
Abstract
Low oxygen tension (LOT) has been reported to promote chondrogenic differentiation and prevent cellular senescence of stem cells. Therefore, the introduction of LOT conditions into conventional tissue engineering processes could further improve the potential of the constructs generated for cartilage repair. The purpose of this study was to elucidate the feasibility of LOT preparation on the chondrogenic differentiation of a scaffold-free tissue-engineered construct (TEC) derived from synovial mesenchymal stem cells (MSCs), construct whose feasibility for cartilage repair has been demonstrated in previous preclinical and clinical studies. Culture of MSCs under LOT conditions prevented cellular senescence and promoted the proliferative capacity of human synovial MSCs. In addition, TEC prepared from human synovial MSCs under LOT conditions (5% O2; LOT-TEC) showed superior in vitro chondrogenic differentiation capacity compared to that prepared under the usual 20% O2 (normal oxygen tension [NOT]; NOT-TEC), with elevated glycosaminoglycan production and elevated levels of chondrogenic marker gene expression. Notably, LOT-TEC differentiated into a hyaline-like cartilaginous tissue of approximately 1 cm in diameter without the detectable presence of fibrous tissue, while conventional NOT-TEC differentiated into a mixture of hyaline-like and fibrocartilaginous tissues. This is the first demonstration of in vitro development of a hyaline-like cartilaginous tissue of an implantable size to chondral lesion that was derived from human MSCs without the use of an exogenous scaffold. The manipulation of oxygen tension is a safe procedure with low cost and, thus, may be a clinically relevant option to improve the quality of TEC-mediated cartilage repair.
Introduction
I
Low oxygen tension (LOT) is one of the potential methods that can be used to promote chondrogenic differentiation,15–19 maintain stemness, 20 and prevent cellular senescence17,19 of MSCs. The effect of LOT on chondrogenesis has been assessed during in vitro chondrogenic differentiation assays using conventional pellet culture systems.17,18 However, little is known how LOT preparation can be efficiently applied to promote the in vivo repair quality of cartilage. Once MSCs are transplanted to the chondral defects in vivo, it is difficult to control the posttransplant cellular microenvironment, including the oxygen tension. Furthermore, oxygen tension in a conventional incubator is 20% and, thus, is much higher than that experienced by cartilage in the normal physiological condition. 21 Thus, conventional culture systems exert an unphysiologically high oxygen tension on the TEC, followed by the drastic reduction in oxygen tension at the time of in vivo implantation within the joint (Fig. 1). Such drastic changes of oxygen tension may also affect cellular functions and potencies. Therefore, we hypothesized that it is a reasonable approach to make the TEC chondrogenically differentiated ex vivo before implantation rather than directly implanting undifferentiated TEC to chondral lesions. Specifically, we wanted to maintain oxygen tension consistently within the physiological range (at 5%; LOT) throughout the culture process from the expansion of human synovial MSCs until the chondrogenic differentiation of the TEC. If LOT promotes ex vivo chondrogenesis of the TEC as we hypothesized, this method could then be directed to clinical applications because LOT cultures can be safely performed with relatively low cost.
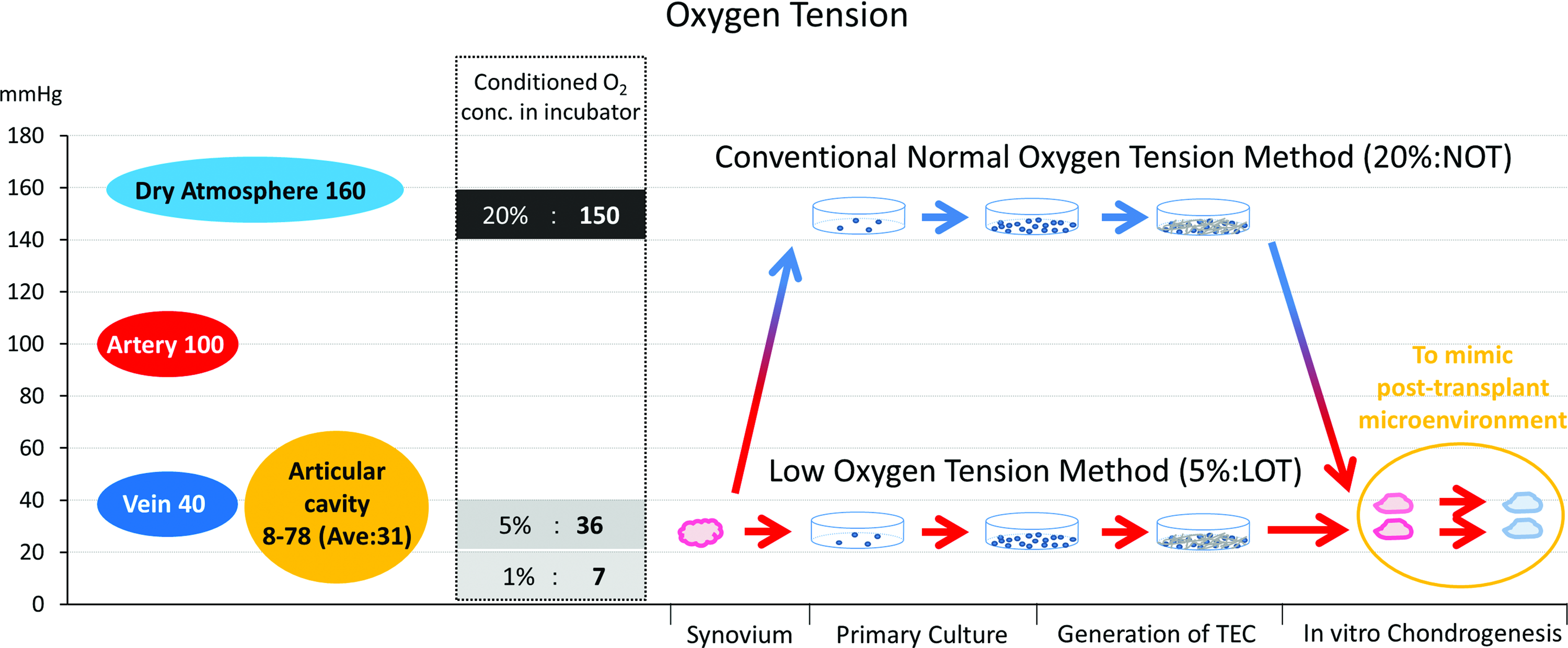
Oxygen tensions in the human body and in standard incubator conditions (left), and a schematic representation of the procedures used in the present study and their relationship to oxygen tension. LOT, low oxygen tension; NOT, normal oxygen tension. Color images available online at www.liebertpub.com/tea
The purpose of the present study was to test our hypothesis that LOT preparation could promote the generation and subsequent chondrogenic differentiation of the TEC to develop larger more hyaline-like cartilaginous tissue constructs that could be applicable to clinically enhance cartilage repair.
