
Abstract
Recent advances in shape memory polymers have enabled the study of programmable, shape-changing, cytocompatible tissue engineering scaffolds. For treatment of bone defects, scaffolds with shape memory functionality have been studied for their potential for minimally invasive delivery, conformal fitting to defect margins, and defect stabilization. However, the extent to which the osteogenic differentiation capacity of stem cells resident in shape memory scaffolds is preserved following programmed shape change has not yet been determined. As a result, the feasibility of shape memory polymer scaffolds being employed in stem cell-based treatment strategies remains unclear. To test the hypothesis that stem cell osteogenic differentiation can be preserved during and following triggering of programmed architectural changes in shape memory polymer scaffolds, human adipose-derived stem cells were seeded in shape memory polymer foam scaffolds or in shape memory polymer fibrous scaffolds programmed to expand or contract, respectively, when warmed to body temperature. Osteogenic differentiation in shape-changing and control scaffolds was compared using mineral deposition, protein production, and gene expression assays. For both shape-changing and control scaffolds, qualitatively and quantitatively comparable amounts of mineral deposition were observed; comparable levels of alkaline phosphatase activity were measured; and no significant differences in the expression of genetic markers of osteogenesis were detected. These findings support the feasibility of employing shape memory in scaffolds for stem cell-based therapies for bone repair.
Introduction
F
Synthetic biomaterials for use as bone grafts have been widely studied in an effort to overcome the limitations of autografts, allografts, and cements. Traditionally, synthetic bone grafts have possessed good osteoconductivity and osseointegration, while lacking osteogenicity and osteoinductivity 6 ; however, recent advances in tissue engineering and material science have made progress in addressing these shortcomings. 7 For example, improved osteoinductivity of synthetic bone grafts has been achieved through incorporation of ceramics, such as hydroxyapatite8,9 and β-tricalcium phosphate,10,11 and growth factors, such as bone morphogenetic proteins.12–14 Additionally, direct incorporation of stem cells into synthetic bone grafts has been shown to improve osteogenicity of the graft.15,16 Prior studies comparing treatments of critical size defects have demonstrated a significant increase in bone regeneration when stem cell-based bone tissue engineering constructs are employed compared with when cell-free constructs are employed.17,18
To improve treatment of bone defects, shape memory polymers (SMPs) have recently been studied for their potential for minimally invasive delivery, conformal fitting to defect margins, and defect stabilization. SMPs are a type of smart material that can undergo programmed changes in shape from a temporarily fixed shape to a permanently memorized shape when exposed to an external stimulus such as heat.19–24 For example, two previous in vitro studies have demonstrated the ability of SMP scaffolds to support osteoblast adhesion, proliferation, and osteogenic gene expression,21,22 but neither study investigated the effect of programmed changes in shape on the behavior of cells seeded in the SMP scaffolds. Moreover, Liu et al. 23 developed a poly(ɛ-caprolactone) scaffold containing hydroxyapatite that was used as a cell-free synthetic bone graft in an in vivo rabbit mandibular critical size defect model. That study found that the SMP scaffold promoted greater bone repair than sham controls. Additionally, we have reported the use of expanding SMP foams and contracting SMP sleeves as synthetic bone grafts in load-bearing applications and as supplementary fixation devices for stabilizing complex fractures, respectively. 25 Collectively, these studies have demonstrated the potential for employing programmable SMP scaffolds in treatment of bone defects.
Although cell-free, biomaterial-based strategies are an active area of research for this and other tissue engineering applications, cell-based strategies that combine cells with biomaterials are likely to remain widely pursued as well.26–28 As a result, because strategies being developed for SMP scaffolds generally involve both a trigger, such as a thermal trigger, and substantial deformation of the material and any cells resident therein, an obvious concern is that triggering and programmed shape change could negatively impact osteogenic differentiation capacity if employed in cell-based therapies.
Despite interest in SMPs for treatment of bone defects, the extent to which stem cell osteogenic differentiation capacity is preserved following programmed shape change has not yet been determined. In this study, we hypothesized that stem cell osteogenic differentiation could be preserved during and following programmed architectural changes in SMP scaffolds. To test this hypothesis, stem cell osteogenic differentiation capacity was assessed before and after heat-triggered scaffold shape change of expanding SMP foam scaffolds and contracting SMP fibrous scaffolds. Differentiation capacity in shape-changing and control scaffolds was then compared by characterizing mineral deposition, protein production, and gene expression.
Material and Methods
Scaffold preparation
Foam scaffold preparation
Porous SMP foam scaffolds, which we have previously studied in vivo in a cell-free approach for grafting of bone defects, 25 were fabricated through a modified porogen-leaching method 29 (Supplementary Methods 1 and 2; Supplementary Data are available online at www.liebertpub.com/tea). An acrylate-based chemistry25,30 was used to prepare scaffolds with a glass transition temperature (Tg) slightly above 37°C (Supplementary Method 2). To improve the relatively poor cell attachment properties of the naïve scaffold surface chemistry, foam scaffolds were coated with polydopamine (Supplementary Method 3) as polydopamine has been shown to promote cell attachment and osteoconductivity.22,31
For cell studies, cylindrical foam plugs of 3.5 mm diameter and 4–4.5 mm height were used. As-prepared foam scaffolds were used as the static control samples and contained a cuboidal pore morphology. Active shape-changing foam scaffolds were programmed into a temporary compressed state by uniaxial compression to 25% strain (Supplementary Method 4). Before cell studies, static and active scaffolds were sterilized through exposure to mononitrogen oxide gas (Supplementary Method 5).
