seen on adult chondrocytes (Fig. 5A). Neither the hMSCs nor the adult chondrocytes showed an increase in proliferation in response to MFAP4 treatment (Fig. 5B). In addition, we further investigated the role of CHRDL1 and MFAP4 to assess whether these factors alter the chondrogenic differentiation potential of hMSCs. We have addressed this question in two different approaches. First, we examined whether a treatment with CHRDL1 or MFAP4 (that does not alter hMSC growth) affects the gene expression of NKx2.5, a stemness-related gene or chondrogenic genes, SOX9, COL2A1, or ACAN in hMSC (Fig. 6 A–D). The treatment with the juvenile factors was performed for 2 days or a week with the factors being replenished daily. No significant change was observed in either the stemness or chondrogenic markers. Second, we examined whether treatment and subsequent expansion with CHRDL1 affects the characteristic cell surface markers on hMSC that are associated with stemness, CD90, and CD105 (Fig. 6E). We observe that treatment with CHRDL1 expands the hMSC, but their stemness markers are retained with no increase in chondrogenic gene expression.
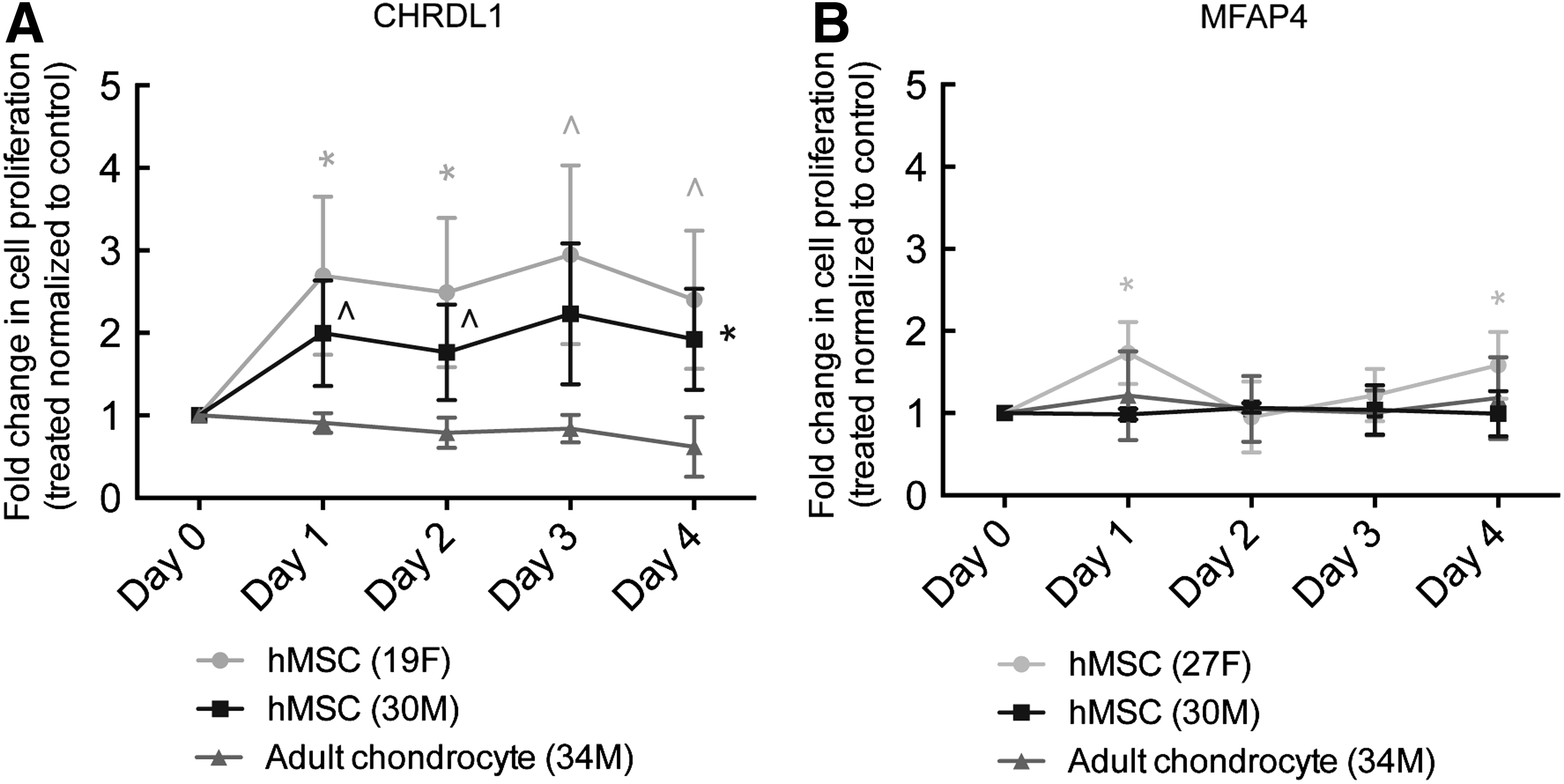
CHRDL1 stimulates proliferation of hMSCs, but MFAP4 does not.