Materials and Methods
LOT and normal oxygen tension culture of human synovial MSCs
The usual oxygen tension in incubators is much higher than that in the native intraarticular environment, 21 and it is also higher than that of the arterial blood (Fig. 1). Conversely, 5% oxygen mimics more closely the physiological cellular microenvironment within a joint. By extrapolation, a method to calculate the partial pressure of PO2 in inspired air (PiO2) in respiratory physiology 22 can be extended to the incubator situation according to the following: the oxygen tension (mmHg) in an incubator can be calculated using the formula (Instruction of Panasonic Healthcare: Multigas Incubator MCO-18M): (760 mmHg [atmospheric pressure] −47 mmHg [saturated vapor pressure at 37°C]) × conditioned O2 concentration (%). This formula is based on Dalton's law of partial pressure in physical chemistry. 23 Because we cannot readily control the postimplant intraarticular microenvironment, LOT preparation alone may exert beneficial effects after clinical implantation of TEC. Therefore, we have evaluated the feasibility of using LOT in TEC preparation by comparing 20% and 5% O2 conditions during cell isolation, expansion, and generation of TEC and subsequently evaluating the effectiveness of chondrogenic differentiation under 5% O2 conditions for both the groups to mimic the posttransplant cellular microenvironment.
Harvest of synovium and isolation of human synovial MSCs
MSCs were isolated and expanded as previously reported. 24 Briefly, synovium was obtained from six human donors at the time of arthroscopic surgery of the knee in accordance with a protocol approved by the Institutional Ethics Committee. The age and gender of the individual patients are listed in Table 1. For isolation of synovial MSCs, synovial specimens were obtained aseptically, rinsed with sterile phosphate-buffered saline (PBS), sheared into small fragments using surgical scissors, and digested with 0.2% collagenase (Collagenase-AOF-Type A; Worthington Biochemical) in high-glucose Dulbecco's modified Eagle's medium (DMEM; Gibco BRL, Life Technologies) for 2 h in a shaking water bath at 37°C. After neutralization of the collagenase using growth medium containing DMEM supplemented with 10% fetal bovine serum (Sigma-Aldrich) and 1% penicillin–streptomycin (Sigma-Aldrich), the cells were collected by centrifugation, resuspended in the growth medium, and subsequently plated in culture dishes. These cells were defined as MSCs. For isolation and expansion, MSCs were cultured in the growth medium at 37°C in a humidified atmosphere of 5% CO2 and 5% or 20% O2, with the medium replaced twice per week. When the cells had reached confluence, they were then washed with PBS, detached with trypsin–EDTA (0.25% trypsin and 1 mM EDTA; Gibco BRL, Life Technologies), and replated at a density of 3.0 × 103 cells/cm2. Regarding the proliferation rate, we calculated the ratio of cell numbers in LOT-MSCs to those in the normal oxygen tension (NOT)-MSCs at each time of passage (from P1 to P10). At the cell inoculation stage for each passage, we used the same number of the cells (3 × 103 cells/cm2) for both the groups. Note: Since LOT-MSCs grew faster than NOT-MSCs, the passaging was always done when the LOT-MSCs became confluent. The result presented was the average of six samples.
Senescence-associated β-galactosidase staining
Cells were cultured in 12-well plates, and the senescence-associated-β-galactosidase (SA-βgal) staining was performed with a Cellular Senescence Assay Kit (Cell Biolabs, Inc.). SA-βgal catalyzes the hydrolysis of X-gal, which produces a blue color. To perform the assays, we divided each well into four areas. A high-power field (×80) was randomly selected from each area, and the cell number was counted. We counted the cell number using three wells in either the NOT or the LOT group (12 fields per each group). Subsequently, the ratio of SA-βgal-stained cells to total cells was calculated using ImageJ software (National Institutes of Health), and the results are shown as mean ± SD. SA-βgal staining was also performed on NOT- and LOT-TEC.
Development of the basic TEC
Human MSCs at passage 3 were plated in 12-well culture dishes at a density of 4.0 × 105 cells/cm2 in the growth medium containing 0.2 mM ascorbate-2-phosphate (Sigma-Aldrich), an optimal concentration based on results from earlier studies, 9 and cultured under 5% or 20% O2 conditions. After 14 days in culture, monolayer complexes of the cultured MSCs and the ECM synthesized by the cells developed into sheet-like constructs, and these constructs could be detached from the substratum by application of shear stress using gentle pipetting. The released constructs were left in suspension to spontaneously form three-dimensional structures by active tissue contraction. These contracted tissues were termed basic tissue-engineered constructs.9,10 The weight and volume of TEC developed in each O2 tension were weighed and measured using an electric balance (AG204; Mettler Toledo) and a plethysmometer (TK-101CMP; UNICON Japan), respectively.
Evaluation of collagen content of the basic TEC
The collagen content of the NOT-TEC and LOT-TEC was evaluated by quantification of hydroxyproline. The measurement of hydroxyproline was performed as previously described. 25 Briefly, the basic TEC was hydrolyzed in 100 mL of 6 N HCl at 105°C for 18 h. The solution was evaporated, and the residue was dissolved in 100 μL distilled water, oxidized with 7% chloramine-T, and reacted with Ehrlich reagent under acidic conditions. The absorbance of the extracted products was assessed and quantified at 558 nm with a spectrophotometer.