Electrospun fibrous scaffold preparation
Aligned fibrous scaffolds were fabricated, as previously described 32 (Supplementary Method 6), by electrospinning a custom-synthesized thermoplastic polyurethane (TPU),33,34 which we have previously studied in vivo for stabilization of bone defects. 25 To prepare static aligned control scaffolds, as-spun aligned scaffolds were further stabilized using a thermal treatment protocol 32 (Supplementary Method 6) to ensure that fiber alignment was maintained. Active scaffolds were programmed to 100% strain (Supplementary Method 7). Before cell studies, static and active scaffolds were sterilized through exposure to mononitrogen oxide gas (Supplementary Method 5).
Cell culture
Human adipose-derived stem cells (hASCs) were used to investigate the effect of programmed shape change on stem cell osteogenic differentiation capacity. hASCs were chosen due to their potential for osteogenic differentiation and because of their research relevance as they are frequently used in bone tissue engineering strategies. 35
Before scaffold experiments, hASCs (Cat no. R7788-115; Life Technologies) were expanded in basal growth medium (Supplementary Method 8) in a 37°C humidified incubator with 5% CO2. Cells were passaged at 80% confluence using TrypLE Express solution (12604-054; Life Technologies). Cells were used at passage 6 for scaffold experiments. Before cell seeding, sterilized scaffolds were immersed in antibiotic–antimycotic solution (A5955; Sigma-Aldrich) overnight at 4°C to ensure the sterility of scaffolds, followed by rinsing in phosphate-buffered saline (PBS). The sterilized scaffolds were hydrated in basal growth medium at room temperature for 20 min.
To determine whether programmed shape change affects stem cell osteogenic differentiation capacity, hASCs were seeded in static and active scaffolds and cultured using a two-phase process. During the first phase, cells were cultured in scaffolds at a cytocompatible temperature (30°C) that does not trigger recovery of active scaffolds; for the second phase, scaffolds were heated to 37°C to trigger recovery of the active scaffolds. For the foam scaffolds, hASCs were seeded into the foam scaffolds using a vacuum-assisted technique (Supplementary Method 9) at a seeding density of 500,000 cells/mL. For the fibrous scaffolds, droplets of a cell suspension were laid on each scaffold with a cell density corresponding to 80,000 cells/cm2 for the total scaffold area of 0.8 cm2. The scaffolds were incubated at 30°C for 5 h, allowing cells to attach to the scaffolds, and extra basal growth medium was added to each sample.
After 24 h of incubation at 30°C, the basal growth medium was switched to osteogenic differentiation medium (Supplementary Method 8) and changed every 2 days. After incubation at 30°C for 5 days, scaffolds were incubated at 37°C, triggering architecture change, for 23 days. Osteogenic differentiation was characterized at three time points: before triggering (30°C for 5 days); 2 days post-triggering (37°C for 2 days); and 23 days post-triggering (37°C for 23 days). Three independent replicates consisting of cell expansion through the scaffold experiments were performed (n = 3; within each repetition, three to six technical replicates were included for different assays).
Scanning electron microscopy
To qualitatively visualize mineral deposition, scanning electron microscopy (SEM) was performed. Scaffolds were sputter coated with gold, and images were acquired in secondary electron mode. Representative images were taken at different areas to visualize scaffold architecture and mineral deposition at three different time points. Moreover, elemental analysis of mineral nodule deposition on the foam scaffolds and the fibrous scaffolds was conducted using energy-dispersive x-ray spectroscopy (EDS, JSM5600; JEOL).
Xylenol orange staining
To detect calcium deposition, xylenol orange (XO) staining—a nondestructive staining method 36 —was used. XO powder (398187; Sigma-Aldrich) was dissolved in deionized water and filtered to make a sterile stock solution of 20 mM and stored at 4°C. The scaffolds were cultured at the final concentration of 40 μM XO in medium overnight before imaging. The XO-containing medium was replaced with fresh Hoechst-containing medium (0.1 mg/mL, Hoechst 33342; Life Technologies) 30 min before imaging to stain for cell nuclei.
Fluorescent images of XO-stained scaffolds were taken using an N3 filter cube and images of cell nuclei taken using an A4 filter cube using an inverted microscope (DMI 4000B; Leica). Brightness and contrast of the micrographs of the fibrous scaffolds were adjusted in ImageJ to improve the contrast between the XO-stained calcium nodules and the background. The pixel intensity of the micrographs in the first two time points was adjusted to 5–100; in the last time point, the pixel intensity of the active and static fibrous scaffolds was adjusted to 75–255 and 44–255, respectively.