Discussion
Although the higher repair potential of juvenile chondrocytes has been reported in the last few years,6,7 the molecular regulators responsible are unknown. Detailed characterization of juvenile and adult chondrocytes is required to dissect the underlying biological differences that lead to the observed increased regenerative capability of juvenile chondrocytes. Identification of the human juvenile chondrocyte-specific factors will not only advance the fundamental understanding of the age-associated loss of regeneration potential, but also provide new strategies for enhancing cartilage regeneration and for engineering superior cartilage tissue. Recently, two studies have described global gene expression profiling of juvenile and adult cartilage in bovine and equine tissues, respectively.23,24 The study in bovine articular cartilage identified genes related to cartilage growth and expansion (COL2A1, COL9A1, MMP2, MMP14, and TGFβ) to be upregulated in bovine juvenile chondrocytes, whereas structural genes (COMP, FN1, TIMP1, TIMP3, and BMP2) were upregulated in bovine adult cartilage. 24 In the equine study, 23 cartilage from young horses (4 years old equivalent to an ∼14-year-old human) and old horses (15 years or older equivalent to human older than 52 years) were utilized for genome-wise RNA sequencing (RNA-seq) to identify gene expression differences. Therefore, the comparison is between adult (since juvenile is generally defined as 13 years or younger) and old cartilage and hence demonstrates the aging-related changes more than the changes associated with the early postnatal development that are reflected in the juvenile and adult cartilage. Nevertheless, upon comparing genes upregulated in the adult versus old cartilage in the equine studies to the genes upregulated in juvenile versus adult cartilage in our human samples, a set of 54 common genes was identified (Supplementary Table S5). It is to be noted that only 54 genes were common out of 600 genes that upregulated 1.5-fold or more in juvenile versus adult cartilage, and such a small overlap likely reflects the fundamental age difference in comparison sets and possibly species differences as well. The study reported multiple ECM genes, including collagen types, biglycan, asporin and various Matrilins, to be decreased in old cartilage, while the expression of Col10a1 and Col25a1 was increased. 23 Another interesting difference was a reduction in multiple Wnt pathway factors with an increase in the Wnt antagonist DKK1 in the older cartilage. 23
Limitation of obtaining normal cartilage samples has generally precluded such studies in human tissues. We had previously established quantitative functional methods to distinguish juvenile and adult chondrocytes that were consistent with previously reported characteristics such as increased ECM production by the juvenile chondrocytes compared to adult chondrocytes. The small cohort of juvenile and adult chondrocyte samples utilized retained the expected functional differences. The average profile of the juvenile and adult chondrocytes clearly showed distinct gene expression patterns, although the general chondrogenic markers (e.g., SOX9, COL2A1, AGAN) were expressed in both populations as described earlier. 16
Many of the genes that were highly expressed in juvenile chondrocytes as compared to the adult chondrocytes were secreted ECM factors. Besides the known ECM factors like col2a1 and col6 that are known to be enriched in juvenile cartilage as compared to adult cartilage, we identified new factors like CHRDL1 and MFAP4 that were preferentially secreted by juvenile chondrocytes. CHRDL1 is interesting since chordin is a known BMP antagonist. 25 The family member chordin-like2 (CHRDL2) has been identified as being expressed exclusively in the cartilage of the developing joint and connective tissue in reproductive organs. 18 The extracellular protein MFAP4 was upregulated in juvenile cartilage at the protein level and there was a notable decrease with aging, unlike the protein expression of CHRDL1 that was expressed only in the juvenile chondrocytes with little or no expression observed in the adult chondrocytes. Our studies have therefore identified new molecular factors that are enriched in juvenile human chondrocytes and undergo an age-dependent decline.
Next, we tried to decipher a molecular basis for the enhanced regenerative potential of juvenile human chondrocytes. One possible reason can be an enrichment of stem cell-like factors in juvenile chondrocytes that allows increased proliferation, and hence more ECM production than the adult chondrocytes. Another possible explanation for a higher regenerative potential for juvenile cartilage could be the secreted paracrine factors that can support stem cell function, survival, or differentiation better than the adult cartilage. To test the first possibility, we tested whether some of the mesenchymal stem cell factors are enriched in the juvenile chondrocytes. We identified 249 genes that are commonly upregulated in juvenile chondrocytes and hMSCs in comparison to adult chondrocytes. Network analyses indicated that these genes were involved in reproductive and developmental pathways suggesting that juvenile chondrocytes potentially retain some mesenchymal stem/progenitor factors that contribute to their regenerative potential. These common genes included HoxB2, a family member of homeobox genes known to play regulatory roles in vertebrate patterning 21 and ADAMTS12, a metalloproteinase implicated in growth plate development as well as cartilage degeneration in arthritis. 22
To test the ability of the newly identified juvenile factors to support stem cell function, we tested their effect on the proliferation of hMSCs. While MFAP4 did not show any significant effect on the proliferation of hMSCs, CHRDL1 showed a remarkable stimulation of hMSC proliferation. These observations were confirmed in hMSC derived from two independent donors. Furthermore, we assessed whether the hMSC retained their stem cell characteristics upon expansion with the juvenile factors and both FACS and gene expression analyses validated that the stem cell markers were retained while there was no increase in the chondrogenic gene expression. The juvenile factors therefore expanded the bonafide mesenchymal stem cell population. Overall, these studies suggest that the juvenile cartilage has a dual advantage toward cartilage regeneration—the intrinsic increase in the growth and ECM generation capability of the juvenile chondrocytes as well as secretion of paracrine factors by the juvenile chondrocytes that stimulate stem cell function.
In summary, these studies provide a first global gene expression snapshot of juvenile chondrocytes in comparison to adult chondrocytes and identify new juvenile chondrocyte factors and their potential stimulatory effects on stem cell function. Future studies will explore the function and significance of the identified factors in detail and provide an increased molecular understanding of age-associated changes in intrinsic cartilage regeneration and repair. Additionally, the identified factors can be potentially useful for the stem cell-based tissue engineering strategies for cartilage regeneration.
Footnotes
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.