Chondrogenic differentiation of TEC
TEC was allowed to reattach to culture dishes and further cultured in growth medium supplemented with 0.2 mM Asc-2P (Sigma-Aldrich) for 1 day. The TEC was then cultured in a chondrogenic medium for another 21 days, with the medium changed three times per week. The chondrogenic medium consisted of DMEM supplemented with 1% insulin–transferrin–selenium supplement (ITS + Premix; Corning, Inc.), 0.2 mM Asc-2P (Sigma-Aldrich), and 200 ng/mL recombinant human BMP2 (Osteopharma, Inc.). To mimic the postimplant microenvironment, chondrogenic differentiation was performed only under the 5% O2 conditions for both the groups. Using the same oxygen tension during chondrogenesis enabled the comparative evaluation of the effectiveness of the two preparatory O2 conditions on the subsequent chondrogenic potential of the two types of TEC (Fig. 1).
Quantitative reverse transcription–polymerase chain reaction
Quantitative reverse transcription–polymerase chain reaction (RT-PCR) was used to evaluate the expression levels for a cellular senescence-associated gene (h-P16), cartilage-specific genes (h-SOX9, h-COL2A1, and h-ACAN), and a housekeeping gene (h-GAPDH). Total RNA from MSCs, basic TEC, and chondrogenic differentiated TEC were extracted with RNeasy Mini Kit for MSCs and RNeasy Fibrous Tissue Kit for TEC (QIAGEN), and complementary DNAs were obtained by reverse transcription of RNA (Superscript III; Invitrogen) with oligo(dT)20 as primers. Gene expression levels were quantified by real-time PCR TaqMan gene expression assays (h-P16; Hs00923894_m1, h-SOX9; Hs01001343_g1, h-COL2A1; Hs00264051_m1, h-ACAN; Hs00153936_m1, h-GAPDH; Hs02758991_g1; Applied Biosystems), and values were then normalized to the corresponding expression levels for h-GAPDH.
Biochemical analysis of sulfated glycosaminoglycan content of TEC
TEC differentiated toward chondrogenesis was digested (at 65°C for 3 h) with 0.01% papain (Sigma-Aldrich) in 0.2 M sodium phosphate buffer (pH 6.4) containing 8 mg/mL sodium acetate, 4 mg/mL EDTA sodium salt, and 0.8 mg/mL cysteine HCl. Subsequently, measurement of glycosaminoglycan (GAG) content was performed using a Blyscan Glycosaminoglycan Assay Kit (Biocolor Life Science Assays). Chondroitin 4-sulfate (Biocolor Life Science Assays) was used as a standard. The content of GAGs (in μg) was expressed as GAG weight per sample and GAG weight per wet weight of the differentiated TEC.
Histology of TEC differentiated toward chondrogenesis
The differentiated TEC was fixed with 4% paraformaldehyde for a day, followed by dehydrating with ethanol, clearing with xylene, and embedding in paraffin. Approximately 3-μm sections were cut and stained with safranin O–fast green and toluidine blue for sulfated GAGs. Serial sections were stained with picrosirius red and observed under Brightfield and polarized light microscopy to assess collagen fibrils in the TEC. Picrosirius red staining can detect type I collagen as a yellow band and type III collagen as a green band simultaneously when observed under polarized microscopy. Type II collagen also stains red with this dye but is not observed under polarized microscopy.26,27 Subsequently, a fibrous morphology becomes apparent. Immunohistochemical (IHC) analyses for type II collagen were performed on sections using primary antibodies from Daiichi Fine Chemical Co. Ltd. (anti-human type II collagen, F-57). Secondary antibodies and avidin–biotinylated enzymes (Vectastain Elite ABC Mouse IgG Kit) were used, followed by the DAB reagent (Dako), and sections were counterstained with hematoxylin.
Statistical analyses
Each in vitro experiment was repeated at least three times using different cell sources with similar results. The data were subjected to an independent-samples t-test using computer-based JMP Pro 11 (SAS Institute). The results are expressed as mean ± SD of triplicate determination. For all analyses, p < 0.05 was used to indicate statistical significance.
Results
Proliferative capacity of human MSCs under NOT and LOT conditions
MSCs expanded in the NOT conditions experienced a slowing in proliferation rate around passage 4, while those cultured in LOT conditions maintained their growth rates (Fig. 2a). The relative proliferation rate under LOT conditions compared to that in NOT conditions was 1.3 times at passage 1, and the ratio gradually increased during the cell culture, with a maximum difference of 1.9 times detected at passages 6 and 7. After passage 8, the ratio tended to decrease as the MSCs in LOT conditions started to exhibit slower proliferation rates (Fig. 2b).

The effect of oxygen tension on the proliferation of synovial MSCs.