Microcomputed tomography
Three-dimensional (3D) analysis of mineral deposition was assessed using microcomputed tomography (μCT 40; Scanco Medical). Scaffolds were rinsed with PBS three times and fixed with 10% neutral buffered formalin overnight. Fixed scaffolds were rinsed with deionized water before drying in an evacuated desiccator overnight. Dried scaffolds were scanned at a nominal voxel size of 6 μm, and images were acquired at 55 kV, 144 μA, and 200 ms integration time. Mineral volume was measured for each scaffold by applying a threshold of 121.67 Hounsfield Units (HUs) for foam scaffolds and −21.48 HUs for fibrous scaffolds. A larger threshold was applied for foam scaffolds as they are more radio-opaque than the fibrous scaffolds.
Mineral volume for each scaffold was normalized by scaffold volume and compared between groups (n = 3). 3D images of mineral deposition were rendered using the 3D viewer in ImageJ. To determine average pore diameter, μCT cross sections were imported into ImageJ and the diameter of 15 pores per scaffold was measured.
Alkaline phosphatase activity
To determine osteogenic protein production, an alkaline phosphatase (ALP) colorimetric assay was performed using p-nitrophenyl phosphate (N1891; Sigma-Aldrich), as per the manufacturer's instructions (Supplementary Method 10). Cell lysates were reacted with p-nitrophenyl phosphate at 37°C for 1 h, and the reaction was stopped by adding 3 N NaOH. The absorbance was measured at 405 nm. ALP concentration was determined using 4-nitrophenol solution and normalized by DNA content.
Real-time quantitative polymerase chain reaction
To determine osteogenic gene expression, real-time quantitative polymerase chain reaction (qPCR) was performed. Total RNA extraction and cDNA analysis were conducted (Supplementary Method 11) before running qPCR. Real-time qPCR was performed in quadruplicate with 100 ng of cDNA in 20 μL reaction volume containing Power SYBR Green Master Mix (Invitrogen) and primers (RealTimePrimers) on Mastercycler ep realplex (Eppendorf).
Expression of the genetic markers of osteogenesis Runx2 and osteocalcin (OC) was examined, with normalization to reference genes B2M and RPL13A for cells cultured in foam and fibrous scaffolds, respectively (n = 3). B2M and RPL13A were determined to be the most stable housekeeping genes for foam and fibrous scaffolds, respectively, before performing qPCR (Supplementary Method 12).
Gene expression fold change was calculated as expression level normalized to the static control group at the first time point based on the method developed by Pfaffl et al. 37 Briefly, a fold change was calculated as the ratio of a target gene of a sample group versus the static control group at the first time point (30°C for 5 days) and expressed in comparison with the most stable housekeeping gene, taking into account the qPCR efficiency. A fold change higher than one indicates gene upregulation, while a fold changer lower than one indicates gene downregulation.
Histology
To visualize mineral deposition in each scaffold, histological analysis was performed using Alizarin red staining. Scaffolds were first rinsed with PBS three times and fixed with 10% neutral buffered formalin overnight. Due to the high porosity of foam scaffolds, thin histological sections could not show the extent of mineral deposition. Instead, entire foam scaffolds were stained with 2% Alizarin red with pH adjusted to 4.2 for 30 s (n = 3). Stained whole samples were extensively rinsed with deionized water. For fibrous scaffolds, fixed scaffolds were dehydrated using a series of ethanol solutions, embedded in paraffin, and sectioned into 3-μm-thick slices. The sections were stained with 2% Alizarin red with pH adjusted to 4.2 for 2 min (n = 2). Sections were then fixed with xylene and mounted. Only samples at the last time point (37°C for 23 days) were collected for staining.
Statistics
Paired t-test and one-way ANOVA with Tukey's HSD post hoc tests were performed for comparisons between active and static groups and comparisons within each group over time, respectively, when analyzing measurements of mineral volume, ALP activity, and gene expression using a commercial statistical analysis package (StatPlus:mac; AnalystSoft, Inc.). Statistical significance was set at p < 0.05.
Results
Scaffold architecture reorganization
Expanding SMP foam scaffolds and contracting SMP fibrous scaffolds underwent a programmed change in architecture upon heating to 37°C (Fig. 1). SMP foam scaffolds recovered 13% of the programmed strain following sterilization, but the programmed compressive strain remained stable at 30°C, as no recovery of the programmed strain was observed over the 5-day period (Fig. 1 Foam). Upon triggering recovery at 37°C, the foam completely recovered within 24 h. We speculate that the recovery during sterilization is due to a combination of programming the foam below its glass transition temperature and exposing the foam to a humid environment during sterilization. Foam scaffolds in the compressed state had a mean pore diameter of 216.9 ± 43.4 μm in the loading axis following the 5-day period at 30°C. Pore diameter increased to 297.3 ± 33.8 μm following recovery at 37°C for 2 days.

SMP foam and fibrous scaffolds are triggered to change shape and architecture under cytocompatible conditions. Programmed foam (left) scaffolds recover 13% of the programmed deformation (25% uniaxial compressive strain) following sterilization and remain stable for 5 days at 30°C (open circle); upon heating to 37°C, foam scaffolds recover all the programmed strain within 24 h (filled circle). Inset SEM micrographs reveal a compressed pore morphology that expands to an open pore morphology upon heating to 37°C. Fibrous (right) scaffolds that are programmed with 100% uniaxial tension recover 11% strain over 5 days at 30°C (open circle); upon heating to 37°C, the fibrous scaffolds recover 53% strain over the first 4 h and gradually recover 90% strain over the first 5 days. Inset SEM micrographs show an initial unaligned fiber orientation during the 5 days at 30°C that changes to an aligned orientation upon recovery at 37°C (filled circle). The much slower recovery rate of fibrous scaffolds compared with the foam scaffolds is due to differences in the scaffold architectures and chemistries. Scale bars for SEM micrographs are foam 500 μm and fibrous 50 μm. SMP, shape memory polymer; SEM, scanning electron microscopy.
Programmed SMP fibrous scaffolds showed no recovery of the programmed strain following sterilization, but recovered 11% of the programmed strain at 30°C over 5 days (Fig. 1 Fibrous). Upon triggering recovery at 37°C, fibrous scaffolds contracted 53% in the first 4 h and recovered 90% over the first 5 days. SEM micrographs show that although 11% strain was recovered at 30°C, fiber architecture remained unaligned (Supplementary Fig. S1). Triggering recovery at 37°C resulted in fibers reorganizing to a uniaxially aligned orientation.
Mineralized nodule formation
Results from SEM, XO staining, and μCT found no qualitative or quantitative difference of mineral deposition between active and static groups for either foam scaffolds or fibrous scaffolds. SEM analysis (Fig. 2) showed qualitatively comparable levels of calcium deposition between active and static samples for both foam scaffolds and fibrous scaffolds, and no difference in mineral nodule formation was qualitatively observed between active and static groups.
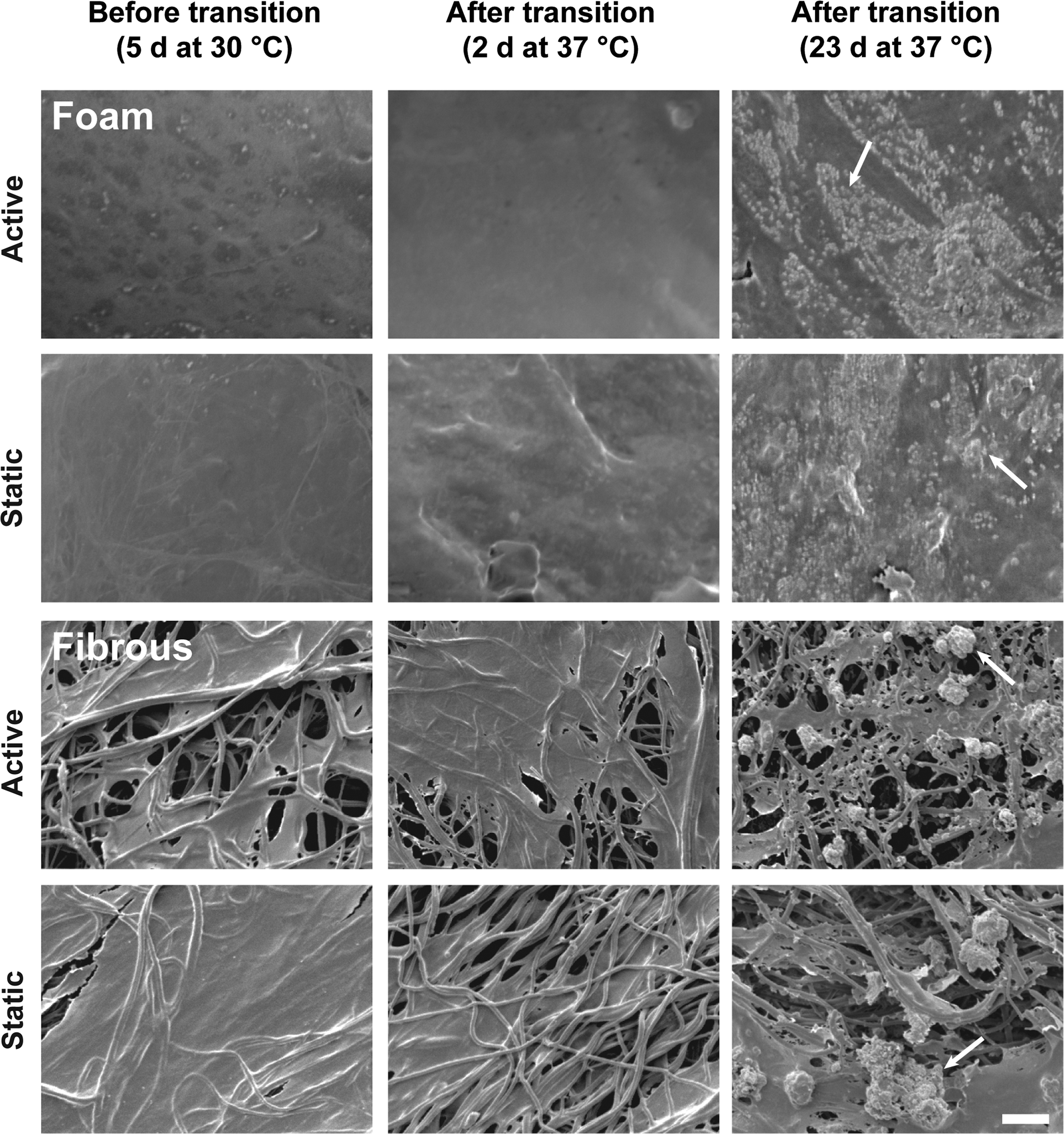
SEM reveals comparable mineral deposition for SMP foam and fibrous scaffolds for all time points. Both foam (top) and fibrous (bottom) scaffolds showed that no observable difference between active and static groups was found for either foam or fibrous scaffolds. No obvious calcium nodules were observed under SEM when cultured at 30°C for 5 days and 37°C for 2 days after triggering of architecture change. After 23 days at 37°C, a significant amount of calcium nodules was observed in both foam and fibrous scaffolds. Arrows highlight representative calcium deposition on each scaffold. Scale bar is 20 μm.