Cellular morphology and staining of human MSCs for the senescence marker β-galactosidase
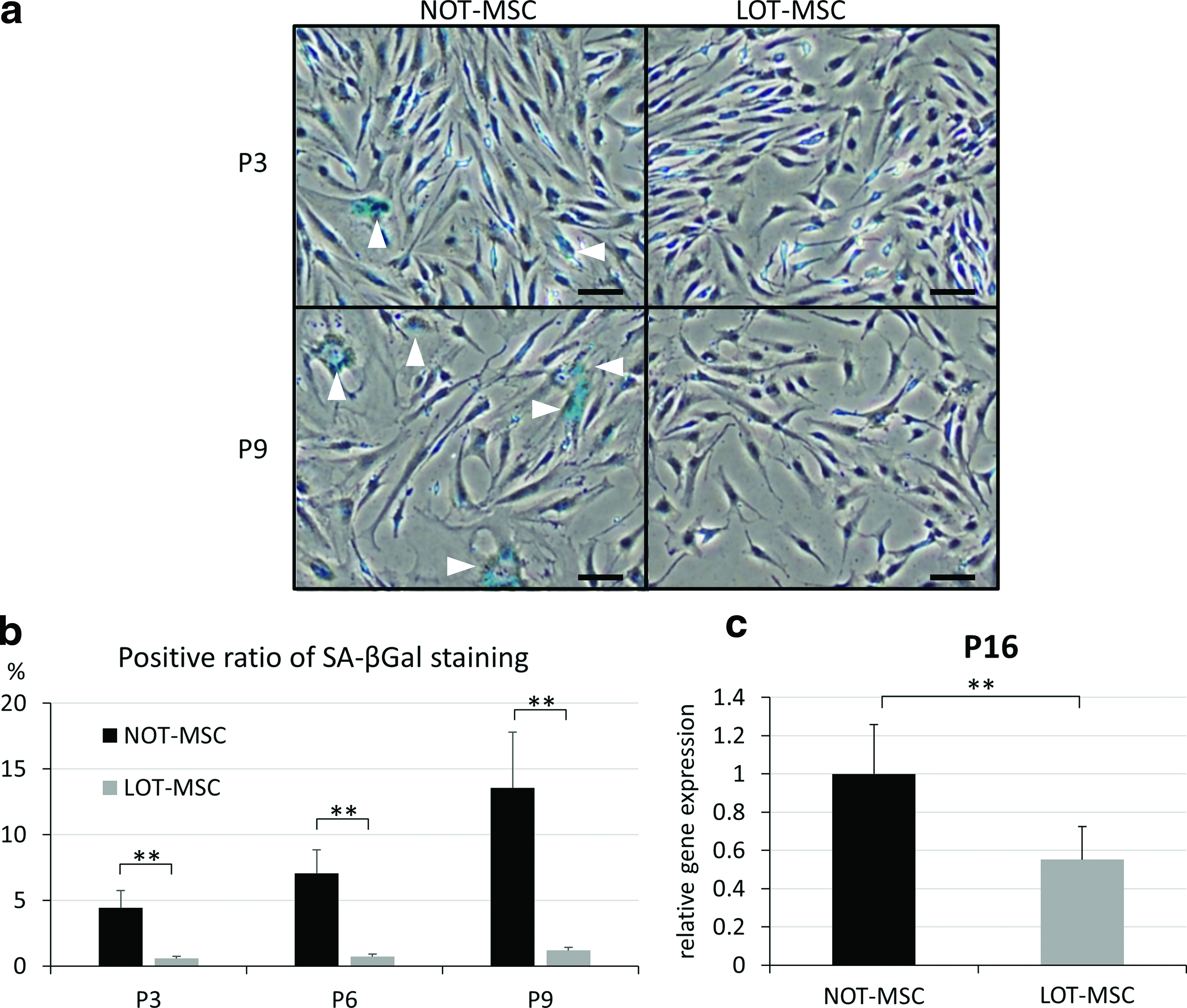
Senescent cells generally enlarge, often doubling in volume, and adopt a flattened morphology. 28 In the present study, the proportion of cells exhibiting an enlarged morphology increased with passage number, and these senescence-associated changes were initiated in earlier passages for MSCs cultured under NOT conditions (Fig. 3a).
Evaluation of cellular senescence of MSCs cultured under NOT and LOT conditions.
Histochemical staining for SA-βgal is commonly used to identify cells undergoing senescence. 29 At passage 3, MSCs cultured under NOT conditions included a small number of enlarged flattened cells, which were a sign of early cellular senescence. In contrast, MSCs cultured under LOT conditions maintained their spindle shape, and most of the cells were negative for SA-βgal staining (Fig. 3a). The percentage of SA-βgal-stained MSCs cultured under NOT and LOT conditions was 4.4% and 0.6% at passage 3 (p < 0.01), 7.0% and 0.7% at passage 6 (p < 0.01), and 13.6% and 1.2% at passage 9 (p < 0.01), respectively (Fig. 3b). These results indicated that culture under the LOT conditions prevented development of cellular senescence in these MSC cultures.
Senescence-related gene expression of human MSCs cultured under LOT conditions
Induction of cellular senescence in MSCs is tightly associated with the upregulation of P16 gene expression.17,28 Compared with MSCs cultured under NOT conditions, the expression levels for P16 were 1.81-fold lower (p < 0.01) in MSCs cultured under LOT conditions at passage 3 (Fig. 3c).
Weight, volume, and hydroxyproline content of human TEC developing under LOT and NOT conditions
The resultant weight of TEC developing in NOT and LOT conditions was 2.20 mg/cm2 and 2.04 g/cm2, respectively. The resultant volume of the TEC was 2.14 and 1.99 cm3/cm2, respectively (Fig. 4a). The gross appearance of the LOT-TEC was nearly identical to that of the NOT-TEC. The hydroxyproline content of the NOT- and the LOT-TEC was 1075 and 1020 ng/cm2, respectively (Fig. 4b). These results for weight, volume, and hydroxyproline content of the basic TEC did not show significant differences between the two groups. Such results indicate that preparation under LOT conditions does not affect tissue formation of the resultant TEC.

Evaluation of NOT-TEC and LOT-TEC before chondrogenic differentiation.
Gene expression in human TEC developed under NOT and LOT conditions
Expression levels for P16 were 2.70-fold lower (p < 0.01) in LOT-TEC compared with the NOT-TEC, indicating a suppression of cellular senescence induction in MSCs in LOT-TEC cultures. Expression levels for SOX9 were 2.60-fold higher (p < 0.01) in LOT-TEC compared with the NOT-TEC. In contrast, COL2A1 gene expression was not detected in the TEC grown under either condition (Fig. 4c). These results indicate that preparation of TEC under LOT conditions alone did not induce chondrogenesis but did increase the chondrogenic potential of the resultant TEC.
Senescence-associated β-galactosidase staining of TEC
It has been suggested by a previous report that senescence-associated βgal expression is also induced by prolonged confluence in culture. 30 Therefore, a protective effect from senescence of LOT on synovial MSCs may potentially be compromised if synovial MSCs were maintained at a high density for a prolonged time, such as within a TEC. To ascertain whether this was the case, NOT-TEC and LOT-TEC were stained for SA-βgal. NOT-TEC was positive for SA-βgal staining (Fig. 5a, c), while LOT-TEC was negative for detectable SA-βgal staining (Fig. 5b, d).