For foam scaffolds, no significant mineral deposition was found at 30°C for 5 days and 37°C for 2 days; however, a significant increase in the number and size of calcium nodules (Supplementary Fig. S2) was observed at 37°C for 23 days. For fibrous scaffolds, calcium nodules were only observed at 37°C for 23 days, with both active and static samples containing large aggregates of calcium nodules (Supplementary Fig. S2). At all time points, cells can be seen spanning several fibers and covering the fibrous scaffolds. Cell nuclear staining showed that cells remained attached and viable on scaffolds before, during, and after triggering architectural changes (Supplementary Fig. S3). Cells covered the fibrous and foam scaffolds at all time points, and at 23 days at 37°C, cells formed sheets spanning the pores for the foam scaffolds.
XO staining also revealed that for both foam scaffolds and fibrous scaffolds, calcium deposition was qualitatively comparable between active and static samples (Fig. 3). No detectable calcium deposition was observed on any samples during the initial 5 days at 30°C; as a comparison, nuclear staining showed a substantial amount of cells in the scaffolds (Supplementary Fig. S3). After triggering at 37°C for 2 days, small calcium nodules were detected on both active and static fibrous scaffolds. After 23 days at 37°C, active and static samples for both foam scaffolds and fibrous scaffolds showed a significant amount of calcium deposition, and no qualitative difference between active and static groups was observed.

XO staining for calcium deposition on SMP foam and fibrous scaffolds shows increasing calcium deposition over time. Both foam (top) and fibrous (bottom) scaffolds showed no observable difference between active and static groups. For either foam or fibrous scaffolds, no calcium deposition is detected during the 5 days of culture at 30°C. Following triggering of architecture change at 37°C for 2 days, small calcium nodules (white specks) are detected in the active and static control groups for both foam and fibrous scaffolds. After 23 days at 37°C, a significant increase in the size and number of calcium nodules is detected for all groups. Scale bar is 200 μm. For foam scaffolds, histogram stretching was performed on the raw images to allow visualization of calcium nodules. Brightness and contrast of the micrographs of fibrous scaffolds were adjusted in ImageJ to improve the contrast between the XO-stained calcium nodules and the background (see the Materials and Methods section for details). XO, xylenol orange.
μCT analysis also demonstrated qualitatively and quantitatively a comparable amount of mineral deposition between active and static groups, with increasing mineral formation being observed over time for both foam scaffolds and fibrous scaffolds (Fig. 4). For foam scaffolds, calcium nodules were observed at all time points, with a noticeable increase in number and size as time progressed. Calcium deposition was concentrated near the outer pores of the foam scaffolds, likely due to a much higher cell density near the surface when compared with the center of the scaffold. No statistical difference (p > 0.25 for all time points) was observed between active and static scaffolds. Both groups showed a trend of increased mineral deposition over time, although the differences were not statistically significant (Supplementary Table S1).

Three-dimensional reconstructions of microcomputed tomography showed mineral deposition over time in foam and fibrous scaffolds. Both foam (top) and fibrous (bottom) scaffolds showed qualitatively comparable amount of mineral deposition in active and static groups. For foam scaffolds, both active and static scaffolds showed minimal minerals before transition (30°C for 5 days) and 2 days after transition (37°C for 2 days), and mineral size was small. A significant increase in mineral volume and size was found on both groups 23 days after transition (37°C for 23 days). For fibrous scaffolds, neither static nor active groups showed mineral deposition at 30°C for 5 days; however, artifacts (pointed by arrow) from the scaffolds were observed. At 37°C for 2 days, both groups showed minimal mineral formation, and minerals appear to begin nucleating from the edges of the scaffolds. At the last time point, scaffolds from both groups were covered by mineral nodules. Scale bar: 1 mm.
For fibrous scaffolds, no calcium nodules were detected at 30°C for 5 days; however, small artifacts from the fiber mat can be observed, which we speculate may result from residual salts from PBS rinsing before μCT scanning. At 37°C for 2 days, calcium nodules can be seen for both active and static scaffolds and appear to begin nucleating from the edges of the fiber mat. At 37°C for 23 days, both active and static scaffolds are completely covered with calcium deposits. No statistical difference (p > 0.15) was found between active and static groups; however, when comparing within group, both groups demonstrated a significant increase in mineral deposition over time (Supplementary Table S2).
ALP activity
ALP protein expression being an early marker of mineral nodule formation—a functional endpoint indication of osteogenic differentiation, 38 quantitative analysis of ALP activity showed no statistical difference (p > 0.1 for fibrous scaffolds and p > 0.15 for foam scaffolds at all time points) between active and static groups of foam scaffolds and fibrous scaffolds (Fig. 5).

ALP activity of cells in active and static foam and fibrous scaffolds. No significant difference in ALP activity was found between active and static scaffolds for both foam (top) and fibrous (bottom) scaffolds. For foam scaffolds, a trend of increased ALP activity was seen after transition (37°C for 2 days and 37°C for 23 days) compared with before transition (30°C for 5 days). However, no statistically significant difference was found. For fibrous scaffolds, static scaffolds demonstrated a trend of increasing ALP activity, yet not statistically significant. Horizontal lines represent averages of three replicates in the groups. ALP, alkaline phosphatase.
Real-time PCR gene expression
Real-time PCR analysis of genetic markers of osteogenesis showed that both foam scaffolds and fibrous scaffolds expressed the osteogenic marker, Runx2, with no significant difference (p > 0.23 for foam scaffolds and p > 0.54 for fibrous scaffolds) between active and static groups found at all time points (Fig. 6). Upregulation of Runx2 expression in foam scaffolds suggests that hASCs differentiated toward an osteogenic lineage over time (p < 0.05). For fibrous scaffolds, no significant trend of Runx2 over time was shown (p > 0.57).
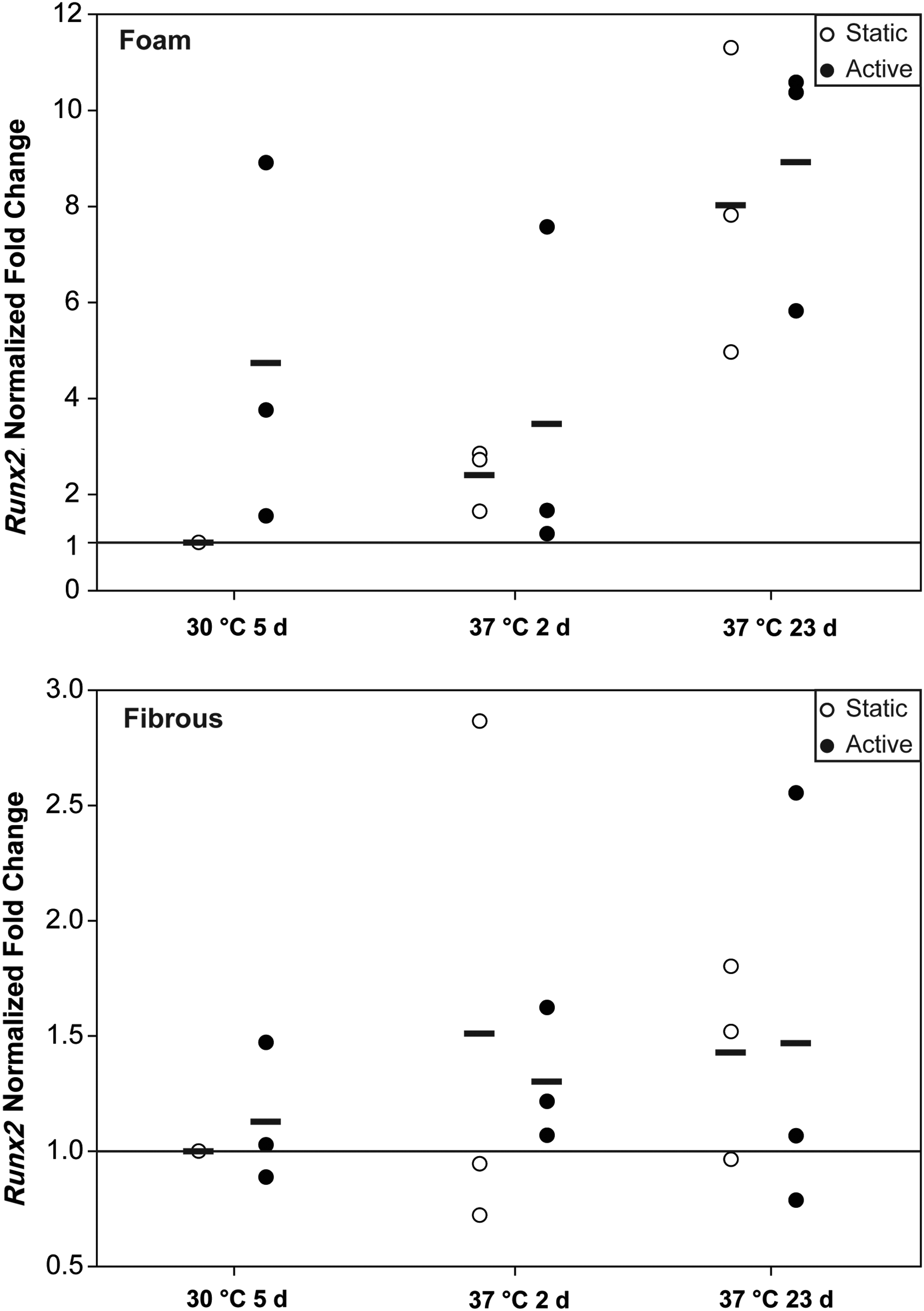
Real-time quantitative polymerase chain reaction expression of osteogenic marker, Runx2, for active and static foam and fibrous scaffolds. There was no difference between active and static scaffolds in both foam (top) and fibrous (bottom) scaffolds across all time points. For both foam and fibrous scaffolds, a trend of increasing (upregulation) Runx2 over time was observed for active and static scaffolds, although significance was not achieved. Horizontal lines indicate averages over 3 independent replicates.