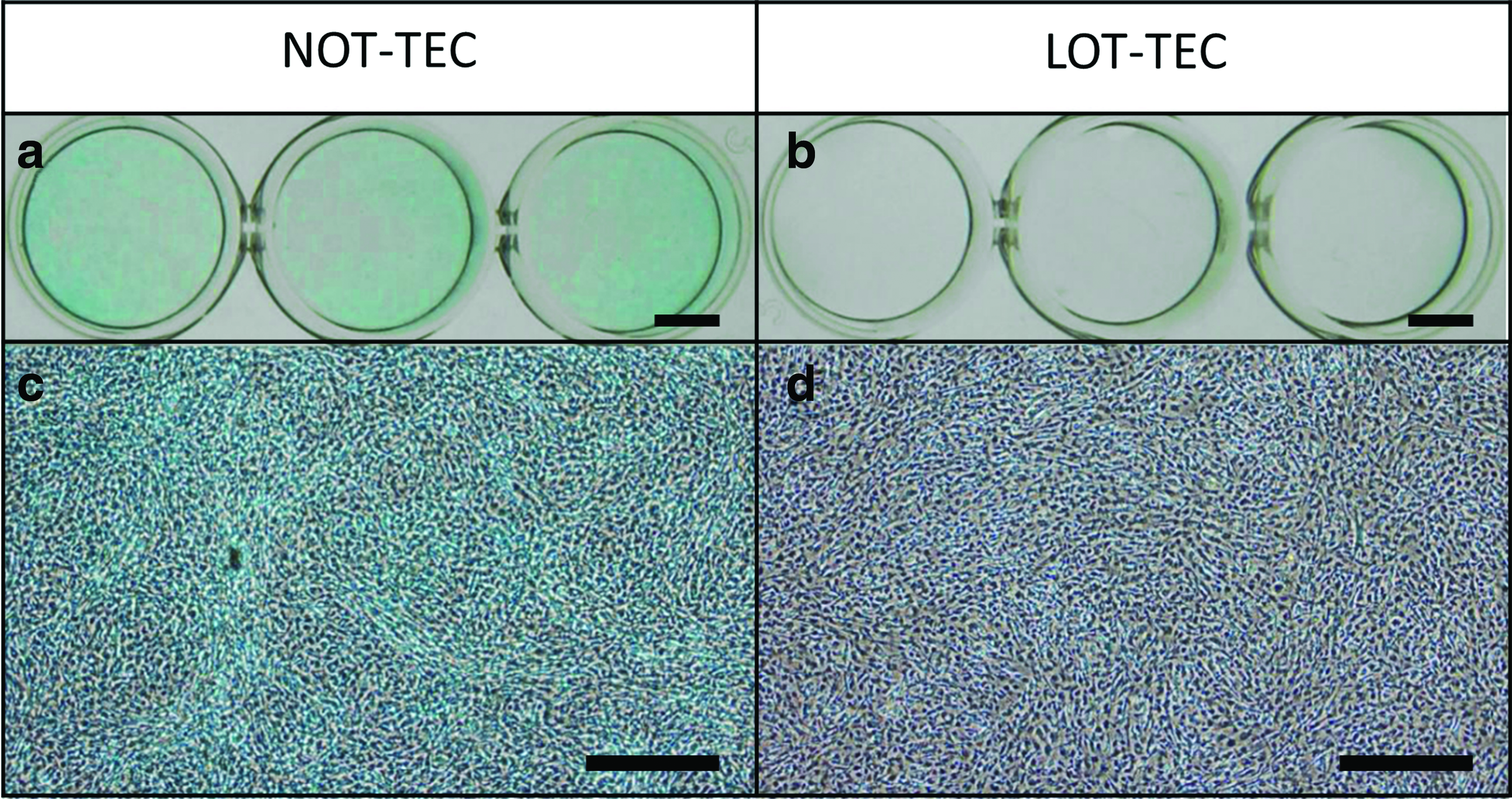
Senescence-associated β-galactosidase staining of NOT- and LOT-TEC derived from P3 MSCs. Staining (blue) was stronger in NOT-TEC
GAG content of TEC subjected to chondrogenic differentiation
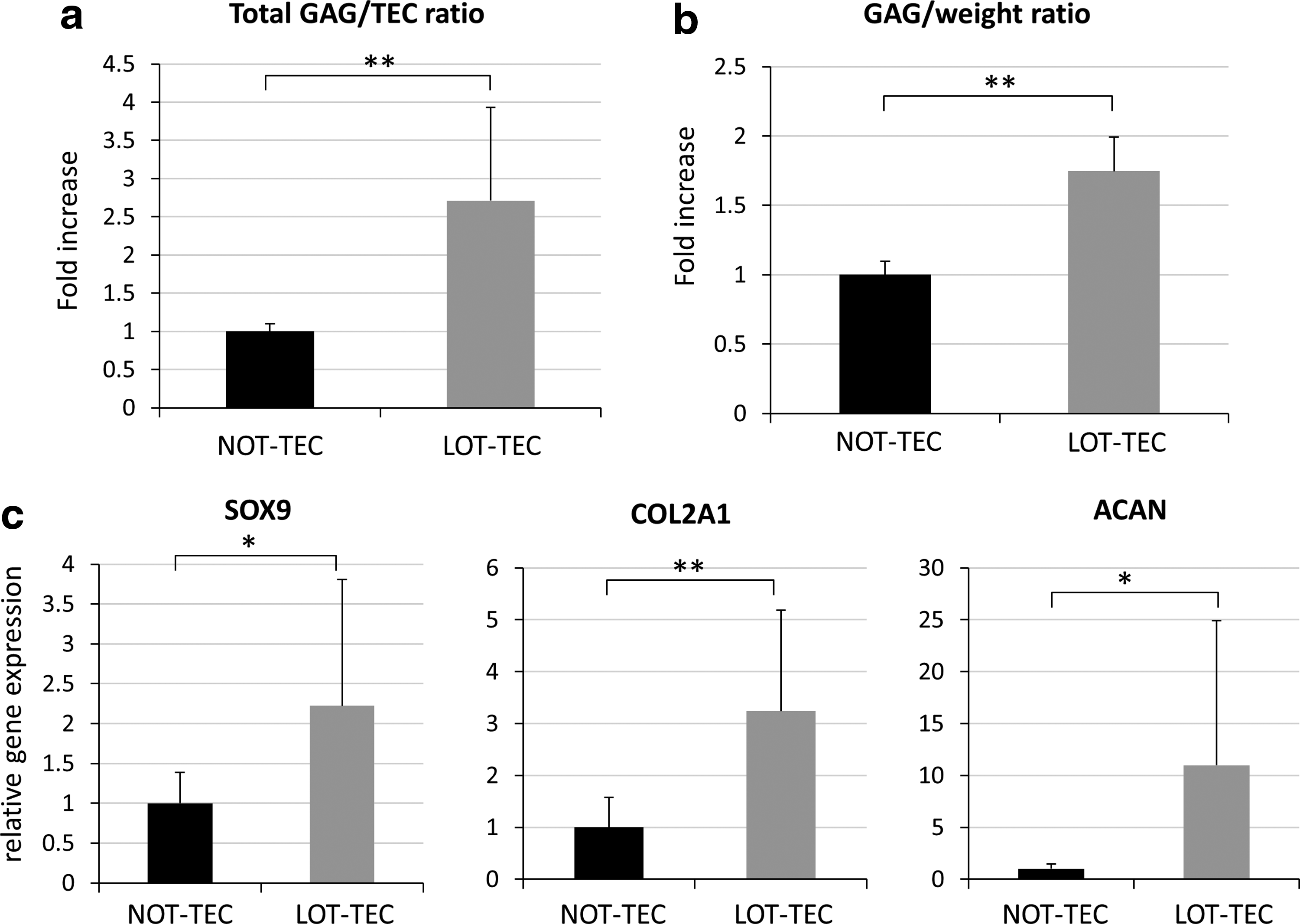
GAG/sample in LOT-TEC following chondrogenic differentiation was 2.71-fold higher (p < 0.01) compared with levels in the NOT-TEC, indicating increased GAG production by those prepared under LOT conditions (Fig. 6a). GAG/wet weight for the LOT-TEC was 1.75-fold higher (p < 0.01) compared with levels in the NOT-TEC, again indicating increased GAG concentrations in the differentiated cartilage-like tissue by preparation under LOT conditions (Fig. 6b). These results indicate that TEC exposed to LOT conditions during development can potentially produce more cartilage-like ECM after implantation in vivo.
Evaluation of chondrogenic differentiated TEC initially prepared under NOT and LOT conditions.
Gene expression of NOT- and LOT-TEC after chondrogenic differentiation
Expression levels for SOX9, COL2A1, and ACAN were 2.22-fold higher (p = 0.039), 3.25-fold higher (p < 0.01), and 11.0-fold higher (p = 0.047), respectively, in LOT-TEC compared with NOT-TEC (Fig. 6c).
Histology of TEC differentiated toward the chondrogenic lineage in vitro
Chondrogenic differentiation was performed under LOT conditions for both the NOT-TEC and LOT-TEC groups to evaluate the efficacy of preparation under LOT conditions. The LOT-TEC exhibited a more hyaline cartilage-like morphology with stronger safranin O and toluidine blue staining than did the NOT-TEC (Figs. 7c–f and 8a, b). These results are complementary to the earlier results regarding the quantification of GAG content in the chondrogenic differentiated TEC. Picrosirius red staining observed under polarized light microscopy revealed bright yellow bands within the NOT-TEC (Figs. 7g and 8g), indicating the presence of fibrous-like tissue within this TEC. In contrast, the LOT-TEC was negative for such bright bands (Figs. 7h and 8h), indicating an absence of a fibrous component and a more hyaline-like cartilaginous morphology. IHC analysis for type II collagen was strongly positive throughout the matrix of LOT-TEC, whereas type II collagen staining was absent in the NOT-TEC (Figs. 7i, j and 8j, k), consistent with the observed presence of the fibrous areas detected as bright bands by picrosirius red staining in the NOT-TEC and, thus likely, the presence of type I collagen. Each staining revealed that the morphology of LOT-TEC was quite similar to that of normal human articular cartilage (Fig. 8c, f, i, l).
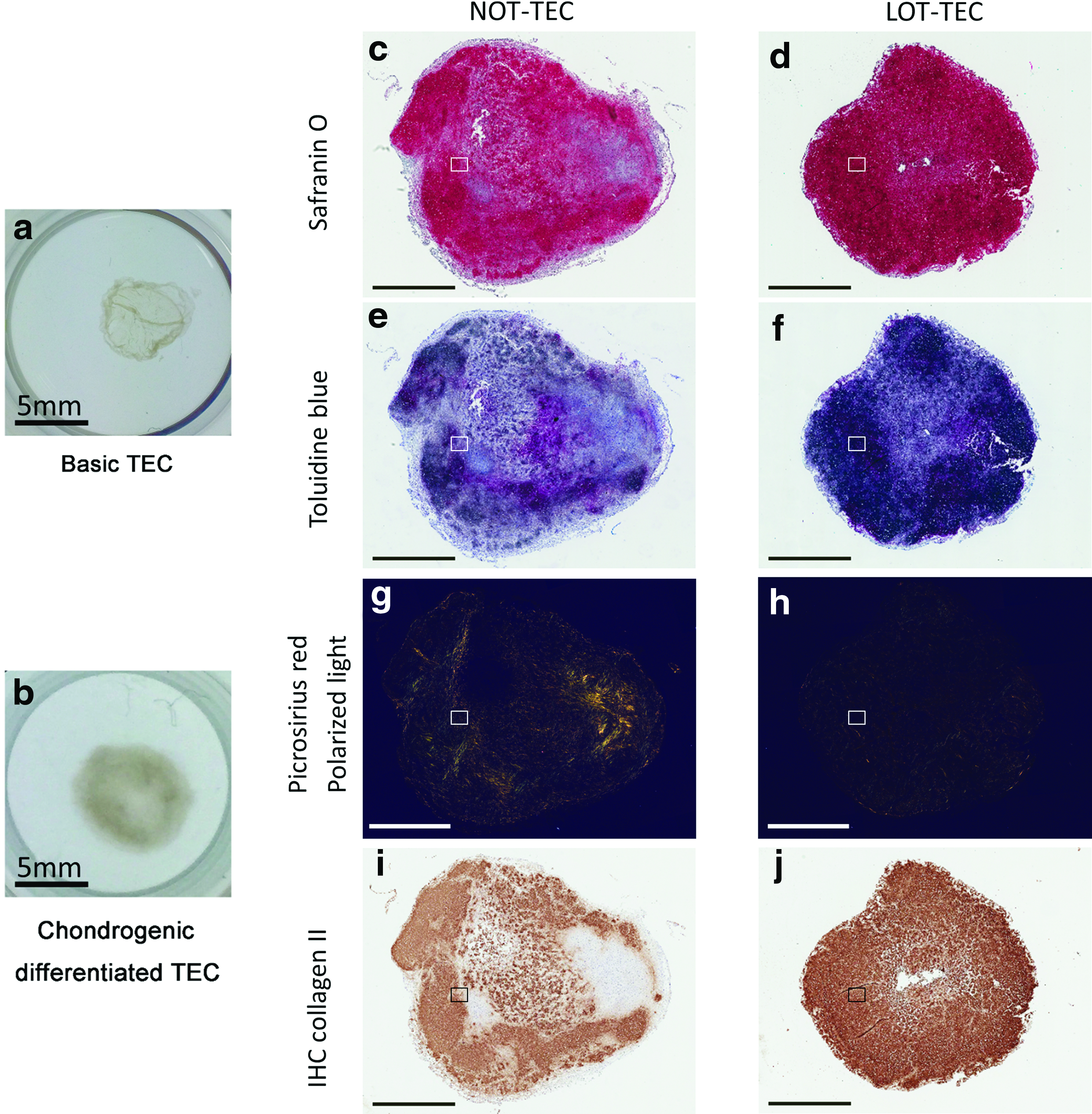
Histological evaluation of chondrogenic differentiated LOT- and NOT-TEC. The cell source was patient 4. Chondrogenic differentiation was performed under 5% oxygen for both the NOT and LOT groups to evaluate the influence of oxygen tension during TEC preparation. During the chondrogenic differentiation of TEC, MSCs produced extracellular matrices, and a sheet-like structure
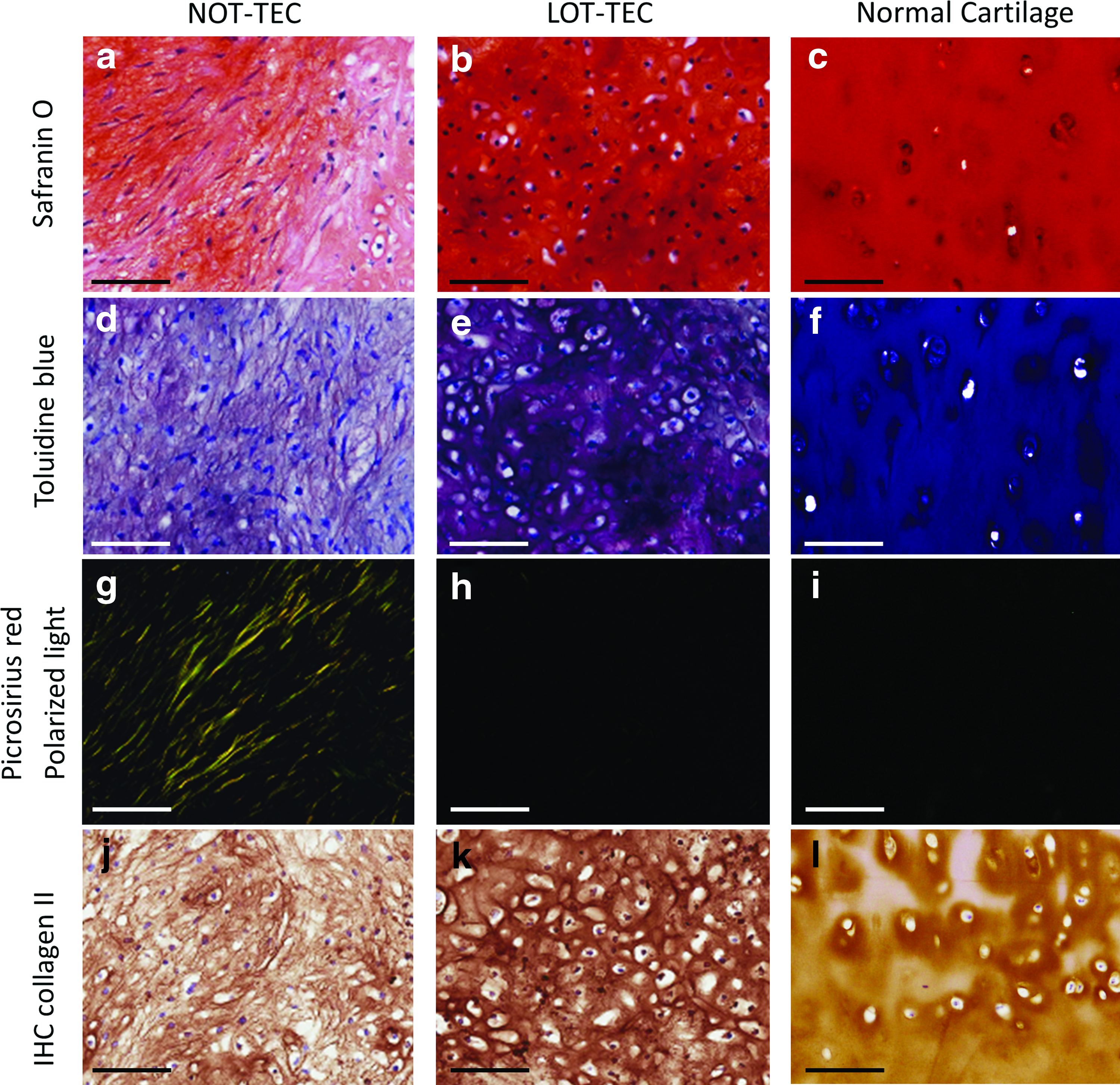
Histological evaluation of chondrogenic differentiated LOT- and NOT-TEC in higher magnification; comparison with human articular cartilage without osteoarthritic change. Safranin O–fast green staining of TEC
Discussion
Primary culture of MSCs has conventionally been performed at oxygen tensions of 150 mmHg, which is higher than that of normal arterial blood (100 mmHg) (Fig. 1). Although the CO2 concentration of a standard incubator is routinely adjusted to mimic the physiological cellular microenvironment, the corresponding oxygen tension is usually not adjusted to tissue-specific physiological ranges. The present study demonstrated that the LOT culture conditions (5% O2) promoted several capacities of synovial MSCs, including (1) prevention of senescence induction, (2) promotion of cellular proliferation, and (3) enhanced chondrogenic differentiation. In contrast, there have been reports on the negative effects of hypoxia regarding the proliferation and differentiation capacity of MSCs. These contradictory data might be related to the PO2 chosen, 31 which is why we chose LOT conditions that more accurately mimicked the intraarticular environment. Severe hypoxia of less than 2% O2 might impede the proliferation and differentiation capacity of MSCs,31–33 and therefore, 5% O2 was used to parallel the oxygen tension reported to be in the articular cavity (8–78 mmHg [average: 30.7 mmHg]) 21 (Fig. 1).