A second characteristic marker, osteocalcin (OC; Supplementary Fig. S4), was also expressed in both foam scaffolds and fibrous scaffolds, with no significant difference between active and static groups for both types of scaffolds observed (p > 0.2 for foam scaffolds and p > 0.12 for fibrous scaffolds) at all time points. A trend of OC downregulation was found in fibrous scaffolds (p < 0.05), but not in foam scaffolds (p = 0.104).
Histology
Alizarin red staining at the final time point (Fig. 7) showed substantial calcium deposition in both foam scaffolds and fibrous scaffolds. The results indicate that qualitatively comparable level of osteogenic differentiation was achieved in both types of scaffolds with or without architecture change. For the foam scaffolds, both active and static groups stained strongly positive for calcium deposition. Foam scaffolds were not sectioned before staining since the high porosity of foam scaffolds resulted in sections with little material. The material itself does not absorb the staining as foam scaffolds with no cells did not stain red (Supplementary Fig. S5). Likewise, for fibrous scaffolds, both active and static samples stained strongly positive, indicating calcium deposition on the scaffolds.
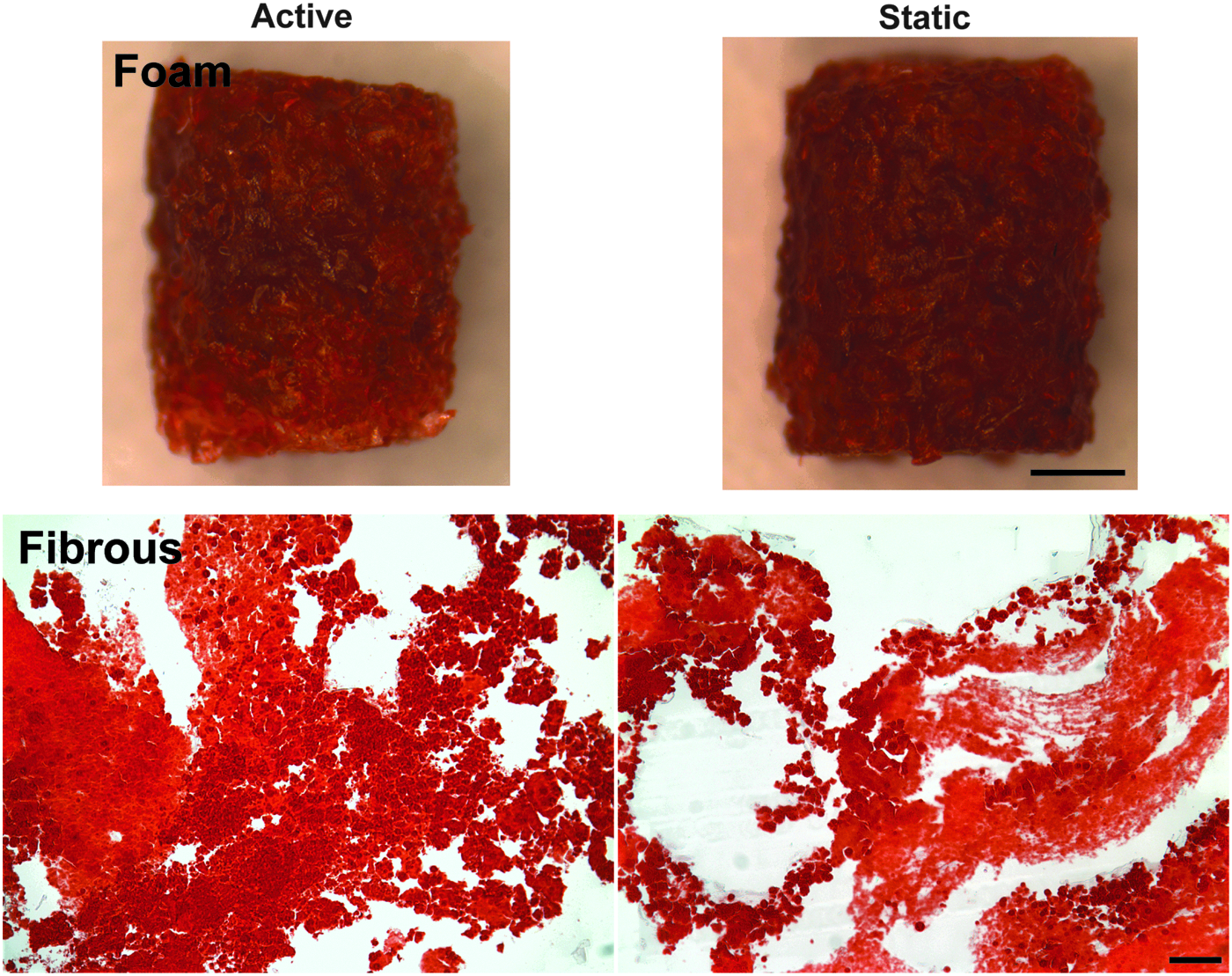
Alizarin red staining of calcium deposition in foam and fibrous scaffolds 4 weeks after culture. Calcium deposition (red stain) was observed in both foam (top) and fibrous (bottom) scaffolds. No apparent differences between active and static scaffolds were observed in either foam or fibrous scaffolds. Both active and static groups in foam and fibrous scaffolds showed significant amount of calcium deposition at the final time point (37°C for 23 days). For foam scaffolds, the scale bar is 1 mm. For fibrous scaffolds, the scale bar is 100 μm. Color images available online at www.liebertpub.com/tea
Discussion
This work is the first to demonstrate differentiation of stem cells cultured in 3D SMP scaffolds during and following programmed architectural changes in SMP scaffolds and found no detrimental effects on stem cell osteogenic differentiation capacity. Osteogenic differentiation was confirmed by mineral deposition, protein production, and gene expression assays. Comparable levels of mineral deposition, protein production, and gene expression between active and static scaffolds were observed.
The mineral deposition assays found qualitatively and quantitatively comparable amounts of mineral deposition for both active and static scaffolds (Figs. 2, 3, and 4). In addition, EDS analysis (Supplementary Fig. S2) showed that the nodules on the SMP scaffolds consisted of high levels of calcium and phosphorus, two key constituents of bone mineral. 39 These results suggest that hASCs differentiated down an osteoblastic lineage and produced mineralized matrix during the course of the 4-week culture.
In examining osteogenic protein production, ALP activity was less conclusively indicative of osteogenic function than was mineral deposition and was similar between the active and static groups. Instead of increasing over time as shown in some previous studies, 40 ALP activity remained unchanged or increased modestly for both SMP scaffolds (Fig. 5). Such relatively flat ALP activity during stem cell osteogenic differentiation has been reported in prior studies.41,42 It is possible that in the present study, the peak ALP activity occurred during the 3-week gap between the second and third time points. A lack of a significant increase in ALP activity may also be attributed to the fetal bovine serum that was used in the induction medium, which has been reported to result in low ALP protein production in hASCs compared with other sera. 43
Results of real-time qPCR showed no significant difference between active and static groups for both SMP scaffolds (Fig. 6 and Supplementary Fig. S4); however, a trend of upregulation of Runx2 expression in foam scaffolds and downregulation of OC expression in fibrous scaffolds over time was found. With respect to the downregulation of OC over time, such downregulation in hASCs has also been shown in previous studies.42,44 As with the results presented in the present study, the prior studies demonstrated successful mineral deposition regardless of the reduced OC expression. These aforementioned studies suggest that although ALP activity and osteogenic gene expression are indicators of osteogenic differentiation,41,42,44 mineralized matrix formation represents successful endpoint outcome of osteogenic differentiation.40,45
This work focused on the extent to which stem cell osteogenic differentiation capacity is preserved following programmed shape change, and it is worth noting that study of other lineage pathways, other stem cell types, and other programmed shape changes may reveal different effects of interaction between stem cell biology and shape memory activation. The response to shape memory activation may be found to be cell-type and lineage pathway specific. Moreover, shape memory materials can be fabricated in diverse architectures and programmed to change shape in a variety of ways, the specifics of which may affect the response of cells resident therein. Therefore, even as the findings here support the feasibility of employing shape memory in scaffolds for stem cell-based therapies for bone repair, many interesting questions related to the effect of shape memory activation on stem cell biology remain to be investigated.
In this study, we have demonstrated successful induction of stem cell osteogenic differentiation following programmed architectural changes in SMP scaffolds with no observed reduction in osteogenic differentiation capacity compared with static controls. The SMP scaffolds used herein have previously been studied in vivo in grafting and stabilizing complex bone defects in a mouse segmental defect model. 25 The findings of the present work support the feasibility of employing shape memory in scaffolds for stem cell-based therapies for bone repair.
Conclusions
In this study, we have demonstrated osteogenic differentiation of adipose-derived stem cells in two different types of programmable SMP scaffolds—foam scaffolds and fibrous scaffolds—upon shape memory-triggered changes in internal architectures under cytocompatible conditions. The foam scaffolds transitioned from a temporarily compressed pore architecture to a permanently open pore architecture; the fibrous scaffolds transitioned from a temporarily unaligned architecture to a permanently uniaxially aligned architecture. The results indicate that stem cells demonstrate comparable level of osteogenic characteristics when in the active and static scaffolds, as indicated by mineral deposition, ALP protein production, and osteogenic gene expression. Promisingly, this present study suggests the feasibility of employing shape memory scaffolds in cell-based strategy for the treatment of bone defects.
Footnotes
Acknowledgments
The authors gratefully thank Professor Megan Oest for microcomputed tomography data analysis. This work was sponsored by DARPA (DP12AP00271 to J.H.H.), and the content of the information does not necessarily reflect the position or the policy of the Government, and no endorsement should be inferred.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.