In many stem cell protocols, MSCs are expanded, usually in 20% O2, for 3–5 weeks of culture.2,12,34,35 The present study provided evidence that cellular senescence had already started during early passages in NOT conditions, and thus, LOT culture conditions for MSCs may prevent the onset of some features of cellular senescence and, thus, could contribute to more sustained cellular proliferation. As a consequence of this sustained MSC proliferation, shorter culture times would be required to generate a clinically useful number of cells. Shortening of culturing duration for cellular expansion could also be clinically beneficial in reducing the risk of contamination of the cultures.
Notably, the LOT-TEC responded to the chondrogenic media with differentiation to a more hyaline-like cartilaginous tissue without the apparent contamination by fibrous tissue development, whereas the NOT-TEC 13 differentiated into a mixture of hyaline and fibrous cartilaginous tissues. The gene expression levels for SOX9 before and after chondrogenic differentiation were significantly higher at either point for the LOT-TEC. It is well known that expression of HIF-1alpha, which is a positive regulator of SOX9, is elevated through LOT. 16 In the present study, it required approximately 3 weeks for cellular expansion under LOT condition. Thus, exposure of MSCs to LOT conditions for such durations may have contributed to the elevated SOX9 expression levels in the LOT-TEC, potentially leading to enhanced chondrogenic differentiation compared to that observed for the NOT-TEC. Because chondrogenic differentiation was performed under LOT condition for both the NOT- and LOT-TEC in this study, NOT-TEC also benefits from the promotive chondrogenic effect of LOT. Therefore, it is noteworthy that LOT-TEC exhibited additional enhancement of chondrogenic capacity from preparation under LOT conditions. Prevention of cellular senescence may be the reason for this additional enhancement of the chondrogenic differentiation capacity of LOT-TEC.
With regard to clinical relevance, the present study is the first demonstration of an in vitro-generated and implantable tissue-engineered hyaline cartilaginous tissue using human MSCs without the use of an exogenous scaffold. 6 Such TEC may be generated over a range of diameters and thicknesses,9,10 which are sufficiently large to match the sizes of in vivo lesions requiring repair. Previous studies to test the effect of LOTs on chondrogenesis of MSCs in vitro used pellet culture,17,18,36 micromass culture, 37 or monolayer culture 15 models. Thus, the size of the resultant differentiated tissues was likely too small to be used clinically as an effective implant. Recently, Yamashita et al. reported generation of scaffold-free hyaline-like cartilaginous tissues using human induced pluripotent stem (iPS) cells, 38 which were larger than those obtained from pellet cultures. However, such approaches using iPS cells require high cost to develop the required chondrogenic masses, and concerns remain regarding safety issues, including the potential contamination of cells differentiated into lineages other than mesoderm, as well as tumorigenicity.14,39 In contrast, the present study succeeded in generating a comparably well-differentiated hyaline-like cartilaginous tissue from synovial MSCs by only lowering the oxygen tension. Since the use of MSCs and their culture under low O2 conditions is a relatively simple and safe procedure, the present LOT-TEC method could be advantageous, with regard to not only safety but also cost-effectiveness in comparison with the use of pluripotent cells, such as iPS cells.
To demonstrate the feasibility of LOT-TEC to effectively repair articular cartilage, transplantation of basic LOT-TEC (before differentiation), as well as transplantation of chondrogenic differentiated LOT-TEC as an artificial hyaline cartilaginous implant, is planned in the near future. We plan to use larger animal models, such as rabbits or pigs, as we have preliminary data indicating that the LOT conditions used in the present study also positively impact MSCs and TEC from these species as well (unpublished data).
In summary, generation of an apparently well-differentiated hyaline-like cartilaginous tissue was achieved by expansion of human synovial MSCs under LOTs, starting from the isolation of the cells from the synovium until being integrated into TEC. Such LOT-TEC could be generated in vitro with variable implantable sizes for subsequent potential in vivo repair of cartilage defects of various sizes. Thus, MSC culture at more physiological oxygen tensions could be a relatively safe and cost-effective procedure, and therefore, with further feasibility studies as well as additional safety studies, this procedure could potentially become a promising method toward facilitating regenerative repair of cartilage.
Footnotes
Acknowledgments
This study was supported by a Grant-in-Aid for Scientific Research (B), Japan Society for the Promotion of Science, Japan. DAH was supported by the Alberta Innovates Health Solutions OA Team Grant.
Disclosure Statement
No competing financial interests exist.